
Contribution of Epigenetic Mechanisms in the Regulation of Environmentally-Induced Polyphenism in Insects


INRAE Bretagne-Normandie-UMR 1349 IGEPP, Domaine de La motte BP 35327, CEDEX, 35653 Le Rheu, France
*
Author to whom correspondence should be addressed.
Insects 2021, 12(7), 649; https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070649
Submission received: 8 June 2021
/
Revised: 8 July 2021
/
Accepted: 12 July 2021
/
Published: 15 July 2021
(This article belongs to the Special Issue Epigenetics in Insects)
Abstract
:Simple Summary
Polyphenism is a widespread phenomenon in insects that allows organisms to produce alternative and discrete phenotypes in response to environmental conditions. Epigenetic mechanisms, including histone post-translational modifications, DNA methylation and non-coding RNAs, are essential mechanisms that can promote rapid and flexible changes in the expression of transcriptional programs associated with the production of alternative phenotypes. This review summarizes knowledge regarding the contribution of those mechanisms in the regulation of the most-studied examples of polyphenism in insects.
Abstract
Many insect species display a remarkable ability to produce discrete phenotypes in response to changes in environmental conditions. Such phenotypic plasticity is referred to as polyphenism. Seasonal, dispersal and caste polyphenisms correspond to the most-studied examples that are environmentally-induced in insects. Cues that induce such dramatic phenotypic changes are very diverse, ranging from seasonal cues, habitat quality changes or differential larval nutrition. Once these signals are perceived, they are transduced by the neuroendocrine system towards their target tissues where gene expression reprogramming underlying phenotypic changes occur. Epigenetic mechanisms are key regulators that allow for genome expression plasticity associated with such developmental switches. These mechanisms include DNA methylation, chromatin remodelling and histone post-transcriptional modifications (PTMs) as well as non-coding RNAs and have been studied to various extents in insect polyphenism. Differential patterns of DNA methylation between phenotypes are usually correlated with changes in gene expression and alternative splicing events, especially in the cases of dispersal and caste polyphenism. Combinatorial patterns of histone PTMs provide phenotype-specific epigenomic landscape associated with the expression of specific transcriptional programs, as revealed during caste determination in honeybees and ants. Alternative phenotypes are also usually associated with specific non-coding RNA profiles. This review will provide a summary of the current knowledge of the epigenetic changes associated with polyphenism in insects and highlights the potential for these mechanisms to be key regulators of developmental transitions triggered by environmental cues.
1. Introduction
Organisms have to cope with the constant fluctuations of their biotic or abiotic environment. When these changes are perceived and integrated, the organism can potentially adjust its phenotype by modifying its physiology accordingly to enhance its ability to face these new environmental conditions. The ability for a genotype to produce different phenotypes in response to fluctuations of external cues is defined as phenotypic plasticity [1]. This plasticity relies on the capacity of the genome to express different combinations of genes and proteins as a response to external signals in order to produce an adjusted phenotype. Phenotypic plasticity can be continuous, with gradual phenotypes being produced depending on the environment, ranging from subtle metabolic adjustments to wider morphological changes at the scale of tissues or even organs [2]. Phenotypic plasticity can also result in the production of discrete phenotypes, often exhibiting extreme morphological, physiological and behavioural differences. Such a discrete phenotypic plasticity is defined as polyphenism [3]. Polyphenisms are widespread in insects [4]. The existence of very discrete developmental stages throughout the life cycle of many insects is the most obvious example. Larval, pupal and adult stages indeed display very different morphological and ecological traits in holometabolous insects (e.g., Coleopteran, Lepidoptera, Hymenoptera, Diptera) thus highlighting the ability of a unique genome to express different gene and protein combinations at various steps of the life cycle. This genome expression plasticity is not triggered by external cues but by internal factors. Remarkably, insects also display other types of polyphenism that are induced by external stimuli. These are referred to as environmentally-induced polyphenism and provide insects with an extraordinary ability to rapidly adapt their phenotype to a wide array of environmental conditions. The signals triggering these environmental changes can be abiotic (e.g., temperature, photoperiod), as is the case for seasonal polyphenism, where insects produce discrete morphs adapted to seasonal constraints (e.g., wing patterning in butterfly or reproductive mode alternation in aphids). However, these signals can also be biotic. For example, dispersal polyphenism relies on the production of alternative morphs that are morphologically and behaviourally adapted to dispersion (e.g., phase polyphenism in locusts and wing polyphenism in aphids). In those cases, an increase in population density and subsequently of tactile stimuli [5] as well as an impoverishment or rarefaction of nutritional resources [6] will trigger the development of alternative phenotypes. In social insects (ants, bees, wasps and termites) where reproductive and non-reproductive tasks are partitioned within the colony (caste polyphenism), the food given to the larva generally determines if it will develop into the worker or queen caste. Although these three types of polyphenisms (i.e., seasonal, dispersal and caste polyphenisms) are induced by different environmental signals, they share common features in their regulatory scheme (Figure 1). External cues first need to be perceived and integrated by specific groups of cells, tissues or organs that are dependent on the nature of the signal itself. This processed signal must then be transduced towards the target tissues/organs that will express alternative genetic programs (summarized in Table 1). In seasonal, dispersal and caste polyphenisms, the neuro-endocrine system is a central integrator and transducer of external cues [7]. The nervous structures, neuropeptides or hormones involved vary between species and polyphenisms, but are well documented [4]. Finally, once the target tissues/organs receive the signal, the genome of the corresponding cells will modify its expression allowing the transcription of new sets of genes and proteins necessary for the production of the alternative phenotype. Such genome-wide expression modifications rely on mechanisms that allow for rapid, flexible changes in transcriptional status of many and distant genes in a coordinated manner, such as epigenetic mechanisms.
Epigenetic mechanisms are major regulators of gene expression in all organisms, including insects [8], and are thus involved in the production of alternative phenotypes. Epigenetics encompasses all the processes of phenotypic variation that are not due to DNA sequence variation. These phenotypic changes might also be heritable across generations. Epigenetic mechanisms involved in the production of alternative phenotypes may occur at different levels. A first level involves the structure of the chromatin. In the nucleus, the DNA containing the coding information is wrapped around histone octamers to form the nucleosomes that correspond to the basic unit of chromatin (Figure 1). Nucleosome distribution and density all along the DNA sequence can affect chromatin accessibility to transcriptional machinery and consequently gene expression levels. Nucleosome stability and distribution is notably controlled by histone post-translational modifications (PTMs). Chromatin remodelling triggered by histone PTMs thus corresponds to a first level of possible epigenetic modifications [9]. DNA methylation is a second major epigenetic mechanism that can affect gene expression levels. DNA methylation involves the transfer of a methyl group onto the C5 position of the cytosine to form 5-methylcytosine (5mC). Changes in DNA methylation levels are associated with gene expression activation or silencing, depending on the organism and the genomic context [10], and have been shown to play key roles in insects (refer to references in [8]). Other changes of the DNA include 5-hydroxymethylcytosine (5hmC), 5-formylcytosine (5fC), 5-carboxylcytosine (5caC) and N 6-methyladenine (6mA), and play critical roles in many biological processes [11,12,13,14,15,16,17,18,19]. A third level for the epigenetic control of alternative phenotypes involves small and long non-coding RNAs (sncRNAs and lncRNAs, respectively). Among sncRNA, miRNAs (for microRNAs) and siRNAs (for small interfering RNAs) are major post-transcriptional gene expression regulators. PiRNAs (for piwi-interacting RNA) and siRNAs can also act as intermediates to direct DNA methylation or histone PTMs at specific genome locations, for example by controlling transposable element expression in germline cells in the case of piRNAs [20]. LncRNAs also share the potential for regulating gene expression at both transcriptional and post-transcriptional levels [21]. Finally, chromatin tridimensional organisation orchestrates gene transcription by regulating chromosome-wide (chromosome territories, A/B compartments) as well as local chromatin topology structures (loops, topologically associating domains) supporting transcription binding factors, such as enhancers or insulators [22,23]. Altogether, the combination of these three major epigenetic mechanisms can have a strong impact on the coordinated regulation of gene expression in the context of alternative phenotype production associated with polyphenism in insects.
In the next sections, we will review the contribution of chromatin regulations, i.e., histone PTMs, DNA methylation and non-coding RNAs in the regulation of insect polyphenisms and their associated transcriptomic programs (summarized in Table 1). Various categories of insect polyphenisms are documented in the literature; however, molecular data detailing the involvement of epigenetic mechanisms can be missing. Moreover, specific phenotypic changes can sometimes be assigned to distinct polyphenisms, for instance, dispersal polyphenism in aphids, which is also a defensive polyphenism when induced by predators [24]. Polyphenism classification can thus be somewhat unclear. In this review, we focus on environmentally-induced polyphenisms in well-studied insect models, classified as three different groups: seasonal, dispersal and caste polyphenisms. For each polyphenism, we start with a brief description of the neuro-endocrine transduction of the triggering signals and follow with a picture of the transcriptional and epigenetic modifications that contribute to the production of alternative phenotypes.
