1. Introduction
A circular economy has become a target objective in the movement towards sustainability. Insects play an important role in this change because their rearing has a small ecological footprint; less land and water is required, they emit fewer greenhouse gases, and they have a high feed conversion efficiency in comparison to traditional animal products. These factors should be considered in tandem with insects’ capacity to transform low-value organic by-products into high-quality food or feed that can be used as animal feed or aqua feed [
1].
The utilization of insect meal as a substitute in fishmeal has been studied in several fish species and insect species. High percentages of substitution have been used for feeding several species without compromising their growth ranges, as in the case of
Salmo salar [
2], yellow catfish [
3] and Nile tilapia [
4], with substitution levels of 100%, 75% and 68%, respectively. However, most of the studies reported 25% as the threshold at which growth indices can be negatively affected [
5]. Among the most studied species are
Hermetia illucens (HI) and
Tenebrio molitor (TM), which, in addition to meeting the approval of the EU as feed ingredients, are produced in sufficient quantities to sustain animal feed manufacturing.
The effect of the inclusion of HI in aqua feed has been analyzed in relation to many fish species and many different issues, such as disease resistance, innate immune response, growth performance, intestinal antioxidant enzymes and amino acid composition [
6]. Similar studies have been carried out for TM [
7,
8].
Sparus aurata, gilthead seabream, is a widely consumed fish in the Mediterranean area. The culture of
Sparus aurata is expanding, as evidenced by 96% of its total production (aquaculture + fisheries) corresponding to aquaculture in 2016. In the period of 2012–2016, cultivated production increased by 32%, and an additional 14% from 2016 to 2017, reaching 94,936 tons and EUR 485 million [
9]; this was mostly fueled by an increase in production in the Mediterranean Sea [
9]. Currently, feeds for seabream are based in fishmeal and soy as protein sources. Nevertheless, sources of soy and fishmeal present environmental problems. To reduce their drawbacks, insect meal can be considered as a potential substitute due to its more sustainable nature and broad acceptance [
10].
The inclusion of both insect species, HI and TM, in
S. aurata feed has been studied previously [
11]. Results pointed out the existence of a limit of substitution, 25–30%, above which growth indices worsened in comparison with fishmeal diets, although two recent studies by Pulido-Rodriguez et al. [
12] and Randazzo et al. [
13] reported similar or even better results with a level of substitution of 40% of vegetable protein mixed with HI meal. Nevertheless, higher percentages of substitution have been reported using other alternative protein sources, such as soy (40%) [
14] or pea protein concentrates (60%) [
15]. One of the main reasons for this limit of insect inclusion is attributed to the low digestibility of chitin and scleroprotein of insect exoskeletons [
10]. Another handicap that limits the inclusion of insects in fish diets is the fatty acid profile, which is different from fishmeal. Insects have a higher percentage of n-6 polyunsaturated fatty acids (
n-6 PUFAs), whereas fishmeal is richer in n-3 highly unsaturated fatty acids (
n-3 HUFAs). In this sense, and trying to overcome this last challenge, Fabrikov et al. [
16] obtained
n-3 HUFAs-rich larvae feeding HI in discards from fisheries, and Liland et al. [
17] produced similar results by feeding larvae with seaweed-enriched media.
Despite the performance of insect meal in relation to fish acceptance or its impact on growth parameters, the effects of high levels of inclusion on digestive and metabolic processes are not well known, and more knowledge is needed to realize the barriers precluding the use of insect meal in aquafeed. In this sense, it is also important to elucidate modifications in the gut microbiome due to the inclusion of insect meal in fish diets. Microbial communities associated with the intestinal tract play a decisive role not only in relation to nutrient assimilation, but regarding immunological responses and disease prevention [
18]. Among these communities, some are considered as persistent and others are understood as transient [
19], depending on their temporality. The presence of transient microbiota is subject to fluctuations caused by environmental factors, whereas persistent communities are less affected by such conditions and only minor quantitative modifications can be observed. Among the most influential factors regarding the composition of transient microbiota are those of a nutritional nature [
20]; these can promote the temporary presence of species more adapted to the properties of substrates used and, consequently, can foster the better utilization of a diet as a whole [
21]. Thus, the study of the gut microbiome and its response to changes in the nutritional composition of feeds can help to interpret the specific roles of different species and even select potential probiotics adapted to the type of nutrients being used. Ultimately, this understanding might encourage the better use of alternative protein sources and allow for the partial and even full replacement of traditional fishmeal-based diets.
Considering the abovementioned discussion, the main aim of this study was to investigate the effect of high levels of replacement of fishmeal with TM or HI meal—including meal with a modified acid profile—on growth and nutritive indices, digestive and metabolic enzymes and the gut microbiota of S. aurata fed with such diets. Elucidating potential correlations between growth parameters and microbial communities was also established as a primary goal.
