
Elevated CO2 Impacts on Plant–Pollinator Interactions: A Systematic Review and Free Air Carbon Enrichment Field Study


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Systematic Review
2.2. Field Experiment
2.2.1. Location
2.2.2. Plant Study System
2.2.3. Environmental Data
2.2.4. Flowering Phenology
2.2.5. Insect Visitation Surveys
2.2.6. Seed Counts
2.2.7. Statistical Analyses
3. Results
3.1. Systematic Review
3.2. BIFOR FACE Field Experiment
3.2.1. Environmental Parameters
3.2.2. Bluebell Flowering Phenology
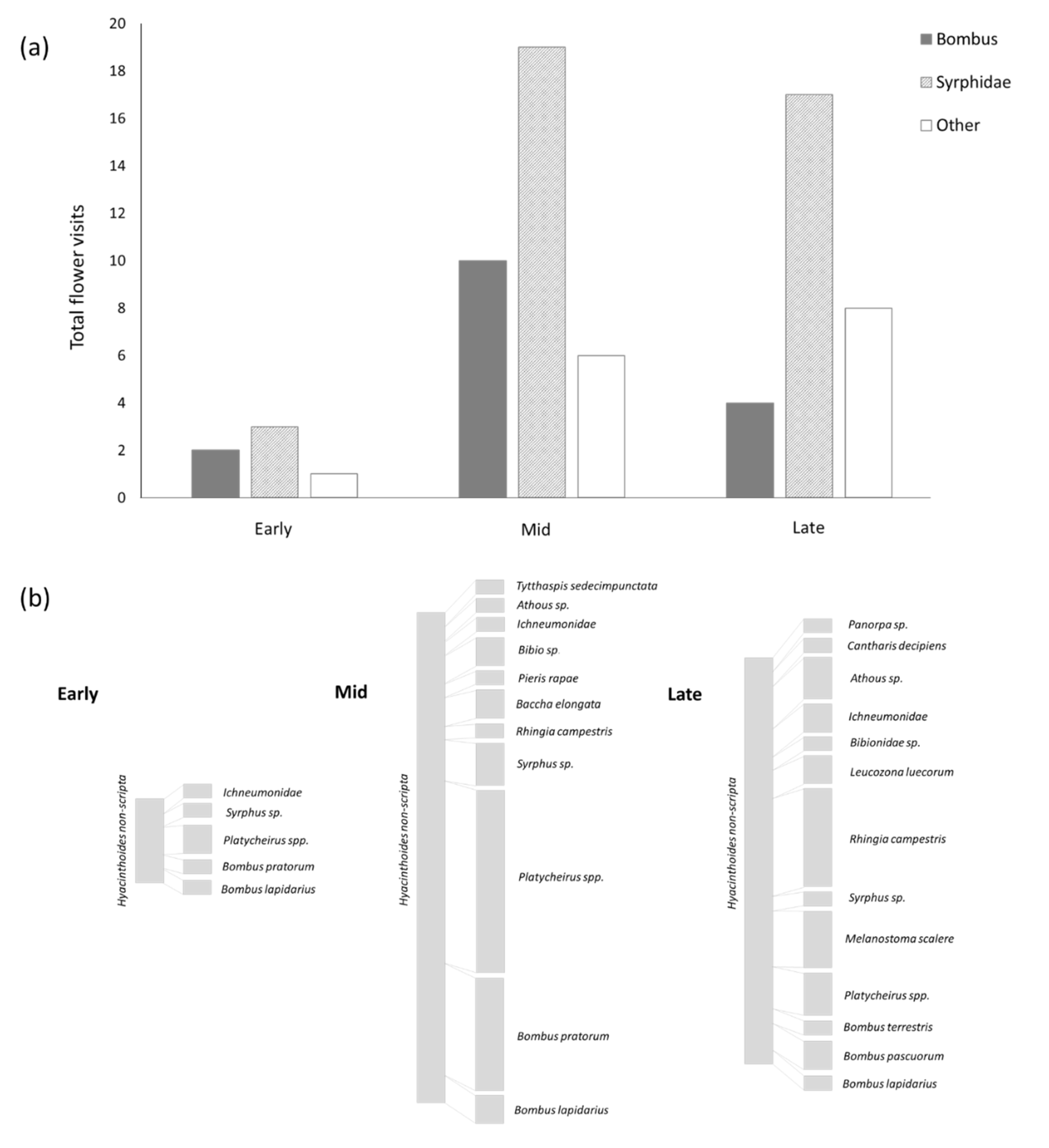
3.2.3. Insect Visitation and Seed Production
4. Discussion
4.1. Systematic Review
4.2. BIFoR FACE Field Experiment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ollerton, J.; Winfree, R.; Tarrant, S. How many flowering plants are pollinated by animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Mortensen, L.O.; Schmidt, N.M.; Høye, T.T.; Damgaard, C.; Forchhammer, M.C. Analysis of trophic interactions reveals highly plastic response to climate change in a tri-trophic High-Arctic ecosystem. Polar Biol. 2016, 39, 1467–1478. [Google Scholar] [CrossRef]
- Hegland, S.J.; Nielsen, A.; Lázaro, A.; Bjerknes, A.L.; Totland, Ø. How does climate warming affect plant-pollinator interac-tions? Ecol. Lett. 2009, 12, 184–195. [Google Scholar] [CrossRef]
- Bale, J.S.; Hayward, S.A.L. Insect overwintering in a changing climate. J. Exp. Biol. 2010, 213, 980–994. [Google Scholar] [CrossRef] [Green Version]
- Owen, E.L.; Bale, J.S.; Hayward, S.A.L. Can winter-active bumblebees survive the cold? Assessing the cold tolerance of Bom-bus terrestris audax and the effects of pollen feeding. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Phillips, B.B.; Shaw, R.F.; Holland, M.J.; Fry, E.L.; Bardgett, R.D.; Bullock, J.M.; Osborne, J.L. Drought reduces floral resources for pollinators. Glob. Chang. Biol. 2018, 24, 3226–3235. [Google Scholar] [CrossRef]
- Memmott, J.; Craze, P.G.; Waser, N.M.; Price, M.V. Global warming and the disruption of plant? pollinator interactions. Ecol. Lett. 2007, 10, 710–717. [Google Scholar] [CrossRef]
- Schenk, M.; Krauss, J.; Holzschuh, A. Desynchronizations in bee–plant interactions cause severe fitness losses in solitary bees. J. Anim. Ecol. 2018, 87, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Powney, G.D.; Carvell, C.; Edwards, M.; Morris, R.K.A.; Roy, H.E.; Woodcock, B.A.; Isaac, N.J.B. Widespread losses of pollinating insects in Britain. Nat. Commun. 