
Mating-Induced Trade-Offs upon Egg Production versus Fertilization and Offspring’s Survival in a Sawfly with Facultative Parthenogenesis

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Library Preparation and Sequencing
2.3. Quality Control and Assembly
2.4. Differential Expression Analysis
2.5. Functional Annotation and Enrichment Analysis of Differentially Expressed Genes (DEGs)
2.6. Validation by qRT-PCR
3. Results
3.1. Sequencing and Assembly
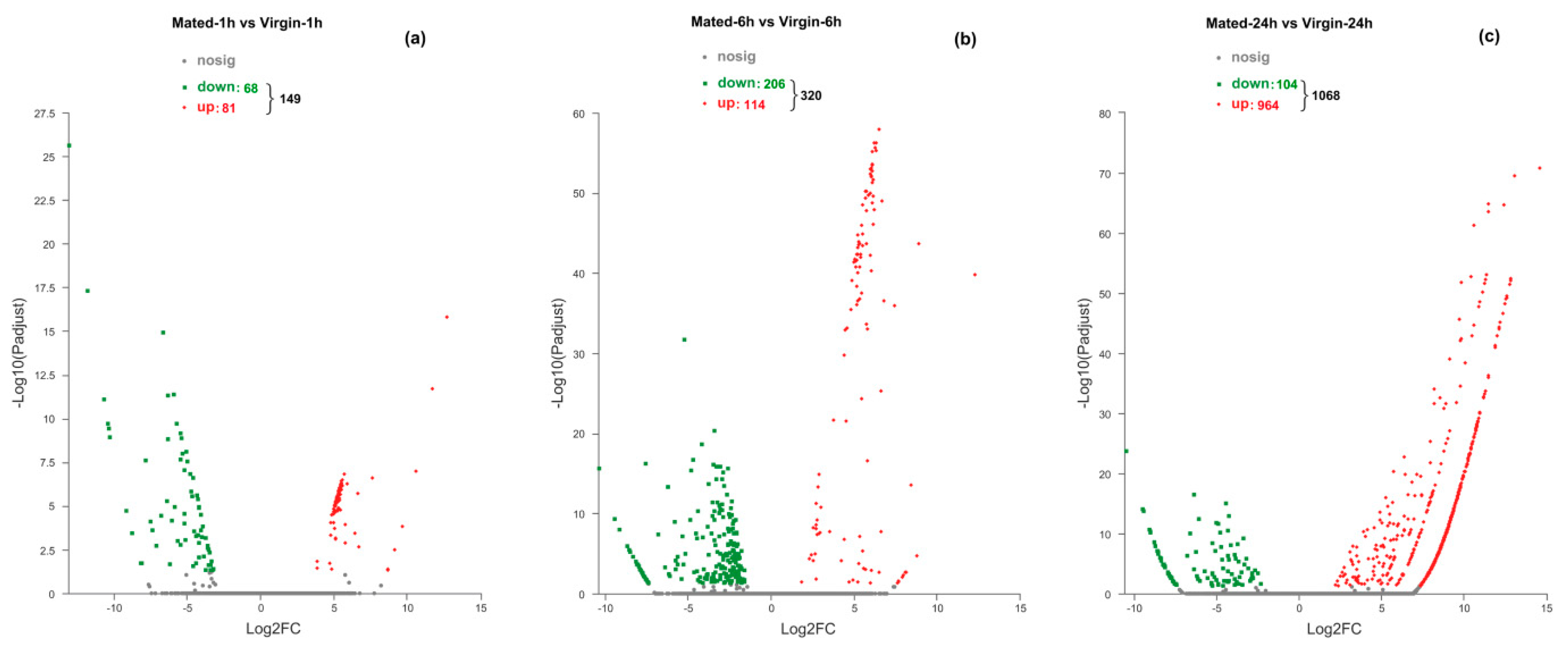
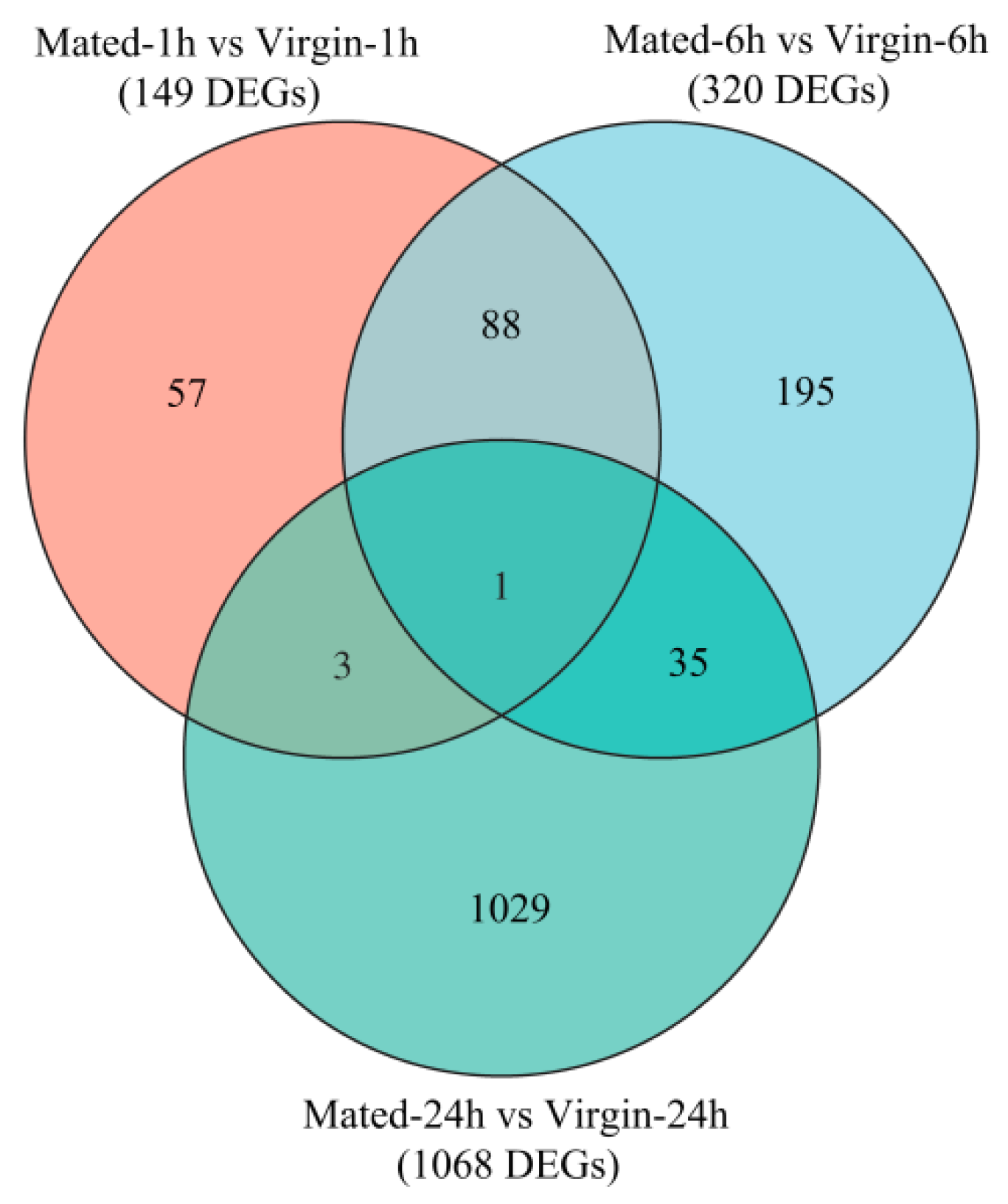
3.2. Outline of Mating-Induced Transcriptional Changes
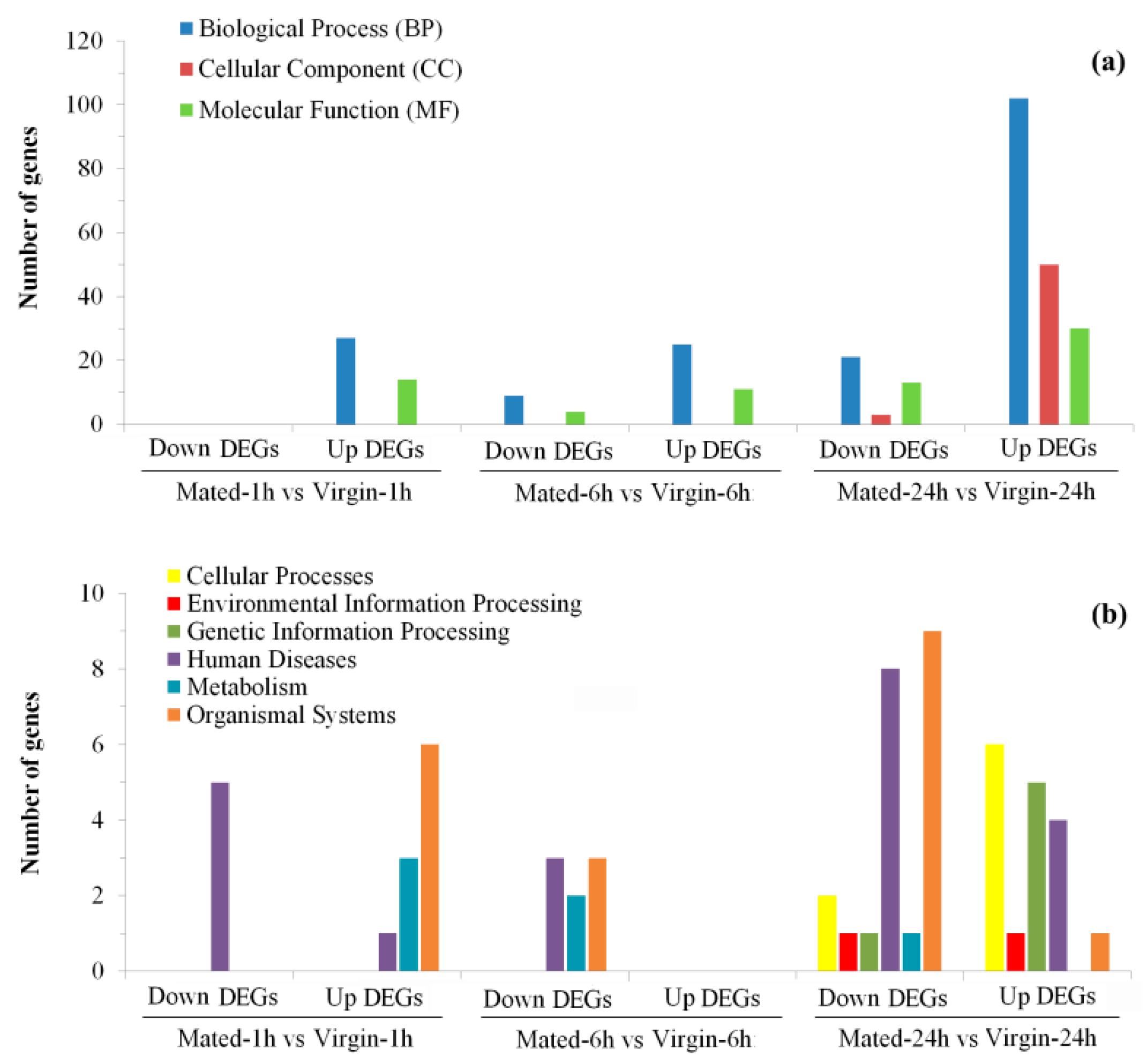
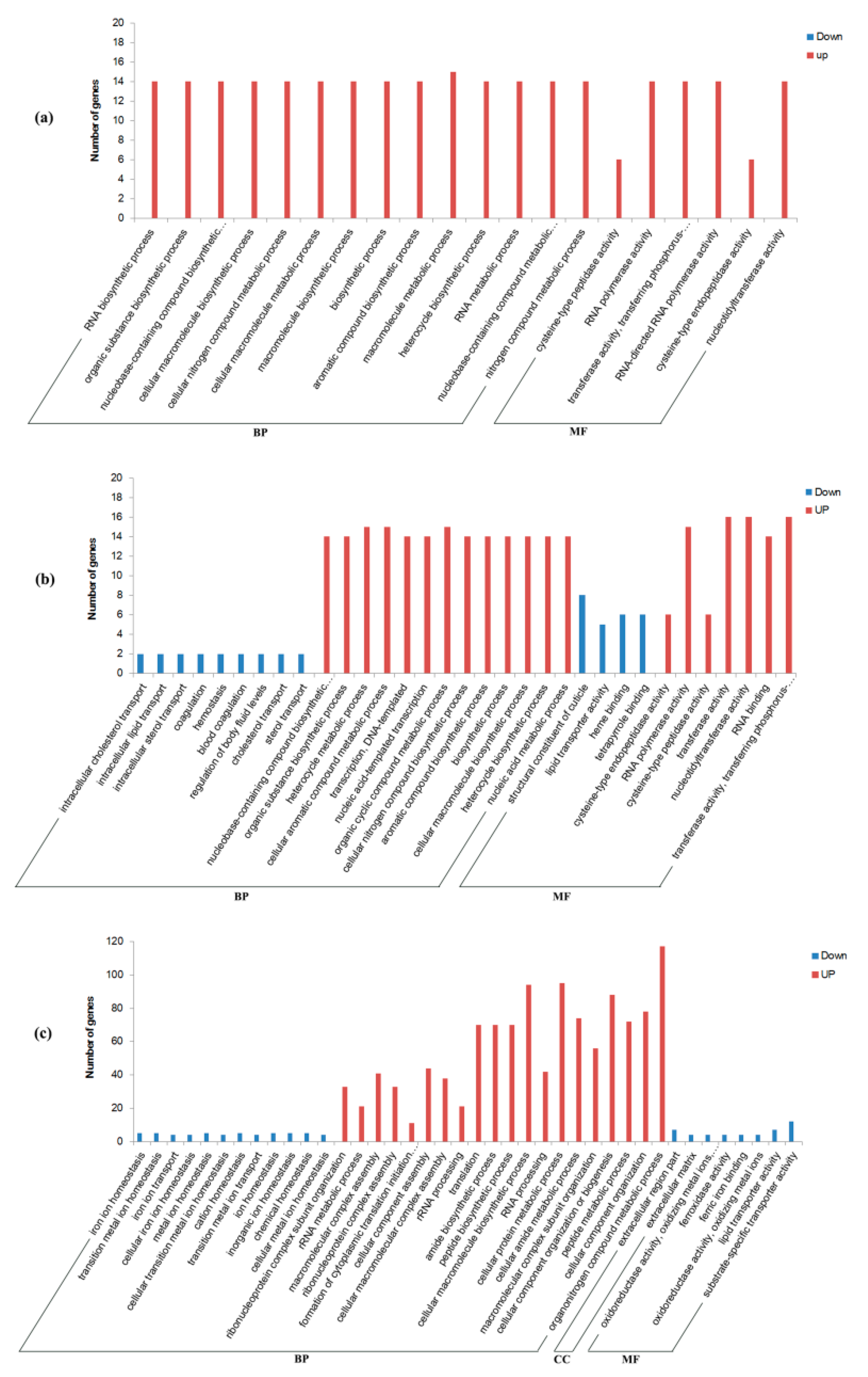
3.3. Transcriptional Changes at 1 h Post-Mating
3.4. Transcriptional Changes at 6 h Post-Mating
3.5. Transcriptional Changes at 24 h Post-Mating
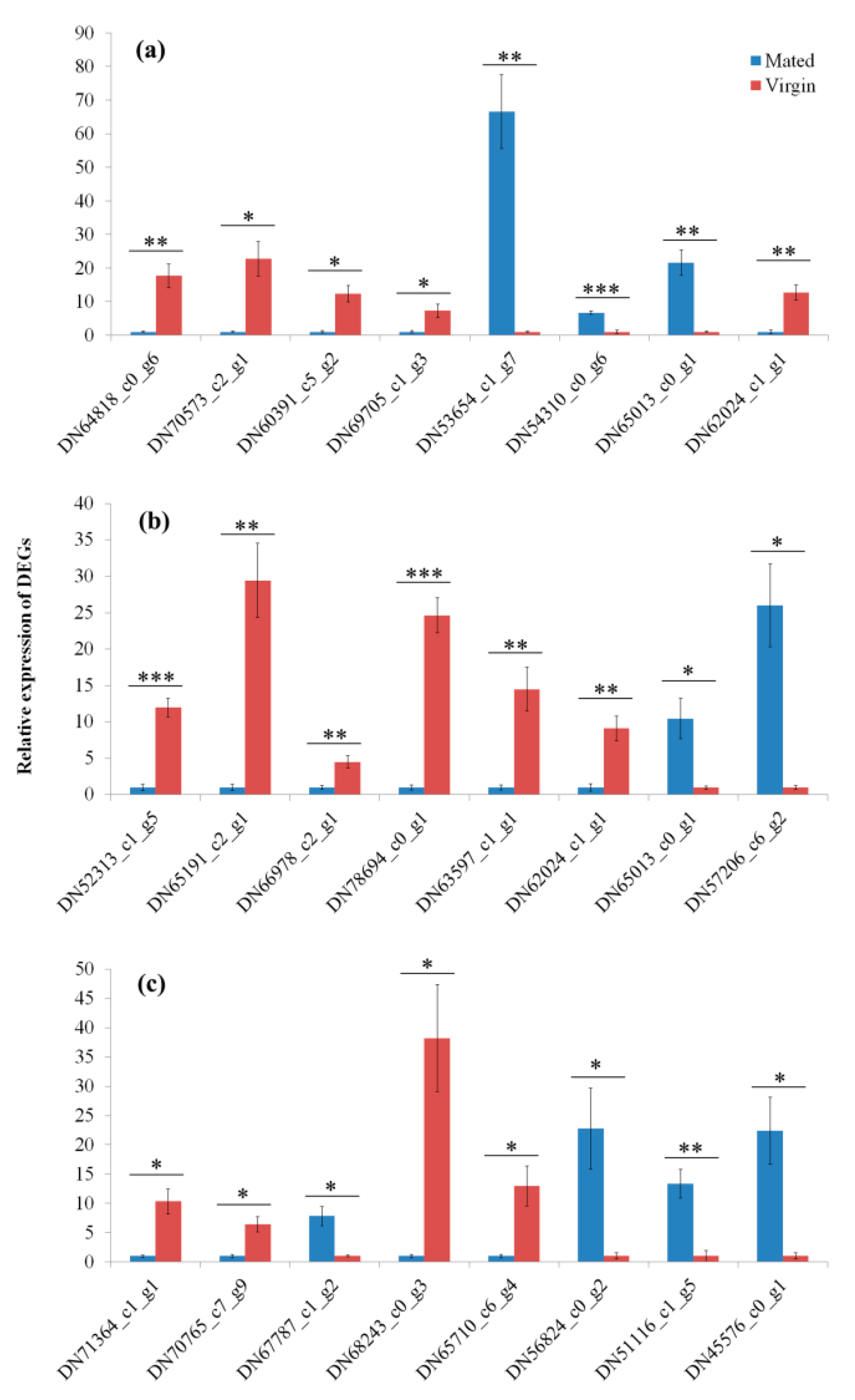
3.6. Validation of RNAseq by qRT-PCR
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schwenke, R.A.; Lazzaro, B.P.; Wolfner, M.F. Reproduction-immunity trade-offs in insects. Annu. Rev. Entomol. 2016, 61, 239–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, M.A.; Flatt, T. Endocrine uncoupling of the trade-off between reproduction and somatic maintenance in eusocial insects. Curr. Opin. Insect Sci. 2016, 16, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, S.R.; Huntsman, E.M.; Flores, M.E.; Snow, J.W. Reproductive potential does not cause loss of heat shock response performance in honey bees. Sci. Rep. 2020, 10, 19610. [Google Scholar] [CrossRef]
- Labbadia, J.; Morimoto, R.I. Repression of the heat shock response is a programmed event at the onset of reproduction. Mol. Cell 2015, 59, 639–650. [Google Scholar] [CrossRef] [Green Version]
- Oku, K.; Price, T.A.R.; Wedell, N. Does mating negatively affect female immune defences in insects? Anim. Biol. 2019, 69, 117–136. [Google Scholar] [CrossRef] [Green Version]
- Stearns, S.C. The Evolution of Life Histories; Oxford University Press: Oxford, UK, 1992. [Google Scholar]
- Flatt, T. Survival costs of reproduction in Drosophila. Exp. Gerontol. 2011, 46, 369–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flatt, T.; Heyland, A.; Rus, F.; Porpiglia, E.; Sherlock, C.; Yamamoto, R.; Garbuzov, A.; Palli, S.R.; Tatar, M.; Silverman, N. Hormonal regulation of the humoral innate immune response in Drosophila melanogaster. J. Exp. Biol. 2008, 211, 2712–2724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Palli, S.R. Identification of a cis-regulatory element required for 20-hydroxyecdysone enhancement of antimicrobial peptide gene expression in Drosophila melanogaster. Insect Mol. Biol. 2009, 18, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Baker, H.G.; Baker, I. Amino-acids in nectar and their evolutionary significance. Nature 1973, 241, 543–545. [Google Scholar] [CrossRef]
- Surendra, K.C.; Olivier, R.; Tomberlin, J.K.; Jha, R.; Khanal, S.K. Bioconversion of organic wastes into biodiesel and animal feed via insect farming. Renew. Energy 2016, 98, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Soller, M.; Bownes, M.; Kubli, E. Mating and sex peptide stimulate the accumulation of yolk in oocytes of Drosophila melanogaster. Eur. J. Biochem. 1997, 243, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.F.; Li, C.; Xu, J.; Liu, J.H.; Ye, H. Male accessory gland secretions modulate female post-mating behavior in the moth Spodoptera litura. J. Insect. Behav. 2014, 27, 105–116. [Google Scholar] [CrossRef]