2. Seasonal Polyphenism
Photoperiod and temperature are the most predictable environmental cues organisms can perceive and integrate to anticipate the arrival of a new season. Some species of insects display a seasonal polyphenism, with discrete morphs each adapted to the different seasonal constraints. The two most studied examples so far are wing patterning polyphenism in butterflies as a response to temperature and the reproductive polyphenism in aphids triggered by autumnal photoperiod shortening.
2.1. Wing Patterning Polyphenism in Butterflies
Many butterfly species exhibit a wing patterning polyphenism. The southern African butterfly, Bicyclus anynana, is the most studied example to date. During the wet season, butterflies exhibit prominent marginal eyespots on the surface of their hind wings while in the dry season these eyespots are reduced (Figure 1). In addition, dry season morphs are less active and brownish in colour, probably to better hide from predators in the dry vegetation [25]. The temperature experienced by larvae in their final stage of development is the main external cue governing this polyphenism, with high temperatures (27 °C) leading to the production of the wet season morph, and lower temperatures (20 °C) leading to the production of the dry season morph. Such a phenotypic switch is triggered by the timing of the hormonal peak of circulating ecdysteroids in the haemolymph. At low temperatures, the ecdysteroid peak occurs later during pupal development, while it occurs earlier at higher temperatures [26]. Little is known about the transcriptomic and epigenetic events related to this polyphenism, apart from the involvement of the distal-less gene in the stipulation of a focal region of cells on the wing, allowing the formation of the eyespot [27] and possibly the regulation by ecdysteroid levels [28].
2.2. Reproductive Polyphenism in Aphids
Aphids are hemipterous insects characterized by complex life cycles on diverse and multiple host plants, including many crops. They display major polyphenic traits during their life cycle, including a dispersal polyphenism, allowing them to potentially escape stressful environments (see below) and a seasonal polyphenism resulting in the production of discrete morphs with distinct reproductive modes. Changes in day length, i.e., photoperiod, are the main trigger of this reproductive mode switch [29,30]. Indeed, during spring and summer (under long photoperiod conditions), aphids reproduce exclusively by viviparous parthenogenesis. In their embryos, only diploid oocytes are produced through apomictic parthenogenesis, ending up with genetically identical offspring. Parthenogenesis thus allows aphid populations to colonize rapidly new habitats (notably crops). At the end of the summer, parthenogenetic females perceive the photoperiod shortening and transduce this predictable signal towards their embryos that change their reproductive fate, ending up with the production of sexual morphs (oviparous females and males) that display true haploid gametogenesis. Hence, the shortening of the photoperiod in aphids induces the production of sexual morphs. The first step of this process consists in the photoperiod signal perception, followed by neuro-endocrine transduction. Physiological studies first revealed that a group of neurosecretory cells (called “Neurosecretory Cells Group I”) within the brain were involved in the reception of the decreasing photoperiodic signal, as micro-cauterization of those cells abolishes the photoperiod response [31]. Pharmacological studies then showed that juvenile hormone was playing an essential role in the neuro-endocrine transduction of the signal [32]. Several transcriptomic studies (genome-wide and candidate gene approaches) identified significant gene expression changes between long and short photoperiod conditions in the heads of aphids. Although the causality of these transcriptomic changes remains unknown, these studies highlight the variety of signalling pathways that might be involved in this process. Indeed, it has been suggested that opsin could be involved in photoreception [33,34] and that the circadian clock might be a component of the photoperiodic clock [35,36]. Additional studies showed that neuropeptides, such as insulin [37], as well as neurotransmitters, such as dopamine [38,39,40], could be involved in the neuro-endocrine transduction of the photoperiodic signal. Nevertheless, little is known so far about the epigenetic modifications that trigger these early transcriptomic changes. The annotation of the gene repertoire of DNA methylation [41] and chromatin remodelling [42] systems nevertheless suggests that these are functional in aphids. A genome-wide comparison of the transcriptome of embryos containing haploid germlines (corresponding to parthenogenetic females) and diploid germlines (corresponding to oviparous females) revealed differentially expressed gene coding for histone proteins (H1 and H2B.3), a histone methylase (Suv4–20H1) and uhrf1, a protein known to bind histone residues. This suggests that chromatin remodelling events might be key triggers of embryonic germline fate switches [43]. The contribution of epigenetic mechanisms during the embryo fate switch has not been investigated yet. Nevertheless, two recent studies aimed at comparing the epigenome of adult parthenogenetic and sexual morphs of aphids. Richard et al. (2017) compared the open chromatin and transcriptomic profiles of parthenogenetic females and males in the pea aphid [44]. They found that the chromatin of the single X chromosome of males was more accessible than the chromatin of the two Xs of females, probably to compensate for the missing second X chromosome transcriptional dose in the male genome. This was positively correlated with an enhanced transcription of thousands of genes on the X chromosome involving male-specific functions. Altogether, these data suggest the existence of a chromosome-wide epigenetic regulation of gene expression in males for dosage compensation as well as for the establishment of male-specific transcriptomic programs. Another study recently compared the genome-wide DNA methylation pattern of parthenogenetic females and males in Myzus persicae [45]. They showed that X chromosomes and autosomes had very different methylome profiles between sexes; autosomal genes are hypomethylated in males compared with females, while X-linked genes are hypermethylated in males. Further analyses are now required to understand the contribution of these epigenetic mechanisms at the onset of the transition between parthenogenetic and sexual reproduction as a direct response to photoperiod shortening signal.
3. Dispersal Polyphenism
When environmental conditions are locally favourable, insect individuals invest preferentially in reproduction rather than dispersal. However, following the degradation of habitat quality, the production of phenotypes suitable for dispersion may be favoured. Dispersal polyphenism is such a possibility and has been described in several insect groups [46]. Phase polyphenism in locusts is one of the most studied examples, whereby a switch from a solitary to a gregarious (and migratory) phenotype occurs in response to increasing individual densities. In aphids, wingless parthenogenetic females are able to produce winged parthenogenetic females in their progeny when exposed to various signals indicative of a deterioration of habitat, including the presence of predators and food quality decrease.
3.1. Phase Polyphenism in Locusts
Locusts are responsible for major economic losses when they form migratory swarms that devastate large crop surfaces in a very limited time. Locusts display solitary and gregarious morphs (known as phase polyphenism), which differ in morphology (body size and colour, Figure 1), neurochemistry, physiology and behaviour [47]. When exposed to crowding, individuals with an initial solitary phenotype can initiate a gradual transition towards a gregarious behavioural phenotype in just a few hours, as behavioural changes rely on short-term neuronal plasticity. In contrast, the development of morphological changes associated with the gregarious phenotype extend over several generations [47]. Two distinct sensory pathways are involved in the phase transition in the desert locust (Schistocerca gregaria): the cerebral pathway, involving visual and olfactory stimuli, and the thoracic pathway, induced by tactile information [48]. Once the crowding signals are perceived and integrated, the neuroendocrine system will initiate the phase transition. Serotonin is a key neurotransmitter involved in the control of phase transition. Indeed, its hemolymph concentration dramatically increases within hours after crowding in the desert locust [49]. Such effect is probably mediated through protein kinase A signalling [50]. Dopamine is also suggested in Locusta migratoria to act as a gregarization factor [51] while juvenile hormone and corazonin are likely to be involved in body colour differences between the solitary and gregarious morphs [52], which harbour brown or yellow colour, respectively (Figure 1). Transcriptomics approaches were also conducted to identify genes differentially expressed between solitary and gregarious phenotypes. First, EST datasets from the migratory locust revealed a repression of anabolic, biosynthetic and muscle-specific gene expression in the hindleg, midgut and head tissues of gregarious versus solitary locusts [53]. A strong upregulation of a subset of juvenile hormone binding proteins was also observed in the head of gregarious locusts, thus providing a link between transcriptional changes and hormonal signalling of crowding. A comparison in the desert locust of central nervous system (CNS) transcriptome during the two phases revealed the upregulation of heat-shock proteins (HSP) and immunity genes in gregarious locusts [54], suggesting that population density increase might induce a general physiological stress state. A reduction of energy metabolism and biosynthesis was also observed in gregarious locusts in another transcriptomic study in the migratory locust [55], together with the overexpression of gene coding for synaptic transport components, neuromodulators receptors and neurotransmitter synthetases, confirming the neuronal modulation of phase transition. A microarray study on fourth instar migratory locusts during phase transition then revealed the differential expression of chemosensory (CSP) and takeout proteins, corresponding to large families of soluble binding proteins secreted in the haemolymph [56]. Gregarious and solitary phenotypes are thus associated with profound transcriptomic modifications even if the causality of these changes remains unknown.
