
Alternative Methods of Ant (Hymenoptera: Formicidae) Control with Emphasis on the Argentine Ant, Linepithema humile


{kind=link}
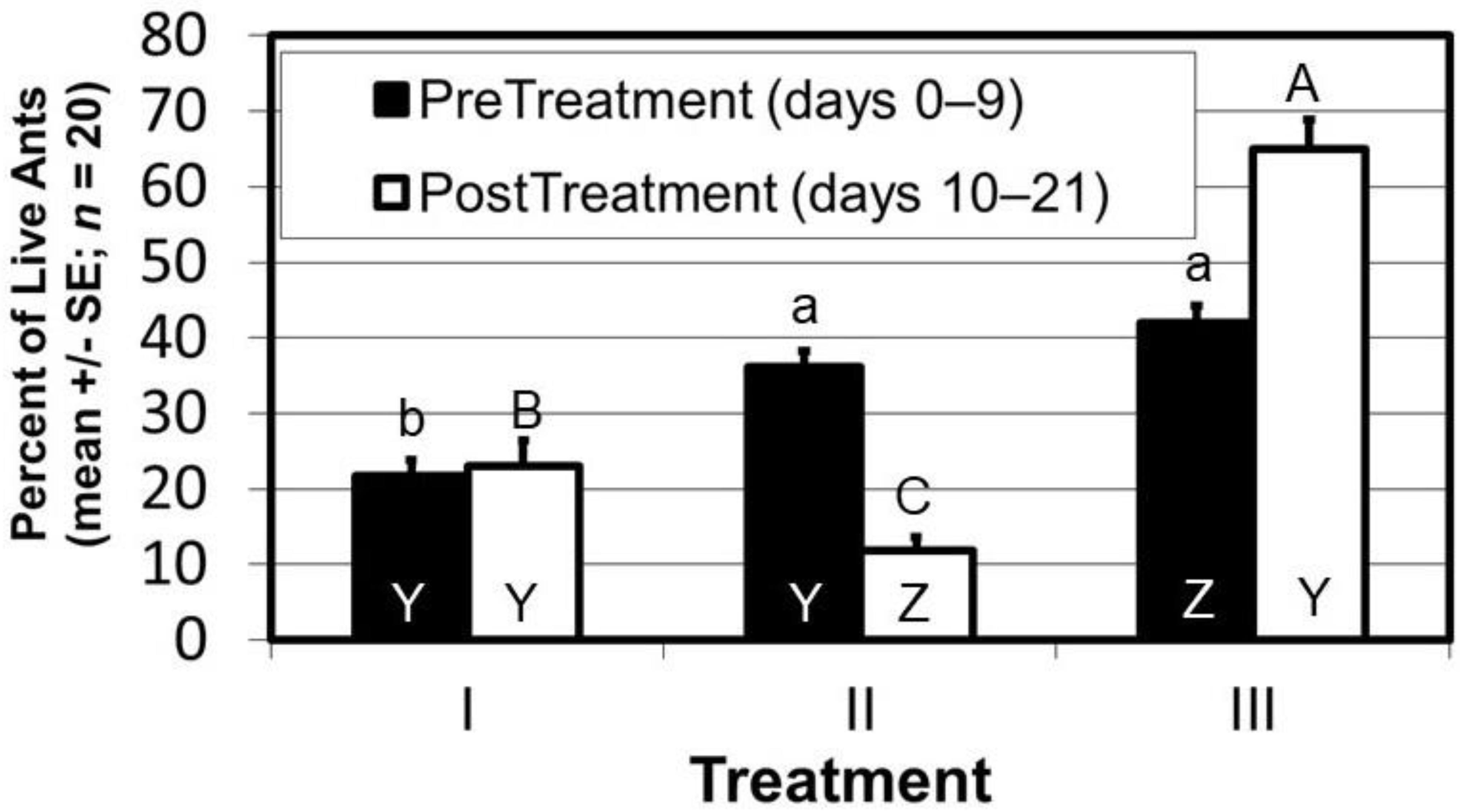{kind=link}
Abstract
:Simple Summary
Abstract
1. Pest Ant Control around Homes
2. Standard Ant Control Methods Can Be Problematic
3. Alternative Methods of Ant Control
3.1. Trail Pheromone
3.2. Use of Behavior-Modifying Chemicals
3.3. Mass Trapping
3.4. Hydrogels, “Virtual” Baiting, and Exceedingly-Low Bait Concentrations
3.5. Food Source Reduction
3.6. Deterrents
3.7. RNA Interference (RNAi)
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vega, S.J.; Rust, M.K. The Argentine ant: A significant invasive species in agricultural, urban and natural environments. Sociobiology 2001, 37, 3–25. [Google Scholar]
- Rust, M.; Reierson, D.A.; Klotz, J.H. Pest management of Argentine ants (Hymenoptera: Formicidae). J. Entomol. Sci. 2003, 38, 159–169. [Google Scholar] [CrossRef]
- Klotz, J.H.; Rust, M.K.; Costa, H.S.; Reierson, D.A.; Kido, K. Strategies for controlling Argentine ants (Hymenoptera: Formicidae) with sprays and baits. J. Agric. Urban Entomol. 2002, 19, 85–94. [Google Scholar]
- Klotz, J.H.; Rust, M.K.; Greenberg, L.; Field, H.C.; Kupfer, K. An evaluation of several urban pest management strategies to control Argentine ants (Hymenoptera: Formicidae). Sociobiology 2007, 50, 391–398. [Google Scholar]
- Klotz, J.H.; Rust, M.K.; Field, H.C.; Greenberg, L.; Kupfer, K. Controlling Argentine ants in residential settings (Hymenoptera: Formicidae). Sociobiology 2008, 51, 579–588. [Google Scholar]
- Klotz, J.H.; Rust, M.K.; Field, H.C.; Greenberg, L.; Kupfer, K. Low impact directed sprays and liquid baits to control Argentine ants (Hymenoptera: Formicidae). Sociobiology 2009, 54, 101–108. [Google Scholar]
- Klotz, J.H.; Rust, M.K.; Greenberg, L.; Robertson, M.A. Developing low risk strategies for Argentine ants (Hymenoptera: Formicidae). Sociobiology 2010, 55, 779–785. [Google Scholar]
- Environmental Hazard and General Labeling for Pyrethroid and Synergized Pyrethrins Non-Agricultural Outdoor Products; U.S. Environmental Protection Agency: Washington, DC, USA, 2013. Available online: https://www.epa.gov/ingredients-used-pesticide-products/environmental-hazard-and-general-labeling-pyrethroid-and (accessed on 5 April 2021).
- Greenberg, L.; Rust, M.K. Chapter 23: Ant control and insecticide runoff around urban homes. In Pesticides in Surface Water: Monitoring Modeling, Risk Assessment, and Management; Goh, K.S., Gan, J., Young, D.F., Luo, Y., Eds.; American Chemical Society: Washington, DC, USA, 2019; pp. 451–473. [Google Scholar] [CrossRef]
- Greenberg, L.; Rust, M.K.; Klotz, J.H.; Haver, D.; Kabashima, J.N.; Bondarenko, S.; Gan, J. Impact of ant control technologies on insecticide runoff and efficacy. Pest Manag. Sci. 2010, 66, 980–987. [Google Scholar] [CrossRef]