- Yapici, N.; Kim, Y.J.; Ribeiro, C.; Dickson, B.J. A receptor that mediates the post-mating switch in Drosophila reproductive behaviour. Nature 2008, 451, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Song, X.Q.; Yu, H.; Fu, D.Y.; Xu, J.; Ye, H. Mating-induced differential expression in genes related to reproduction and immunity in Spodoptera litura (Lepidoptera: Noctuidae) female moths. J. Insect Sci. 2020, 20, 10. [Google Scholar] [CrossRef]
- Wigby, S.; Suarez, S.S.; Lazzaro, B.P.; Pizzari, T.; Wolfner, M.F. Sperm success and immunity. Curr. Top. Dev. Biol. 2019, 135, 287–313. [Google Scholar]
- Okada, K.; Suzaki, Y.; Sasaki, R.; Katsuki, M. Fitness costs of polyandry to female cigarette beetle Lasioderma serricorne. Behav. Ecol. Sociobiol. 2017, 71, 86. [Google Scholar] [CrossRef]
- Zhou, S.; Mackay, T.F.C.; Anholt, R.R.H. Transcriptional and epigenetic responses to mating and aging in Drosophila melanogaster. BMC Genom. 2014, 15, 927. [Google Scholar] [CrossRef] [Green Version]
- Dalton, J.E.; Kacheria, T.S.; Knott, S.R.V.; Lebo, M.S.; Nishitani, A.; Sanders, L.E.; Stirling, E.J.; Winbush, A.; Arbeitman, M.N. Dynamic, mating-induced gene expression changes in female head and brain tissues of Drosophila melanogaster. BMC Genom. 2010, 11, 541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, Z.; Liu, Y.; Yang, J.; Xie, W.; Wang, S.; Wu, Q.; Zhou, X.; Pang, B.; Zhang, Y. Transcriptomic analysis of mating responses in Bemisia tabaci MED females. Insects 2020, 11, 308. [Google Scholar] [CrossRef]
- Li, Y.-H.; Xu, J. Geographical Distribution, Biological Characteristics and Diapause Mechanism of Cephalica Chuxiongica; China Forestry Publishing House: Beijing, China, 2020; p. 226. [Google Scholar]
- Fan, J.-X. Study on Biological Characteristics and Control of Cephalcia Chuxiongica Xiao; Southwest Forestry University: Kunming, China, 2017. [Google Scholar]
- Yu, H.; Du, C.-M.; Shi, M.-R.; Feng, L.; Fu, D.-Y.; Xu, J.; Li, Y.-H. The diversity and function of intestinal microorganisms in four geographic Cephalcia chuxiongica (a pine defoliator) populations. J. Appl. Entomol. 2021, 145, 394–405. [Google Scholar] [CrossRef]
- Taeger, A.; Blank, S.M.; Liston, A.D. World catalog of Symphyta (Hymenoptera). Zootaxa 2010, 2580, 3–1064. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N. A Systematic Study of Cephalcia (Hymenoptera: Pamphiliidae) from China; Central South University of Forestry and Technology: Changsha, China, 2019. [Google Scholar]
- Baine, Q.; Looney, C.; Smith, D.R.; Schiff, N.M.; Goulet, H.; Redford, A.J. About Sawflies. Available online: https://idtools.org/id/sawfly/sawflies_biology.php (accessed on 5 March 2021).
- Yan, Z.-L.; Ma, H.-F.; Mao, Y.-L.; Liu, L. Gravid females of Cephalcia chuxiongica (Hymenoptera, Pamphiliidae) are attracted to egg-carrying needles of Pinus yunnanensis. J. Hymenopt. Res. 2018, 65, 157–166. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Pertea, G.; Huang, X.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S.; Lee, Y.; White, J.; Cheung, F.; Parvizi, B.; et al. TIGR gene indices clustering tools (TGICL): A software system for fast clustering of large EST datasets. Bioinformatics 2003, 19, 651–652. [Google Scholar] [CrossRef] [Green Version]