4. Discussion
In recent years, the replacement of fishmeal in aquafeed has been studied widely. Specifically, in feed for
S. aurata, the level of inclusion of HI or TM meal without compromising growth has been established as being over 25–30% [
11,
38]. Higher replacement percentages need to be studied thoroughly to deepen knowledge regarding the digestive and metabolic responses of fish to insect meal ingestion. In this experiment, diets with high levels of substitution of fishmeal with insect meal (50:50) were studied, as was HI meal with a modified fatty acid profile according to the method described by Fabrikov et al. [
16].
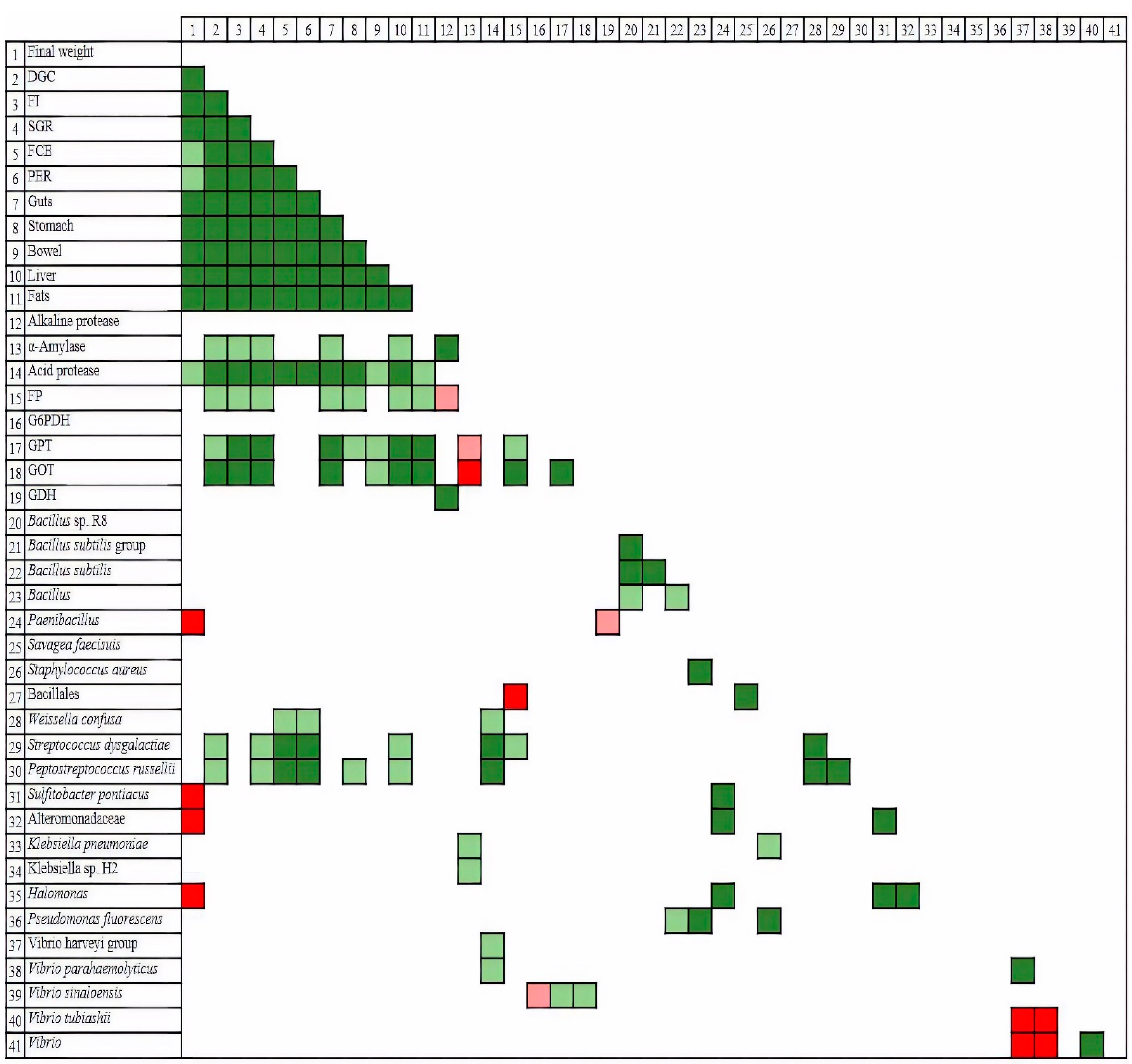
Nutritional and growth indices (
Table 4) showed better results for the control diet in all the indices determined, followed by T50. Piccolo et al. [
38] obtained similar growth rates for
S. aurata fed with a fishmeal-based diet and a diet including a 25% replacement with TM meal, which was lower than that in our experiment. Pulido-Rodriguez et al. [
12] and Randazzo et al. [
13] also obtained similar results with inclusion of levels of 32.4%. Other than different percentages of fishmeal substitution, these discrepancies might be related to varying specific growth rates, which were around 2 in our experiment versus values closer to 0.5 in Piccolo et al. [
35] and to 1.5 in Pulido-Rodriguez et al. [
12] and Randazzo et al. [
13]; these values are in accordance with the different initial weights used, i.e., 100 g and 48.8 g, respectively, versus 7 g in our experiment. FCE and PER also worsened in the T50 diet in comparison to C treatment, whereas no differences were observed in the study by Piccolo et al. [
38] at a replacement level of 25%.