As major regulators of rapid changes in gene expression, epigenetic mechanisms are likely to play an important role in phase transition in a tissue-specific manner. So far, DNA methylation is the most studied epigenetic mechanism in the context of phase polyphenism in locusts. It was first demonstrated that the Schistocerca gregaria genome is relatively highly methylated (1.3–1.4%) compared with other insects (less than 1% in drosophila [57]). More precisely, 5mC (i.e., methylated cytosine) are found in both coding regions and repetitive elements [58]. Such trends were confirmed in the Locusta migratoria genome [59]. A methylome analysis of brain tissues from fourth instar solitary and gregarious L. migratoria locusts first revealed that methylation in coding regions was lower and more variable overall than in the rest of the genome. Intergenic repetitive elements were especially highly and stably methylated, and introns were more methylated than exons [60]. 90 genes were then identified as differentially methylated between the two phenotypes, including genes coding for signal transduction proteins and some others involved in microtubule cytoskeleton organization, and thus hypothesized to contribute to synaptic plasticity, since these cellular components are known to play a crucial role in neuronal functions in animals. The functional role of DNA methylation in the control of phase transition is nevertheless not demonstrated yet. Regarding the precise role of chromatin remodelling, little is known on their exact role in phase polyphenism. Histone marks may play a role since H3 histones are more phosphorylated in brains of gregarious than of solitary individuals [47]. Small RNAs transcriptome profiling during the two phases allowed for the identification of 185 miRNAs family candidates, thousands of endogenous siRNAs, some of them of transposon origin, as well as many piRNAs [61]. Interestingly, comparison of small RNA expression patterns of the two phases revealed that longer small RNAs are expressed more abundantly in the solitary individuals, suggesting a specific role in transcript level regulation during this phase. Taken together, the precise role of epigenetic mechanisms in the regulation of phase polyphenism requires further investigation and probably at a tissue-specific level, considering the variety of neuronal and morphological modifications that are at stake during this process.
3.2. Wing Polyphenism in Aphids
During the parthenogenetic part of their life cycle, aphids display a dispersal polyphenism when wingless females produce winged morphs by parthenogenesis in response to various environmental cues, including population density (or crowding), the physiological state of the plants they feed on and the presence of predators or parasites [62]. Winged and wingless parthenogenetic females differ in several morphological, physiological and behavioural characteristics, notably the presence or absence of a functional flight apparatus as well as differences in the level of cuticle sclerotization in the head and thorax [63]. Once crowding signals are perceived, the neuro-endocrine system plays an essential role in its transduction, juvenile hormone being shown to be implicated [6]. A recent transcriptomic comparison of parthenogenetic female heads of the pea aphid submitted or not to a crowding treatment revealed profound differences in gene expression associated with the maternal signalling inducing the winged morphs [64]. Some significant transcriptional changes were found for genes involved in odorant binding, neurotransmitter transport and hormonal signalling. Interestingly, genes from the dopamine and ecdysone pathway were found as differentially expressed, suggesting a role for these molecules in maternal signalling too. The role of ecdysone was then more precisely investigated. Indeed, pharmacological approaches (ecdysone feeding, ecdysone analog and ecdysone receptor antagonist injections) combined with RNAi directed against Ecdysone Receptor (EcR) functionally validated the role of ecdysone signalling in wing polyphenism: an increase in ecdysone signalling is associated with a higher production of wingless individuals, while a decrease in ecdysone signalling promotes the production of winged progeny [65]. Regarding the contribution of epigenetic mechanisms in wing polyphenism, little is known so far apart from a handful of genes involved in chromatin remodelling that are differentially expressed in the heads of crowded and uncrowded parthenogenetic females. This suggests a role for this mechanism in the maternal signalling of wing polyphenism [64]. A more recent study investigated the extent of alternative gene splicing between winged and wingless parthenogenetic females of the pea aphid, at both embryo and adult stages [66]. Overall, they found that 28% and 27% of the expressed genes displayed alternative splicing in winged and wingless adult females, respectively, and 25% for winged-destined and unwinged-destined embryos. When comparing these phenotypes, 332 differentially spliced sites were identified between winged and wingless adults but only 13 between winged and unwinged-destined embryos. The majority of differentially spliced genes were nevertheless not differentially expressed. This study suggests a potential role for alternative splicing in the expression of environmentally induced phenotypes. The causality of these differences is not yet known, but DNA methylation might be a good candidate to explain these splicing patterns (see below). Altogether, epigenetic data are so far very limited in the case of dispersal polyphenism and further investigations are required.
4. Caste Polyphenism
Eusocial insects (i.e., bees, wasps, ants and some termites) display a highly sophisticated organization within the colony, with the presence of different groups of individuals that differ in their morphology and behaviour, which are referred to as caste. The reproductive caste includes queens and males, and the non-reproductive caste includes workers (and soldiers in some species). Caste morphology is generally determined early during larvae development in response to environmental cues (see Lavanchy and Schwander, 2019 [67]). Interestingly, the worker caste is in charge of various tasks, including nursing, foraging, nest maintenance, defence or policing [68]. Within the non-reproductive caste, behaviour can sometimes change in an age-specific manner, such as the nursing to foraging transition in honeybees [69]. In other species (e.g., some ants and termites), non-reproducing individuals can differ in morphology, the largest one being specialized in defence tasks (soldiers or major workers) and the small ones (minor workers) in other tasks. In this section, we will mainly focus on the regulation of caste polyphenism in honeybees and ants. Unlike seasonal and dispersal polyphenisms, the contribution of epigenetic mechanisms in the regulation of caste polyphenism has been well documented in the last few years, especially DNA methylation, chromatin regulation and transcription factors. The contribution of non-coding RNAs is also being considered.
4.1. Nutrient Sensing and Neuro-Endocrine Signalling as Key Triggers of Caste Differentiation
Depending on the environmental stimuli perceived at larval or nymphal stages, individuals can be routed towards alternative developmental programs and produce distinct phenotypes once they are adults. They are especially able to produce either reproductive (queen) or non-reproductive (workers) phenotypes. In ants, the amount and composition (including specific chemical compounds) of larval diet is a widespread stimulus that triggers caste fate determination. In Myrmica rubra, a highly nutritive diet during larval growth, can induce queen development while starvation can prevent it [70,71]. However, other signals, such as temperature or pheromone, can also influence developmental trajectories [72]. Internal factors, such as genetic variations, can also play a role in caste determination; polymorphism in sequences that regulate gene expression (Transcription factors binding sites or ncRNAs) can be used to modulate thresholds that define an individual’s developmental trajectory [73]. More intriguing is the case of social hybridogenesis in some ant species, an unusual form of reproduction found in hybrids between different species. Indeed, queens are produced by parthenogenesis or from intra-lineage mating while workers are only produced from hybrid crosses between genetically divergent lineages [67]. Nevertheless, little is known about the neuro-endocrine transduction of this nutritional signal towards the target tissues apart from a recent study demonstrating that the neuropeptide corazonin was involved in the control of social behaviour as well as caste identity in ants [74].
In honeybees, larvae fed with royal jelly differentiate into queen, while larvae that do not receive this diet develop as workers. Interestingly, it has been shown that royalactin, a 57 kDa protein present in royal jelly, is necessary to induce the differentiation of honeybee larvae into queens [75]. This protein promotes the increase in body size and the development of ovaries. More precisely, royalactin activates the Epidermal growth factor receptor (EGRF), which then turns on the InR substrate (IRS), a well-known target of insulin/insulin-like growth factor (IGF) signalling pathway, but also TOR nutrient-signalling pathway. Juvenile hormone signalling also appears to play an essential role in honeybee caste determination, especially by promoting ovarian development [76]. Diet sensed by larvae are thus transduced by the neuro-endocrine system to promote distinct caste fates later during development. Epigenetic mechanisms then act as key regulators in targeted cell types to establish alternative developmental programs.
4.2. DNA Methylation Patterns Changes during Caste Polyphenism
Eusocial insect genomes, including bees, ants, some wasps and termites, harbour the two key DNA methyltransferases, DNMT1-responsible for maintenance of DNA methylation and DNMT3, involved in de novo DNA methylation [68]. In contrast, at least one or both genes are missing in non-social insects, such as Drosophila, silkworm and red flour beetle. These findings raised a strong interest for the functional role of this epigenetic mechanism in the regulation of caste polyphenism. Kucharski et al. (2008) first showed that RNAi silencing of DNMT3 in honeybee worker larvae resulted in the production of queen-like ovaries, which suggested that DNA methylation might be necessary for inhibiting queen development [77]. Elango et al. further demonstrated that the A. mellifera genome displays a bimodal distribution of CpG content across genes sequences (also found in A. pisum [78]), with functionally distinct low and high CpG methylated genes; the latter displaying a strong bias towards caste-specificity expression [79]. Following these observations, DNA methylation patterns in the brain of adult honeybee workers and queens were compared [80]. Nearly all methylated cytosines were located in gene exons. Genes (#550) were identified as differentially methylated between castes, thus revealing a potential role for DNA methylation in the regulation of gene expression. A strong correlation between methylation patterns and splicing sites was also found, suggesting a role for DNA methylation in alternative splicing. Later on, DNA methylation patterns were compared between larval and adult stages in honeybee workers and queens [81]. This analysis revealed that larval stages displayed five times more differentially methylated genes compared to adult stages, and that a significant proportion of these genes were upregulated in worker larvae compared to queen larvae. Among those genes, several are involved in metabolic and signalling pathways, including juvenile hormone and insulin, two hormones/peptides known to regulate caste determination. Strong correlations between DNA methylation and alternative gene splicing were also found in this study, and particularly for the alk gene, a receptor tyrosine kinase acting as an important regulator of metabolism, thus linking diet experienced by the larvae and caste determination. RNAi knockdown of DNMT3 also induced some changes in gene splicing patterns, thus confirming the role of DNA methylation in the regulation of alternative splicing [82].
