
New Fossil Tingidae (Hemiptera: Heteroptera) from the Mid-Cretaceous of Myanmar, with Remarks on the Phylogenetic Relationships within the Family


{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Terminology
2.2. Taxon Sampling and Character Choice
2.3. Phylogenetic Analysis
3. Results
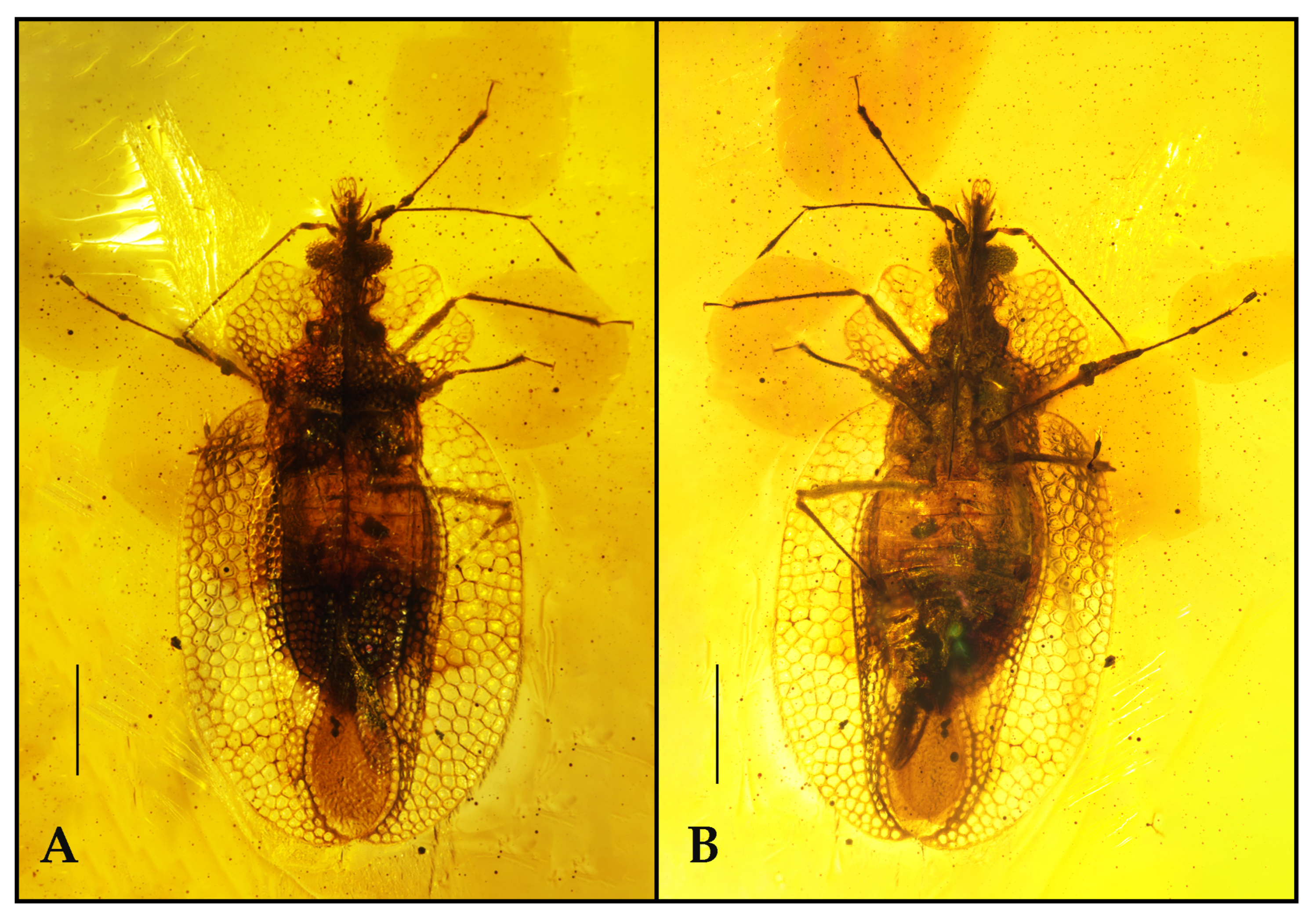
3.1. Systematic Paleontology
Description
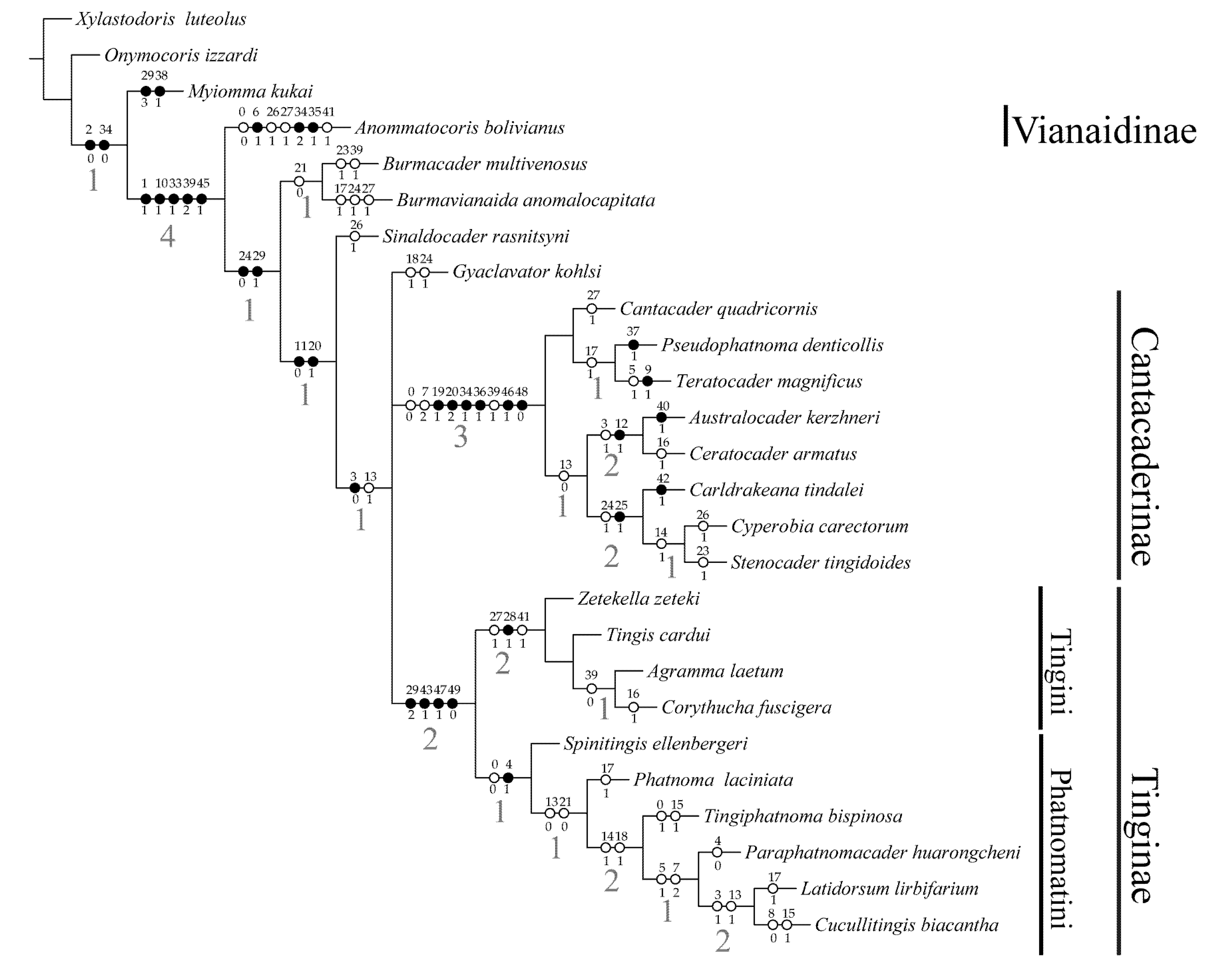
3.2. Phylogenetic Analysis
4. Discussion
4.1. Clade Burmacader Multivenosus + Burmavianaida Anomalocapitata and Clade Sinaldocader Rasnitsyni
4.2. Clade Gyaclavator Kohlsi + Tinginae+ Cantacaderinae
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guidoti, M.; Montemayor, S.I.; Guilbert, E. Lace Bugs (Tingidae). In True Bugs (Heteroptera) of the Neotropics; Panizzi, A.R., Grazia, J., Eds.; Springer Science + Business Media Dordrecht: Dordrecht, The Netherlands, 2015; Volume 2, pp. 395–419. [Google Scholar]
- Schuh, R.T.; Weirauch, C. True Bugs of the World (Hemiptera: Heteroptera). Classification and Natural History, 2nd ed.; Siri Scientific Press: Manchester, UK, 2020; pp. 1–768. [Google Scholar]
- Drake, C.J.; Ruhoff, F.A. Lacebugs of the world: A catalog (Hemiptera: Tingidae). Bull. United States Natl. Mus. 1965, 243, 1–634. [Google Scholar] [CrossRef]
- Lis, B. Phylogeny and classification of Cantacaderini [Cantacaderidae stat. nov.] (Hemiptera: Tingoidea). Ann. Zool. 1999, 49, 157–196. [Google Scholar]
- Schuh, R.T.; Cassis, G.; Guilbert, E. Description of the first recent macropterous species of Vianaidinae (Heteroptera: Tingidae) with comments on the phylogenetic relationships of the family within the Cimicomorpha. J. N. Y. Entomol. Soc. 2006, 114, 38–53. [Google Scholar] [CrossRef]
- Guilbert, E.; Damgaard, J.; D’Haese, C. Phylogeny of the lacebugs (Insecta: Heteroptera: Tingidae) using morphological and molecular data. Syst. Entomol. 2014, 39, 431–441. [Google Scholar] [CrossRef]
- Wappler, T.; Guilbert, E.; Labandeira, C.C.; Hörnschemeyer, T.; Wedmann, S. Morphological and behavioral convergence in Extinct and Extant bugs: The systematics and biology of a new unusual fossil lace bug from the Eocene. PLoS ONE 2015, 10, e0133330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilbert, E.; Heiss, E. New lacebugs (Hemiptera: Heteroptera: Tingidae) from Upper Cretaceous Burmese amber. Cretac. Res. 2019, 94, 72–79. [Google Scholar] [CrossRef]
