
From Phenology and Habitat Preferences to Climate Change: Importance of Citizen Science in Studying Insect Ecology in the Continental Scale with American Red Flat Bark Beetle, Cucujus clavipes, as a Model Species


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Citizen Scientific Data from iNaturalist
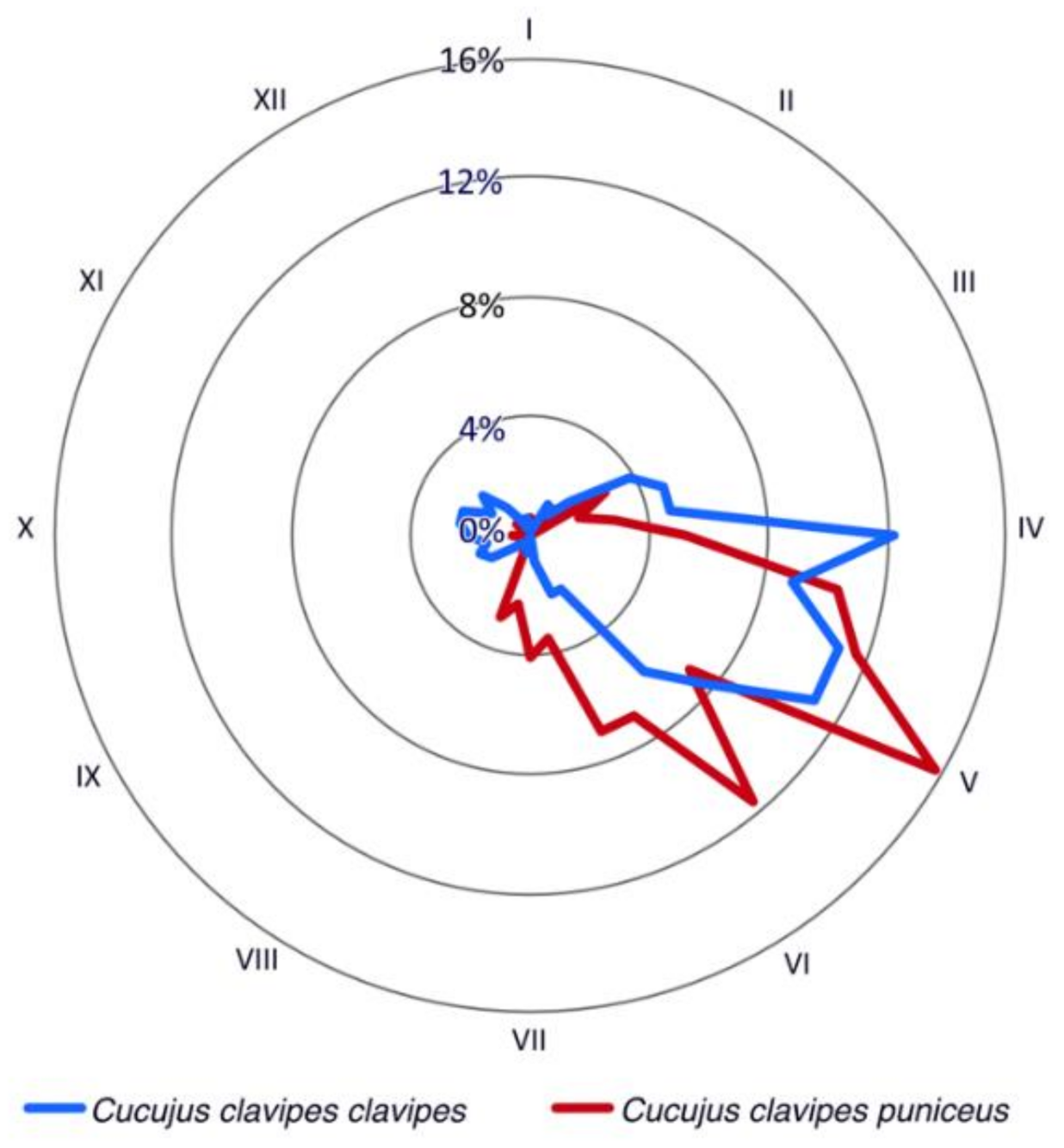
2.2. Phenology of Adults
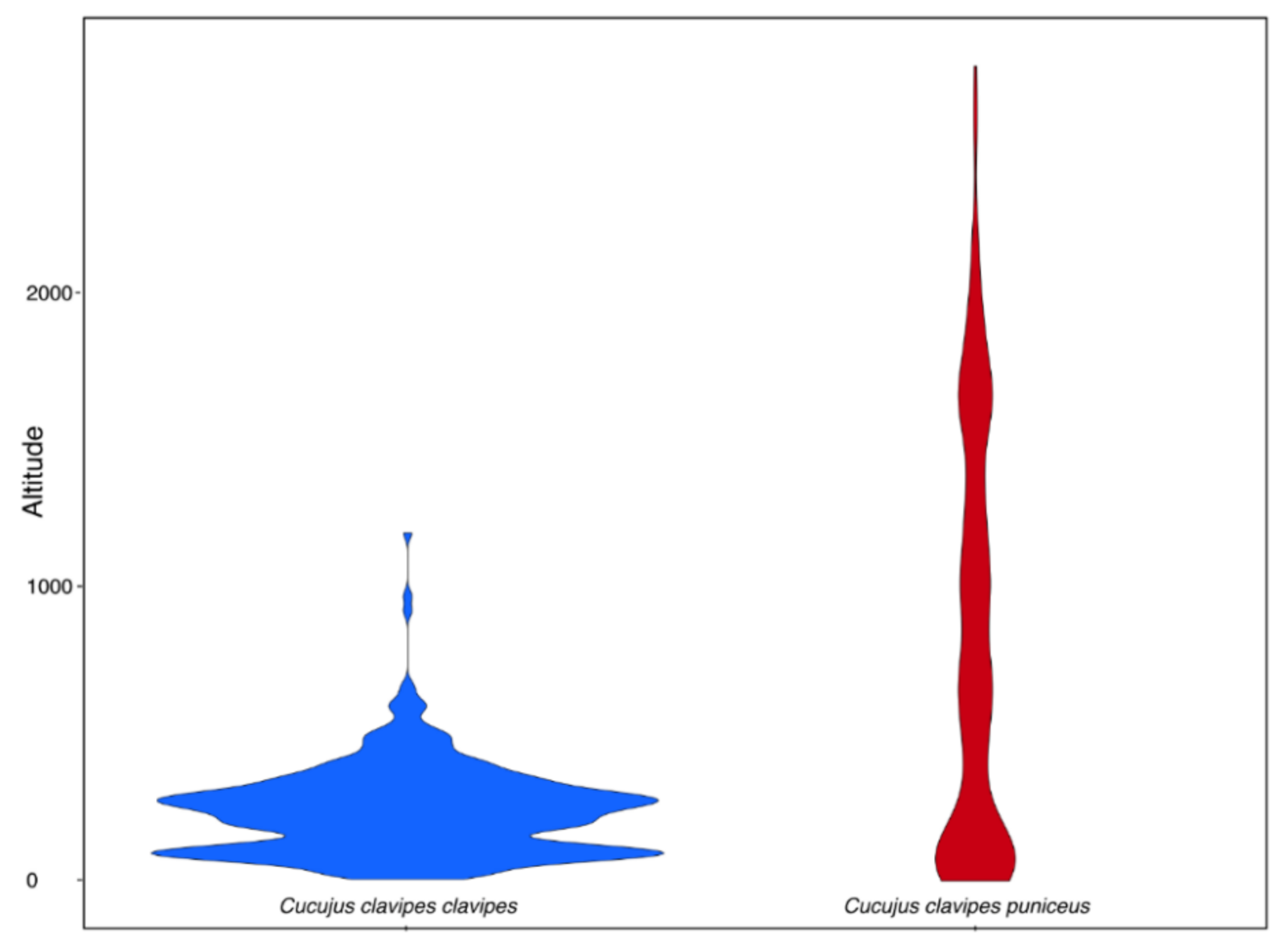
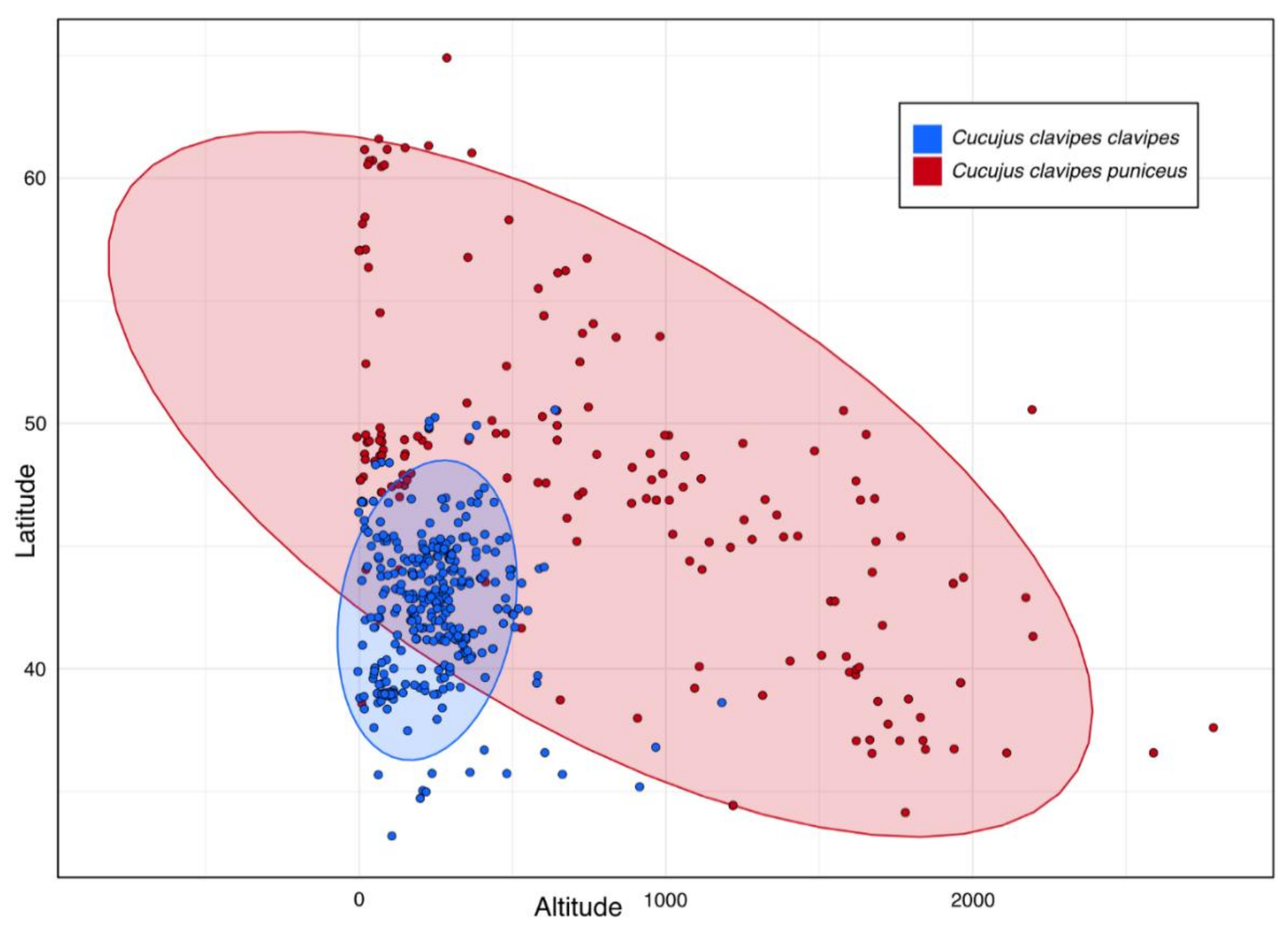
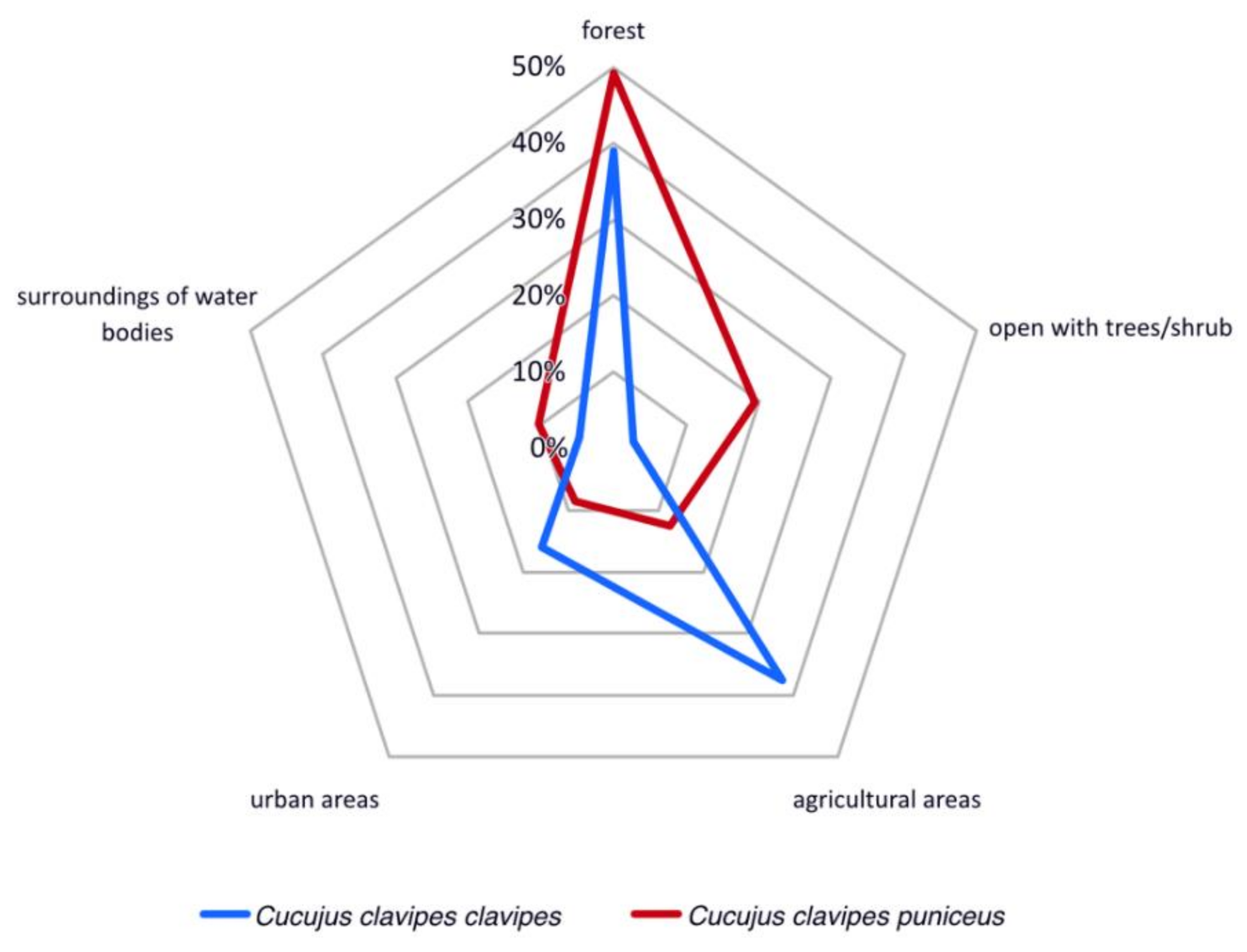
2.3. Habitat Preferences
2.4. Impact of Climate Change
3. Results
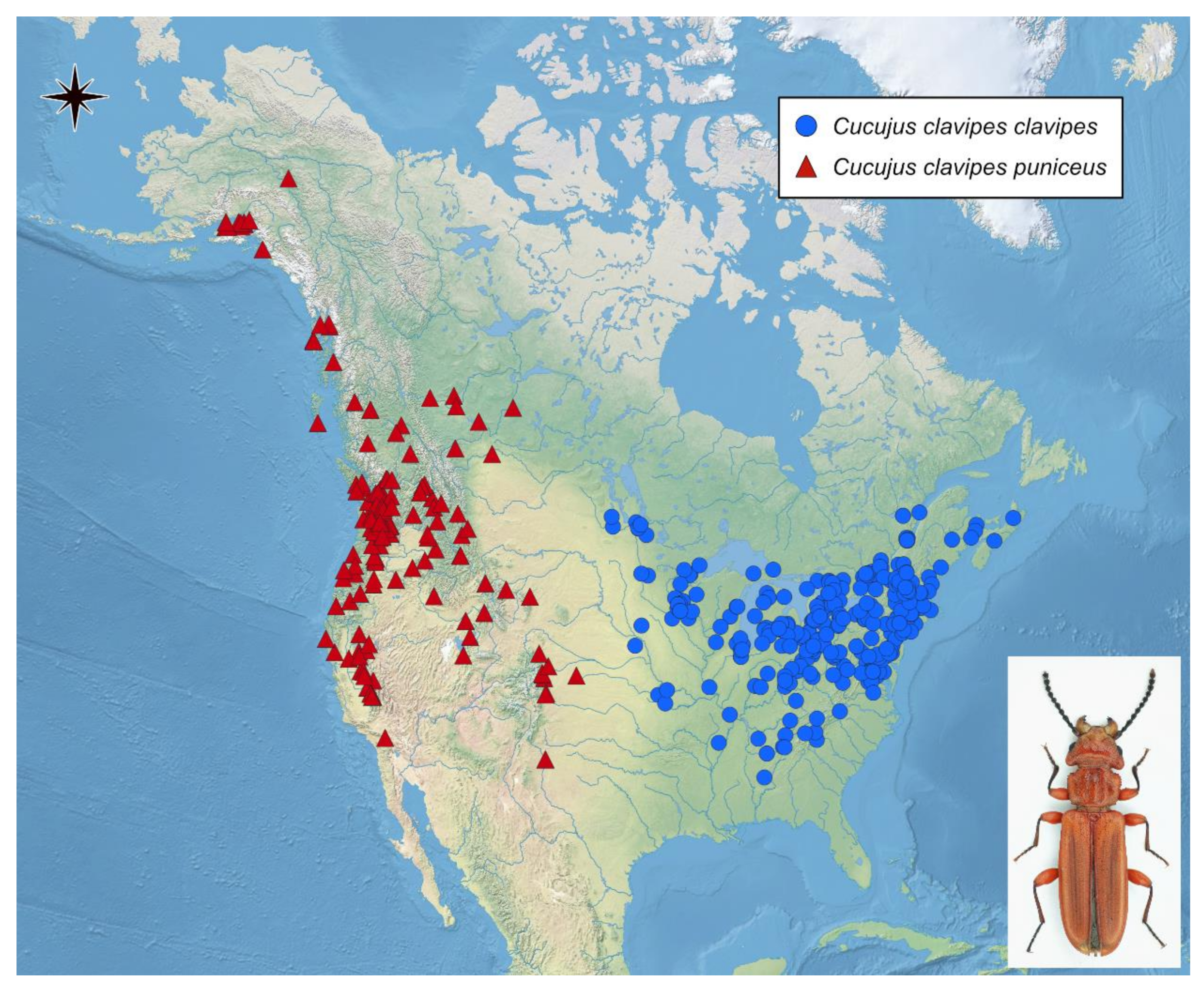
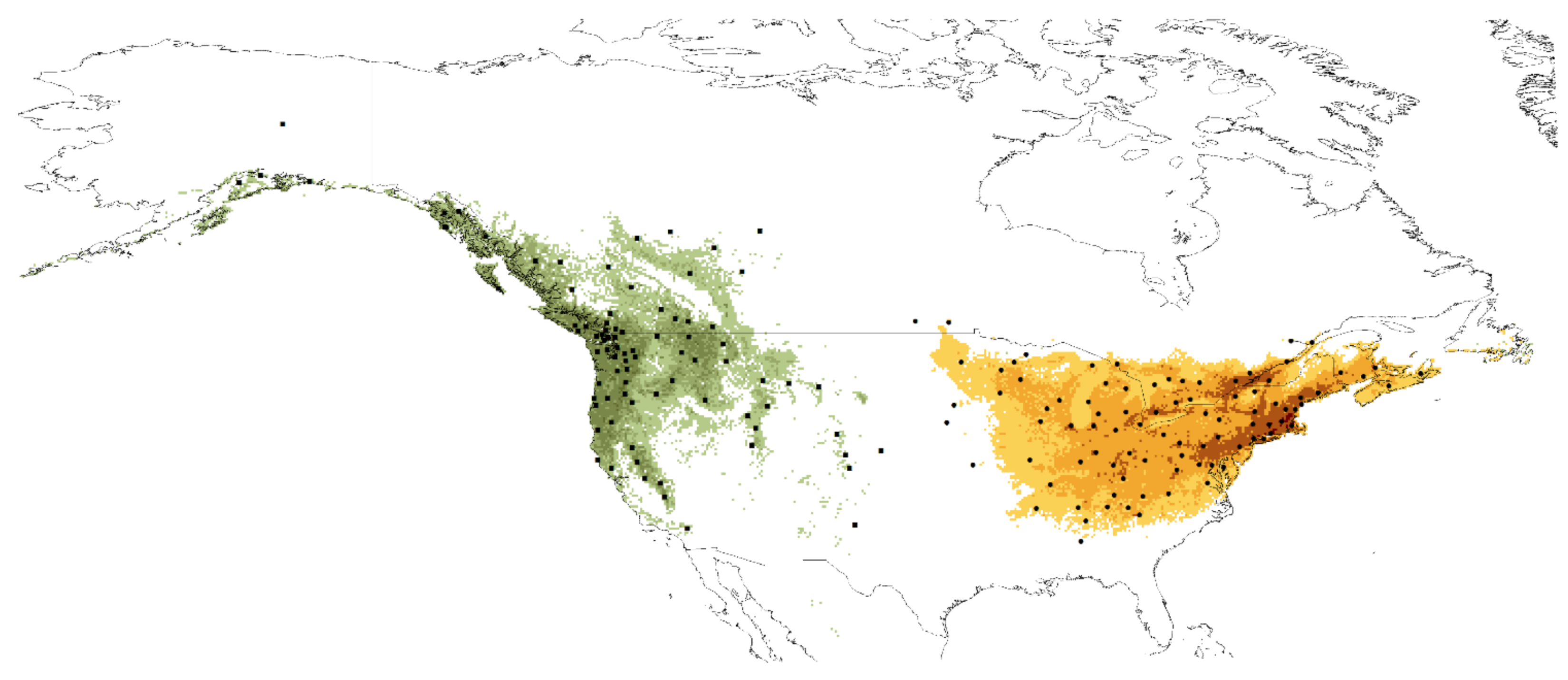
3.1. Activity of Adult Beetles of Cucujus clavipes and C. v. puniceus
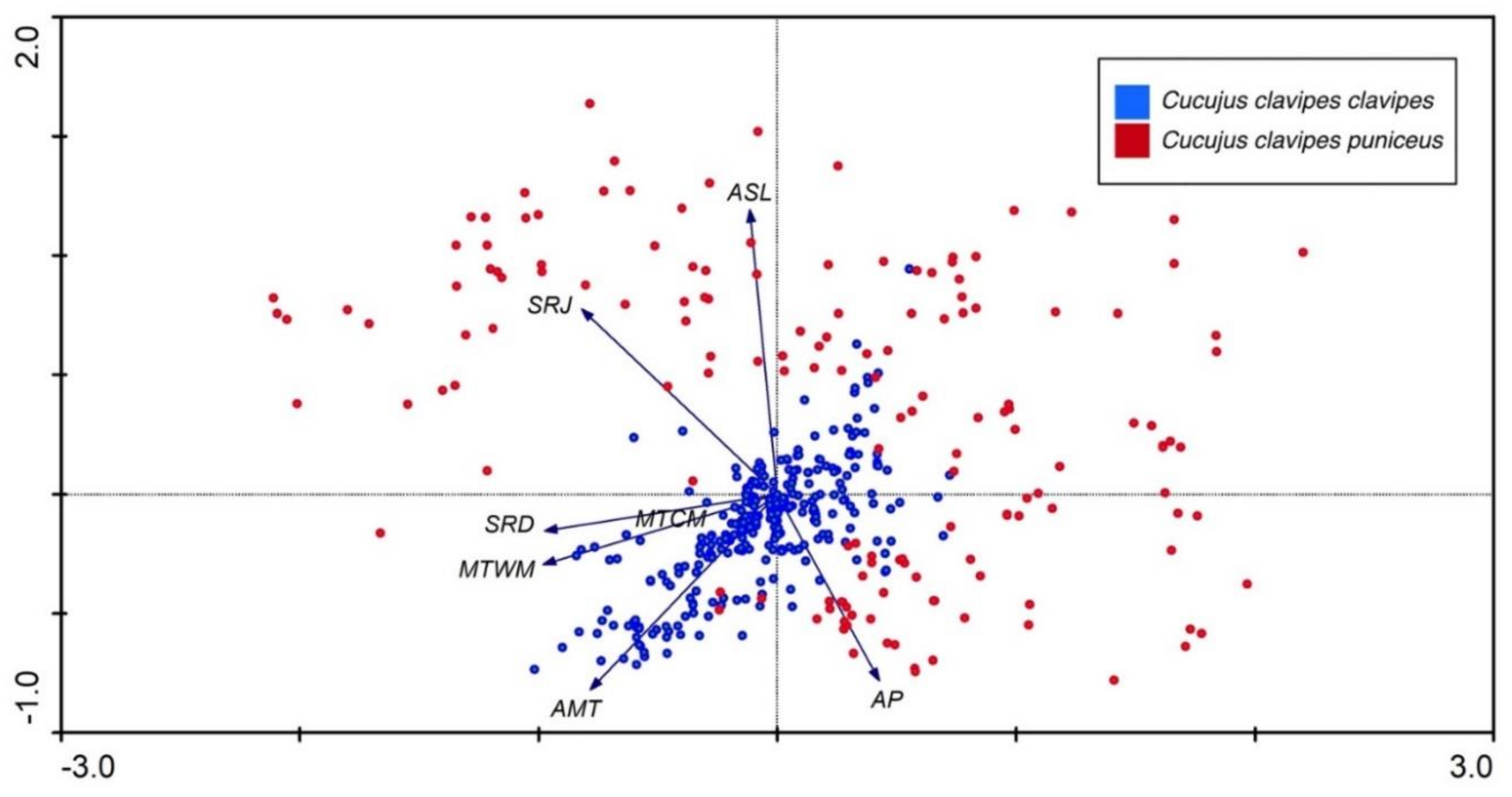
3.2. Environmental Parameters vs. Present Distribution of Cucujus clavipes Subspecies
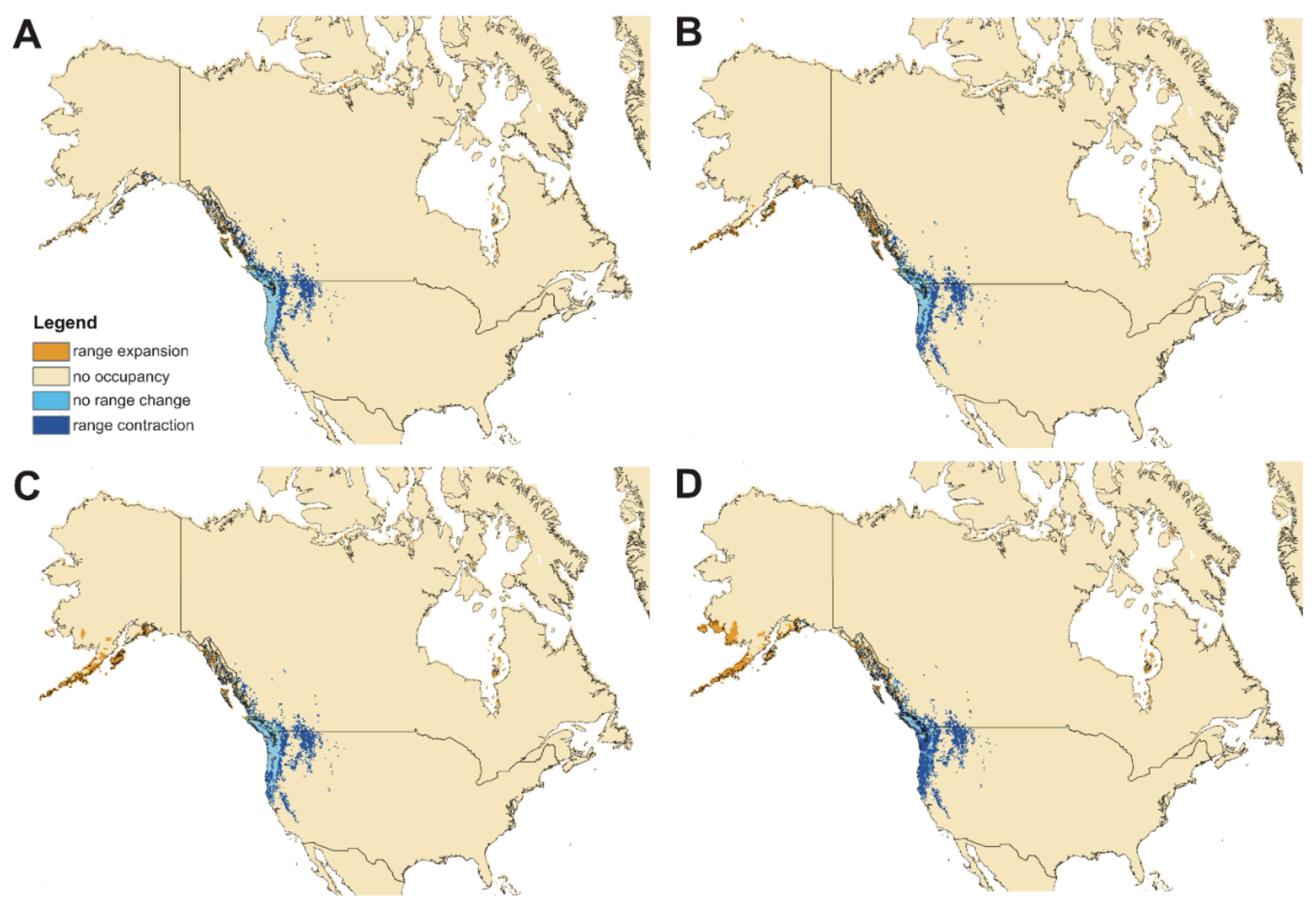
3.3. ENM—Models Evaluation, Limiting Factors and Range Overlap between Subspecies and Impact of Global Warning
3.4. Impact of Global Warming
4. Discussion
4.1. Phenological Activity of Cucujus clavipes Subspecies
4.2. Habitat Preferences of Cucujus clavipes
4.3. The Future of Cucujus clavipes Because of Climate Change
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gura, T. Citizen science: Amateur experts. Nature 2013, 496, 259–261. [Google Scholar] [CrossRef] [Green Version]
- Hand, E. Citizen science: People power. Nature 2010, 466, 685–687. [Google Scholar] [CrossRef] [Green Version]
- Miller-Rushing, A.; Primack, R.; Bonney, R. The history of public participation in ecological research. Front. Ecol. Environ. 2012, 10, 285–290. [Google Scholar] [CrossRef]
- Kobori, H.; Dickinson, J.L.; Washitani, I.; Sakurai, R.; Amano, T.; Komatsu, N.; Kitamura, W.; Takagawa, S.; Koyama, K.; Ogawara, T.; et al. Citizen science: A new approach to advance ecology, education and conservation. Ecol. Res. 2016, 31, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Richter, C.F.; Lortie, C.J.; Kelly, T.L.; Filazzola, A.; Nunes, K.A.; Sarkar, R. Online but not remote: Adapting field-based ecology laboratories for online learning. Ecol. Evol. 2020. [Google Scholar] [CrossRef]
- Unger, S.; Rollins, M.; Tietz, A.; Dumais, H. iNaturalist as an engaging tool for identifying organisms in outdoor activities. J. Biol. Educ. 2020. [Google Scholar] [CrossRef]
- Wilson, J.; Jisming-See, S.; Brandon-Mong, G.; Lim, A.; Lim, V.; Lee, P.; Sing, K. Citizen science: The first peninsular Malaysia butterfly count. Biodivers. Data J. 2015, 3, e7159. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, L.M.; Bachman, S.P. The role of citizen science in a global assessment of extinction risk in palms (Arecaceae). Bot. J. Linn. Soc. 2016, 182, 543–550. [Google Scholar] [CrossRef] [Green Version]
- Sumner, S.; Bevan, P.; Hart, A.G.; Isaac, N.J.B. Mapping species distributions in 2 weeks using citizen science. Insect Conserv. Divers. 2019, 12, 382–388. [Google Scholar] [CrossRef]
- Girardello, M.; Chapman, A.; Dennis, R.; Kaila, L.; Borges, P.A.V.; Santangeli, A. Gaps in butterfly inventory data: A global analysis. Biol. Conserv. 2019, 236, 289–295. [Google Scholar] [CrossRef]
- Skejo, J.; Gupta, S.K.; Chandra, K.; Panhwar, W.A.; Franjević, D. Oriental macropterous leaf-mimic pygmy grasshoppers—genera Oxyphyllum and Paraphyllum (Orthoptera: Tetrigidae) and their taxonomic assignment. Zootaxa 2019, 4590, 546–560. [Google Scholar] [CrossRef] [PubMed]
- La Sorte, F.A.; Somveille, M. The island biogeography of the eBird citizen-science programme. J. Biogeogr. 2020, 48, 628–638. [Google Scholar] [CrossRef]
- Rose, K.; Franjević, D. Online social media tells a story of Anaselina, Paraselina, and Selivinga (Orthoptera, Tetrigidae), rare Australian pygmy grasshoppers. ZooKeys 2020, 948, 107–119. [Google Scholar] [CrossRef]
- Webster, R.P.; de Tonnancour, P.; Sweeney, J.D.; Webster, V.L.; Kostanowicz, C.A.; Hughes, C.; Anderson, R.S.; Klymko, J.; Chantal, C.; Vigneault, R. New Coleoptera records from eastern Canada, with additions to the fauna of Manitoba, British Columbia, and Yukon Territory. ZooKeys 2020, 946, 53–112. [Google Scholar] [CrossRef]
- Silva, D.P.; Hall, H.G.; Ascher, J.S. Predicting the distribution range of a recently described, habitat specialist bee. J. Insect Conserv. 2020, 24, 671–680. [Google Scholar] [CrossRef]
- Jesus, M.D.; Zapelini, C.; Schiavetti, A. Can citizen science help delimit the geographical distribution of a species? The case of the Callistoctopus sp. (eastern octopus) on the Brazilian coast. Ethnobiol. Conserv. 2021, 10, 03. [Google Scholar] [CrossRef]
- Bathori, F.; Pfliegler, W.; Zimmerman, C.-U.; Tartally, A. Online image databases as multi-purpose resources: Discovery of a new host ant of Rickia wasmannii Cavara (Ascomycota, Laboulbeniales) by screening AntWeb.org. J. Hymenopt. Res. 2017, 61, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Sheard, J.K.; Sanders, N.J.; Gundlach, C.; Schär, S.; Larsen, R.S. Monitoring the influx of new species through citizen science: The first introduced ant in Denmark. PeerJ 2020, 8, e8850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skejo, J.; Deranja, M.; Adžić, K. Pygmy hunchback of New Caledonia: Notredamia dora gen. n. et sp. n.—A New Cladonotin (Caelifera: Tetrigidae) genus and species from Oceania. Entomol. News 2020, 129, 170–185. [Google Scholar] [CrossRef]
- Hadjiconstantis, M.; Zoumides, C. First records of the pest leaf beetle Chrysolina (Chrysolinopsis) americana (Linnaeus, 1758) (Coleoptera, Chrysomelidae) in Cyprus—A study initiated from social media. Biodivers. Data J. 2021, 9, e61349. [Google Scholar] [CrossRef]
- Hartop, E.A.; Brown, B.V.; Disney, R.H.L. Opportunity in our ignorance: Urban biodiversity study reveals 30 new species and one new nearctic record for Megaselia (Diptera: Phoridae) in Los Angeles (California, USA). Zootaxa 2015, 3941, 451–484. [Google Scholar] [CrossRef]
- Hiller, T.; Haelewaters, D. A case of silent invasion: Citizen science confirms the presence of Harmonia axyridis (Coleoptera, Coccinellidae) in Central America. PLoS ONE 2019, 14, e0220082. [Google Scholar] [CrossRef] [Green Version]