Fish fed with HI meal, no matter the level of replacement, showed worse nutritional and growth indices, which agrees with results obtained by Karapanagiotidis et al. [
11]. Nevertheless, values observed for FCR and PER in the cases of H30 and H50M diets seem to be slightly better than those from H50. In the case of H50M, this result might be a consequence of the improved fatty acids profile of HI; however, in the case of H30, it could be due to the lower level of insect inclusion. The influence of insect meal on growth can also be illustrated with morphometric indices (
Table 4). Results obtained for these indices confirmed what the nutritional parameters pointed out, with HI diets producing lower values. However, no differences between C and T50 treatments were found in this case, which support the idea of the better nutritive quality of TM meal in comparison with HI for
S. aurata juveniles, especially when the latter does not improve nutritional profile.
Data regarding HI substitution levels higher than 30% in
S. aurata feeding are scarce, if non-existent, although some studies have been published with other species as subjects. Thus, Reyes et al. [
39] described similar results for
Dicentrarchus labrax, reporting a better acceptance of diets containing a 30% inclusion in comparison to those with a 50% replacement. In contrast, no differences in nutritional and growth indices were detected for growth responses between fish fed with diets including 30% HI meal or 50% TM meal, showing a slightly better nutritive utilization of HI by
D. labrax than by
S. aurata.
The response of fish to specific diets is dependent on digestibility, which, in turn, is primarily associated with protein assimilation [
38,
40]. As such, the differences in growth and nutritional indices between HI- and TM-based diets could be due to the different digestibility of feeds obtained for
S. aurata (CDA H30: 79.13; CDA H50: 79.16; CDA T50: 93.20). These values of digestibility are in agreement with those obtained in
D. labrax for HI [
6] and higher than those obtained in TM [
41]. The different digestibility in these two insect species contrast with those reported by Marono et al. [
42], who obtained an in vitro crude protein digestibility lower than in the present study but that was similar among different samples of HI and TM. Chitin content could also affect digestibility, and TM meal contains less chitin than HI meal (
Table 1), which may partly explain the different digestibility indices measured in each case. The high CDA found for H50M (CDA H50M: 91.57) might be attributable to microbiological factors, because chitin concentrations must be similar. According to the results, H50M-fed fish showed the most similar microbiome to those undergoing C treatment, and both were different from the rest of the diets. Such differentiation was mostly based on the higher population of members of the genus