- Storey, J.D. The positive false discovery rate: A Bayesian interpretation and the q-value. Ann. Stat. 2003, 31, 2013–2035. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Burke, T.; Waring, G.L.; Popodi, E.; Minoo, P. Characterization and sequence of follicle cell genes selectively expressed during vitelline membrane formation in Drosophila. Dev. Biol. 1987, 124, 441–450. [Google Scholar] [CrossRef]
- Feyereisen, R. Insect CYP genes and P450 enzymes. In Insect Molecular Biology and Biochemistry; Gilbert, L.I., Ed.; Academic Press: San Diego, CA, USA, 2012; pp. 236–316. [Google Scholar]
- Upadhyay, S.K.; Singh, H.; Dixit, S.; Mendu, V.; Verma, P.C. Molecular characterization of vitellogenin and vitellogenin receptor of Bemisia tabaci. PLoS ONE 2016, 11, e0155306. [Google Scholar]
- Van Rompay, L.; Borghgraef, C.; Beets, I.; Caers, J.; Temmerman, L. New genetic regulators question relevance of abundant yolk protein production in C. elegans. Sci. Rep. 2015, 5, 16381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audsley, N.; Matthews, H.J.; Price, N.R.; Weaver, R.J. Allatoregulatory peptides in Lepidoptera, structures, distribution and functions. J. Insect Physiol. 2008, 54, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Pietrzyk, A.J.; Jaskolski, M.; Bujacz, G. Structural studies of juvenile hormone binding proteins. In Juvenile Hormones and Juvenoids: Modeling Biological Effects and Environmental Fate; Devillers, J., Ed.; CRC Press-Taylor & Francis Group: Boca Raton, FL, USA, 2013; pp. 291–309. [Google Scholar]
- Saurabh, S.; Vanaphan, N.; Wen, W.; Dauwalder, B. High functional conservation of takeout family members in a courtship model system. PLoS ONE 2018, 13, e0204615. [Google Scholar] [CrossRef]
- Ge, L.Q.; Zheng, S.; Gu, H.T.; Zhou, Y.K.; Zhou, Z.; Song, Q.S.; Stanley, D. Jinggangmycin-induced UDP-glycosyltransferase 1-2-like is a positive modulator of fecundity and population growth in Nilaparvata lugens (Stal) (Hemiptera: Delphacidae). Front. Physiol. 2019, 10, 747. [Google Scholar] [CrossRef] [PubMed]
- Deady, L.D.; Shen, W.; Mosure, S.A.; Spradling, A.C.; Sun, J. Matrix metalloproteinase 2 is required for ovulation and corpus luteum formation in Drosophila. Plos Genet. 2015, 11, e1004989. [Google Scholar] [CrossRef] [Green Version]
- Saldivar-Hernandez, A.; Gonzalez-Gonzalez, M.E.; Sanchez-Tusie, A.; Maldonado-Rosas, I.; Lopez, P.; Trevino, C.L.; Larrea, F.; Chirinos, M. Human sperm degradation of zona pellucida proteins contributes to fertilization. Reprod. Biol. Endocrinol. 2015, 13, 99. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Bansal, P.; Ganguly, A.; Bhandari, B.; Chakrabarti, K. Human zona pellucida glycoproteins: Functional relevance during fertilization. J. Reprod. Immunol. 2009, 83, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Bhandari, B.; Shrestha, A.; Biswal, B.K.; Palaniappan, C.; Malhotra, S.S.; Gupta, N. Mammalian zona pellucida glycoproteins: Structure and function during fertilization. Cell Tissue Res. 2012, 349, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Guzeloglu-Kayisli, O.; Lalioti, M.D.; Aydiner, F.; Sasson, I.; Ilbay, O.; Sakkas, D.; Lowther, K.M.; Mehlmann, L.M.; Seli, E. Embryonic poly(A)-binding protein (EPAB) is required for oocyte maturation and female fertility in mice. Biochem. J. 2012, 446, 47–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galagovsky, D.; Katz, M.J.; Acevedo, J.M.; Sorianello, E.; Glavic, A.; Wappner, P. The Drosophila insulin-degrading enzyme restricts growth by modulating the PI3K pathway in a cell-autonomous manner. Mol. Biol. Cell 2014, 25, 916–924. [Google Scholar] [CrossRef]