In ants, whole-body DNA methylation patterns were examined in two species (Camponotus floridanus and Harpegnathos saltator), confirming that methylated cysotines were strongly enriched in the exons of active genes. As seen in bees, changes in exonic DNA also correlated with alternative splicing, such as exon skipping. Overall, few changes in methylation levels were observed between castes in both ant species. Interestingly, several genes conserved between the two species displayed caste-specific changes in DNA methylation, especially genes involved in reproduction, telomere maintenance and non-coding RNA metabolism [83]. It was also reported that the termite Zootermopsis nevadensis displayed some of the highest levels of DNA methylation found in insects: 12% of the genomic CpGs dinucleotides and 58% of the exonic CpGs [84]. Interestingly, strong differences in methylation between castes were observed. Differentially methylated genes were also more frequently alternatively spliced, and preferentially involved in developmental-related functions. In addition, intergenic differentially methylated regions were enriched for multiple transcription factor binding sites, suggesting an interplay between DNA methylation and transcriptional regulation of gene expression [84]. Altogether, it appears that the global trends related to the role of DNA methylation in the regulation of caste polyphenism are shared between bees, ants and possibly termites.
4.3. Contribution of Transcription Factors and Chromatin Remodelling Events in Caste Polyphenism
In bees, a few indirect cues first suggested a role of chromatin regulation and histone PTMs in the control of caste polyphenism. Mass spectrometry approaches showed that histone H3.1, H3.3 and H4 were extensively modified by lysine methylation and acetylation in honeybees [85]. The authors then compared histone PTMs within queen ovaries and 96-h-old whole larvae. They observed similar global profiles of histone PTMs in both samples as well as some specific combinatorial patterns of lysine methylation on H3K27 and H3K36 residues more frequently identified in histones extracted from queen ovaries than from larvae. Interestingly, the comparison of brain methylomes from queen and worker castes [80] allowed for the identification of the differential methylation of specific histone genes. They found that only intron-containing histone variants were methylated, whereas intronless canonical histone genes were not. This suggests that the interplay between DNA methylation and histone profiles might be important for modulating chromatin accessibility and gene expression in the context of caste polyphenism. Interestingly, it has been shown that royal jelly, known to induce the queen phenotype, has histone deacetylase inhibitor activity (HDACi). Indeed, the fatty acid (E)-10-hydroxy-2-decenoic acid (10HDA), accounts for up to 5% of royal jelly and harbours this HDACi activity, which suggests that the inhibition of histone deacetylation is important for queen caste development [86]. More recently, a genome-wide comparison of various histone PTMs between queen-destined and worker-destined larvae highlighted extensive differences in H3K4me3, H3K27ac as well as H3K36me3 [87]. Many of these were positively correlated with caste-specific gene expression. Intronic H3K27 acetylation was also found to be a trigger of worker caste differentiation. This study thus demonstrates the role of chromatin modifications in the establishment and maintenance of caste-specific transcriptional programs at an early stage of larval development in the honeybee.
In ants, regulatory elements and genome-wide histone PTMs related to caste polyphenism also received strong attention [88]. First, genome sequence comparisons of eight eusocial (one honeybee and seven ant species) and 22 solitary insects (spanning different insect orders) revealed that the transcription factor binding sites (TFBS) within the promoters of orthologous genes are more divergent among eusocial insects than between solitary and eusocial insects [89]. Genes displaying the most important evolutionary changes are involved in neuroendocrine signalling and are differentially expressed between castes in Camponotus floridanus and Harpegnathos saltator species. These genes contain some binding motifs for neuronal-related TFs in their promoters, including cyclic AMP response element binding protein (CREB), empty spiracles (EMS) and grainyhead (GRH), suggesting that neuronal gene networks have been the target of regulatory rewiring in the course of evolution. In Camponotus floridanus, genome-wide comparison of various histone PTMs between two female workers (major and minor) and male phenotypes showed that H3K27ac discriminates between these different castes, but also explains most of the differential gene expression between castes [90]. The genes showing these correlated changes in gene expression and H3K27ac patterns are involved in muscle development and neuronal regulation as well as sensory response. Interestingly, binding sites for CBP, which is the transcriptional coactivator CREB binding protein, show important variations in H3K27ac patterns between castes. Considering the role of CBP as a major acetyltransferase of histone H3 lysine 27 residues in insects [91], this suggests that gene expression changes associated with caste polyphenism establishment might be mediated by differential recruitment of CBP to chromatin.
4.4. Non-Coding RNAs and Caste Polyphenism
The role of small RNAs and long non-coding RNAs in the regulation of gene expression during caste polyphenism has been investigated mainly in ants and honeybees. In particular, miRNAs are well-known post-transcriptional regulators of gene expression. In ants, respectively 96 and 159 miRNA genes were annotated within Camponotus floridanus and Harpegnathos saltator genomes. Interestingly, the minor and major C. floridanus worker castes displayed differential expression of miRNAs; mir-64 being, for example, upregulated in minor worker castes [92]. Later on, small RNA profiling allowed for the identification of 20 novel miRNAs in C. floridanus, 12 of them displaying stage- and caste-specific expression [89]. Small RNA profiling in honeybees unveiled the differential expression of specific miRNA genes in the brain associated with age-dependent behavioural changes [93] as well as behavioural plasticity [94,95]. Small RNAs profiling in bee larval food also revealed that worker jelly displays an increased miRNA complexity and abundance compared with royal jelly. These results indicate a potential important role for miRNAs in the regulation of gene expression in caste polyphenism [96]. Similar patterns of caste-specific expression of miRNAs were also reported in the bumblebee Bombus terrestris [97].
Other classes of small RNAs, including siRNAs and piRNAs have not received much attention yet. Long non-coding RNAs (lncRNAs) are also important regulators of gene expression that can act transcriptionally or post-transcriptionally [98]. For instance, honeybees display two lncRNAs (lncov1 and lncov2) with a potential role in regulating the size of the worker ovaries [99]. Altogether, non-coding RNAs profiling revealed caste-specific expression in various species, but very little is known on their exact functional role in the context of polyphenism.