- Du, S.; Yao, Y. A new genus and species of Tingidae (Heteroptera: Cimicomorpha) from Myanmar, with the analysis of the evolution of hood, carinae and paranota. Zool. Syst. 2018, 43, 283–293. [Google Scholar] [CrossRef]
- Heiss, E.; Golub, V.B. A new genus and species of Tingidae from Cretaceous Burmese amber (Hemiptera, Heteroptera). Palaeoentomology 2021, 4, 137–140. [Google Scholar] [CrossRef]
- Heiss, E.; Guilbert, E. Two new genera and species of Tingidae from Cretaceous amber from Myanmar (Burma) (Hemiptera: Heteroptera). Zootaxa 2013, 3736, 379–386. [Google Scholar] [CrossRef]
- Heiss, E.; Guilbert, E. A new species of Tingidae from Cretaceous amber from Myanmar (Burma). (Hemiptera: Heteroptera). Zootaxa 2018, 4457, 339–345. [Google Scholar] [CrossRef]
- Heiss, E.; Guilbert, E. New species of Tingiphatnoma from Cretaceous Burmese amber (Heteroptera, Tingidae). Palaeoentomology 2019, 2, 340–344. [Google Scholar] [CrossRef]
- Heiss, E.; Guilbert, E. A new species of Burmacader Heiss & Guilbert, 2013 from Cretaceous Burmese amber (Hemiptera, Heteroptera, Tingidae). Palaeoentomology 2021, 4, 30–33. [Google Scholar] [CrossRef]
- Maksoud, S.; Azar, D.; Huang, D.Y. Tingiphatnoma andreneli sp. nov., a new tingid bug from the mid-Cretaceous Burmese amber (Heteroptera, Tingidae). Palaeoentomology 2019, 2, 576–580. [Google Scholar] [CrossRef]
- Souma, J.; Yamamoto, S.; Takahashi, Y. Discovery of the Smallest Lace Bug from Mid-Cretaceous of Northern Myanmar Supports the Hypothesis of a Miniaturization Phenomenon of Insects in Kachin Amber (Hemiptera: Heteroptera: Tingidae). Taxonomy 2021, 1, 60–68. [Google Scholar] [CrossRef]
- Souma, J.; Yamamoto, S.; Takahashi, Y. A new species of the fossil lace bug genus Burmacader (Hemiptera: Heteroptera: Tingidae) from mid-Cretaceous Kachin amber of northern Myanmar, with implications of male genital structures. Cretac. Res. 2021, 126, 104911. [Google Scholar] [CrossRef]
- Heiss, E.; Guilbert, E. New synonymy in Burmese amber Tingidae (Hemiptera, Heteroptera). Palaeoentomology 2021, 4, 202. [Google Scholar] [CrossRef]