Bacillus, especially, to those belonging to the
Bacillus subtilis group, whose chitinolytic properties are well recognized [
43].
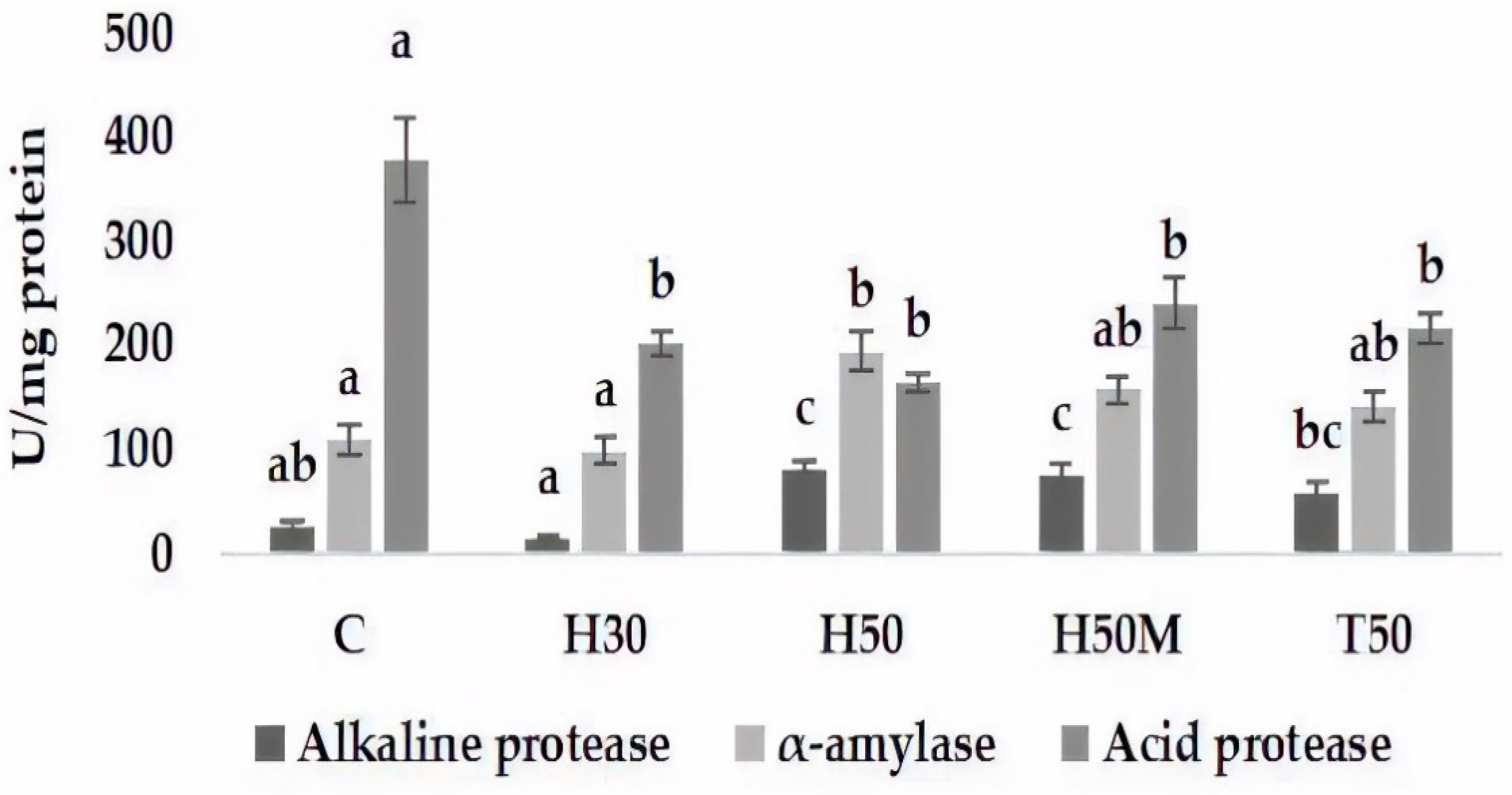
On the other hand, digestibility is also related to digestive enzyme activities. In this respect, the results indicate that insect meals induced changes in digestive enzyme activities, decreasing the activity of acid protease and increasing alkaline protease and alpha-amylase, especially for H50 and H50M. In trout fed with HI or TM meal at 15% or 30% fishmeal replacement, an increase in alkaline protease activity was also observed, although no affectation of acid protease activity was detected [
44]. Conversely, decreases in alkaline protease and trypsin activities were described in
Argyrosomus regius fed with a diet with 30% fishmeal replacement by TM [
45], whereas the inclusion of HI in
D. labrax feed did not affect intestinal protease activities [
6]. The digestibility of protein from insects can be highly variable, depending on the proportion of chitin [
40]. Coutinho et al. [
45] explain these changes as a slowdown in intestinal transit due to chitin that causes decreases in digestive enzyme activities. Nevertheless, protein digestibility is mostly influenced by the amino acid contents in ADF fractions [
46]. Hence, changes in digestive protease activities could be associated with the skleroprotein of an insect’s exoskeleton, in addition to the chitin effect, as proposed by Coutinho et al. [
45]. On the other hand, there is a clear connection between enzyme activity and dietary habits [
47]; both may influence and be influenced by the intestinal microbiome [
48]. Thus, the detection of different enzyme responses due to changes in feed and microbial communities should not be surprising.
Alpha-amylase activity showed a tendency to increase activity at high levels of insect inclusion. The results described by other authors vary among fish or insect species. For
D. labrax fed with a 45% replacement with HI [
6], or
Argyrosomus regius fed with a diet with 30% fishmeal replacement with TM [
45], no changes in alpha-amylase activity were detected; however, in
Oreochromis mossambicus and
Clarias gariepinus fed with
Imbrasia belina, an increased amylase activity was found [
49]. In rainbow trout
(Oncorhychus mykiss), the inclusion of 30% HI meal produced a reduction in alpha-amylase activity, whereas no effect was observed when TM meal was used. Reasons for such varying responses are not clear, especially due to the absence of data dealing with 1,4-α-glucose polymer content, i.e., the substrate for this enzyme.