- Jaskuła, R.; Michalski, M.; Marris, J.W.M. First records of the Palaestes abruptus Sharp, 1899 and P. nicaraguae Sharp, 1899 (Coleoptera: Cucujidae) from South America, with a checklist of flat bark beetles from the continent. Biodivers. Data J. 2021, 9, e62576. [Google Scholar] [CrossRef]
- Alvarado-Cardenas, L.O.; Chavez-Hernandez, M.G.; Leon, J.F.P. Gonolobus naturalistae (Apocynaceae; Asclepiadoideae; Gonolobeae; Gonolobinae), a new species from Mexico. Phytotaxa 2020, 472, 249–258. [Google Scholar] [CrossRef]
- Amorim, D.D.; Brown, B.V. Urban Scatopsidae (Diptera) of Los Angeles, California, United States. Insect Syst. Divers. 2020, 4, 1. [Google Scholar] [CrossRef]
- Adhikari, B.; Wood, J.R.I. Thunbergia kasajuana, a new species of Acanthaceae from Nepal. Kew Bull. 2020, 75, 26. [Google Scholar] [CrossRef]
- Cadena-Castañeda, O.J.; Díaz, C.J.A.; Rodríguez, N.O.P.; García, A.G. Studies on raspy crickets: New Triaenogryllacris species (Orthoptera: Gryllacrididae). Zootaxa 2020, 4896, 239–250. [Google Scholar] [CrossRef]
- Moonlight, P.; Hollands, R.; Cano, A.; Purvis, D. A new species of tuberosus Begonia (Begoniaceae) from Andean Peru. Edinb. J. Bot. 2020, 77, 145–159. [Google Scholar] [CrossRef]
- Winterton, S.L. A new bee-mimicking stiletto fly (Therevidae) from China discovered on iNaturalist. Zootaxa 2020, 4816, 361–369. [Google Scholar] [CrossRef]
- Coxen, C.L.; Frey, J.K.; Carleton, S.A.; Collins, D.P. Species distribution models for a migratory bird based on citizen science and satellite tracking data. Glob. Ecol. Conserv. 2017, 11, 298–311. [Google Scholar] [CrossRef]
- Schubert, S.C.; Manica, L.T.; Guaraldo, A.D. Revealing the potential of a huge citizen-science platform to study bird migration. EMU Austral Ornithol. 2019, 119, 364–373. [Google Scholar] [CrossRef]
- DeGroote, L.W.; Hingst-Zaher, E.; Moreira-Lima, L.; Whitacre, J.V.; Slyder, J.B.; Wenzel, J. Citizen science data reveals the cryptic migration of the Common Potoo Nyctibius griseus in Brazil. Ibis 2020, 163, 380–389. [Google Scholar] [CrossRef]
- Weisshaupt, N.; Lehtiniemi, T.; Koistinen, J. Combining citizen science and weather radar data to study large-scale bird movements. Ibis 2020, 193, 728–736. [Google Scholar] [CrossRef]
- de Camargo Barbosa, K.V.; Develey, P.F.; Ribeiro, M.C.; Jahn, A.E. The contribution of citizen science to research on migratory and urban birds in Brazil. Ornithol. Res. 2021. [Google Scholar] [CrossRef]
- Katušić, L.; Jelaska, S.D.; Šerić Jelaska, L. Monitoring of saproxylic beetles in Croatia: Following the path of the stag beetle. Nat. Conserv. 2017, 19, 39–56. [Google Scholar] [CrossRef] [Green Version]
- Méndez, M.; de Jaime, C.; Alcántara, M.A. Habitat description and interannual variation in abundance and phenology of the endangered beetle Lucanus cervus L. (Coleoptera) using citizen science monitoring. J. Insect. Conserv. 2017, 21, 907–915. [Google Scholar] [CrossRef]
- Zapponi, L.; Cini, A.; Bardiani, M.; Hardersen, S.; Maura, M.; Maurizi, E.; Redolfi De Zan, L.; Audisio, P.; Bologna, M.A.; Carpaneto, G.M.; et al. Citizen science data as an efficient tool for mapping protected saproxylic beetles. Biol. Conserv. 2017, 208, 139–145. [Google Scholar] [CrossRef]
- Wilson, J.S.; Pan, A.D.; General, D.E.M.; Koch, J.B. More eyes on the prize: An observation of a very rare, threatened species of Philippine Bumble bee, Bombus irisanensis, on iNaturalist and the importance of citizen science in conservation biology. J. Insect Conserv. 2020, 24, 727–729. [Google Scholar] [CrossRef]
- Pawson, S.M.; Sullivan, J.J.; Grant, A. Expanding general surveillance of invasive species by integrating citizens as both observers and identifiers. J. Pest Sci. 2020, 93, 1155–1166. [Google Scholar] [CrossRef]
- Purkart, A.; Depa, Ł.; Kollár, J.; Suvák, M.; Holecová, M.; Goffová, K.; Országhová, Z. Citizen science reveals the current distribution of the new plant pest Aphis nerii in Slovakia. Plant Prot. Sci. 2020, 56, 101–106. [Google Scholar] [CrossRef]
- Crall, A.W.; Newan, G.J.; Jarnevich, C.S.; Stohlgren, T.J.; Waller, D.M.; Graham, J. Improving and integrating data on invasive species collected by citizen scientists. Biol. Invasions 2010, 12, 3419–3428. [Google Scholar] [CrossRef]
- Gallo, T.; Waitt, D. Creating a successful citizen science model to detect and report invasive species. BioScience 2011, 61, 459–465. [Google Scholar] [CrossRef] [Green Version]
- Maistrello, L.; Dioli, P.; Bariselli, M.; Mazzoli, G.L.; Giacalone-Forini, I. Citizen science and early detection of invasive species: Phenology of first occurrences of Halyomorpha halys in Southern Europe. Biol. Invasions 2016, 18, 3109–3116. [Google Scholar] [CrossRef]
- Goczał, K.; Rossa, R.; Sweeney, J.; Tofilski, A. Citizen monitoring of invasive species: Wing morphometry as a tool for detection of alien Tetropium species. J. Appl. Entomol. 2017, 141, 496–506. [Google Scholar] [CrossRef]
- Mannino, A.M.; Balistreri, P. Citizen science: A successful tool for monitoring invasive alien species (IAS) in Marine Protected Areas. The case study of the Egadi Islands MPA (Tyrrhenian Sea, Italy). Biodiversity 2018, 19, 42–48. [Google Scholar] [CrossRef]
- Roy-Dufresne, E.; Saltré, F.; Cooke, B.D.; Mellin, C.; Mutze, G.; Cox, T.; Fordham, D.A. Modeling the distribution of a wide-ranging invasive species using the sampling efforts of expert and citizen scientists. Ecol. Evol. 2019, 9, 11053–11063. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.Y.; McQuillan, P.B.; McDonald, E.; Hawkins, C. Citizen science reveals the Palaearctic poison hemlock moth Agonopterix alstroemeriana (Clerck) (Lepidoptera: Depressariidae) has established in Australia. N. Z. Entomol. 2020, 43, 86–92. [Google Scholar] [CrossRef]
- Johnson, B.A.; Mader, A.D.; Dasgupta, R.; Kumar, P. Citizen science and invasive alien species: An analysis of citizen science initiatives using information and communications technology (ICT) to collect invasive alien species observations. Glob. Ecol. Conserv. 2020, 21, e00812. [Google Scholar] [CrossRef]