- Jiang, W.; Soeprono, A.; Rust, M.K.; Gan, J. Ant control efficacy of pyrethroids and fipronil on outdoor concrete surfaces. Pest Manag. Sci. 2014, 70, 271–277. [Google Scholar] [CrossRef]
- Gan, J.; Bondarenko, S.; Oki, L.; Haver, D.; Li, J.X. Occurrence of fipronil and its biologically active derivatives in urban residential runoff. Environ. Sci. Technol. 2012, 46, 1489–1495. [Google Scholar] [CrossRef]
- Klotz, J.H.; Greenberg, L.; Storey, H.H.; Williams, D.F. Alternative control strategies for ants around homes. J. Entomol. Sci. 1997, 14, 249–257. [Google Scholar]
- Suckling, D.; Peck, R.; Manning, L.; Stringer, L.; Cappadonna, J.; El-Sayed, A. Pheromone disruption of Argentine ant trail integrity. J. Chem. Ecol. 2008, 34, 1602–1609. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Nishisue, K.; Sunamura, E.; Suzuki, S.; Sakamoto, H.; Fukumoto, T.; Terayama, M.; Tatsuki, S. Trail-following disruption in the invasive Argentine ant with a synthetic trail pheromone component (Z)-9-hexadecenal. Sociobiology 2009, 54, 139–152. [Google Scholar]
- Suckling, D.M.; Peck, R.W.; Stringer, L.D.; Snook, K.; Banko, P.C. Trail pheromone disruption of Argentine ant trail formation and foraging. J. Chem. Ecol. 2010, 36, 122–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishisue, K.; Sunamura, E.; Tanaka, Y.; Sakamoto, H.; Suzuki, S.; Fukumoto, T.; Terayama, M.; Tatsuki, S. Long-term field trial to control the invasive Argentine ant (Hymenoptera: Formicidae) with synthetic trail pheromone. J. Econ. Entomol. 2010, 103, 1784–1789. [Google Scholar] [CrossRef]
- Suckling, D.M.; Stringer, L.D.; Corn, J.E. Argentine ant trail pheromone disruption is mediated by trail concentration. J. Chem. Ecol. 2011, 37, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Sunamura, E.; Suzuki, S.; Nishisue, K.; Sakamoto, H.; Otsuka, M.; Utsumi, Y.; Mochizuki, F.; Fukumoto, T.; Ishikawa, Y.; Terayama, M.; et al. Combined use of a synthetic trail pheromone and insecticidal bait provides effective control of an invasive ant. Pest Manag. Sci. 2011, 67, 1230–1236. [Google Scholar] [CrossRef] [PubMed]
- Choe, D.H.; Villafuerte, D.B.; Tsutsui, N.D. Trail pheromone of the Argentine ant, Linepithema humile (Mayr) (Hymenoptera: Formicidae). PLoS ONE 2012, 7, e45016. [Google Scholar] [CrossRef] [PubMed]
- Westermann, F.L.; Suckling, D.M.; Lester, P.J. Disruption of foraging by a dominant invasive species to decrease its competitive ability. PLoS ONE 2014, 9, e90173. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, L.; Klotz, J.H. Argentine ant (Hymenoptera: Formicidae) trail pheromone enhances consumption of liquid sucrose solution. J. Econ. Entomol. 2000, 93, 119–122. [Google Scholar] [CrossRef]