- Bourouh, M.; Dhaliwal, R.; Rana, K.; Sinha, S.; Guo, Z.; Swan, A. Distinct and overlapping requirements for Cyclins A, B, and B3 in Drosophila female meiosis. G3 Genes Genomes Genet. 2016, 6, 3711–3724. [Google Scholar] [CrossRef] [Green Version]
- McCleland, M.L.; Farrell, J.A.; O’Farrell, P.H. Influence of cyclin type and dose on mitotic entry and progression in the early Drosophila embryo. J. Cell Biol. 2009, 184, 639–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vardy, L.; Pesin, J.A.; Orr-Weaver, T.L. Regulation of Cyclin A protein in meiosis and early embryogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 1838–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegler, R.; Van Antwerpen, R. Lipid uptake by insect oocytes. Insect Biochem. Molec. 2006, 36, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.M.; Wang, P.; Brown, C.A.; Zilinski, C.A.; Matzuk, M.M. Zygote arrest 1 (Zar1) is an evolutionarily conserved gene expressed in vertebrate ovaries. Biol. Reprod. 2003, 69, 861–867. [Google Scholar] [CrossRef]
- Burkart, A.D.; Xiong, B.; Baibakov, B.; Jimenez-Movilla, M.; Dean, J. Ovastacin, a cortical granule protease, cleaves ZP2 in the zona pellucida to prevent polyspermy. J. Cell Biol. 2012, 197, 37–44. [Google Scholar] [CrossRef] [Green Version]
- He, W.X.; Wu, M.; Liu, Z.; Li, Z.; Wang, Y.; Zhou, J.; Yu, P.; Zhang, X.J.; Zhou, L.; Gui, J.F. Oocyte-specific maternal Slbp2 is required for replication-dependent histone storage and early nuclear cleavage in zebrafish oogenesis and embryogenesis. RNA 2018, 24, 1738–1748. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Patocka, J.; Kuca, K. Insect antimicrobial peptides, a mini review. Toxins 2018, 10, 461. [Google Scholar] [CrossRef]
- Roy, S.; Aravind, P.; Madhurantakam, C.; Ghosh, A.K.; Sankaranarayanan, R.; Das, A.K. Crystal structure of a fungal protease inhibitor from Antheraea mylitta. J. Struct. Biol. 2009, 166, 79–87. [Google Scholar] [CrossRef]
- Ragan, E.J.; An, C.; Jiang, H.; Kanost, M.R. Roles of haemolymph proteins in antimicrobial defences of Manduca sexta. In Insect Infection and Immunity: Evolution, Ecology, and Mechanisms; Rolff, J., Reynolds, S.E., Eds.; Oxford University Press: Oxford, UK, 2009; pp. 34–48. [Google Scholar]
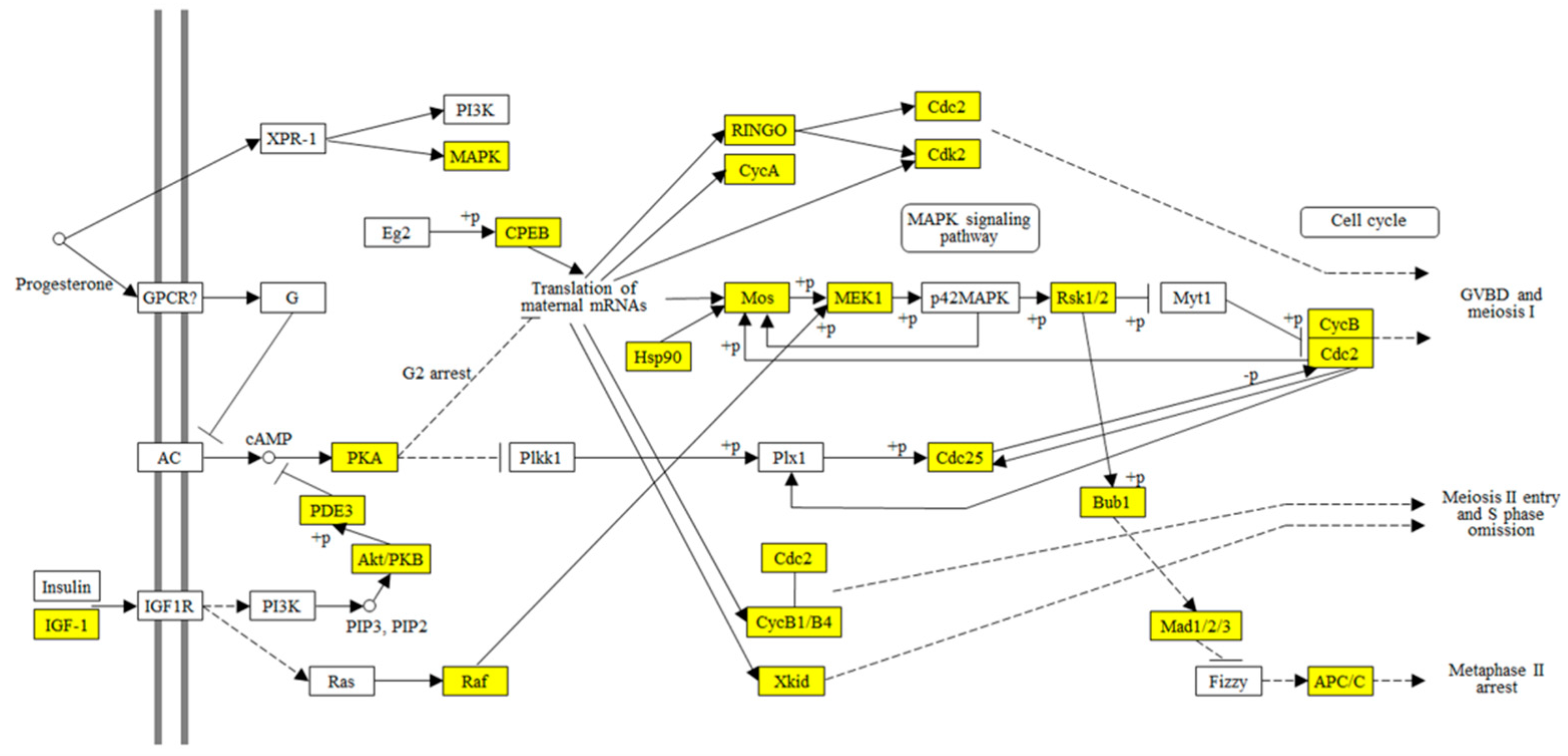
- Perez, L.H.; Antonio, C.; Flament, S.; Vernos, I.; Nebreda, A.R. Xkid chromokinesin is required for the meiosis I to meiosis II transition in Xenopus laevis oocytes. Nat. Cell Biol. 2002, 4, 737–742. [Google Scholar] [CrossRef]
- Schmitt, A.; Nebreda, A.R. Signalling pathways in oocyte meiotic maturation. J. Cell Sci. 2002, 115, 2457–2459. [Google Scholar] [CrossRef]
- Litscher, E.S.; Wassarman, P.M. A Guide to Zona Pellucida Domain Proteins; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Lu, F.-K.; Tang, S.-M.; Shen, X.-J.; Wang, N.; Zhao, Q.-L.; Zhang, G.-Z.; Guo, X.-J. Molecular cloning and characterization of hatching enzyme-like gene in the silkworm, Bombyx mori. Mol. Biol. Rep. 2010, 37, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Innocenti, P.; Morrow, E.H. Immunogenic males: A genome-wide analysis of reproduction and the cost of mating in Drosophila melanogaster females. J. Evol. Biol. 2009, 22, 964–973. [Google Scholar] [CrossRef] [PubMed]