4.5. Conclusions
Polyphenism appears to be an extraordinarily efficient way for organisms to cope with the various and predictable fluctuations of their environment by producing better-adapted phenotypes. This relies on the genome ability to switch rapidly from a developmental program to another. Insects display a remarkable variety of polyphenisms, ranging from seasonal adaptations, the ability to optimize their dispersal strategies and the division of labour within a colony. In all cases, epigenetic mechanisms are key actors of this genome expression plasticity (summarized in Table 1); however, their precise role in the context of polyphenisms remains to be elucidated, i.e., if they trigger the changes, or simply facilitate or maintain them. Caste polyphenism has thus far received the main attention and especially in well-studied organisms, such as ants and bees, several studies pointing out the role of DNA methylation and alternative splicing in the establishment of caste-specific transcriptional programs underlying alternative phenotypes. The investigation of genome-wide chromatin dynamics also revealed that specific combinations of histone PTMs were strongly associated with the expression of distinct sets of genes, thereby with caste identity. In the case of seasonal and dispersal polyphenism, evidence for the functional role of epigenetic mechanisms in the regulation of alternative phenotypes are so far limited, with the notable exception of the characterization of the role of DNA methylation during the phase polyphenism in locusts. Various transcriptomic studies comparing alternative phenotype production identified many putative candidate genes that could play a role in neuro-endocrine transduction and epigenetic regulation. Nevertheless, their causality is not demonstrated yet, in part because these non-model organisms often suffer from the absence of efficient functional validation tools. DNA methylation analyses is relatively easy to develop; however, the challenge in the near future will rely on setting up molecular techniques (such as ChIP-seq or ATAC-seq for example) to implement genome-wide chromatin dynamics analysis, allowing to reveal the epigenomic identity of alternative phenotypes. In addition, these studies will have to be investigated at the critical steps (and potentially in specific tissues) when (and where) developmental switches occur rather than on final phenotypes. This will then allow for identifying the key factors that are responsive to the environmental signals and initiate the cascades of gene expression changes that finally contribute to the establishment of the different phenotypes.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fordyce, J.A. The evolutionary consequences of ecological interactions mediated through phenotypic plasticity. J. Exp. Biol. 2006, 209, 2377–2383. [Google Scholar] [CrossRef] [Green Version]
- Via, S.; Gomulkiewicz, R.; De Jong, G.; Scheiner, S.; Schlichting, C.D.; Van Tienderen, P.H. Adaptive phenotypic plasticity: Consensus and controversy. Trends Ecol. Evol. 1995, 10, 212–217. [Google Scholar] [CrossRef]
- Mayr, E. Animal Species and Evolution; Harvard University Press: Cambridge, MA, USA; Oxford University Press: London, UK, 1963; 797p. [Google Scholar]
- Simpson, S.J.; Sword, G.; Lo, N. Polyphenism in Insects. Curr. Biol. 2011, 21, R738–R749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roessingh, P.; Bouaïchi, A.; Simpson, S.J. Effects of sensory stimuli on the behavioural phase state of the desert locust, Schistocerca gregaria. J. Insect Physiol. 1998, 44, 883–893. [Google Scholar] [CrossRef] [Green Version]
- Braendle, C.; Davis, G.K.; Brisson, J.; Stern, D. Wing dimorphism in aphids. Heredity 2006, 97, 192–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, G.; Le Trionnaire, G.; Danchin, E.; Sentis, A. Epigenetics and insect polyphenism: Mechanisms and climate change impacts. Curr. Opin. Insect Sci. 2019, 35, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Glastad, K.; Hunt, B.G.; Goodisman, M.A. Epigenetics in Insects: Genome Regulation and the Generation of Phenotypic Diversity. Annu. Rev. Entomol. 2019, 64, 185–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannister, A.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Klose, R.J.; Bird, A.P. Genomic DNA methylation: The mark and its mediators. Trends Biochem. Sci. 2006, 31, 89–97. [Google Scholar] [CrossRef]
- Shi, D.-Q.; Ali, I.; Tang, J.; Yang, W.-C. New Insights into 5hmC DNA modification: Generation, distribution and function. Front. Genet. 2017, 8, 100. [Google Scholar] [CrossRef]
- Wion, D.; Casadesús, J. N6-methyl-adenine: An epigenetic signal for DNA-protein interactions. Nat. Rev. Microbiol. 2006, 4, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Kriaucionis, S.; Heintz, N. The Nuclear DNA Base 5-Hydroxymethylcytosine Is Present in Purkinje Neurons and the Brain. Science 2009, 324, 929–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahiliani, M.; Koh, K.P.; Shen, Y.; Pastor, W.A.; Bandukwala, H.; Brudno, Y.; Agarwal, S.; Iyer, L.M.; Liu, D.R.; Aravind, L.; et al. Conversion of 5-Methylcytosine to 5-Hydroxymethylcytosine in Mammalian DNA by MLL Partner TET1. Science 2009, 324, 930–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.-F.; Li, B.-Z.; Li, Z.; Liu, P.; Wang, Y.; Tang, Q.; Ding, J.; Jia, Y.; Chen, Z.; Li, L.; et al. Tet-Mediated Formation of 5-Carboxylcytosine and Its Excision by TDG in Mammalian DNA. Science 2011, 333, 1303–1307. [Google Scholar] [CrossRef] [Green Version]
- Ito, S.; Shen, L.; Dai, Q.; Wu, S.C.; Collins, L.B.; Swenberg, J.A.; He, C.; Zhang, Y. Tet Proteins Can Convert 5-Methylcytosine to 5-Formylcytosine and 5-Carboxylcytosine. Science 2011, 333, 1300–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffeneder, T.; Hackner, B.; Truß, M.; Münzel, M.; Müller, M.; Deiml, C.A.; Hagemeier, C.; Carell, T. The Discovery of 5-Formylcytosine in Embryonic Stem Cell DNA. Angew. Chem. 2011, 123, 7146–7150. [Google Scholar] [CrossRef]
- Fu, Y.; He, C. Nucleic acid modifications with epigenetic significance. Curr. Opin. Chem. Biol. 2012, 16, 516–524. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhu, Y.; Luo, G.-Z.; Wang, X.; Yue, Y.; Wang, X.; Zong, X.; Chen, K.; Yin, H.; Fu, Y.; et al. Abundant DNA 6mA methylation during early embryogenesis of zebrafish and pig. Nat. Commun. 2016, 7, 13052. [Google Scholar] [CrossRef]
- Kim, V.N.; Han, J.; Siomi, M.C. Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell Biol. 2009, 10, 126–139. [Google Scholar] [CrossRef]
- Engreitz, J.M.; Ollikainen, N.; Guttman, M. Long non-coding RNAs: Spatial amplifiers that control nuclear structure and gene expression. Nat. Rev. Mol. Cell Biol. 2016, 17, 756–770. [Google Scholar] [CrossRef] [Green Version]
- Bonev, B.; Cavalli, G. Organization and function of the 3D genome. Nat. Rev. Genet. 2016, 17, 661–678. [Google Scholar] [CrossRef] [PubMed]
- Lieberman-Aiden, E.; Van Berkum, N.L.; Williams, L.; Imakaev, M.; Ragoczy, T.; Telling, A.; Amit, I.; Lajoie, B.R.; Sabo, P.J.; Dorschner, M.O.; et al. Comprehensive Mapping of Long-Range Interactions Reveals Folding Principles of the Human Genome. Science 2009, 326, 289–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uriel, Y.; Abram, P.K.; Gries, G. Parasitoid pressure does not elicit defensive polyphenism in the green peach aphid. Ecol. Entomol. 2021, 46, 668–676. [Google Scholar] [CrossRef]
- Brakefield, P.M.; Frankino, W.A. Polyphenisms in Lepidoptera: Multidisciplinary approaches to studies of evolution and development. In Phenotypic Plast Insects Mech Consequences; Science Publishers: New York, NY, USA, 2009; pp. 337–368. [Google Scholar]