- Werenkraut, V.; Baudino, F.; Roy, H.E. Citizen science reveals the distribution of the invasive harlequin ladybird (Harmonia axyridis Pallas) in Argentina. Biol. Invasions 2020, 22, 2915–2921. [Google Scholar] [CrossRef]
- Encarnação, J.; Teodósio, M.A.; Morais, P. Citizen Science and Biological Invasions: A Review. Front. Environ. Sci. 2021, 8, 602980. [Google Scholar] [CrossRef]
- Johansson, F.; Heino, J.; Coiffard, P.; Svanbäck, R.; Wester, J.; Bini, L.M. Can information from citizen science data be used to predict biodiversity in stormwater ponds? Sci. Rep. 2020, 10, 9380. [Google Scholar] [CrossRef]
- Kagawa, O.; Uchida, S.; Yamazaki, D.; Osawa, Y.; Ito, S.; Chiba, S. Citizen science via social media revealed conditions of symbiosis between a marine gastropod and an epibiotic alga. Sci. Rep. 2020, 10, 19647. [Google Scholar] [CrossRef] [PubMed]
- Nowak, K.; Berger, J.; Panikowski, A.; Reid, D.G.; Jacob, A.L.; Newman, G.; Young, N.E.; Beckmann, J.P.; Richards, S.A. Using community photography to investigate phenology: A case study of coat molt in the mountain goat (Oreamnos americanus) with missing data. Ecol. Evol. 2020, 10, 13488–13499. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.J.; Vise, C.; Krishnamoorthy, M.A.; Kingston, T.; Venter, S. Citizen science confirms the rarity of fruit bat pollination of baobab (Adansonia digitata) flowers in Southern Africa. Diversity 2020, 12, 106. [Google Scholar] [CrossRef] [Green Version]
- Schultz, C.B.; Brown, L.M.; Pelton, E.; Crone, E.E. Citizen science monitoring demonstrates dramatic declines of monarch butterflies in western North America. Biol. Conserv. 2017, 214, 343–346. [Google Scholar] [CrossRef]
- Bried, J.; Ries, L.; Smith, B.; Patten, M.; Abbott, J.; Ball-Damerow, J.; Cannings, R.; Cordero-Rivera, A.; Córdoba-Aguilar, A.; De Marco, P., Jr.; et al. Towards global volunteer monitoring of odonate abundance. BioScience 2020, 70, 914–923. [Google Scholar] [CrossRef]
- Michielini, J.P.; Dopman, E.B.; Crone, E.E. Changes in flight period predict trends in abundance of Massachusetts butterflies. Ecol. Lett. 2021, 24, 249–257. [Google Scholar] [CrossRef]
- Young, B.E.; Dodge, N.; Hunt, P.D.; Ormes, M.; Schlesinger, M.D.; Shaw, H.Y. Using citizen science data to support conservation in environmental regulatory contexts. Biol. Conserv. 2019, 237, 57–62. [Google Scholar] [CrossRef]
- Brown, B.B.; Hunter, L.; Santos, S. Bird-window collisions: Different fall and winter risk and protective factors. PeerJ 2020, 8, e9401. [Google Scholar] [CrossRef]
- Duan, H.; Xia, S.; Yu, X.; Liu, Y.; Teng, J.; Dou, Y. Using citizen science data to inform the relative sensitivity of waterbirds to natural versus human-dominated landscapes in China. Ecol. Evol. 2020, 10, 7233–7241. [Google Scholar] [CrossRef] [PubMed]
- Irga, P.J.; Dominici, L.; Torpy, F.R. The mycological social network a way forward for conservation of fungal biodiversity. Environ. Conserv. 2020, 47, 243–250. [Google Scholar] [CrossRef]
- Margules, C.; Boedhihartono, A.K.; Langston, J.D.; Riggs, R.A.; Sari, D.A.; Sarkar, S.; Sayer, J.A.; Supriatna, J.; Winarni, N.L. Transdisciplinary science for improved conservation outcomes. Environ. Conserv. 2020, 47, 224–233. [Google Scholar] [CrossRef]
- Papafitsoros, K.; Panagopoulou, A.; Schofield, G. Social media reveals consistently disproportionate tourism pressure on a threatened marine vertebrate. Anim. Conserv. 2020. [Google Scholar] [CrossRef]
- Grol, M.G.G.; Vercelloni, J.; Kenyon, T.M.; Bayraktarov, E.; van den Berg, C.P.; Harris, D.; Loder, J.A.; Mihaljević, M.; Rowland, P.I.; Roelfsema, C.M. Conservation value of a subtropical reef in south-eastern Queensland, Australia, highlighted by citizen-science efforts. Mar. Freshw. Res. 2021, 72, 1–13. [Google Scholar] [CrossRef]
- Kirchhoff, C.; Callaghan, C.T.; Keith, D.A.; Indiarto, D.; Taseski, G.; Ooi, M.K.I.; Le Breton, T.D.; Mesaglio, T.; Kingsford, R.T.; Cornwell, W.K. Rapidly mapping fire effects on biodiversity at a large-scale using citizen science. Sci. Total Environ. 2021, 755, 142348. [Google Scholar] [CrossRef] [PubMed]
- Reed, M. Scientific citizens, smartphones and social media—Reshaping the socio-spatial networks of participation: Insects, soil and food. Morav. Geogr. Rep. 2020, 28, 61–67. [Google Scholar] [CrossRef]
- Ciceoi, R.; Bǎdulescu, L.-A.; Gutue, M.; Mardare, E.Ş.; Pomohaci, C.M. Citizen-generated data on invasive alien species in Romania: Trends and challenges. Acta Zool. Bulg. 2017, 9, 255–260. [Google Scholar]
- Spear, D.M.; Pauly, G.B.; Kaiser, K. Citizen science as a tool for augmenting museum collection data from urban areas. Front. Ecol. Evol. 2017, 5, 86. [Google Scholar] [CrossRef] [Green Version]
- Heberling, J.M.; Isaac, B.L. iNaturalist as a tool to expand the research value of museum specimens. Appl. Plant Sci. 2018, 6, e1193. [Google Scholar] [CrossRef] [Green Version]
- Brooks, D.R.; Nocera, J.J. Bumble bee (Bombus spp.) diversity differs between forested wetlands and clearcuts in the Acadian forest. Can. J. For. Res. 2020, 50, 1399–1404. [Google Scholar] [CrossRef]
- Marshall, B.M.; Freed, P.; Vitt, L.; Bernardo, P.; Vogel, G.; Lotzkat, S.; Franzen, M.; Hallermann, J.; Sage, R.D.; Bush, B.; et al. An inventory of online reptile images. Zootaxa 2020, 4896, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Seregin, A.P.; Bochkov, D.A.; Shner, J.V.; Garin, E.V.; Pospelov, I.N.; Prokhorov, V.E.; Golyakov, P.V.; Mayorov, S.R.; Svirin, S.A.; Khimin, A.N.; et al. “Flora of Russia” on iNaturalist: A dataset. Biodivers. Data J. 2020, 8, e59249. [Google Scholar] [CrossRef]
- Thomas, M.C. A revision of Pediacus Shuckard (Coleoptera: Cucujidae) for America north of Mexico, with notes on other species. Insecta Mundi 2003, 17, 157–177. [Google Scholar]
- Lee, C.F.; Satô, M. A review of the genus Cucujus Fabricius (Insecta: Cucujoidea: Cucujidae) from Taiwan, Japan, and China, with descriptions of two new species and the larvae of Cucujus mniszechi Grouvelle. Zool. Stud. 2007, 46, 311–321. [Google Scholar]