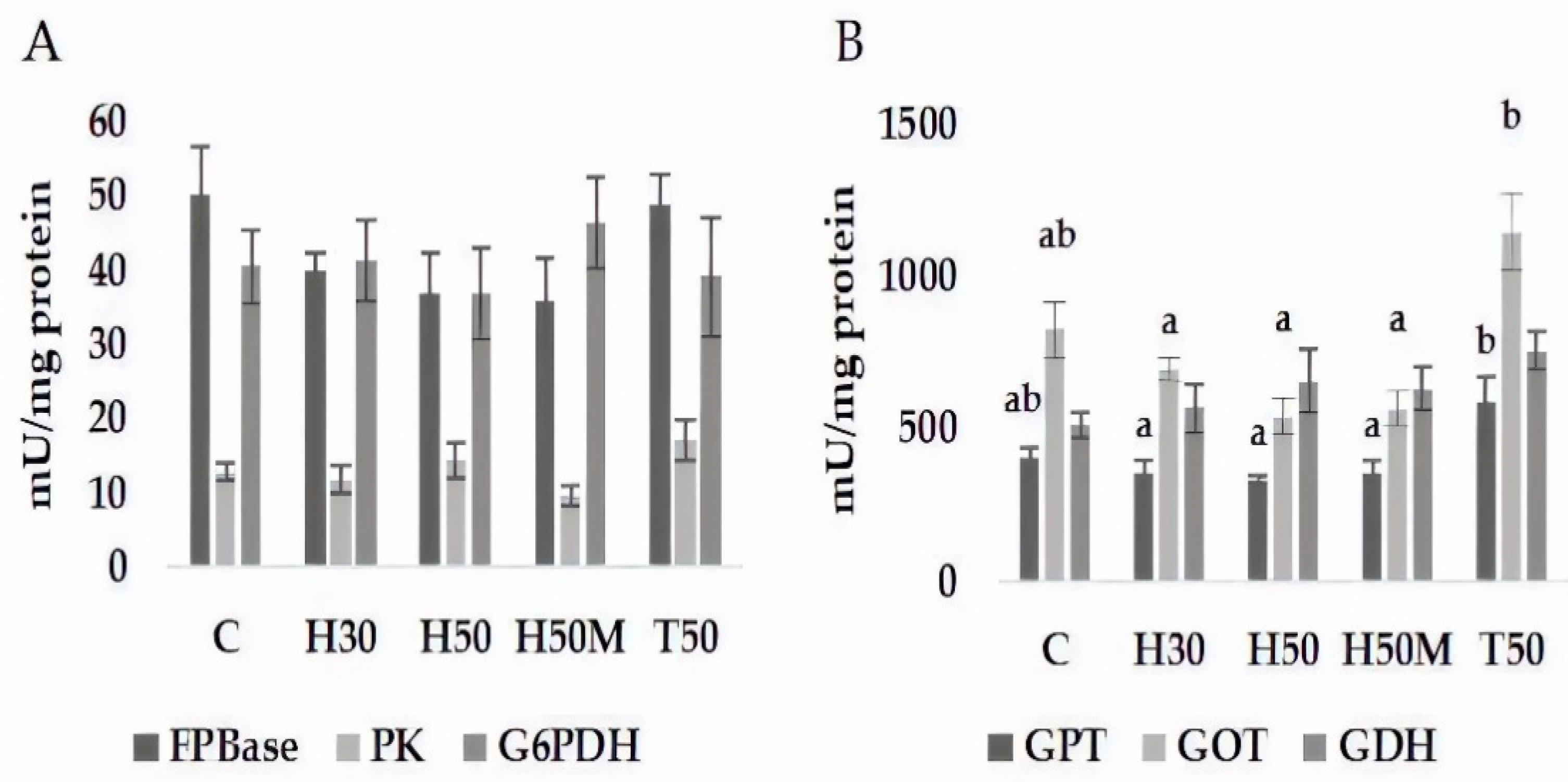
On the other hand, the inclusion of insect meal as an ingredient in diets did not seem to alter the intermediary metabolism of glucose or amino acids, because none of the analyzed enzymes involved in such processes showed significant changes in comparison to the C diet. Similar results were found for rainbow trout [
44] and sea trout (
Salmo trutta m.
trutta) at low levels of replacement of fishmeal with TM meal [
50]. Insect species induced changes in the Vmax of GPT between HI treatments regarding T50. Fabrikov et al. [
5] studied the adaptation of amino acid catabolism enzymes to HI or TM meal inclusion in the diet among different species, including
S. aurata. They found that Vmax exhibited an increase no matter the insect used for the formulation of the diet. However, in the present study, results showed different trends related to different insect species. Thus, although TM replicated the results from Fabrikov et al. [
5], i.e., the Vmax of GPT increased in value, HI tended to reduce activity. In the case of GOT, Fabrikov et al. [
5] did not find changes in the Vmax when HI meal was included in the diet, whereas an increase was observed when fishmeal was partially substituted by TM meal. These different responses could be attributed to the inclusion level—15% and 30% replacements in the experiment by Fabrikov et al. [
5] and 50% in this assay—which might have led to a reduction in feed intake in HI groups. This fact, as well as lower digestibility, potentially reduces the availability of amino acids to be deaminated and, consequently, decreases GOT activity. In T50-fed fish, feed intake is higher than in HI groups, with the subsequent increase in amino acid availability being utilized as energy or for gluconeogenesis purposes, promoting both transaminase activities (GPT and GOT).