2019, 10, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Curtis, P.S.; Wang, X. A meta-analysis of elevated CO 2 effects on woody plant mass, form, and physiology. Oecologia 1998, 113, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Pringle, E.G. Integrating plant carbon dynamics with mutualism ecology. New Phytol. 2016, 210, 71–75. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Springer, C.J.; Ward, J.K. Flowering time and elevated atmospheric CO2. New Phytol. 2007, 176, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Simpson, G.G.; Gendall, A.R.; Dean, C. When to Switch to Flowering. Annu. Rev. Cell Dev. Biol. 1999, 15, 519–550. [Google Scholar] [CrossRef] [PubMed]
- Lesica, P.; Kittelson, P. Precipitation and temperature are associated with advanced flowering phenology in a semi-arid grassland. J. Arid. Environ. 2010, 74, 1013–1017. [Google Scholar] [CrossRef]
- Haslett, J.R. Adult feeding by holometabolous insects: Pollen and nectar as complementary nutrient sources for Rhingia cam-pestris Diptera: Syrphidae. Oecologia 1989, 81, 361–363. [Google Scholar] [CrossRef] [PubMed]
- López-Cubillos, S.; Hughes, L. Effects of elevated carbon dioxide (CO2) on flowering traits of three horticultural plant species. Aust. J. Crop. Sci. 2016, 10, 1523–1528. [Google Scholar] [CrossRef]
- Ziska, L.H.; Pettis, J.S.; Edwards, J.; Hancock, J.E.; Tomecek, M.B.; Clark, A.; Dukes, J.S.; Loladze, I.; Polley, H.W. Rising at-mospheric CO2 is reducing the protein concentration of a floral pollen source essential for North American bees. Proc. R. Soc. B: Biol. Sci. 2016, 283, 20160414. [Google Scholar] [CrossRef] [Green Version]
- Ziska, L.H.; Caulfield, F.A. Rising CO2 and pollen production of common ragweed (Ambrosia artemisiifolia L.), a known allergy-inducing species: Implications for public health. Funct. Plant Biol. 2000, 27, 893. [Google Scholar] [CrossRef]
- LaDeau, S.L.; Clark, J.S. Pollen production by Pinus taeda growing in elevated atmospheric CO2. Funct. Ecol. 2006, 20, 541–547. [Google Scholar] [CrossRef]
- Gardener, M.C.; Gillman, M.P. The taste of nectar–a neglected area of pollination ecology. Oikos 2002, 98, 552–557. [Google Scholar] [CrossRef]
- Pacini, E.; Nepi, M.; Vesprini, J.L. Nectar biodiversity: A short review. Plant Syst. Evol. 2003, 238, 7–21. [Google Scholar] [CrossRef]
- Lake, J.C.; Hughes, L. Nectar Production and Floral Characteristics of Tropaeolum majus L. Grown in Ambient and Elevated Carbon Dioxide. Ann. Bot. 1999, 84, 535–541. [Google Scholar] [CrossRef] [Green Version]