- Lee, C.F.; Pütz, A. A new species of Cucujus Fabricius, 1775 from China and key to the east–Palaearctic species of the genus (Coleoptera: Cucujidae). Entomol. Z. 2008, 118, 211–213. [Google Scholar]
- Horák, J.; Chobot, K. Worldwide distribution of saproxylic beetles of the genus Cucujus Fabricius, 1775 (Coleoptera: Cucujidae). In Proceedings of the 5th Symposium and Workshop on the Conservation of Saproxylic Beetles, Lüneberg, Germany, 14–16 June 2008; Buse, J., Alexander, K., Ranius, T., Assmann, T., Eds.; Pensoft Publishers: Moscow, Russia, 2009; pp. 189–206. ISBN 97895464250722. [Google Scholar]
- Lee, J.; Thomas, M.C. Clarification of the taxonomic status of Cucujus clavipes with descriptions of the larvae of C. c. clavipes and C. c. puniceus (Coleoptera: Cucujidae). Fla. Entomol. 2011, 94, 145–150. [Google Scholar] [CrossRef]
- Bonacci, T.; Mazzei, A.; Horák, J.; Brandmayr, P. Cucujus tulliae sp. n.—An endemic Mediterranean saproxylic beetle from genus Cucujus Fabricius, 1775 (Coleoptera, Cucujidae), and keys for identification of adults and larvae native to Europe. ZooKeys 2012, 212, 63–79. [Google Scholar] [CrossRef]
- Marris, J.W.M.; Ślipiński, A. A revision of the Pediacus Shuckard 1839 (Coleoptera: Cucujidae) of Asia and Australasia. Zootaxa 2014, 3754, 32–58. [Google Scholar] [CrossRef]
- Bussler, H. Cucujus muelleri sp. n. aus den kaspischen Gebirgswaldern des Iran (Coleoptera: Cucujidae). Nachr. Der Bayer. Entomol. 2017, 66, 54–58. [Google Scholar]
- Marris, J.W.M. A revision of the flat bark beetle genus Platisus Erichson, 1842 (Coleoptera: Cucujidae). Austral Entomol. 2017, 56, 277–295. [Google Scholar] [CrossRef]
- Háva, J.; Zahradník, P.; Růžička, T. A new species of genus Cucujus Fabricius, 1775 (Coleoptera: Cucujidae) from China. Nat. Som. 2019, 33, 129–134. [Google Scholar] [CrossRef]
- Zhao, M.-Z.; Zhang, J.-K. Contribution to the knowledge of the genus Cucujus Fabricius (Coleoptera, Cucujidae) from China. Zootaxa 2019, 4544, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Zwick, A.; Ślipiński, A.; Marris, J.W.M.; Thomas, M.C.; Pang, H. A comprehensive phylogeny of flat bark beetles (Coleoptera: Cucujidae) with a revised classification and a new South American genus. Syst. Entomol. 2020, 45, 248–268. [Google Scholar] [CrossRef]
- Jaskuła, R.; Michalski, M.; Acal, D.A. First record of the family Cucujidae (Insecta: Coleoptera) from Vietnam with a checklist and a key to species currently known from Indochinese Peninsula. Orient. Insects 2020. [Google Scholar] [CrossRef]
- Fabricius, J.C. Species Insectorum, Exhibentes Eorum Differentias Specificas, Synonyma Auctorum, Loca Natalia, Metamorphosin, Adjectis Observationibus, Descriptionibus; Tom. 1. C. E. Bohnii, Hamburg-Kiel; NCSU Libraries: Raleigh, NC, USA, 1781; p. 566. [Google Scholar] [CrossRef] [Green Version]
- Mannercheim, G.C.G. Beitrag zur Kaefer-Fauna der Aleutischen Inseln, der Inseln Sitkha und Neu-Californiens. Bull. De La Société Impériale Des Nat. De Moscou 1843, 16, 175–314. [Google Scholar]
- DeLeon, D. An annotated list of the parasites, predators, and other associated fauna of the mountain pine beetle in western white pine and lodgepole pine. Can. Entomol. 1934, 66, 51–61. [Google Scholar] [CrossRef]
- Smith, D.B.; Sears, M.K. Mandibular structure and feeding habits of the morphologically similar coleopterous larvae: Cucujus clavipes (Cucujidae), Dendroides canadensis (Pyrochroidae) and Pytho depressus (Salpingidae). Can. Entomol. 1982, 114, 173–175. [Google Scholar] [CrossRef]
- Duman, J.G. Chance in overwintering mechanism of the cucujid beetle, Cucujus clavipes. J. Insect Physiol. 1984, 30, 235–239. [Google Scholar] [CrossRef]
- Kukal, J.G.; Duman, J.G. Switch in the overwintering strategy of two insect species and latitudinal differences in cold hardiness. Can. J. Zool. 1989, 67, 825–827. [Google Scholar] [CrossRef]
- Hammond, H.E.J. Arthropod biodiversity from Populus coarse woody material in north-central Alberta: A review of taxa and collection methods. Can. Entomol. 1997, 129, 1009–1033. [Google Scholar] [CrossRef]
- Hammond, J.H.E.; Langor, D.W.; Spence, J.R. Early colonization of Populus wood by saproxylic beetles (Coleoptera). Can. J. For. Res. 2001, 31, 1175–1183. [Google Scholar] [CrossRef]
- Kennedy, A.A.; McCullough, D.G. Phenology of the larger European pine shoot beetle Tomicus piniperda (L.) (Coleoptera: Scolytidae) in relation to native bark beetles and natural enemies in pine stands. Environ. Entomol. 2002, 31, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Bennett, V.A.; Sformo, T.; Walters, K.; Toien, O.; Jeannet, K.; Hochstrasser, R.; Pan, Q.; Serianni, A.S.; Barnes, B.M.; Duman, J.G. Comparative overwintering physiology of Alaska and Indiana populations of the beetle Cucujus clavipes (Fabricius): Roles of antifreeze proteins, polyols, dehydration and diapause. J. Exp. Biol. 2005, 208, 4467–4477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, J.M.; Spence, J.R.; Langor, D.W. Influence of boreal forest succession and dead wood qualities on saproxylic beetles. Agric. For. Entomol. 2007, 9, 3–16. [Google Scholar] [CrossRef]
- Dollin, P.E.; Majka, C.G.; Duinker, P.N. Saproxylic beetle (Coleoptera) communities and forest management practices in coniferous stands in southwestern Nova Scotia, Canada. ZooKeys 2008, 2, 291–336. [Google Scholar] [CrossRef] [Green Version]
- Majka, C.G. The flat bark beetles (Coleoptera: Silvanidae, Cucujidae, Laemophloeidae) of Atlantic Canada. ZooKeys 2008, 2, 221–238. [Google Scholar] [CrossRef]
- Sformo, T.; Walters, K.; Jeannet, K.; Wowk, B.; Fahy, G.M.; Barnes, B.M.; Duman, J.G. Deep supercooling, vitrification and limited survival to −100 °C in the Alaskan beetle Cucujus clavipes puniceus (Coleoptera: Cucujidae) larvae. J. Exp. Biol. 2010, 213, 502–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrasco, M.A.; Buechler, S.A.; Arnold, R.A.; Sformo, T.; Barnes, B.M.; Duman, J.G. Elucidating the biochemical overwintering adaptations of larval Cucujus clavipes puniceus, a nonmodel organism, via high throughput proteomics. J. Proteome Res. 2011, 10, 4634–4646. [Google Scholar] [CrossRef]