4.1. Gut Microbiome Response to Dietary Insect Inclusion
Previous studies have reported variations in the gastrointestinal microbiota of fish due to modifications in their diets [
51]. In this case, the shaping of the microbiota is a logical response of the gut environment to variations in the nutritional composition of the diet, given the plasticity of bacterial communities and their rapid adaptation to new conditions [
21]. In relation to the partial replacement of fishmeal with insect meal, some studies described an increase in the diversity of the intestinal microbiome [
52], although opposite results have been obtained depending on insect and fish species, insect life-cycle stage or replacement level [
53,
54]. In the present work, different responses seem to be preferably related to the level of replacement rather than insect species, as revealed by the β diversity. Curiously, greater differences with regard to C diet were detected at the lowest level of inclusion, H30, which may seem contradictory, although this result has previously been reported [
53]. Microbial community structures are dependent on several factors, and the characteristics of diet is just one of them. Environmental, phylogenetic and physiological interactions come in a wide range of conditions that, in turn, can create particular habitats which affect and modulate the composition of the gut microbiome [
55]. In this sense, insect meal shows properties that can affect its acceptance by fish [
56], as is the case of chitin content and fat profile. Regarding the latter, HI is characterized by a high saturated fatty acid content, as well as an unbalanced fatty acid profile [
54] that can negatively affect fish performance. Therefore, it is common practice to provide defatted or fatty-modified meals with the potential to reduce undesirable effects and improve meal composition [
16]. Concerning the gut microbiome, our results seem to suggest that such treatments may promote a nutritional profile more akin to that of fishmeal, which caused a lower impact in bacterial communities in the intestinal tract. Therefore, these two diets, C and H50M, clustered together. The almost complete lack of studies comparing the effect of full-fat and defatted insect meal on the fish gut microbiome has made it difficult to validate such a hypothesis, although it has been described in relation to other species [
57]. Moreover, several documents point out the higher nutritional quality and digestibility of meals with a modified fat profile [
58].
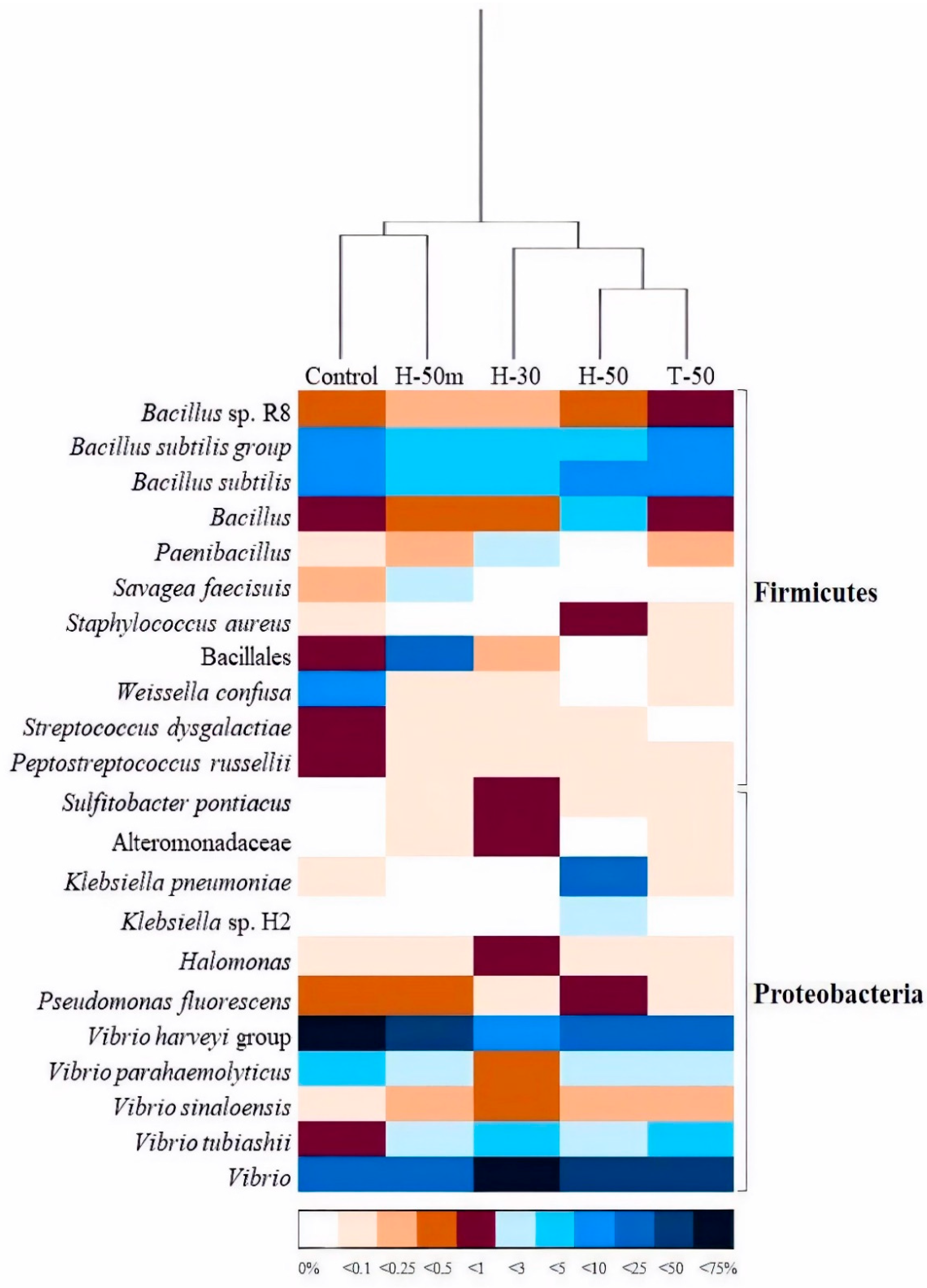
The dominance of
Vibrio and