- Welzel, K.F.; Choe, D.H. Development of a pheromone-assisted baiting technique for Argentine ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2016, 109, 1303–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choe, D.; Hwan, K.T.; Lopez, C.M.; Campbell, K. Pheromone-assisted techniques to improve the efficacy of insecticide sprays against Linepithema humile (Hymenoptera: Formicidae). J. Econ. Entomol. 2014, 107, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Soeprono, A.M.; Rust, M.K. Effect of horizontal transfer of barrier insecticides to control Argentine ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2004, 97, 1675–1681. [Google Scholar] [CrossRef]
- Choe, D.H.; Rust, M.K. Horizontal transfer of insecticides in laboratory colonies of the Argentine ant (Hymenoptera: Formicidae). J. Econ. Entomol. 2008, 101, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Wiltz, B.; Suiter, D.; Gardner, W. Activity of bifenthrin, chlorfenapyr, fipronil, and thiamethoxam against Argentine ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2009, 102, 2279–2288. [Google Scholar] [CrossRef] [PubMed]
- Choe, D.H.; Millar, J.G.; Rust, M.K. Chemical signals associated with life inhibit necrophoresis in Argentine ants. Proc. Natl. Acad. Sci. USA 2009, 106, 8251–8255. [Google Scholar] [CrossRef] [Green Version]
- Wilson, E.O.; Durlach, N.I.; Roth, L.M. Chemical releaser of necrophoric behavior in ants. Psyche 1958, 65, 108–114. [Google Scholar] [CrossRef]
- Gochnour, B.M.; Suiter, D.R.; Davis, J.W.; Huang, Q. Use of an Argentine ant, Linepithema humile, semiochemical to deliver an acute toxicant. Insects 2018, 9, 171. [Google Scholar] [CrossRef] [Green Version]
- Wiltz, B.A.; Suiter, D.R.; Gardner, W.A.; Berisford, C.W. A novel delivery method for ant (Hymenoptera: Formicidae) toxicants. Midsouth Entomol. 2010, 3, 79–88. [Google Scholar]
- Buczkowski, G. The Trojan horse approach for managing invasive ants: A study with Asian needle ants, Pachycondyla chinensis. Biol. Invasions 2016, 18, 507–515. [Google Scholar] [CrossRef]
- Buczkowski, G. Prey-baiting as a conservation tool: Selective control of invasive ants with minimal non-target effects. Insect Conserv. Divers. 2017, 10, 302–309. [Google Scholar] [CrossRef]
- Buczkowski, G. Trap-treat-release: Horizontal transfer of fipronil in field colonies of black carpenter ants, Camponotus pennsylvanicus. Pest Manag. Sci. 2019, 75, 2195–2201. [Google Scholar] [CrossRef] [PubMed]
- Buczkowski, G.; Mothapo, N.P.; Wossler, T.C. Let them eat termites-prey-baiting provides effective control of Argentine ants, Linepithema humile, in a biodiversity hotspot. J. Appl. Entomol. 2017, 1–9. [Google Scholar] [CrossRef]
- Newell, W.; Barber, T. The Argentine ant. US Dep. Agric. Bur. Entomol. Bull. 1913, 122, 3–98. [Google Scholar]