- Will, T.; Schmidtberg, H.; Skaljac, M.; Vilcinskas, A. Heat shock protein 83 plays pleiotropic roles in embryogenesis, longevity, and fecundity of the pea aphid Acyrthosiphon pisum. Dev. Genes. Evol. 2017, 227, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, Y.; Teramura, K.; Takahashi, K.H. Heat shock proteins mediate trade-offs between early-life reproduction and late survival in Drosophila melanogaster. Physiol. Entomol. 2014, 39, 304–312. [Google Scholar] [CrossRef]
- Jiang, H.; Vilcinskas, A.; Kanost, M.R. Immunity in lepidopteran insects. In Invertebrate Immunity; Soderhall, K., Ed.; Springer: Boston, MA, USA, 2010; Volume 708, pp. 181–204. [Google Scholar]
- Xu, J.; Wang, Q. Seminal fluid reduces female longevity and stimulates egg production and sperm trigger oviposition in a moth. J. Insect Physiol. 2011, 57, 385–390. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GeneID | log2FoldChange | Padj | Annotation | Function | Reference |
|---|---|---|---|---|---|
| Mated-1 h vs. Virgin-1 h | |||||
| DN64818_c0_g6 | −3.51325 | 0.02376 | Follicle cell protein 3C | Encodes a major protein component of the vitelline membrane. | [33] |
| DN64030_c0_g1 | 9.66278 | 0.00015 | CYP2C (Cytochrome P450 family) | Detoxification and the biosynthesis of hormones. | [34] |
| Mated-6 h vs. Virgin-6 h | |||||
| DN65980_c5_g3 | −9.14964 | 1.05 × 10−8 | Vitellogenin-2 | Plays vital role in oocytes and embryo development in insects. | [35,36] |
| N55390_c2_g3 | −7.75002 | 0.00289 | Vitellogenin-A2-like | ||
| N52188_c2_g1 | −6.37534 | 0.00062 | Vitellogenin-A2-like | ||
| DN71364_c1_g1 | −2.39804 | 2.43 × 10−11 | Vitellogenin-like | ||
| DN59614_c4_g1 | −2.34561 | 1.55 × 10−10 | Vitellogenin-like | ||
| DN78694_c0_g1 | −3.45314 | 0.03932 | Haemolymph juvenile hormone binding protein | Protect the labile hormone molecules from degradation by esterase. | [37,38] |
| DN63882_c0_g1 | −2.75932 | 0.00064 | |||
| DN65663_c1_g1 | −2.48229 | 1.04 × 10−8 | |||
| DN65191_c2_g1 | −2.34190 | 0.02528 | Protein takeout-like | Takeout is part of a large gene family found throughout insects with roles in metabolism, circadian behavior, aging, and male courtship behavior. | [39] |
| DN69612_c3_g1 | −1.58633 | 0.03611 | Protein takeout-like | ||
| DN66978_c2_g1 | −2.71257 | 0.00840 | UDP-glucosyltransferase | Positive modulator of fecundity. | [40] |
| DN71428_c0_g1 | −8.16014 | 9.90 × 10−5 | Matrix metalloproteinase-16 | Some members of matrix metalloproteinase may play roles in corpus luteum formation, follicular development, and ovulation. | [41] |
| DN69614_c4_g4 | −2.86011 | 0.04696 | CYP4 (Cytochrome P450 family) | Detoxification and the biosynthesis of hormones. | [34] |
| DN52313_c1_g5 | −2.81364 | 3.33 × 10−14 | CYP4 (Cytochrome P450 family) | ||
| DN69106_c0_g1 | −2.15773 | 0.00170 | CYP4 (Cytochrome P450 family) | ||
| DN53829_c6_g2 | −2.00810 | 0.01412 | CYP4 (Cytochrome P450 family) | ||
| Mated-24 h vs. Virgin-24 h | |||||
| DN65980_c5_g3 | −9.44195 | 1.71 × 10−14 | Vitellogenin-2 | Plays vital role in oocytes and embryo development in insects. | [35,36] |
| DN71047_c1_g1 | −8.99780 | 8.17 × 10−11 | Vitellogenin-A2-like | ||
| DN55390_c2_g3 | −8.55112 | 1.39 × 10−7 | Vitellogenin-A2-like | ||
| DN52188_c2_g1 | −8.51720 | 2.32 × 10−7 | Vitellogenin-A2-like | ||
| DN49694_c0_g1 | −2.51669 | 0.00038 | Vitellogenin-like | ||