Bacillus representatives in the fish gut microbiome is a common reality [
19]. Members of the
V. harveyi group, which is dominant in the C/H50M cluster, are associated with pathogenic processes [
59]. Thus, the handover from this group to
Vibrio sp. in the H30/H50/T50 cluster can be considered as positive, because species other than pathogens have been described in the genus, as in the case of
V. alginolyticus [
60], which has been postulated as a probiotic in aquaculture. The second largest microbial group was the
Bacillus genus and its relatives. The importance of these bacteria relies on their enzyme activity and, to a lesser extent, their immunostimulant ability and antimicrobial production [
61]. Taking into account their beneficial activities, and their presence in all treatments, it is clear that they have a positive impact on fish performance and wellbeing.
Apart from these dominant groups, other species were differentially present in the intestinal tract because of insect meal replacement. Bacteria such as
Streptococcus dysgalactiae,
Peptostreptococcus russellii, and especially
Weisella confusa, reached relatively important population densities in the C samples, although they were practically absent in the rest of the treatments.
W. confusa is a lactic acid bacterium which is associated with some infectious processes in human beings [
62], but not in fish [
63]. Moreover, it has been postulated as a probiotic for some species in aquaculture [
64].
P. russellii has been positively associated with intestinal homeostasis in different species [
65], although there is no literature about this in relation to fish. In contrast,
S. dysgalactiae have been described as pathogenic for both human and fish [
66]. However,
Sulfitobacter pontiacus and members of the family Alteromonadaceae, absent in C-fed fishes, were detected in insect meal samples, especially H30. The first is a recognized sulfite and thiosulfate-oxidizing bacterium that might promote organic matter degradation in aquatic environments [
67], whereas the latter comprises both pathogenic and potentially probiotic species [
68,
69].
In short, the replacement of fishmeal with insect meal seems to promote a redistribution of the most dominant bacterial populations, Vibrionaceae and Bacillaceae, whereas it modifies the identity of minor species. Nevertheless, and taking into account the diversity of conditions in which assays were performed, more information dealing with the impact of dietary modifications on intestinal bacterial communities is needed to attain a better understanding of the particular factors that rule the microbial response.