- Gordon, D.M.; Moses, L.; Falkovitz-Halpern, M.; Wong, E.H. Effect of weather on infestation of buildings by the invasive Argentine ant, Linepithema humile (Hymenoptera: Formicidae). Am. Midl. Nat. 2001, 146, 321–328. [Google Scholar] [CrossRef]
- Schilman, P.E.; Lighton, J.R.B.; Holway, D.A. Water balance in the Argentine ant (Linepithema humile) compared with five common native ant species from southern California. Physiol. Entomol. 2007, 32, 1–7. [Google Scholar] [CrossRef]
- Walters, A.C.; Mackay, D.A. An experimental study of the relative humidity preference and survival of the Argentine ant, Linepithema humile (Hymenoptera, Formicidae): Comparisons with a native Iridomyrmex species in South Australia. Insectes Soc. 2003, 50, 355–360. [Google Scholar] [CrossRef]
- Heller, N.E.; Gordon, D.M. Seasonal spatial dynamics and causes of nest movement in colonies of the invasive Argentine ant (Linepithema humile). Ecol. Entomol. 2006, 31, 499–510. [Google Scholar] [CrossRef]
- Silverman, J.; Nsimba, B. Soil-free collection of Argentine ants (Hymenoptera: Formicidae) based on food-directed brood and queen movement. Fla. Entomol. 2000, 83, 10–16. [Google Scholar] [CrossRef]
- Scocco, C.; Suiter, D.; Gardner, W. Repellency of five essential oils to Linepithema humile (Hymenoptera: Formicidae). J. Entomol. Sci. 2012, 47, 150–159. [Google Scholar] [CrossRef]
- Wiltz, B.; Suiter, D.; Gardner, W. Deterrency and toxicity of essential oils to Argentine and red imported fire ants (Hymenoptera: Formicidae). J. Entomol. Sci. 2007, 42, 239–249. [Google Scholar] [CrossRef]
- Silverman, J.; Sorenson, C.E.; Waldvogel, M.G. Trap mulching Argentine ants. J. Econ. Entomol. 2006, 99, 1757–1760. [Google Scholar] [CrossRef] [PubMed]
- Enzmann, B.L.; Kepheim, K.M.; Wang, T.B.; Nonacs, P. Giving them what they want: Manipulating Argentine ant activity patterns with water. J. Appl. Entomol. 2012, 136, 588–595. [Google Scholar] [CrossRef]
- Buczkowski, G.; Roper, E.; Chin, D. Polyacrylamide hydogels: An effective tool for delivering liquid baits to pest ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2014, 107, 748–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buczkowski, G.; Roper, E.; Chin, D.; Mothapo, N.; Wossler, T. Hydrogel baits with low-dose thiamethoxam for sustainable Argentine ant management in commercial orchards. Entomol. Exp. Appl. 2014, 153, 183–190. [Google Scholar] [CrossRef]
- Rust, M.K.; Soeprono, A.; Wright, S.; Greenberg, L.; Choe, D.H.; Boser, C.L.; Cory, C.; Hanna, C. Laboratory and field evaluations of polyacrylamide hydrogel baits against Argentine ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2015, 108, 1228–1236. [Google Scholar] [CrossRef]