- Oostra, V.; de Jong, M.; Invergo, B.M.; Kesbeke, F.; Wende, F.; Brakefield, P.M.; Zwaan, B.J. Translating environmental gradients into discontinuous reaction norms via hormone signalling in a polyphenic butterfly. Proc. R. Soc. B Biol. Sci. 2010, 278, 789–797. [Google Scholar] [CrossRef] [Green Version]
- Brakefield, P.M.; Gates, J.; Keys, D.; Kesbeke, F.; Wijngaarden, P.J.; Monteiro, A.; French, V.; Carroll, S.B. Development, plasticity and evolution of butterfly eyespot patterns. Nature 1996, 384, 236–242. [Google Scholar] [CrossRef]
- Brakefield, P.M.; Koch, K. The Regulation of Phenotypic Plasticity of Eyespots in the Butterfly Bicyclus anynana. Am. Nat. 1998, 152, 853. [Google Scholar] [CrossRef]
- Tagu, D.; Sabater-Muñoz, B.; Simon, J.-C. Deciphering reproductive polyphenism in aphids. Invertebr. Reprod. Dev. 2005, 48, 71–80. [Google Scholar] [CrossRef]
- Le Trionnaire, G.; Wucher, V.; Tagu, D. Genome expression control during the photoperiodic response of aphids. Physiol. Entomol. 2013, 38, 117–125. [Google Scholar] [CrossRef]
- Steel, C.G.; Lees, A.D. The role of neurosecretion in the photoperiodic control of polymorphism in the aphid Megoura viciae. J. Exp. Biol. 1977, 67, 117–135. [Google Scholar] [CrossRef]
- Hardie, J.; Lees, A.D. The induction of normal and teratoid viviparae by a juvenile hormone and kinoprene in two species of aphids. Physiol. Entomol. 1985, 10, 65–74. [Google Scholar] [CrossRef]
- Gao, N.; von Schantz, M.; Foster, R.G.; Hardie, J. The putative brain photoperiodic photoreceptors in the vetch aphid, Megoura viciae. J. Insect Physiol. 1999, 45, 1011–1019. [Google Scholar] [CrossRef]
- Collantes-Alegre, J.M.; Mattenberger, F.; Barberà, M.; Martínez-Torres, D. Characterisation, analysis of expression and localisation of the opsin gene repertoire from the perspective of photoperiodism in the aphid Acyrthosiphon pisum. J. Insect Physiol. 2018, 104, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Cortes, T.; Ortiz-Rivas, B.; Martínez-Torres, D.; Cortes, T.; Ortiz-Rivas, B.; Martínez-Torres, D. Identification and characterization of circadian clock genes in the pea aphid Acyrthosiphon pisum. Insect Mol. Biol. 2010, 19, 123–139. [Google Scholar] [CrossRef]
- Barberà, M.; Collantes-Alegre, J.M.; Martínez-Torres, D. Characterisation, analysis of expression and localisation of circadian clock genes from the perspective of photoperiodism in the aphid Acyrthosiphon pisum. Insect Biochem. Mol. Biol. 2017, 83, 54–67. [Google Scholar] [CrossRef] [Green Version]
- Barberà, M.; Cañas-Cañas, R.; Martínez-Torres, D. Insulin-like peptides involved in photoperiodism in the aphid Acyrthosiphon pisum. Insect Biochem. Mol. Biol. 2019, 112, 103185. [Google Scholar] [CrossRef] [PubMed]
- Le Trionnaire, G.; Francis, F.; Jaubert-Possamai, S.; Bonhomme, J.; De Pauw, E.; Gauthier, J.-P.; Haubruge, E.; Legeai, F.; Prunier-Leterme, N.; Simon, J.-C.; et al. Transcriptomic and proteomic analyses of seasonal photoperiodism in the pea aphid. BMC Genom. 2009, 10, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallot, A.; Rispe, C.; Leterme, N.; Gauthier, J.-P.; Jaubert-Possamai, S.; Tagu, D. Cuticular proteins and seasonal photoperiodism in aphids. Insect Biochem. Mol. Biol. 2010, 40, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Trionnaire, G.L.; Hudaverdian, S.; Richard, G.; Tanguy, S.; Gleonnec, F.; Prunier-Leterme, N.; Gauthier, J.-P.; Tagu, D. Dopamine pathway characterization during the reproductive mode switch in the pea aphid. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Walsh, T.K.; Brisson, J.; Robertson, H.M.; Gordon, K.; Jaubert-Possamai, S.; Tagu, D.; Edwards, O.R. A functional DNA methylation system in the pea aphid, Acyrthosiphon pisum. Insect Mol. Biol. 2010, 19, 215–228. [Google Scholar] [CrossRef]
- Rider, S.D., Jr.; Srinivasan, D.G.; Hilgarth, R.S. Chromatin-remodelling proteins of the pea aphid, Acyrthosiphon pisum (Harris). Insect Mol. Biol. 2010, 19, 201–214. [Google Scholar] [CrossRef]
- Gallot, A.; Shigenobu, S.; Hashiyama, T.; Jaubert-Possamai, S.; Tagu, D. Sexual and asexual oogenesis require the expression of unique and shared sets of genes in the insect Acyrthosiphon pisum. BMC Genom. 2012, 13, 76. [Google Scholar] [CrossRef] [Green Version]
- Richard, G.; Legeai, F.; Prunier-Leterme, N.; Bretaudeau, A.; Tagu, D.; Jaquiéry, J.; Le Trionnaire, G. Dosage compensation and sex-specific epigenetic landscape of the X chromosome in the pea aphid. Epigenet. Chromatin 2017, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Mathers, T.C.; Mugford, S.T.; Percival-Alwyn, L.; Chen, Y.; Kaithakottil, G.; Swarbreck, D.; Hogenhout, S.A.; Van Oosterhout, C. Sex-specific changes in the aphid DNA methylation landscape. Mol. Ecol. 2019, 28, 4228–4241. [Google Scholar] [CrossRef] [Green Version]
- Harrison, R.G. Dispersal Polymorphisms in Insects. Annu. Rev. Ecol. Syst. 1980, 11, 95–118. [Google Scholar] [CrossRef]
- Ernst, U.; Van Hiel, B.; Depuydt, G.; Boerjan, B.; De Loof, A.; Schoofs, L. Epigenetics and locust life phase transitions. J. Exp. Biol. 2015, 218, 88–99. [Google Scholar] [CrossRef] [Green Version]
- Burrows, M.; Rogers, S.M.; Ott, S.R. Epigenetic remodelling of brain, body and behaviour during phase change in locusts. Neural Syst. Circuits 2011, 1, 11. [Google Scholar] [CrossRef] [Green Version]
- Rogers, S.M.; Matheson, T.; Sasaki, K.; Kendrick, K.; Simpson, S.J.; Burrows, M. Substantial changes in central nervous system neurotransmitters and neuromodulators accompany phase change in the locust. J. Exp. Biol. 2004, 207 Pt 20, 3603–3617. [Google Scholar] [CrossRef] [Green Version]
- Ott, S.R.; Verlinden, H.; Rogers, S.M.; Brighton, C.H.; Quah, P.S.; Vleugels, R.K.; Verdonck, R.; Broeck, J.V. Critical role for protein kinase A in the acquisition of gregarious behavior in the desert locust. Proc. Natl. Acad. Sci. USA 2012, 109, E381–E387. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Guo, W.; Guo, X.; Wang, X.; Kang, L. Modulation of behavioral phase changes of the migratory locust by the catecholamine metabolic pathway. Proc. Natl. Acad. Sci. USA 2011, 108, 3882–3887. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S. Corazonin and locust phase polyphenism. Appl. Entomol. Zool. 2006, 41, 179–193. [Google Scholar] [CrossRef] [Green Version]
- Kang, L.; Chen, X.; Zhou, Y.; Liu, B.; Zheng, W.; Li, R.; Wang, J.; Yu, J. The analysis of large-scale gene expression correlated to the phase changes of the migratory locust. Proc. Natl. Acad. Sci. USA 2004, 101, 17611–17615. [Google Scholar] [CrossRef] [Green Version]
- Badisco, L.; Ott, S.R.; Rogers, S.M.; Matheson, T.; Knapen, D.; Vergauwen, L.; Verlinden, H.; Marchal, E.; Sheehy, M.R.J.; Burrows, M.; et al. Microarray-based transcriptomic analysis of differences between long-term gregarious and solitarious desert locusts. PLoS ONE 2011, 6, e28110. [Google Scholar]
- Chen, S.; Yang, P.; Jiang, F.; Wei, Y.; Ma, Z.; Kang, L. De Novo Analysis of Transcriptome Dynamics in the Migratory Locust during the Development of Phase Traits. PLoS ONE 2010, 5, e15633. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Wang, X.; Ma, Z.; Xue, L.; Han, J.; Yu, D.; Kang, L. CSP and Takeout Genes Modulate the Switch between Attraction and Repulsion during Behavioral Phase Change in the Migratory Locust. PLoS Genet. 2011, 7, e1001291. [Google Scholar] [CrossRef] [PubMed]