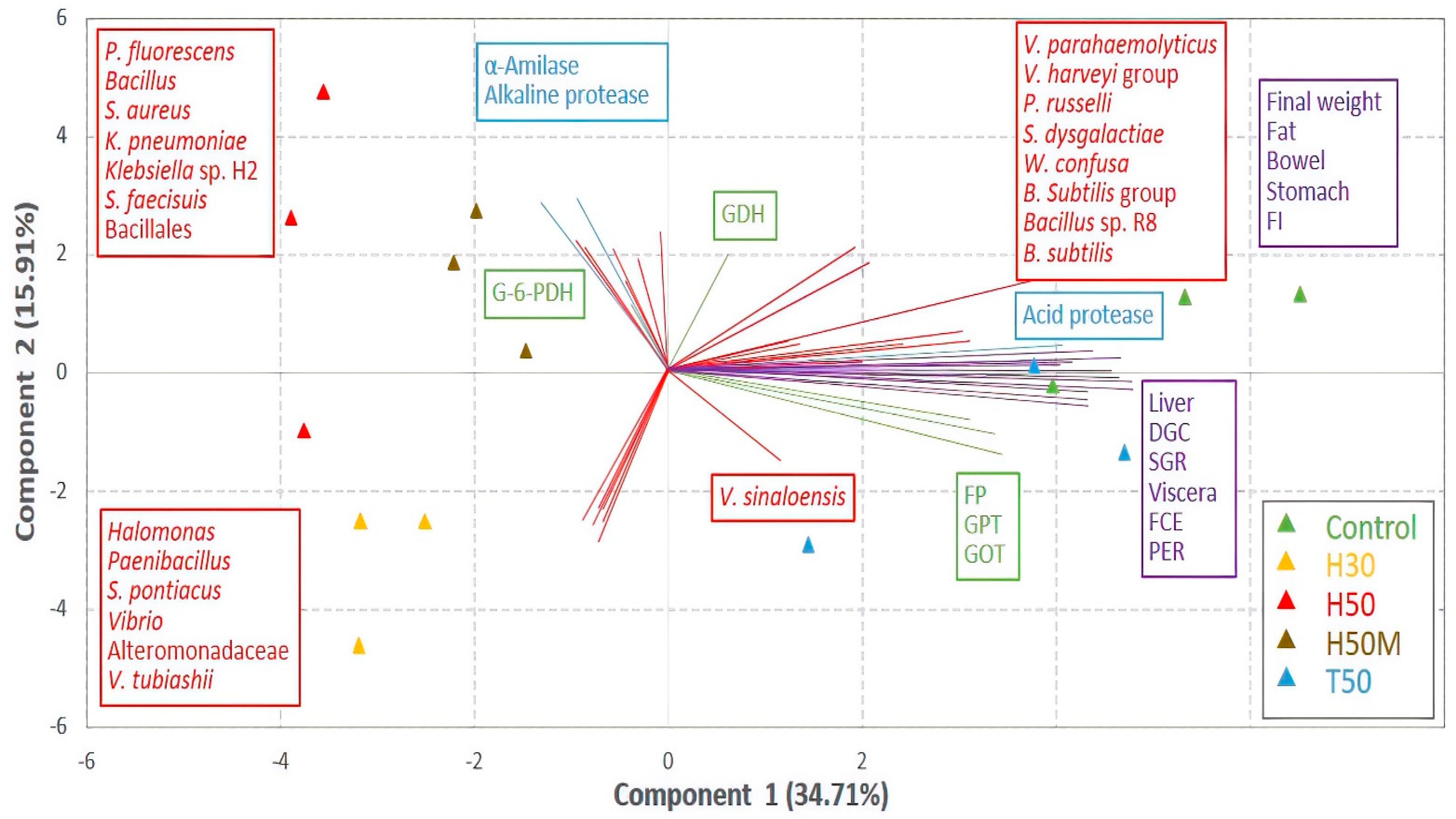
4.2. Gut Microbiome/Fish Performance Correlations
Gut microorganisms play different roles with respect to host performance and health. Beneficial bacteria can excrete enzymes that contribute to assimilating nutrients, reducing the availability of attachment sites for pathogens and synthesizing antimicrobial compounds [
18]. The so-called persistent microbiota, which comprise those bacteria residing permanently in the gut, are considered as main protagonists of the symbiotic relation between the gut microbiome and a host [
70]. Transient species, mostly associated with external environmental and nutritional factors, are thought to play some significant roles depending on their colonization ability [
71]. Thus, species able to survive the conditions in the gut and adhere to intestinal mucosa may become permanent residents [
72] and perform primary functionality [
73]. The result described in this paper showing the grouping of T50 and C samples, and leaving treatment most microbiologically similar to C, H50M, out of this group, may be explained by this theory. Moreover, the evolutionary closeness between species in T50 and C, which shows the possibility of sharing metabolic properties, sustains such a hypothesis. The positive relationship between both diets and fish performance was evident by means of pooling with all the biometric parameters and most of the metabolic enzymes. Relevant microorganisms in the gut microbiome of these treatments, such as the
Bacillus subtilis group, were also in this group, as well as
W. confusa,
S. dysgalactiae and
P. russellii. These latter three were statistically correlated with many of the biometric parameters, such DGC, SGR, FCE and PER, reflecting their involvement in the beneficial utilization of a diet by fish. Thus far, none of them have been considered as key players in fish performance.
S. dysgalactiae has even been associated with pathogenic processes [
66]. Nevertheless, it has been reported that this bacterium could be a member of the persistent gut microbiome in fish, although its specific origin is not yet clear [
74]. On the other hand, it has been referenced as a bacteriocin-producing bacteria [
75], which is a desirable characteristic on account of its ability to prevent the growth of some other pathogens. For its part,
W. confusa, which is also an antibacterial compound producer [
76], has shown potential to positively impact fish growth performance, as well as stimulate the activity of some digestive enzymes [
77]. Similarly,
P. russelli has been associated with beneficial effects in terms of fish health and growth, especially regarding activities dealing with protein degradation [
78], as elucidated in this paper.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}