- Sformo, T.; McIntyre, J.; Walters, K.R., Jr.; Barnes, B.M.; Duman, J. Probability of freezing in the freeze-avoiding beetle larvae Cucujus clavipes puniceus (Coleoptera: Cucujidae) from interior Alaska. J. Insect Physiol. 2011, 57, 1170–1177. [Google Scholar] [CrossRef]
- Carrasco, M.A.; Buechler, S.A.; Arnold, R.A.; Sformo, T.; Barnes, B.M.; Duman, J.G. Investigating the deep supercooling ability of an Alaskan beetle, Cucujus clavipes puniceus, via high throughput proteomics. J. Proteome Res. 2012, 75, 1220–1234. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.M.; Duman, J.G. Upper lethal temperatures in three cold-tolerant insects are higher in winter than in summer. J. Exp. Biol. 2017, 220, 2726–2732. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, Y. A taxonomic study of Cucujus Fabricius, 1775 from Asia (Coleoptera: Cucujidae), with descriptions of new species and notes on morphological classification. Insect Syst. Evol. 2020. [Google Scholar] [CrossRef]
- Tozer, B.; Sandwell, D.T.; Smith, W.H.F.; Olson, C.; Beale, J.R.; Wessel, P. Global bathymetry and topography at 15 arc sec: SRTM15+. Earth Space Sci. 2019, 6, 1847–1864. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 20 January 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 20 January 2021).
- Ter Braak, C.J.F. CANOCO—A FORTRAN Program for Canonical Community Ordination by [Partial][Detrended][Canonical] Correspondence Analysis, Principal Components Analysis and Redundancy Analysis (Version 2.1); DLO Agricultural Mathematics Group: Wageningen, The Netherlands, 1987; p. 95. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002; p. 499. [Google Scholar]
- Brown, J.L. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. A maximum entropy approach to species distribution modeling. ICML ’04. In Proceedings of the Twenty-First International Conference on Machine Learning, Banff, Canada, 4–8 July 2004; ACM: New York, NY, USA, 2004; pp. 655–662. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1 km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hengl, T.; de Jesus, J.M.; MacMillan, R.A.; Batjes, N.H.; Heuvelink, G.B.M.; Ribeiro, E.; Samuel-Rosa, A.; Kempen, B.; Leenaars, J.G.B.; Walsh, M.G.; et al. SoilGrids1km—Global Soil Information Based on Automated Mapping. PLoS ONE 2014, 9, e105992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barve, N.; Barve, V.; Jimenez-Valverde, A.; Lira-Noriega, A.; Maher, S.P.; Peterson, A.T.; Soberón, J.; Villalobos, F. The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol. Mod. 2011, 222, 1810–1819. [Google Scholar] [CrossRef]
- O’Neill, B.C.; Kriegler, E.; Riahi, K.; Ebi, K.L.; Hallegatte, S.; Carter, T.R.; Mathur, R.; van Vuuren, D.P. A new scenario frame-work for climate change research: The concept of shared socioeconomic pathways. Clim. Chang. 2014, 122, 387–400. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.B.; Aneja, V.P.; Kang, D.; Arya, S.P. Modelling and analysis of the atmospheric nitrogen deposition in North Carolina. Int. J. Glob. Environ. Issues 2006, 6, 231–252. [Google Scholar] [CrossRef] [Green Version]
- Mason, S.J.; Graham, N.E. Areas beneath the relative operating characteristics (ROC) and relative operating levels (ROL) curves statistical significance and interpretation. Q. J. R. Meteorol. Soc. 2002, 128, 2145–2166. [Google Scholar] [CrossRef]
- Evangelista, P.H.; Kumar, S.; Stohlgren, T.J.; Jarnevich, C.S.; Crall, A.W.; Norman III, J.B.; Barnett, D.T. Modelling invasion for a habitat generalist and a specialist plant species. Divers Distrib. 2008, 14, 808–817. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Horák, J.; Chobot, K. Phenology and notes on the behaviour of Cucujus cinnaberinus: Points for understanding the conservation of the saproxylic beetle. North-West. J. Zool. 2011, 7, 352–355. [Google Scholar]
- Marczak, D. Zgniotek cynobrowy Cucujus cinnaberinus w Kampinoskim Parku Narodowym i uwagi do jego monitoringu. Studia i Materiały CEPL w Rogowie 2016, 49A, 142–152. [Google Scholar]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Amack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob. Chang. Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Menéndez, R. How are insects responding to global warming. Tijdschr. Voor Entomol. 2007, 150, 355–365. [Google Scholar]
- Lieth, H. (Ed.) Phenology and Seasonality Modeling. Ecological Studies Series; Springer: New York, NY, USA, 1974; p. 444. [Google Scholar]
- Tauber, M.J.; Tauber, C.A. Insect seasonality: Diapause maintenance, termination, and postdiapause development. Annu. Rev. Entomol. 1976, 21, 81–107. [Google Scholar] [CrossRef]
- Denno, R.F.; Dingle, H. Insect Life History Patterns: Habitat and Geographic Variation; Springer: New York, NY, USA, 1981; p. 225. [Google Scholar]
- Brown, V.K.; Hodek, I. (Eds.) Diapause and Life Cycle Strategies in Insects; W. Junk Publisher: Berlin, Germany, 1983; p. 283. [Google Scholar]
- Scott, J.A.; Epstein, M.E. Factors affecting phenology in a temperate insect community. Am. Midl. Nat. 1987, 117, 103–118. [Google Scholar] [CrossRef]
- Alonso, C. Variation in herbivory by Yponomeuta mahalebella on its only host plant Prunus mahaleb along an elevational gradient. Ecol. Entomol. 1999, 24, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Hodkinson, I.D. Terrestrial insects along elevations gradients. Biol. Rev. 2005, 80, 489–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazzari, S.M.N.; Lazzarotto, C.M. Distribuição altitudinal e sazonal de afídeos (Hemiptera, Aphididae) na Serra do Mar, Paraná, Brasil. Rev. Bras. De Zool. 2005, 22, 891–897. [Google Scholar] [CrossRef] [Green Version]
- Merrill, R.M.; Gutiérrez, D.; Lewis, O.T.; Gutiérrez, J.; Díez, S.B.; Wilson, R.J. Combined effects of climate and biotic interactions on the elevational range of a phytophagous insect. J. Anim. Ecol. 2008, 77, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Buchholz, L. Zgniotek cynobrowy Cucujus cinnaberinus (Scopoli, 1763). In Monitoring Gatunków Zwierząt. Przewodnik Metodyczny. Część Druga; Makomaska-Juchewicz, M., Baran, P., Eds.; Biblioteka Monitoringu Środowiska, Inspekcja Ochrony Środowiska: Kraków, Poland, 2012; pp. 419–446, 522. [Google Scholar]