| DN59614_c4_g1 | −2.30309 | 0.02439 | Vitellogenin-like | ||
| DN71364_c1_g1 | −2.31618 | 0.02594 | Vitellogenin-like | ||
| DN54912_c7_g1 | 9.02029 | 1.51 × 10−11 | Vitellogenin receptor | ||
| DN58744_c7_g1 | 9.28253 | 8.58 × 10−14 | Zona pellucida sperm-binding protein (ZP) 1-like | ||
| DN32813_c0_g1 | 9.32544 | 3.72 × 10−14 | Zp1 | ||
| DN67043_c1_g1 | 11.27687 | 2.07 × 10−52 | Zp1 | ||
| DN44634_c0_g2 | 10.29005 | 2.97 × 10−23 | Zp2 | ||
| DN93387_c0_g1 | 7.70158 | 0.002245 | Zp3 | ||
| DN50223_c1_g2 | 8.04870 | 5.28 × 10−5 | Zp3 | ||
| DN12342_c0_g1 | 8.89036 | 1.66 × 10−10 | Zp3 | ||
| DN63118_c0_g5 | 9.43726 | 3.94 × 10−15 | Zp3 | ||
| DN42788_c0_g1 | 9.81002 | 1.13 × 10−18 | Zp3 | ||
| DN44618_c0_g1 | 9.87890 | 1.89 × 10−52 | Zp3 | The zona pellucida is a glycoprotein layer surrounding the plasma membrane of oocytes. It is a vital constitutive part of the oocyte and functions in primary binding and induction of the sperm acrosome reaction. | [42,43,44] |
| DN43523_c0_g1 | 10.0620 | 5.22 × 10−21 | Zp3 | ||
| DN22329_c0_g1 | 10.18637 | 3.23 × 10−22 | Zp3 | ||
| DN47551_c0_g1 | 10.48415 | 1.26 × 10−43 | Zp3 | ||
| DN48592_c0_g2 | 11.49147 | 9.56 × 10−37 | Zp3 | ||
| DN63118_c0_g2 | 11.49933 | 2.70 × 10−64 | Zp3 | ||
| DN47452_c0_g3 | 11.88468 | 1.13 × 10−41 | Zp3 | ||
| DN49748_c0_g1 | 12.50758 | 6.61 × 10−49 | Zp3 | ||
| DN100350_c0_g1 | 12.60210 | 5.74 × 10−50 | Zp3 | ||
| DN63118_c0_g1 | 13.09691 | 3.27 × 10−70 | Zp3 | ||
| DN67043_c2_g1 | 7.96939 | 4.36 × 10−26 | Zp4 | ||
| DN67043_c1_g2 | 8.81695 | 6.03 × 10−10 | Zp4 | ||
| DN72297_c0_g1 | 9.81754 | 9.66 × 10−19 | Zp4 | ||
| DN67043_c2_g3 | 10.63913 | 5.50 × 10−62 | Zp4 | ||
| DN67258_c0_g10 | 10.78996 | 1.26 × 10−28 | Zp4 | ||
| DN50530_c5_g14 | 9.970503 | 3.83 × 10−20 | ZP domain-containing protein-like | ||
| DN46494_c0_g1 | 12.88190 | 4.01 × 10−53 | ZP domain-containing protein-like | ||
| DN64127_c3_g3 | 9.64628 | 4.61 × 10−17 | Embryonic poly(A)-binding protein A | Required for oocyte maturation and female fertility. | [45] |
| DN82911_c0_g1 | 8.20682 | 7.04 × 10−6 | Insulinase | Destroys or inactivates insulin. Elevated insulin promotes oogenesis and inhibits immune responses. | [1,46] |
| DN49876_c0_g1 | 8.44020 | 2.62 × 10−7 | Insulinase | ||
| DN46576_c0_g1 | 8.18360 | 9.56 × 10−6 | G1/S-specific cyclin-E-like | Oocyte maturation and the onset of embryogenesis. | [47,48,49] |
| DN49363_c0_g1 | 9.49947 | 1.08 × 10−15 | G2/mitotic-specific cyclin-A | ||
| DN49147_c0_g1 | 11.21366 | 1.90 × 10−33 | G2/mitotic-specific cyclin-B | ||
| DN15801_c0_g1 | 9.55009 | 3.72 × 10−16 | G2/mitotic-specific cyclin-B | ||
| DN47388_c2_g1 | 10.19335 | 2.76 × 10−22 | G2/mitotic-specific cyclin-B-like | ||
| DN49534_c0_g1 | 9.76583 | 2.58 × 10−46 | G2/mitotic-specific cyclin-B-like isoform X4 | ||
| DN8940_c0_g1 | 10.33337 | 1.07 × 10−23 | Cell division cycle protein 20 homolog | ||
| DN50119_c0_g2 | −6.06256 | 3.33 × 10−13 | Apolipophorin III | Plays roles in lipid uptake by insect oocytes. | [50] |
| DN71428_c0_g1 | −8.39452 | 1.39 × 10−6 | Matrix metalloproteinase-16 | Some members of matrix metalloproteinase may play roles in corpus luteum formation, follicular development, and ovulation. | [41] |
| DN56705_c2_g7 | −3.79775 | 0.01004 | Matrix metalloproteinase-24-like | ||