- Bezemer, T.M.; Jones, T.H. Plant-Insect Herbivore Interactions in Elevated Atmospheric CO2: Quantitative Analyses and Guild Effects. Oikos 1998, 82, 212. [Google Scholar] [CrossRef]
- Gifford, R.M.; Barrett, D.J.; Lutze, J.L. The effects of elevated [CO2] on the C:N and C:P mass ratios of plant tissues. Plant Soil 2000, 224, 1–14. [Google Scholar] [CrossRef]
- Vaudo, A.D.; Farrell, L.M.; Patch, H.M.; Grozinger, C.M.; Tooker, J.F. Consistent pollen nutritional intake drives bumble bee (Bombus impatiens) colony growth and reproduction across different habitats. Ecol. Evol. 2018, 8, 5765–5776. [Google Scholar] [CrossRef] [PubMed]
- Dolezal, A.G.; Toth, A.L. Feedbacks between nutrition and disease in honey bee health. Curr. Opin. Insect Sci. 2018, 26, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, M.A.; Burkle, L.A.; Manson, J.S.; Runyon, J.B.; Trowbridge, A.M.; Zientek, J. Global change effects on plant–insect interactions: The role of phytochemistry. Curr. Opin. Insect Sci. 2017, 23, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Norby, R.J.; De Kauwe, M.G.; Domingues, T.F.; Duursma, R.A.; Ellsworth, D.S.; Goll, D.S.; Lapola, D.M.; Luus, K.A.; Mac-Kenzie, A.R.; Medlyn, B.E.; et al. Model–data synthesis for the next generation of forest free-air CO2 enrichment (FACE) experiments. New Phytol. 2016, 209, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drake, J.E.; Macdonald, C.A.; Tjoelker, M.; Crous, K.Y.; Gimeno, T.E.; Singh, B.; Reich, P.B.; Anderson, I.C.; Ellsworth, D.S. Short-term carbon cycling responses of a mature eucalypt woodland to gradual stepwise enrichment of atmospheric CO2 concentration. Glob. Chang. Biol. 2016, 22, 380–390. [Google Scholar] [CrossRef]
- James, K.L.; Randall, N.P.; Haddaway, N.R. A methodology for systematic mapping in environmental sciences. Environ. Évid. 2016, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Hart, K.M.; Curioni, G.; Blaen, P.; Harper, N.J.; Miles, P.; Lewin, K.F.; Nagy, J.; Bannister, E.J.; Cai, X.M.; Thomas, R.M.; et al. Characteristics of free air carbon dioxide enrichment of a northern temperate mature forest. Glob. Chang. Biol. 2019, 26, 1023–1037. [Google Scholar] [CrossRef] [Green Version]
- Corbet, S.A. Fruit and seed production in relation to pollination and resources in bluebell, Hyacinthoides non-scripta. Oecologia 1998, 114, 349–360. [Google Scholar] [CrossRef]
- Faegri, K.; Van Der Pijl, L. Principles of Pollination Ecology, 3rd ed.; Pergamon Press: Oxford, UK, 2013. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3. Available online: https://svn.r-project.org/R-packages/trunk/nlme/ (accessed on 22 February 2020).