- Parmesan, C.; Ryrholm, N.; Stefanescu, C.; Hill, J.K.; Thomas, C.D.; Descimon, H.; Huntley, B.; Kaila, L.; Kullberg, J.; Tammaru, T.; et al. Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature 1999, 399, 579–583. [Google Scholar] [CrossRef]
- Roy, D.B.; Sparks, T.H. Phenology of British butterflies and climate change. Glob. Chang. Biol. 2000, 6, 407–416. [Google Scholar] [CrossRef]
- Forister, M.L.; Shapiro, A.M. Climatic trends and advancing spring flight of butterflies in lowland California. Glob. Chang. Biol. 2003, 9, 1130–1135. [Google Scholar] [CrossRef]
- Bartomeus, I.; Ascher, J.S.; Wagner, D.; Danforth, B.N.; Colla, S.; Kornbluth, S.; Winfree, R. Climate-associated phenological advances in bee pollinators and bee-pollinated plants. Proc. Natl. Acad. Sci. USA 2011, 108, 20654–20659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamond, S.E.; Frame, A.M.; Martin, R.A.; Buckley, L.B. Species’ traits predict phenological responses to climate change in butterflies. Ecology 2011, 92, 1005–1012. [Google Scholar] [CrossRef]
- Guo, K.; Sun, O.J.; Kang, L. The responses of insects to global warming. In Recent Advances in Entomological Research; Liu, T., Kang, L., Eds.; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar] [CrossRef]
- Brooks, S.J.; Self, A.; Toloni, F.; Sparks, T. Natural history museum collections provide information on phenological change in British butterflies since the late-nineteenth century. Int. J. Biometeorol. 2014, 58, 1749–1758. [Google Scholar] [CrossRef]
- Kharouba, H.M.; Paquette, S.R.; Kerr, J.T.; Vellend, M. Predicting the sensitivity of butterfly phenology to temperature over the past century. Glob. Chang. Biol. 2014, 20, 504–514. [Google Scholar] [CrossRef]
- Bell, J.R.; Alderson, L.; Izera, D.; Kruger, T.; Parker, S.; Pickup, J.; Shortall, C.R.; Taylor, M.S.; Verrier, P.; Harrington, R. Long-term phenological trends, species accumulation rates, aphid traits and climate: Five decades of change in migrating aphids. J. Anim. Ecol. 2015, 84, 21–34. [Google Scholar] [CrossRef]
- Sato, Y.; Sato, S. Spring Temperature Predicts the Long-term Molting Phenology of Two Cicadas, Cryptotympana facialis and Graptopsaltria nigrofuscata (Hemiptera: Cicadidae). Ann. Entomol. Soc. Am. 2015, 108, 494–500. [Google Scholar] [CrossRef] [Green Version]
- Forrest, J.R.K. Complex responses of insect phenology to climate change. Curr. Opin. Insect Sci. 2016, 17, 49–54. [Google Scholar] [CrossRef]
- Thomsen, P.F.; Jørgensen, P.S.; Bruun, H.H.; Pedersen, J.; Riis-Nielsen, T.; Jonko, K.; Słowińska, I.; Rahbek, C.; Karsholt, O. Resource specialists lead local insect community turnover associated with temperature—Analysis of an 18-year full-seasonal record of moths and beetles. J. Anim. Ecol. 2016, 85, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Pureswaran, D.S.; Roques, A.; Battisti, A. Forest Insects and Climate Change. Curr. For. Rep. 2018, 4, 35–50. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, P.; Ammunét, T.; Barton, M.; Battisti, A.; Eigenbrode, S.D.; Jepsen, J.U.; Kalinkat, G.; Neuvonen, S.; Niemelä, P.; Terblanche, J.S.; et al. Complex responses of global insect pests to climate warming. Front. Ecol. Environ. 2020, 18, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.C. Family 82. Cucujidae Latreille 1802. In American Beetles: Polyphaga: Scarabaeoidea through Cuculionoidea; Arnett, R.H., Jr., Thomas, M.C., Skelley, P.E., Frank, J.H., Eds.; CRC Press: Boca Raton, FL, USA, 2002; Volume 2, pp. 329–330. [Google Scholar]
- Evans, A.V.; Hogue, J.N. Field Guide to Beetles of California, Flat Bark Beetles (Cucujidae); University of California Press: Berkeley, CA, USA; Los Angeles, CA, USA; London, UK, 2006. [Google Scholar]
- Horák, J.; Chobot, K.; Kohutka, A.; Gebauer, R. Possible factors influencing distribution of a threatened saproxylic beetle Cucujus cinnaberinus (Scopoli, 1763) (Coleoptera: Cucujidae). Coleopt. Bull. 2008, 62, 437–440. [Google Scholar] [CrossRef]
- Horák, J.; Vávrová, E.; Chobot, K. Habitat preferences influencing populations, distribution and conservation of the endangered saproxylic beetle Cucujus cinnaberinus (Coleoptera: Cucujidae) at the landscape level. Eur. J. Entomol. 2010, 107, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Mazzei, A.; Bonacci, T.; Contarini, E.; Zetto, T.; Brandmayr, P. Rediscovering the ‘umbrella species’ candidate Cucujus cinnaberinus (Scopoli, 1763) in Southern Italy (Coleoptera Cucujidae), and notes on bionomy. Ital. J. Zool. 2011, 78, 264–270. [Google Scholar] [CrossRef] [Green Version]
- Horák, J.; Chumanová, E.; Hilszczański, J. Saproxylic beetle thrives on the openness in management: A case study on the ecological requirements of Cucujus cinnaberinus from Central Europe. Insect Conserv. Divers. 2012, 5, 403–413. [Google Scholar] [CrossRef]
- Goczał, J.; Rossa, R. Dead wood complexity shapes the pattern of the occurrence of threatened saproxylic beetle Cucujus cinnaberinus. Pol. J. Ecol. 2017, 65, 158–165. [Google Scholar] [CrossRef]
- Vrezec, A.; Ambrožič, S.; Kobler, A.; Kapla, A.; de Groot, M. Cucujus cinnaberinus (Scopoli, 1763) at its terra typica in Slovenia: Historical overview, distribution patterns and habitat selection. Nat. Conserv. 2017, 19, 219–229. [Google Scholar] [CrossRef]
- Bełcik, M.; Goczał, L.; Ciach, M. Large-scale habitat model reveals a key role of large trees and protected areas in the metapopulation survival of the saproxylic specialist Cucujus Cinnaberinus. Biodivers. Conserv. 2018, 28, 3851–3871. [Google Scholar] [CrossRef] [Green Version]