- Takayama, S.; Dhahbi, J.; Roberts, A.; Mao, G.; Heo, S.-J.; Pachter, L.; Martin, D.I.K.; Boffelli, D. Genome methylation in Drosophila melanogaster is found at specific short motifs and is independent of DNMT2 activity. Genome Res. 2014, 24, 821–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falckenhayn, C.; Boerjan, B.; Raddatz, G.; Frohme, M.; Schoofs, L.; Lyko, F. Characterization of genome methylation patterns in the desert locust Schistocerca gregaria. J. Exp. Biol. 2013, 216, 1501–1515. [Google Scholar] [CrossRef] [Green Version]
- Robinson, K.L.; Tohidi-Esfahani, D.; Lo, N.; Simpson, S.J.; Sword, G.A. Evidence for Widespread Genomic Methylation in the Migratory Locust, Locusta migratoria (Orthoptera: Acrididae). PLoS ONE 2011, 6, e28167. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Fang, X.; Yang, P.; Jiang, X.; Jiang, F.; Zhao, D.; Li, B.; Cui, F.; Wei, J.; Ma, C.; et al. The locust genome provides insight into swarm formation and long-distance flight. Nat. Commun. 2014, 5, 2957. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Chen, S.; Yang, P.; Ma, Z.; Kang, L. Characterization and comparative profiling of the small RNA transcriptomes in two phases of locust. Genome Biol. 2009, 10, R6. [Google Scholar] [CrossRef] [Green Version]
- Müller, C.B.; Williams, I.S.; Hardie, J. The role of nutrition, crowding and interspecific interactions in the development of winged aphids. Ecol. Entomol. 2001, 26, 330–340. [Google Scholar] [CrossRef]
- Ishikawa, A.; Miura, T. Transduction of high-density signals across generations in aphid wing polyphenism. Physiol. Èntomol. 2013, 38, 150–156. [Google Scholar] [CrossRef]
- Vellichirammal, N.N.; Madayiputhiya, N.; Brisson, J.A. The genomewide transcriptional response underlying the pea aphid wing polyphenism. Mol. Ecol. 2016, 25, 4146–4160. [Google Scholar] [CrossRef] [Green Version]
- Vellichirammal, N.N.; Gupta, P.; Hall, T.A.; Brisson, J.A. Ecdysone signaling underlies the pea aphid transgenerational wing polyphenism. Proc. Natl. Acad. Sci. USA 2017, 114, 1419–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grantham, M.E.; Brisson, J. Extensive Differential Splicing Underlies Phenotypically Plastic Aphid Morphs. Mol. Biol. Evol. 2018, 35, 1934–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavanchy, G.; Schwander, T. Hybridogenesis. Curr. Biol. 2019, 29, 539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; Simola, D.F.; Bonasio, R.; Liebig, J.; Berger, S.; Reinberg, D. Eusocial insects as emerging models for behavioural epigenetics. Nat. Rev. Genet. 2014, 15, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Nguyen, T.; Mattila, H.R.; Rodriguez-Zas, S.L.; Seeley, T.D.; Robinson, G.E. Molecular Determinants of Scouting Behavior in Honey Bees. Science 2012, 335, 1225–1228. [Google Scholar] [CrossRef]
- Brian, M.V. Studies of caste differentiation in Myrmica rubra L. 4. Controlled larval nutrition. Insectes Sociaux 1956, 3, 369–394. [Google Scholar] [CrossRef]
- Brian, M.V. Caste differentiation and division of labour. In Social Insects; Elsevier Inc.: Amsterdam, The Netherlands, 1979; Volume 1. [Google Scholar]
- Chittka, A.; Wurm, Y.; Chittka, L. Epigenetics: The making of ant castes. Curr. Biol. 2012, 22, R835–R838. [Google Scholar] [CrossRef] [Green Version]
- Beshers, S.N.; Fewell, J.H. Models of division of labor in social insects. Annu. Rev. Entomol. 2001, 46, 413–440. [Google Scholar] [CrossRef] [Green Version]
- Gospocic, J.; Shields, E.J.; Glastad, K.; Lin, Y.; Penick, C.; Yan, H.; Mikheyev, A.; Linksvayer, T.; Garcia, B.A.; Berger, S.; et al. The Neuropeptide Corazonin Controls Social Behavior and Caste Identity in Ants. Cell 2017, 170, 748–759. [Google Scholar] [CrossRef]
- Kamakura, M. Royalactin induces queen differentiation in honeybees. Nature 2011, 473, 478–483. [Google Scholar] [CrossRef]
- Mutti, N.S.; Dolezal, A.G.; Wolschin, F.; Mutti, J.S.; Gill, K.S.; Amdam, G.V. IRS and TOR nutrient-signaling pathways act via juvenile hormone to influence honey bee caste fate. J. Exp. Biol. 2011, 214, 3977–3984. [Google Scholar] [CrossRef] [Green Version]
- Kucharski, R.; Maleszka, R.; Foret, S. Nutritional Control of Reproductive Status in Honeybees via DNA Methylation. Science 2008, 319, 1827–1830. [Google Scholar] [CrossRef] [Green Version]
- Glastad, K.; Hunt, B.G.; Yi, S.V.; Goodisman, M.A.D. DNA methylation in insects: On the brink of the epigenomic era. Insect Mol. Biol. 2011, 20, 553–565. [Google Scholar] [CrossRef]
- Elango, N.; Hunt, B.; Goodisman, M.A.D.; Yi, S.V. DNA methylation is widespread and associated with differential gene expression in castes of the honeybee, Apis mellifera. Proc. Natl. Acad. Sci. USA 2009, 106, 11206–11211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyko, F.; Foret, S.; Kucharski, R.; Wolf, S.; Falckenhayn, C.; Maleszka, R. The honey bee epigenomes: Differential methylation of brain DNA in queens and workers. PLoS Biol. 2010, 8, e1000506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foret, S.; Kucharski, R.; Pellegrini, M.; Feng, S.; Jacobsen, S.E.; Robinson, G.E.; Maleszka, R. DNA methylation dynamics, metabolic fluxes, gene splicing, and alternative phenotypes in honey bees. Proc. Natl. Acad. Sci. USA 2012, 109, 4968–4973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li-Byarlay, H.; Li, Y.; Stroud, H.; Feng, S.; Newman, T.C.; Kaneda, M.; Hou, K.K.; Worley, K.C.; Elsik, C.; Wickline, S.A.; et al. RNA interference knockdown of DNA methyl-transferase 3 affects gene alternative splicing in the honey bee. Proc. Natl. Acad. Sci. USA 2013, 110, 12750–12755. [Google Scholar] [CrossRef] [Green Version]
- Bonasio, R.; Li, Q.; Lian, J.; Mutti, N.S.; Jin, L.; Zhao, H.; Zhang, P.; Wen, P.; Xiang, H.; Ding, Y.; et al. Genome-wide and Caste-Specific DNA Methylomes of the Ants Camponotus floridanus and Harpegnathos saltator. Curr. Biol. 2012, 22, 1755–1764. [Google Scholar] [CrossRef] [Green Version]
- Glastad, K.; Gokhale, K.; Liebig, J.; Goodisman, M.A.D. The caste- and sex-specific DNA methylome of the termite Zootermopsis nevadensis. Sci. Rep. 2016, 6, 37110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickman, M.J.; Kucharski, R.; Maleszka, R.; Hurd, P.J. Extensive histone post-translational modification in honey bees. Insect Biochem. Mol. Biol. 2013, 43, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Spannhoff, A.; Kim, Y.K.; Raynal, N.J.M.; Gharibyan, V.; Su, M.; Zhou, Y.; Li, J.; Castellano, S.; Sbardella, G.; Issa, J.-P.; et al. Histone deacetylase inhibitor activity in royal jelly might facilitate caste switching in bees. EMBO Rep. 2011, 12, 238–243. [Google Scholar] [CrossRef] [Green Version]
- Wojciechowski, M.; Lowe, R.; Maleszka, J.; Conn, D.; Maleszka, R.; Hurd, P.J. Phenotypically distinct female castes in honey bees are defined by alternative chromatin states during larval development. Genome Res. 2018, 28, 1532–1542. [Google Scholar] [CrossRef] [Green Version]
- Bonasio, R. The role of chromatin and epigenetics in the polyphenisms of ant castes. Brief. Funct. Genom. 2014, 13, 235–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simola, D.F.; Wissler, L.; Donahue, G.; Waterhouse, R.; Helmkampf, M.; Roux, J.; Nygaard, S.; Glastad, K.; Hagen, D.E.; Viljakainen, L.; et al. Social insect genomes exhibit dramatic evolution in gene composition and regulation while preserving regulatory features linked to sociality. Genome Res. 2013, 23, 1235–1247. [Google Scholar] [CrossRef] [Green Version]
- Simola, D.F.; Ye, C.; Mutti, N.S.; Dolezal, K.; Bonasio, R.; Liebig, J.; Reinberg, D.; Berger, S.L. A chromatin link to caste identity in the carpenter ant Camponotus floridanus. Genome Res. 2013, 23, 486–496. [Google Scholar] [CrossRef] [Green Version]