- Heiss, E.; Golub, V.B.; Popov, J.A. A new subfamily, genus and species of Tingidae (Hemiptera: Heteroptera) from Burmese amber. Z. Arb.gem. Österr. Entomol. 2015, 67, 1–9. [Google Scholar]
- Golub, V.B.; Heiss, E. Three new species of the lacebug genus Tingiometra from Cretaceous Burmese amber (Hemiptera: Heteroptera: Tingidae: Tingiometrinae). Zootaxa 2020, 4767, 531–542. [Google Scholar] [CrossRef]
- Poinar, J.G.; Vega, F.E. A lacewing bug, Paleoanomala aptenus gen. et sp. nov. (Hemiptera: Tingidae), in mid-Cretaceous Burmese amber. Biosis Biol. Syst. 2020, 1, 65–71. [Google Scholar] [CrossRef]
- Ross, A.J. Burmese (Myanmar) amber checklist and bibliography 2018. Palaeoentomology 2019, 2, 22–84. [Google Scholar] [CrossRef]
- Ross, A.J. Supplement to the Burmese (Myanmar) amber checklist and bibliography, 2019. Palaeoentomology 2020, 3, 103–118. [Google Scholar] [CrossRef]
- Ross, A.J. Supplement to the Burmese (Myanmar) amber checklist and bibliography, 2020. Palaeoentomology 2021, 4, 57–76. [Google Scholar] [CrossRef]
- Westerweel, J.; Roperch, P.; Licht, A.; Dupont-Nivet, G.; Win, Z.; Poblete, F.; Ruffet, G.; Swe, H.H.; Thi, M.K.; Aung, D.W. Burma Terrane part of the Trans-Tethyan arc during collision with India according to palaeomagnetic data. Nat. Geosci. 2019, 12, 863–868. [Google Scholar] [CrossRef]
- Yu, T.; Kelly, R.; Mu, L.; Ross, A.; Kennedy, J.; Broly, P.; Xia, F.; Zhang, H.; Wang, B.; Dilcher, D. An ammonite trapped in Burmese amber. Proc. Nati. Acad. Sci. USA 2019, 116, 11345–11350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, A.; Mellish, C.; York, P.; Crighton, B. Burmese amber. In Biodiversity of Fossils in Amber from the Major World Deposits; David, P., Ed.; Siri Scientific Press: Manchester, UK, 2010; pp. 208–235. [Google Scholar]
- Shi, G.; Grimaldi, D.A.; Harlow, G.E.; Wang, J.; Wang, J.; Yang, M.; Lei, W.; Li, Q.; Li, X. Age constraint on Burmese amber based on U–Pb dating of zircons. Cretac. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Walker, J.D.; Geissman, J.W.; Bowring, S.A.; Babcock, L.E. The Geological Society of America time scale. Geol. Soc. Am. Bul. 2012, 125, 259–272. [Google Scholar] [CrossRef]