| DN37027_c0_g1 | 11.90835 | 5.60 × 10−42 | Zygote arrest protein 1 | Ovary-specific maternal factor that plays essential roles during the oocyte-to-embryo transition. | [51] |
| DN94405_c0_g1 | 12.16383 | 4.47 × 10−45 | Zygote arrest protein 1 | ||
| DN84630_c0_g1 | 8.95337 | 5.28 × 10−11 | Astacin-like; Ovastacin | Cleaves ZP2 and prevents polyspermy; degrades egg envelope and releases the embryo from its intracapsular life. | [52] |
| DN47329_c0_g1 | 10.45829 | 5.41 × 10−25 | Oocyte-specific histone RNA stem-loop-binding protein 2 | Plays a role in the storage of replication-dependent histone mRNAs and proteins during oogenesis. | [53] |
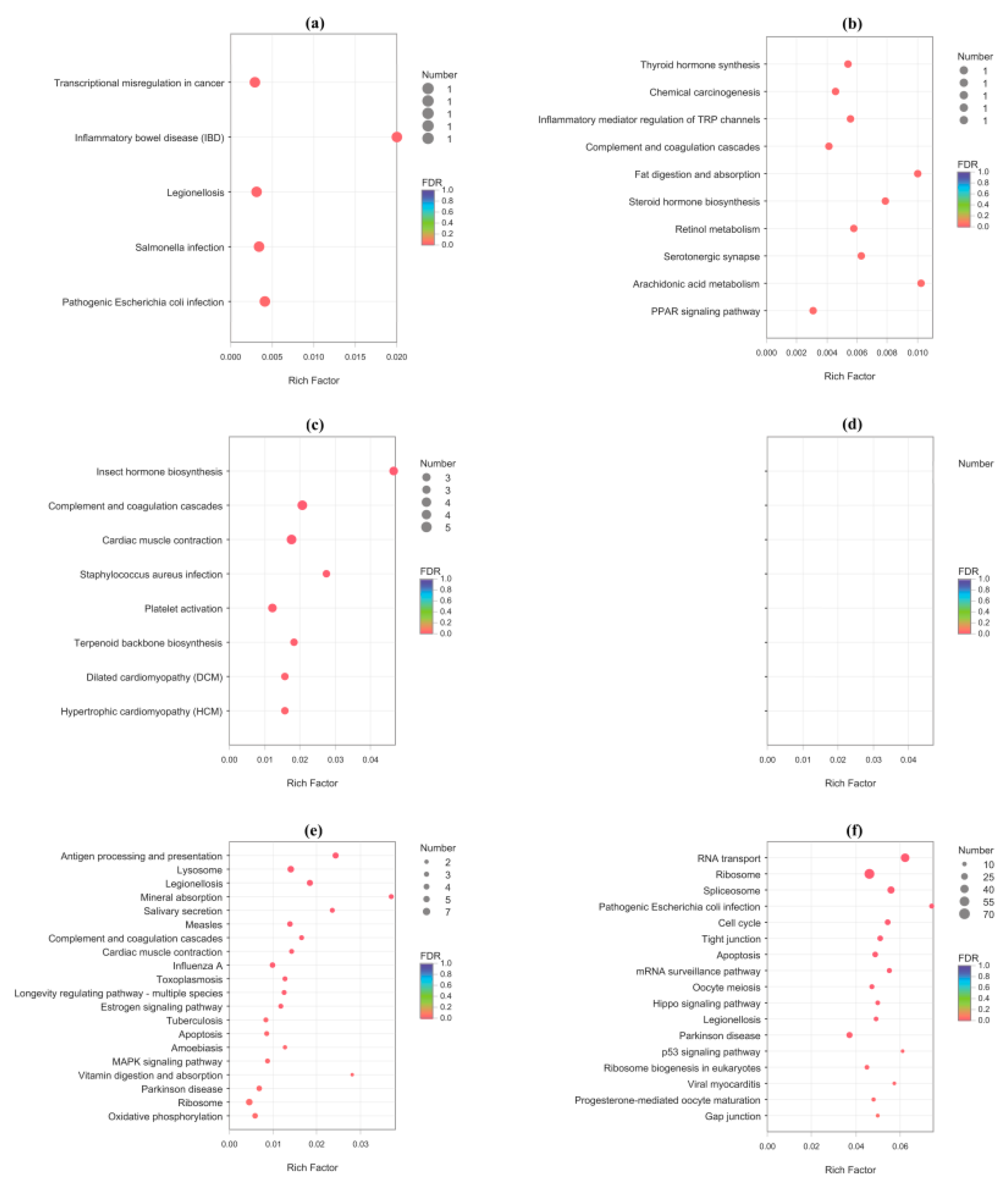
| Pathway | Padj | Number of Genes | First Category | Second Category | Function |
|---|---|---|---|---|---|
| Mated-1 h vs. Virgin-1 h: Up regulated DEGs | |||||
| Steroid hormone biosynthesis | 0.03602 | 1 | Metabolism | Lipid metabolism | Egg maturation |
| Mated-24 h vs. Virgin-24 h: Down regulated DEGs | |||||
| Estrogen signaling pathway | 0.01301 | 4 | Organismal Systems | Endocrine system | Fecundity |
| Mated-24 h vs. Virgin-24 h: Up regulated DEGs | |||||
| Oocyte meiosis | 0.01082 | 18 | Cellular Processes | Cell growth and death | Egg maturation |
| Progesterone-mediated oocyte maturation | 0.03177 | 13 | Organismal Systems | Endocrine system | Egg maturation |
| GeneID | log2FoldChange | Padj | Annotation | Function | Reference |
|---|---|---|---|---|---|
| Mated-1 h vs. Virgin-1 h | |||||
| DN70573_c2_g1 | −5.42272 | 1.27 × 10−9 | Defensin | Insect antimicrobial peptides. | [54] |
| DN60391_c5_g2 | −5.02886 | 2.84 × 10−8 | Hymenoptaecin | ||
| Mated-6 h vs. Virgin-6 h | |||||
| DN63597_c1_g1 | −3.53279 | 4.53 × 10−12 | Phenoloxidase | Phenoloxidase involved in defensive melanization and production of oxidative free radicals. | [1] |
| Mated-24 h vs. Virgin-24 h | |||||
| DN45974_c0_g1 | −6.77749 | 5.08 × 10−7 | Megourin | Insect antimicrobial peptides. | [54] |
| DN45254_c0_g1 | −6.34763 | 4.15 × 10−17 | Fungal protease inhibitor-1-like | Against fungal infection. | [55] |
| DN71273_c1_g5 | −2.95068 | 0.00896 | C-type lysozyme | Defends against bacterial infection by hydrolyzing the bacterial cell walls and causing bacterial lysis. | [56] |
| DN51648_c2_g8 | −6.63010 | 9.23 × 10−11 | Heat shock protein 27.2 (sHsp family) | Heat shock and stress response. | [4] |
| DN47336_c0_g1 | −5.01800 | 1.63 × 10−12 | Heat shock protein 27.2 (sHsp family) | ||
| DN71108_c1_g3 | −4.29401 | 3.40 × 10−11 | Heat shock protein 27.2 (sHsp family) | ||
| DN56298_c0_g1 | −3.69216 | 0.02212 | Heat shock protein 27.2 (sHsp family) | ||
| DN70765_c7_g9 | −4.25919 | 1.24 × 10−13 | Heat shock protein 70-like (Hsp70 family) | ||
| DN57953_c3_g2 | −3.39554 | 5.58 × 10−10 | Heat shock protein 70-like (Hsp70 family) | ||
| DN62526_c5_g1 | −6.22937 | 1.06 × 10−7 | Heat shock protein 70 cognate (Hsp70 family) | ||
| DN62526_c5_g4 | −4.59306 | 0.04497 | Heat shock protein 70 cognate (Hsp70 family) | ||
| DN67613_c0_g10 | 10.89725 | 7.42 × 10−30 | Heat shock protein 70 (Hsp70 family) | ||
| DN67613_c0_g5 | 8.479329 | 6.81 × 10−21 | Heat shock protein 70 (Hsp70 family) | ||
| DN67090_c3_g8 | 8.29444 | 2.51 × 10−6 | Heat shock protein 70 domain containing protein (Hsp70 family) | ||
| DN64447_c3_g1 | 7.39234 | 0.03894 | Heat shock protein 75 (Hsp90 family) | ||
| DN64447_c4_g1 | 3.92852 | 4.91 × 10−11 | Heat shock protein 90 (Hsp90 family) | ||