- Tay, J.W.; Hoddle, M.S.; Mulchandani, A.; Choe, D.H. Development of an alginate hydrogel to deliver aquaeous bait for pest ant management. Pest Manag. Sci. 2017, 73, 2028–2038. [Google Scholar] [CrossRef]
- Tay, J.W.; Choe, D.H.; Mulchandani, A.; Rust, M.K. Hydrogels: From controlled release to a new bait delivery for insect pest management. J. Econ. Entomol. 2020, 113, 2061–2068. [Google Scholar] [CrossRef]
- Milosavljevic, I.; Hoddle, M.S. Laboratory screening of selected synthetic and organic insecticides for efficacy against Argentine ants when incorporated into alginate hydrogel beads, 2021. Arthropod Manag. Tests 2021, 46, 1–3. [Google Scholar] [CrossRef]
- McCalla, K.A.; Tay, J.W.; Mulchandani, A.; Choe, D.H.; Hoddle, M.S. Biodegradable alginate hydrogel bait delivery system effectively controls high-density populations of Argentine ant in commercial citrus. J. Pest Sci. 2020, 93, 1031–1042. [Google Scholar] [CrossRef]
- Boser, C.L.; Hanna, C.; Faulkner, K.R.; Cory, C.; Randall, J.M.; Morrison, S.A. Argentine ant management in conservation areas: Results of a pilot study. Monogr. West. N. Am. Nat. 2014, 7, 518–530. [Google Scholar] [CrossRef] [Green Version]
- Choe, D.H.; Vetter, R.S.; Rust, M.K. Development of virtual bait stations to control Argentine ants (Hymenoptera: Formicidae) in environmentally sensitive habitats. J. Econ. Entomol. 2010, 103, 1761–1769. [Google Scholar] [CrossRef] [PubMed]
- Brightwell, R.; Silverman, J. Effects of honeydew-producing hemipteran denial on local Argentine ant distribution and boric acid bait performance. J. Econ. Entomol. 2009, 102, 1170–1174. [Google Scholar] [CrossRef]
- Brightwell, R.; Bambara, S.B.; Silverman, J. Combined effect of hemipteran control and liquid bait on Argentine ant populations. J. Econ. Entomol. 2010, 103, 1790–1796. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [Green Version]
- Regnault-Roger, C.; Philogene, B.J.R. Past and current prospects for the use of botanicals and plant allelochemicals in integrated pest management. Pharm. Biol. 2008, 46, 41–52. [Google Scholar] [CrossRef]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential oils in insect control: Low-risk products in a high-stakes world. Annu. Rev. Entomol. 2012, 37, 405–424. [Google Scholar] [CrossRef]
- Guerra, M.D.; Suiter, D.R.; Scocco, C.M. Topical toxicity of nine essential oils to Camponotus pennsylvanicus (Hymenoptera: Formicidae). Sociobiology 2011, 58, 419–426. [Google Scholar]
- Appel, A.G.; Gehret, M.J.; Tanley, M.J. Repellency and toxicity of mint oil granules to red imported fire ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2004, 97, 575–580. [Google Scholar] [CrossRef]
- Vogt, J.T.; Shelton, T.G.; Merchant, M.E.; Russell, S.A.; Tanley, M.J.; Appel, A.G. Efficacy of three citrus oil formulations against Solenopsis invicta Buren (Hymenoptera: Formicidae), the red imported fire ant. J. Agric. Urban Entomol. 2002, 19, 159–171. [Google Scholar]