- Bonacci, T.; Mazzei, A.; Naccarato, A.; Elliani, R.; Tagarelli, A.; Brandmayr, P. Beetles “in red”: Are the endangered flat bark beetles Cucujus cinnaberinus and C. haematodes chemically protected? (Coleoptera: Cucujidae). Eur. Zool. J. 2018, 85, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Bonacci, T.; Rovito, M.; Horák, J.; Brandmayr, P. Artificial Feeding and Laboratory Rearing of Endangered Saproxylic Beetles as a Tool for Insect Conservation. J. Insect Sci. 2020, 20, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Barnosky, A.D.; Matzke, N.; Tomiya, S.; Wogan, G.O.U.; Swarts, B.; Quental, T.B.; Marshall, C.; McGuire, J.L.; Lindsey, E.L.; Maguire, K.C.; et al. Has the Earth’s sixth mass extinction already arrived? Nature 2011, 471, 51–57. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human–induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [Green Version]
- Ceballos, G.; Ehrlich, P.R.; Dirzo, R. Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc. Natl. Acad. Sci. USA 2017, 114, E6089–E6096. [Google Scholar] [CrossRef] [Green Version]
- Hughes, L. Biological consequences of global warming: Is the signal already apparent? Trends Ecol. Evol. 2000, 15, 56–61. [Google Scholar] [CrossRef]
- Walther, G.-R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.-M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Walther, G.-R.; Beißner, S.; Burga, C.A. Trends in the upward shift of alpine plants. J. Veg. Sci. 2005, 16, 541–548. [Google Scholar] [CrossRef]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Ann. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef] [Green Version]
- Lenoir, J.; Gégout, J.C.; Marquet, P.A.; de Ruffray, P.; Brisse, H. A Significant Upward Shift in Plant Species Optimum Elevation During the 20th Century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef]
- Schweiger, O.; Biesmeijer, J.C.; Bommarco, R.; Hickler, T.; Hulme, P.E.; Klotz, S.; Kühn, I.; Moora, M.; Nielsen, A.; Ohlemüller, R.; et al. Multiple stressors on biotic interactions: How climate change and alien species interact to affect pollination. Biol. Rev. 2010, 85, 777–795. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Rasmont, P.; Franzén, M.; Lecocq, T.; Harpke, A.; Roberts, S.; Biesmeijer, J.; Castro, L.; Cederberg, B.; Dvorak, L.; Fitzpatrick, Ú.; et al. Climatic risk and distribution atlas of European bumblebees. BioRisk 2015, 10, 1–236. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef] [PubMed]
- Varela, S.; Lobo, J.M.; Hortal, J. Using species distribution models in paleobiogeography: A matter of data, predictors and concepts. Palaeogeogr. Palaeoclim.. Palaeoecol. 2011, 310, 451–463. [Google Scholar] [CrossRef]
- Lima-Ribeiro, M.S.; Varela, S.; Nógues-Bravo, D.; Diniz-Filho, J.A.F. Potential suitable areas of Giant Ground Sloths dropped before its extinction in South America: The evidences from bioclimatic envelope modelling. Nat. Conserv. 2012, 10, 145–151. [Google Scholar] [CrossRef]
- Barrientos, R.; Kvist, L.; Barbosa, A.; Valera, F.; Khoury, F.; Varela, S.; Moreno, E. Refugia, colonization and diversification of an arid-adapted bird: Coincident patterns between genetic data and ecological niche modelling. Mol. Ecol. 2014, 23, 390–407. [Google Scholar] [CrossRef] [PubMed]
- Kukwa, M.; Kolanowska, M. Glacial refugia and the prediction of future habitat coverage of the South American lichen species Ochrolechia austroamericana. Sci. Rep. 2016, 6, 38779. [Google Scholar] [CrossRef] [Green Version]
- Biella, P.; Bogliani, G.; Cornalba, M.; Manino, A.; Neumayer, J.; Porporato, M.; Rasmont, P.; Milanesi, P. Distribution patterns of the cold adapted bumblebee Bombus alpinus in the Alps and hints of an uphill shift (Insecta: Hymenoptera: Apidae). J. Insect Conserv. 2017, 21, 357–366. [Google Scholar] [CrossRef]
- Wilson, C.D.; Roberts, D.; Reid, N. Applying species distribution modeling to identify areas of high conservation value for endangered species: A case study using Margaritifera margaritifera (L.). Biol. Conserv. 2011, 144, 821–829. [Google Scholar] [CrossRef]
- Koch, R.; Almeida-Cortez, J.S.; Kleinschmit, B. Revealing areas of high nature conservation importance in a seasonally dry tropical forest in Brazil: Combination of modelled plant diversity hot spots and threat patterns. J. Nat. Conserv. 2017, 35, 24–39. [Google Scholar] [CrossRef]
- Spiers, J.A.; Oatham, M.P.; Rostant, L.V.; Farrell, A.D. Applying species distribution modelling to improving conservation based decisions: A gap analysis of Trinidad and Tobago’s endemic vascular plants. Biodivers. Conserv. 2018, 27, 2931–2949. [Google Scholar] [CrossRef]
- Nóbrega, C.C.; De Marco, P., Jr. Unprotecting the rare species: A niche-based gap analysis for odonates in a core Cerrado area. Divers. Distrib. 2011, 17, 491–505. [Google Scholar] [CrossRef]
- Silva, D.P.; Aguiar, A.J.C.; Melo, G.A.R.; Anjos-Silva, E.J.; De Marco, P., Jr. Amazonian species within the Cerrado savanna: New records and potential distribution for Aglae caerulea (Apidae: Euglossini). Apidologie 2013, 44, 673–683. [Google Scholar] [CrossRef]
- Estay, S.A.; Labra, F.A.; Sepulveda, R.D.; Bacigalupe, L.D. Evaluating Habitat Suitability for the Establishment of Monochamus spp. through Climate-Based Niche Modeling. PLoS ONE 2014, 9, e102592. [Google Scholar] [CrossRef] [PubMed]
- Pérez-De la, O.N.B.; Espinosa-Zaragoza, S.; López-Martínez, V.; Hight, S.D.; Varone, L. Ecological niche modeling to calculate ideal sites to introduce a natural enemy: The case of Apanteles opuntiarum (Hymenoptera: Braconidae) to control Cactoblastis cactorum (Lepidoptera: Pyralidae) in North America. Insects 2020, 11, 454. [Google Scholar] [CrossRef] [PubMed]
- Raxworthy, C.; Martinez-Meyer, E.; Horning, N.; Nussbaum, R.A.; Schneider, G.E.; Ortega-Huerta, M.A.; Peterson, A.T. Predicting distributions of known and unknown reptile species in Madagascar. Nature 2003, 426, 837–841. [Google Scholar] [CrossRef] [Green Version]
- Silva, D.P.; Andrade, A.F.A.; Oliveira, J.P.J.; Morais, D.M.; Vieira, J.E.A.; Engel, M.S. Current and future ranges of an elusive North American insect using species distribution models. J. Insect Conserv. 2019, 23, 175–186. [Google Scholar] [CrossRef]