- Zuur, A.F.; Ieno, E.N. A protocol for conducting and presenting results of regression-type analyses. Methods Ecol. Evol. 2016, 7, 636–645. [Google Scholar] [CrossRef]
- Culley, T.M.; Weller, S.G.; Sakai, A.K. The evolution of wind pollination in angiosperms. Trends Ecol. Evol. 2002, 17, 361–369. [Google Scholar] [CrossRef]
- He, J.S.; Wolfe-Bellin, K.S.; Bazzaz, F.A. Leaf-level physiology, biomass, and reproduction of Phytolacca americana under con-ditions of elevated CO2 and altered temperature regimes. Int. J. Plant Sci. 2005, 166, 615–622. [Google Scholar] [CrossRef]
- Schemske, D.W.; Willson, M.F.; Melampy, M.N.; Miller, L.J.; Verner, L.; Schemske, K.M.; Best, L.B. Flowering ecology of some spring woodland herbs. Ecology. 1978, 59, 351–366. [Google Scholar] [CrossRef]
- Wang, D.; Heckathorn, S.A.; Wang, X.; Philpott, S.M. A meta-analysis of plant physiological and growth responses to tem-perature and elevated CO2. Oecologia 2012, 169, 1–13. [Google Scholar] [CrossRef]
- Corbet, S.A. Spatiotemporal patterns in the flowering of bluebell, Hyacinthoides non-scripta (Hyacinthaceae). Flora-Morphol. Distrib. Funct. Ecol. Plants 1999, 194, 345–356. [Google Scholar] [CrossRef]
- Corbet, S.A.; Tiley, C.F. Insect visitors to flowers of bluebell (Hyacinthoides non-scripta). Entomolog. Month. Mag. 1999, 135, 133–141. [Google Scholar]
- Richman, S.K.; Levine, J.M.; Stefan, L.; Johnson, C.A. Asynchronous range shifts drive alpine plant–pollinator interactions and reduce plant fitness. Glob. Chang. Biol. 2020, 26, 3052–3064. [Google Scholar] [CrossRef] [PubMed]
- Rafferty, N.E.; Ives, A.R. Effects of experimental shifts in flowering phenology on plant-pollinator interactions. Ecol. Lett. 2010, 14, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Kharouba, H.M.; Ehrlén, J.; Gelman, A.; Bolmgren, K.; Allen, J.M.; Travers, S.E.; Wolkovich, E.M. Global shifts in the pheno-logical synchrony of species interactions over recent decades. Proc. Natl. Acad. Sci. USA 2018, 115, 5211–5216. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Array | Treatment | Patch Size (m2) | Total Number of Racemes | Total Number of Flowers | Total Number of Fruits |
|---|---|---|---|---|---|
| 1 | eCO2 | 0.71 | 60 | 257 | 167 |
| 2 | Ambient | 1.60 | 88 | 581 | 405 |
| 3 | Ambient | 6.03 | 150 | 933 | 613 |
| 4 | eCO2 | 11.31 | 230 | 1495 | 985 |
| 5 | Ambient | 2.90 | 96 | 536 | 285 |
| 6 | eCO2 | 12.57 | 250 | 1879 | 1279 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crowley, L.M.; Sadler, J.P.; Pritchard, J.; Hayward, S.A.L. Elevated CO2 Impacts on Plant–Pollinator Interactions: A Systematic Review and Free Air Carbon Enrichment Field Study. Insects 2021, 12, 512. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12060512
Crowley LM, Sadler JP, Pritchard J, Hayward SAL. Elevated CO2 Impacts on Plant–Pollinator Interactions: A Systematic Review and Free Air Carbon Enrichment Field Study. Insects. 2021; 12(6):512. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12060512
Chicago/Turabian StyleCrowley, Liam M., Jonathan P. Sadler, Jeremy Pritchard, and Scott A. L. Hayward. 2021. "Elevated CO2 Impacts on Plant–Pollinator Interactions: A Systematic Review and Free Air Carbon Enrichment Field Study" Insects 12, no. 6: 512. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12060512