- Chen, S.; Deng, S.W.; Shih, C.K.; Zhang, W.W.; Zhang, P.; Ren, D.; Zhu, Y.N.; Gao, T.P. The earliest Timematids in Burmese amber reveal diverse tarsal pads of stick insects in the mid-Cretaceous. Insect Sci. 2019, 26, 945–957. [Google Scholar] [CrossRef]
- Page, R.D.M. NDE: NEXUS Data Editor 0.5.0. University of Glasgow [WWW document]. 2001. Available online: http://taxonomy.zoology.gla.ac.uk/rod/NDE/nde.html (accessed on 18 February 2011).
- Nixon, K.C. WinClada ver. 1.00.08; Published by the author: Ithaca, NY, USA, 2002; Available online: http://www.cladistics.com/ (accessed on 18 February 2011).
- Goloboff, P.A. NoName (NONA), version 2.0; Program and Documentation; Fundación Instituto Miguel Lillo: Tucumán, Argentina, 1997. [Google Scholar]
- Goloboff, P.A.; Catalano, S.A. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 2016, 32, 221–238. [Google Scholar] [CrossRef]
- Popov, Y.A.; Golub, V.B. A new tribe, genus and four species of lace bugs (Hemiptera: Heteroptera: Tingidae) from the Upper Cretaceous of Southwestern Kazakhstan. Paleontol. J. 2019, 53, 155–164. [Google Scholar] [CrossRef]
- Montagna, M.; Strada, L.; Dioli, P.; Tintori, A. The Middle Triassic Lagerstätten of Monte San Giorgio reveals the oldest lace bugs (Hemiptera: Tingidae): Archetingis ladinica gen. n. sp. n. Riv. Ital. Paleontol. Strat. 2018, 124, 35–44. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Tang, H.; Ren, D.; Yao, Y. New Fossil Tingidae (Hemiptera: Heteroptera) from the Mid-Cretaceous of Myanmar, with Remarks on the Phylogenetic Relationships within the Family. Insects 2021, 12, 887. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12100887
Wang Y, Tang H, Ren D, Yao Y. New Fossil Tingidae (Hemiptera: Heteroptera) from the Mid-Cretaceous of Myanmar, with Remarks on the Phylogenetic Relationships within the Family. Insects. 2021; 12(10):887. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12100887
Chicago/Turabian StyleWang, Yuxin, Huiying Tang, Dong Ren, and Yunzhi Yao. 2021. "New Fossil Tingidae (Hemiptera: Heteroptera) from the Mid-Cretaceous of Myanmar, with Remarks on the Phylogenetic Relationships within the Family" Insects 12, no. 10: 887. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12100887