- Gräff, J.; Tsai, L.-H. Histone acetylation: Molecular mnemonics on the chromatin. Nat. Rev. Neurosci. 2013, 14, 97–111. [Google Scholar] [CrossRef]
- Bonasio, R.; Zhang, G.; Ye, C.; Mutti, N.S.; Fang, X.; Qin, N.; Donahue, G.; Yang, P.; Li, Q.; Li, C.; et al. Genomic Comparison of the Ants Camponotus floridanus and Harpegnathos saltator. Science 2010, 329, 1068–1071. [Google Scholar] [CrossRef] [Green Version]
- Behura, S.K.; Whitfield, C.W. Correlated expression patterns of microRNA genes with age-dependent behavioural changes in honeybee. Insect Mol. Biol. 2010, 19, 431–439. [Google Scholar] [CrossRef]
- Greenberg, J.K.; Xia, J.; Zhou, X.; Thatcher, S.R.; Gu, X.; Ament, S.A.; Newman, T.C.; Green, P.J.; Zhang, W.; Robinson, G.E.; et al. Behavioral plasticity in honey bees is associated with differences in brain microRNA transcriptome. Genes Brain Behav. 2012, 11, 660–670. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Peng, W.; Li, Z.; Li, W.; Li, L.; Pan, J.; Zhang, S.; Miao, Y.; Chen, S.; Su, S. Next-generation small RNA sequencing for microRNAs profiling in Apis mellifera: Comparison between nurses and foragers. Insect Mol. Biol. 2012, 21, 297–303. [Google Scholar] [CrossRef]
- Guo, X.; Su, S.; Skogerboe, G.; Dai, S.; Li, W.; Li, Z.; Liu, F.; Ni, R.; Guo, Y.; Chen, S.; et al. Recipe for a Busy Bee: MicroRNAs in Honey Bee Caste Determination. PLoS ONE 2013, 8, e81661. [Google Scholar] [CrossRef] [PubMed]
- Collins, D.H.; Mohorianu, I.; Beckers, M.; Moulton, V.; Dalmay, T.; Bourke, A.F.G. MicroRNAs associated with caste determination and differentiation in a primitively eusocial insect. Sci. Rep. 2017, 7, 45674. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Humann, F.C.; Tiberio, G.J.; Hartfelder, K. Sequence and Expression Characteristics of Long Noncoding RNAs in Honey Bee Caste Development—Potential Novel Regulators for Transgressive Ovary Size. PLoS ONE 2013, 8, e78915. [Google Scholar] [CrossRef] [Green Version]
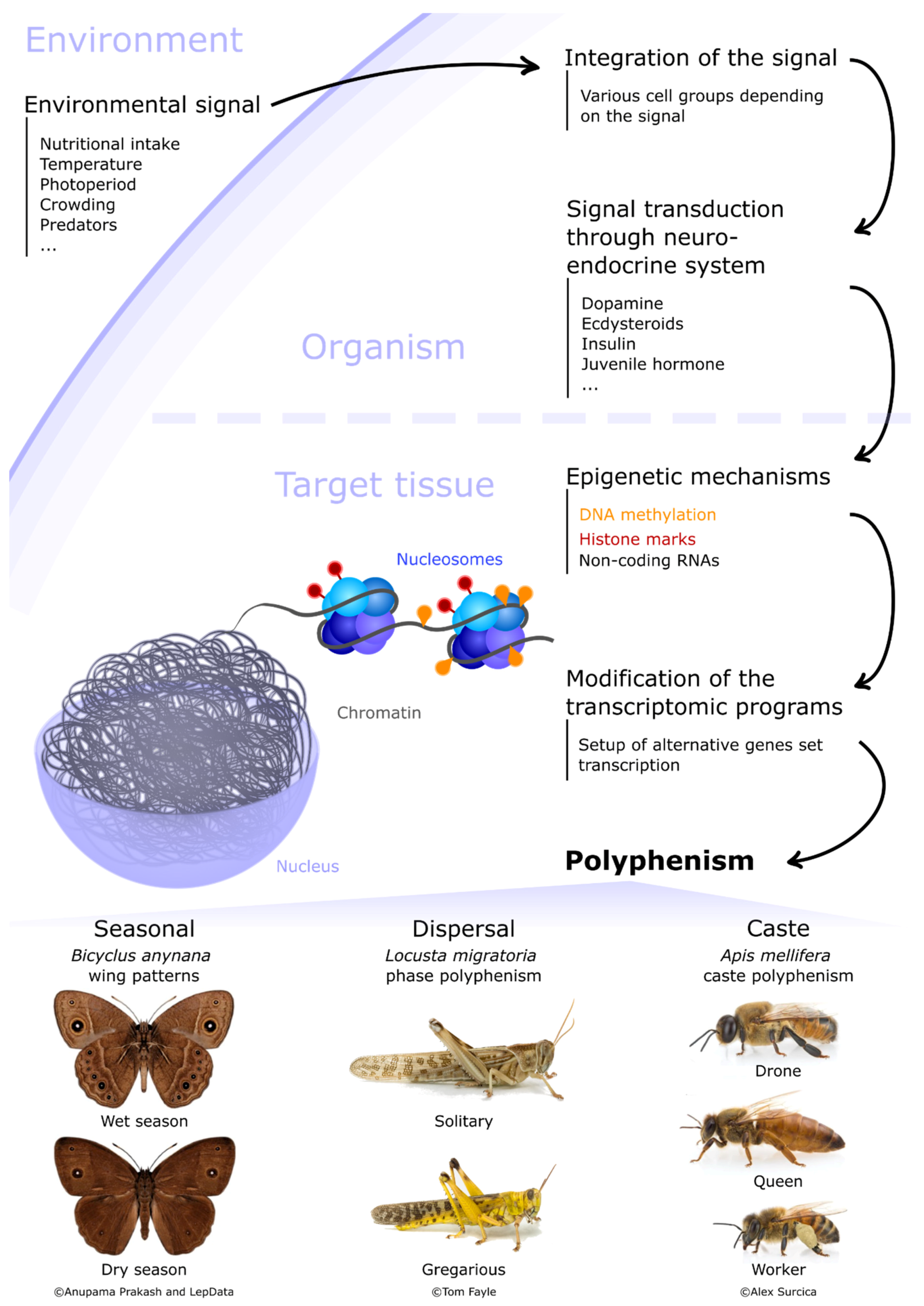
Figure 1.
Insect polyphenism regulation framework. Environmental signal(s) are first perceived and integrated by specific groups of cells. This signal is then transduced to the target tissues/cells by the neuro-endocrine system, often relying on multiple hormones and neuropeptides. Epigenetic mechanisms mainly involving chromatin regulation and DNA methylation are then activated in specific cells of the target tissue(s). These mechanisms then modulate gene expression, ending up with the setup of alternative transcriptional programs underlying the production of discrete phenotypes. Examples of discrete phenotypes for three types of polyphenism are illustrated here: (i) the seasonal polyphenism of Bicyclus anynana with wet (top) and dry (bottom) season morphs (credits: Anupama Prakash, University of Singapore and LepData, 2018); (ii) the phase polyphenism of Locusta migratoria, with a solitary (top) and a gregarious (bottom) male (credits: Tom Fayle, University of Cambridge); (iii) the caste polyphenism of Apis mellifera with drone (top), queen (middle) and worker (bottom) morphs (credits: Alex Surcica, Digital Museum of Natural History).
Figure 1.
Insect polyphenism regulation framework. Environmental signal(s) are first perceived and integrated by specific groups of cells. This signal is then transduced to the target tissues/cells by the neuro-endocrine system, often relying on multiple hormones and neuropeptides. Epigenetic mechanisms mainly involving chromatin regulation and DNA methylation are then activated in specific cells of the target tissue(s). These mechanisms then modulate gene expression, ending up with the setup of alternative transcriptional programs underlying the production of discrete phenotypes. Examples of discrete phenotypes for three types of polyphenism are illustrated here: (i) the seasonal polyphenism of Bicyclus anynana with wet (top) and dry (bottom) season morphs (credits: Anupama Prakash, University of Singapore and LepData, 2018); (ii) the phase polyphenism of Locusta migratoria, with a solitary (top) and a gregarious (bottom) male (credits: Tom Fayle, University of Cambridge); (iii) the caste polyphenism of Apis mellifera with drone (top), queen (middle) and worker (bottom) morphs (credits: Alex Surcica, Digital Museum of Natural History).


{kind=link}
Table 1.
Summary of the epigenetic mechanisms involved in the regulation of environmentally-induced polyphenism in insects.
Table 1.
Summary of the epigenetic mechanisms involved in the regulation of environmentally-induced polyphenism in insects.
| Insect Species | Phenotypes | Environmental Signal(s) | Neuro-Endocrine System | Epigenetic Mechanism Studied | Key References | ||||
|---|---|---|---|---|---|---|---|---|---|
| Chromatin Regulation | DNA Methylation | sRNAs | lncRNAs | ||||||
| Seasonal polyphenism | Bicyclus anynana (Nymphalida) | Wings spots patterns | Temperature | Ecdysteroids | - | - | - | - | - |
| Acyrthosiphon pisum Myzus persicae (Aphidida) | Clonal Sexual | Photoperiod | Dopamine, Insulin, Juvenile hormone | Morph-specific open chromatin profile of the X chromosome | Morph-specific DNA methylation profile of the X chromosome | - | - | [44,45] | |
| Dispersal polyphenism | Schistocerca gregaria Locusta migratoria (Acrididae) | Solitary Gregarious | Visual, olfactory and physical contacts | Corazonin, Dopamine, Juvenile hormone, Serotonin | H3 phosphorylation in gregarious locusts brains | Differentially methylated genes between phases | Phase-specific sRNAs profiles | - | [47,58,59,60,61] |
| Acyrthosiphon pisum (Aphididae) | Winged Wingless | Crowding, Predators, Food quality | Ecdysone, Juvenile hormone | - | - | - | - | [65] | |
| Caste polyphenism | Apis mellifera (Apidae) | Queen Worker | Larval diet, Genetic factors | Insulin, Juvenile hormone | HDACi in royal jelly Caste-specific patterns of H3K4me3, H3K27ac, H3K36me3 | Differentially methylated genes between castes. Correlation between methylation and alternative splicing | Caste-specific miRNAs profiles | Caste specific lncRNAs (lncov1, lncov2) | [75,79,80,81,82,85,86,87,93,94,95,96,99] |
| Camponotus floridanus Harpegnathos saltator (Formicidae) | Queen Male Worker (Gamergate) | Larval diet Pheromones Genetic factors | Corazonin | H3K27ac strongly associated with caste identity Differential binding of CBP | Differentially methylated genes between castes. Correlation between methylation and alternative splicing | Caste-specific miRNAs profiles | - | [83,89,90,92] | |
| Zootermopsis nevadensis (Archotermopsidae) | Queen Worker | Larval diet | Juvenile hormone | - | Differentially methylated genes between castes. Correlation between methylation and alternative splicing | - | - | [84] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Richard, G.; Jaquiéry, J.; Le Trionnaire, G. Contribution of Epigenetic Mechanisms in the Regulation of Environmentally-Induced Polyphenism in Insects. Insects 2021, 12, 649. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070649
AMA Style
Richard G, Jaquiéry J, Le Trionnaire G. Contribution of Epigenetic Mechanisms in the Regulation of Environmentally-Induced Polyphenism in Insects. Insects. 2021; 12(7):649. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070649
Chicago/Turabian StyleRichard, Gautier, Julie Jaquiéry, and Gaël Le Trionnaire. 2021. "Contribution of Epigenetic Mechanisms in the Regulation of Environmentally-Induced Polyphenism in Insects" Insects 12, no. 7: 649. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070649
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.