| Pathway | Padj | Number of Genes | First Category | Second Category | Function |
|---|---|---|---|---|---|
| Mated-1 h vs. Virgin-1 h: Down regulated DEGs | |||||
| Inflammatory bowel disease (IBD) | 0.02764 | 1 | Human Diseases | Immune disease | Defense |
| Legionellosis | 0.02981 | 1 | Human Diseases | Infectious disease: bacterial | Defense |
| Salmonella infection | 0.03185 | 1 | Human Diseases | Infectious disease: bacterial | Defense |
| Pathogenic Escherichia coli infection | 0.03341 | 1 | Human Diseases | Infectious disease: bacterial | Defense |
| Mated-1 h vs. Virgin-1 h: Up regulated DEGs | |||||
| Inflammatory mediator regulation of TRP channels | 0.03394 | 1 | Organismal Systems | Sensory system | Defense |
| Complement and coagulation cascades | 0.03397 | 1 | Organismal Systems | Immune system | Defense |
| Mated-6 h vs. Virgin-6 h: Down regulated DEGs | |||||
| Complement and coagulation cascades | 0.00061 | 5 | Organismal Systems | Immune system | Defense |
| Staphylococcus aureus infection | 0.00791 | 3 | Human Diseases | Infectious disease: bacterial | Defense |
| Platelet activation | 0.01418 | 4 | Organismal Systems | Immune system | Defense |
| Mated-24 h vs. Virgin-24 h: Down regulated DEGs | |||||
| Antigen processing and presentation | 0.00018 | 6 | Organismal Systems | Immune system | Defense |
| Legionellosis | 0.00046 | 6 | Human Diseases | Infectious disease: bacterial | Defense |
| Measles | 0.00319 | 5 | Human Diseases | Infectious disease: viral | Defense |
| Complement and coagulation cascades | 0.00620 | 4 | Organismal Systems | Immune system | Defense |
| Influenza A | 0.00987 | 5 | Human Diseases | Infectious disease: viral | Defense |
| Toxoplasmosis | 0.01130 | 4 | Human Diseases | Infectious disease: parasitic | Defense |
| Tuberculosis | 0.02795 | 4 | Human Diseases | Infectious disease: bacterial | Defense |
| Amoebiasis | 0.02986 | 3 | Human Diseases | Infectious disease: parasitic | Defense |
| Staphylococcus aureus infection | 0.04516 | 2 | Human Diseases | Infectious disease: bacterial | Defense |
| Longevity regulating pathway - multiple species | 0.01139 | 4 | Organismal Systems | Aging | Aging |
| Mated-24 h vs. Virgin-24 h: Up regulated DEGs | |||||
| Pathogenic Escherichia coli infection | 6.25 × 10−5 | 18 | Human Diseases | Infectious disease: bacterial | Defense |
| Legionellosis | 0.01178 | 16 | Human Diseases | Infectious disease: bacterial | Defense |
| Viral myocarditis | 0.02943 | 10 | Human Diseases | Cardiovascular disease | Defense |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, H.; Shi, M.-R.; Xu, J.; Chen, P.; Liu, J.-H. Mating-Induced Trade-Offs upon Egg Production versus Fertilization and Offspring’s Survival in a Sawfly with Facultative Parthenogenesis. Insects 2021, 12, 693. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080693
Yu H, Shi M-R, Xu J, Chen P, Liu J-H. Mating-Induced Trade-Offs upon Egg Production versus Fertilization and Offspring’s Survival in a Sawfly with Facultative Parthenogenesis. Insects. 2021; 12(8):693. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080693
Chicago/Turabian StyleYu, Hong, Min-Rui Shi, Jin Xu, Peng Chen, and Jian-Hong Liu. 2021. "Mating-Induced Trade-Offs upon Egg Production versus Fertilization and Offspring’s Survival in a Sawfly with Facultative Parthenogenesis" Insects 12, no. 8: 693. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080693