- Rödder, D.; Schmidtlein, S.; Veith, M.; Lötters, S. Alien invasive slider turtle in unpredicted habitat: A matter of niche shift or of predictors studied? PLoS ONE 2009, 4, e7843. [Google Scholar] [CrossRef] [PubMed]
- Gallien, L.; Muenkemueller, T.; Albert, C.H.; Boulangeat, I.; Thuiller, W. Predicting potential distributions of invasive species: Where to go from here? Divers. Distrib. 2010, 16, 331–342. [Google Scholar] [CrossRef]
- Hortal, J.; Roura-Pascual, N.; Sanders, N.J.; Rahbek, C. Understanding (insect) species distributions across spatial scales. Ecography 2010, 33, 51–53. [Google Scholar] [CrossRef]
- Kolanowska, M.; Konowalik, K. Niche conservatism and future changes in the potential area coverage of Arundina graminifolia, an invasive orchid species from Southeast Asia. Biotropica 2014, 46, 157–165. [Google Scholar] [CrossRef]
- Olivera, L.; Minghetti, E.; Montemayor, S. Ecological niche modeling (ENM) of Leptoglossus clypealis a new potential global invader: Following in the footsteps of Leptoglossus occidentalis? Bull. Entomol. Res. 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.C.; Silva, D.P.; De Marco Jr, P.; Melo, G.A.R. Species conservation under future climate change: The case of Bombus bellicosus, a potentially threatened South American bumblebee species. J. Insect Conserv. 2015, 19, 33–43. [Google Scholar] [CrossRef]
- LeConte, J.L. Synopsis of the Cucujides of the United States. Proc. Acad. Nat. Sci. Phila. 1854, 7, 73–79. [Google Scholar]
- LeConte, J.L. Classification of the Coleoptera of North America. Prepared for the Smithsonian Institution. Part 1. Smithson. Misc. Collect. 1861, 136, 286. [Google Scholar]
- LeConte, J.L. List of the Coleoptera of North America. Prepared for the Smithsonian Institution. Part 1. Smithson. Misc. Collect. 1863, 140, 77. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| bio19 | 0.496 | 0.550 | 0.640 | 0.415 | 0.591 | 0.179 | 0.465 | 0.433 | 0.549 | 0.599 | 0.318 | 0.930 | 0.898 | 0.859 | 0.390 | 0.910 | 0.873 | 0.720 | x |
| bio18 | 0.553 | 0.826 | 0.824 | 0.760 | 0.848 | 0.252 | 0.802 | 0.819 | 0.380 | 0.845 | 0.353 | 0.916 | 0.913 | 0.859 | 0.691 | 0.912 | 0.870 | x | |
| bio17 | 0.527 | 0.636 | 0.684 | 0.537 | 0.686 | 0.165 | 0.582 | 0.582 | 0.457 | 0.701 | 0.291 | 0.951 | 0.866 | 0.997 | 0.394 | 0.875 | x | ||
| bio16 | 0.561 | 0.757 | 0.805 | 0.656 | 0.785 | 0.245 | 0.704 | 0.688 | 0.490 | 0.782 | 0.368 | 0.979 | 0.997 | 0.857 | 0.657 | x | |||
| bio15 | 0.444 | 0.863 | 0.820 | 0.894 | 0.827 | 0.244 | 0.908 | 0.766 | 0.342 | 0.773 | 0.347 | 0.556 | 0.672 | 0.376 | x | ||||
| bio14 | 0.509 | 0.619 | 0.664 | 0.527 | 0.668 | 0.152 | 0.569 | 0.564 | 0.441 | 0.683 | 0.274 | 0.938 | 0.849 | x | |||||
| bio13 | 0.567 | 0.763 | 0.810 | 0.661 | 0.790 | 0.258 | 0.709 | 0.698 | 0.492 | 0.787 | 0.381 | 0.973 | x | ||||||
| bio12 | 0.568 | 0.724 | 0.774 | 0.618 | 0.764 | 0.215 | 0.667 | 0.660 | 0.490 | 0.770 | 0.343 | x | |||||||
| bio11 | 0.836 | 0.411 | 0.539 | 0.123 | 0.487 | 0.838 | 0.199 | 0.535 | 0.779 | 0.555 | x | ||||||||
| bio10 | 0.806 | 0.951 | 0.944 | 0.760 | 0.991 | 0.364 | 0.833 | 0.936 | 0.612 | x | |||||||||
| bio9 | 0.816 | 0.508 | 0.625 | 0.188 | 0.569 | 0.551 | 0.275 | 0.441 | x | ||||||||||
| bio8 | 0.725 | 0.891 | 0.860 | 0.744 | 0.923 | 0.381 | 0.803 | x | |||||||||||
| bio7 | 0.398 | 0.919 | 0.829 | 0.990 | 0.891 | 0.102 | x | ||||||||||||
| bio6 | 0.592 | 0.234 | 0.355 | 0.056 | 0.308 | x | |||||||||||||
| bio5 | 0.741 | 0.981 | 0.958 | 0.826 | x | ||||||||||||||
| bio4 | 0.297 | 0.856 | 0.753 | x | |||||||||||||||
| bio3 | 0.751 | 0.961 | x | ||||||||||||||||
| bio2 | 0.656 | x | |||||||||||||||||
| bio1 | x | ||||||||||||||||||
| bio1 | bio2 | bio3 | bio4 | bio5 | bio6 | bio7 | bio8 | bio9 | bio10 | bio11 | bio12 | bio13 | bio14 | bio15 | bio16 | bio17 | bio18 | bio19 |
| Cucujus clavipes clavipes | Cucujus clavipes puniceus | ||
|---|---|---|---|
| Variable | Percent Contribution | Variable | Percent Contribution |
| bio1 | 44.3 | bio1 | 35.9 |
| bio12 | 23.9 | bio9 | 20.2 |
| srad07 | 7.8 | land cover | 10.6 |
| srad12 | 7.3 | bio10 | 10.4 |
| bio6 | 4.3 | soil class | 5.6 |
| Cucujus clavipes clavipes | |||||
| scenario | −1 (range expansion) | 0 (absent in both) | 1 (present in both) | 2 (range contraction) | Change |
| ssp126 | 92,386.29 | 19,997,564 | 204,237.6 | 788,769.7 | −70.13% |
| ssp245 | 518,292.9 | 19,571,658 | 192,326.2 | 800,681.2 | −28.44% |
| ssp370 | 569,134.4 | 19,520,816 | 159,206.6 | 833,800.8 | −26.65% |
| ssp585 | 580,755.3 | 19,509,195 | 133,059.5 | 859,947.8 | −28.12% |
| Cucujus clavipes puniceus | |||||
| scenario | −1 (range expansion) | 0 (absent in both) | 1 (present in both) | 2 (range contraction) | Change |
| ssp126 | 109,236.6 | 20,422,890 | 210,338.6 | 340,492.9 | −41.98% |
| ssp245 | 161,530.7 | 20,370,596 | 175,766.4 | 375,065.1 | −38.77% |
| ssp370 | 213,824.9 | 20,318,301 | 191,454.6 | 359,376.9 | −26.42% |
| ssp585 | 218,763.8 | 20,313,363 | 72,630.73 | 478,200.7 | −47.10% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaskuła, R.; Kolanowska, M.; Michalski, M.; Schwerk, A. From Phenology and Habitat Preferences to Climate Change: Importance of Citizen Science in Studying Insect Ecology in the Continental Scale with American Red Flat Bark Beetle, Cucujus clavipes, as a Model Species. Insects 2021, 12, 369. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12040369
Jaskuła R, Kolanowska M, Michalski M, Schwerk A. From Phenology and Habitat Preferences to Climate Change: Importance of Citizen Science in Studying Insect Ecology in the Continental Scale with American Red Flat Bark Beetle, Cucujus clavipes, as a Model Species. Insects. 2021; 12(4):369. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12040369
Chicago/Turabian StyleJaskuła, Radomir, Marta Kolanowska, Marek Michalski, and Axel Schwerk. 2021. "From Phenology and Habitat Preferences to Climate Change: Importance of Citizen Science in Studying Insect Ecology in the Continental Scale with American Red Flat Bark Beetle, Cucujus clavipes, as a Model Species" Insects 12, no. 4: 369. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12040369