- Meissner, H.E.; Silverman, J. Effects of aromatic cedar mulch on the Argentine ant and the odorous house ant (Hymenoptera: Formicidae). J. Econ. Entomol. 2001, 94, 1526–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meissner, H.E.; Silverman, J. Effect of aromatic cedar mulch on Argentine ant (Hymenoptera: Formicidae) foraging activity and nest establishment. J. Econ. Entomol. 2003, 96, 850–855. [Google Scholar] [CrossRef] [PubMed]
- Sisk, C.B.; Shorey, H.H.; Gerber, R.G.; Gaston, L.K. Semiochemicals that disrupt foraging by the Argentine ant (Hymenoptera: Formicidae): Laboratory bioassays. J. Econ. Entomol. 1996, 89, 381–385. [Google Scholar] [CrossRef]
- Shorey, H.H.; Gaston, L.K.; Gerber, R.G.; Phillips, P.A.; Wood, D.L. Disruption of foraging by Argentine ants, Iridomyrmex humilis (Mayr) (Hymenoptera: Formicidae), in citrus trees through the use of semiochemicals and related chemicals. J. Chem. Ecol. 1992, 18, 2131–2141. [Google Scholar] [CrossRef]
- Shorey, H.H.; Gaston, L.K.; Gerber, R.G.; Sisk, C.B.; Phillips, P.A. Formulating farnesol and other ant-repellent semiochemicals for exclusion of Argentine ants (Hymenoptera: Formicidae) from citrus trees. Environ. Entomol. 1996, 25, 114–119. [Google Scholar] [CrossRef]
- Sen, G.; Blau, H. A brief history of RNAi: The silence of the genes. FASEB J. 2006, 20, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Welzel, K.F. Exploration of Argentine ant (Linepithema humile) Biology for Pest Management. Ph.D. Thesis, University of California Riverside, Riverside, CA, USA, September 2017. Available online: https://escholarship.org/uc/item/2fn009vr (accessed on 23 May 2021).
- Meng, J.; Lei, J.; Davitt, A.; Holt, J.R.; Huang, J.; Gold, R.; Vargo, E.L.; Tarone, A.M.; Zhu-Salzman, K. Suppressing tawny crazy ant (Nylanderia fulva) by RNAi technology. Insect Sci. 2020, 27, 113–121. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suiter, D.R.; Gochnour, B.M.; Holloway, J.B.; Vail, K.M. Alternative Methods of Ant (Hymenoptera: Formicidae) Control with Emphasis on the Argentine Ant, Linepithema humile. Insects 2021, 12, 487. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12060487
Suiter DR, Gochnour BM, Holloway JB, Vail KM. Alternative Methods of Ant (Hymenoptera: Formicidae) Control with Emphasis on the Argentine Ant, Linepithema humile. Insects. 2021; 12(6):487. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12060487
Chicago/Turabian StyleSuiter, Daniel R., Benjamin M. Gochnour, Jacob B. Holloway, and Karen M. Vail. 2021. "Alternative Methods of Ant (Hymenoptera: Formicidae) Control with Emphasis on the Argentine Ant, Linepithema humile" Insects 12, no. 6: 487. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12060487