Effects of Acetamiprid at Low and Median Lethal Concentrations on the Development and Reproduction of the Soybean Aphid Aphis glycines


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Laboratory Aphid Population and Chemical Agents
2.2. Preparation of Culture Medium
2.3. Selection of Acetamiprid Concentration
2.4. Dose–Response Bioassay
2.5. Life–History Study
2.6. Reproductive Parameter Calculation
3. Results
3.1. Dose–Response Bioassay with the First Instar A. glycines Nymphs
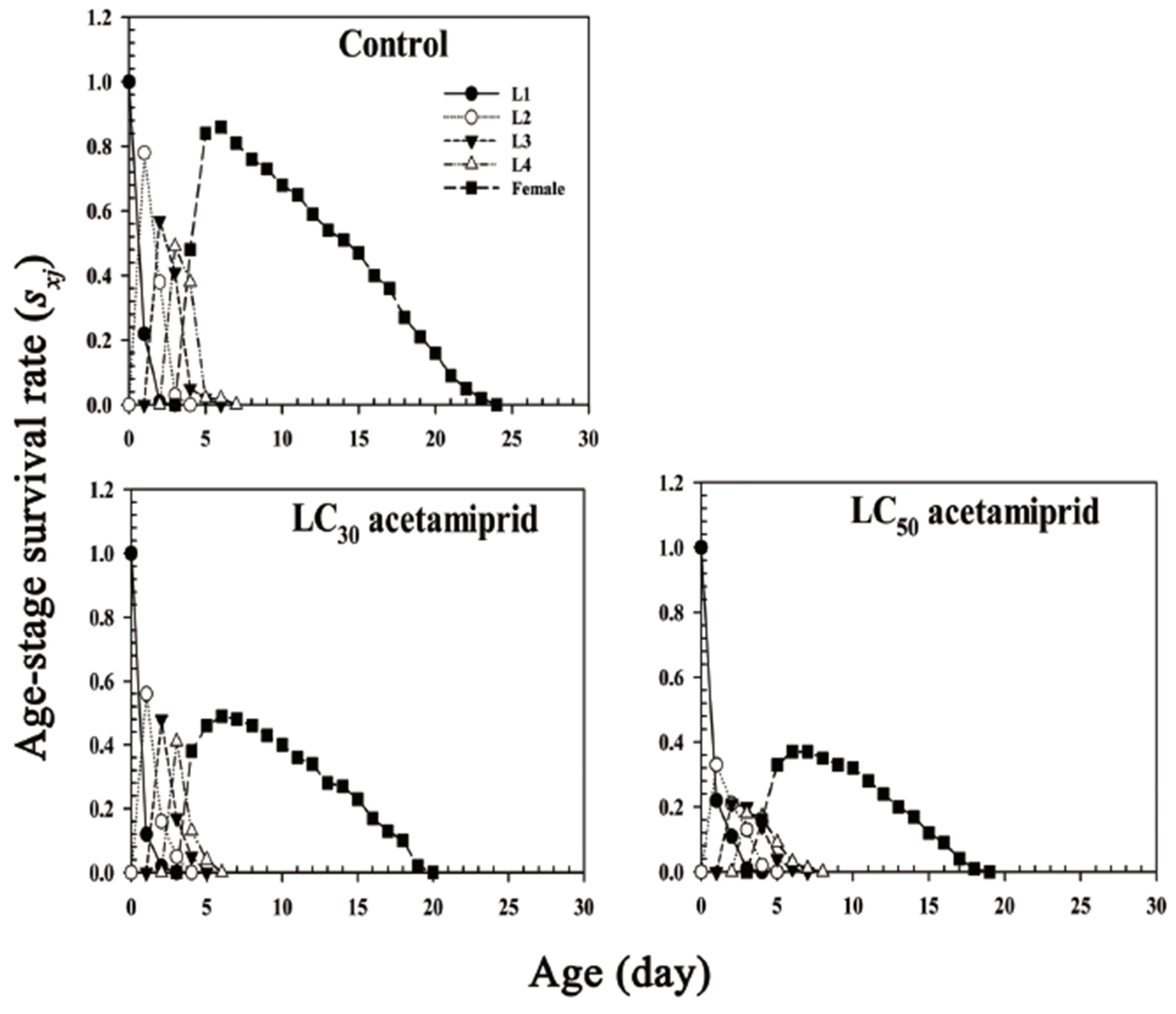
3.2. Life–History Traits
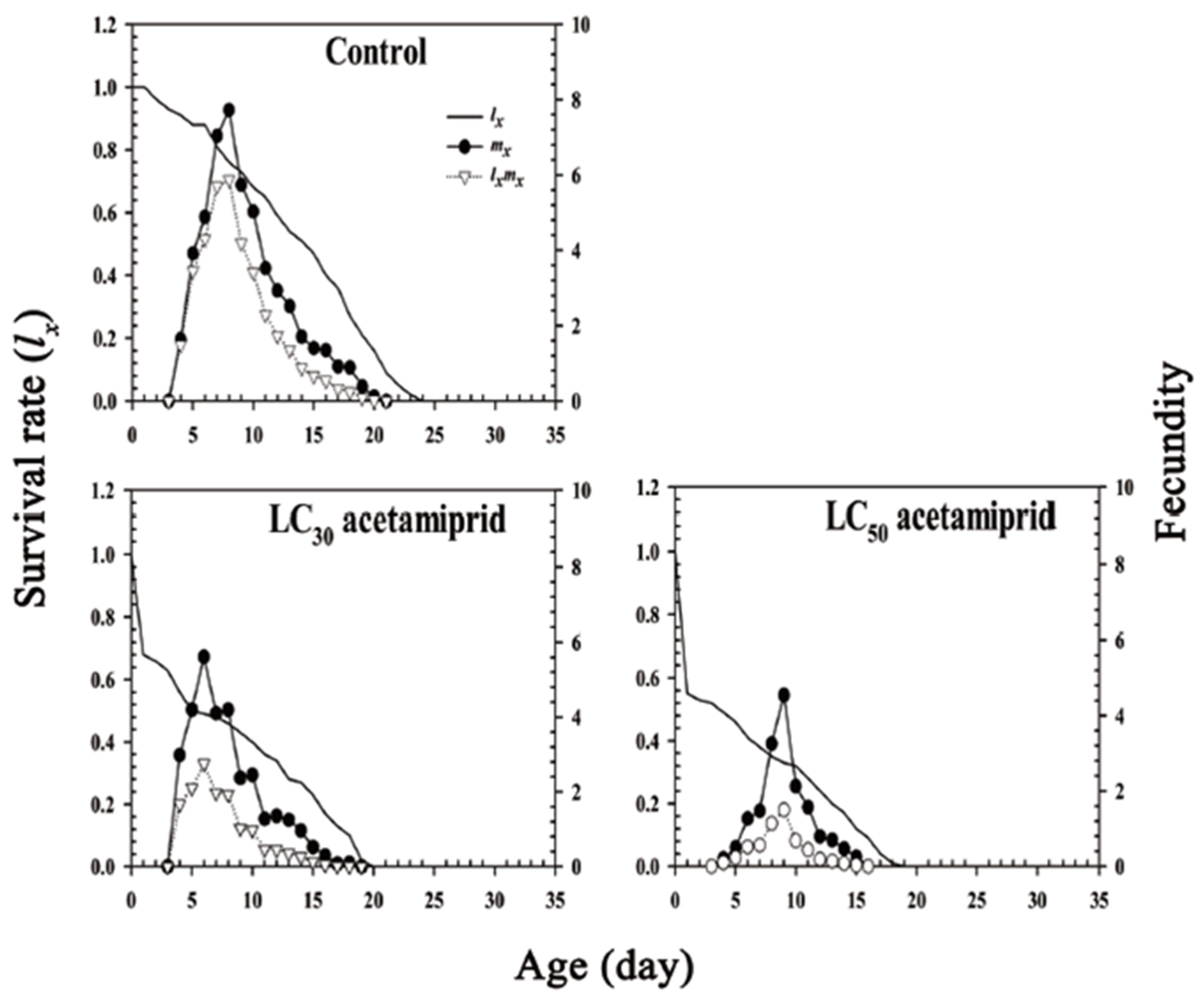
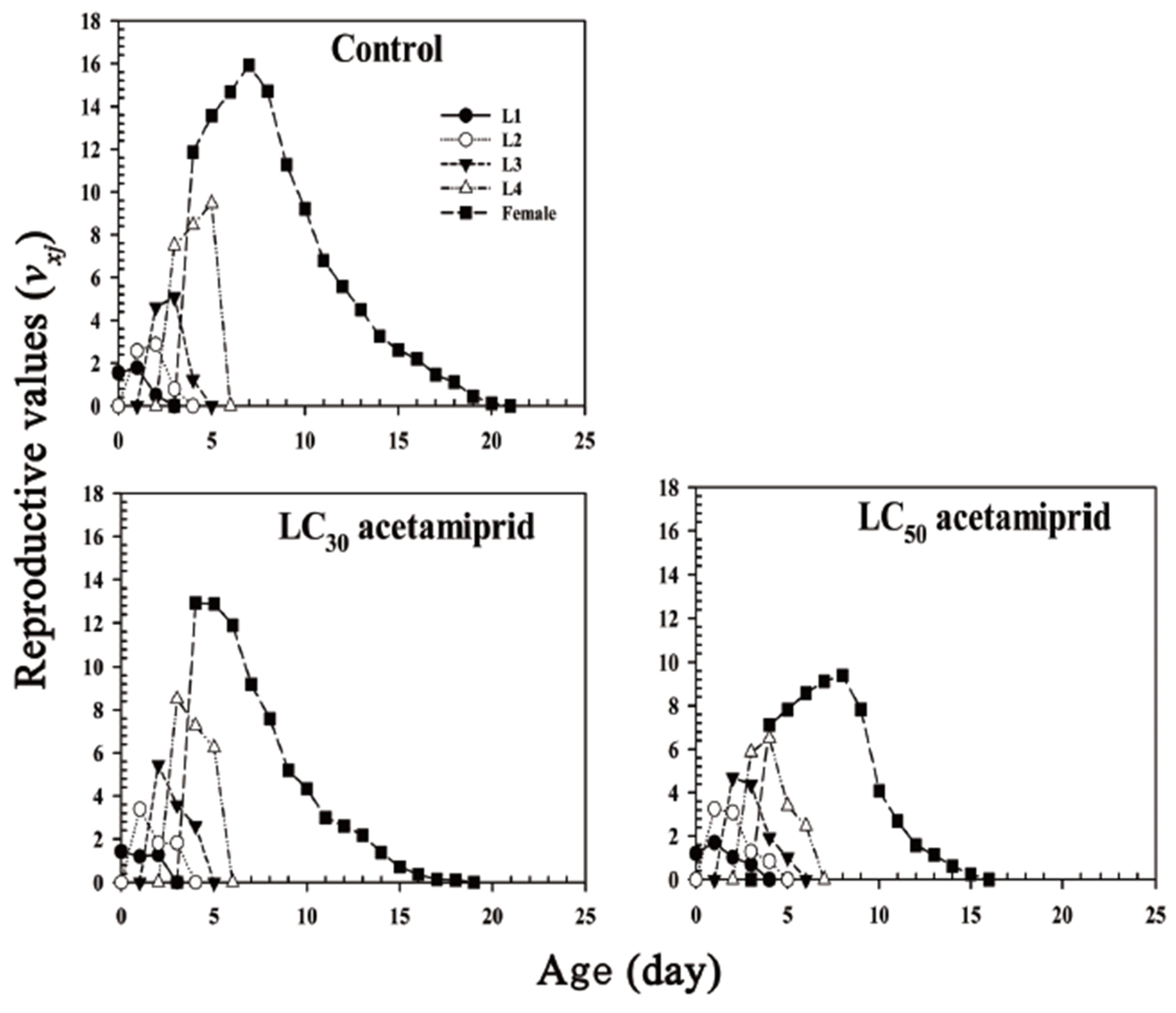
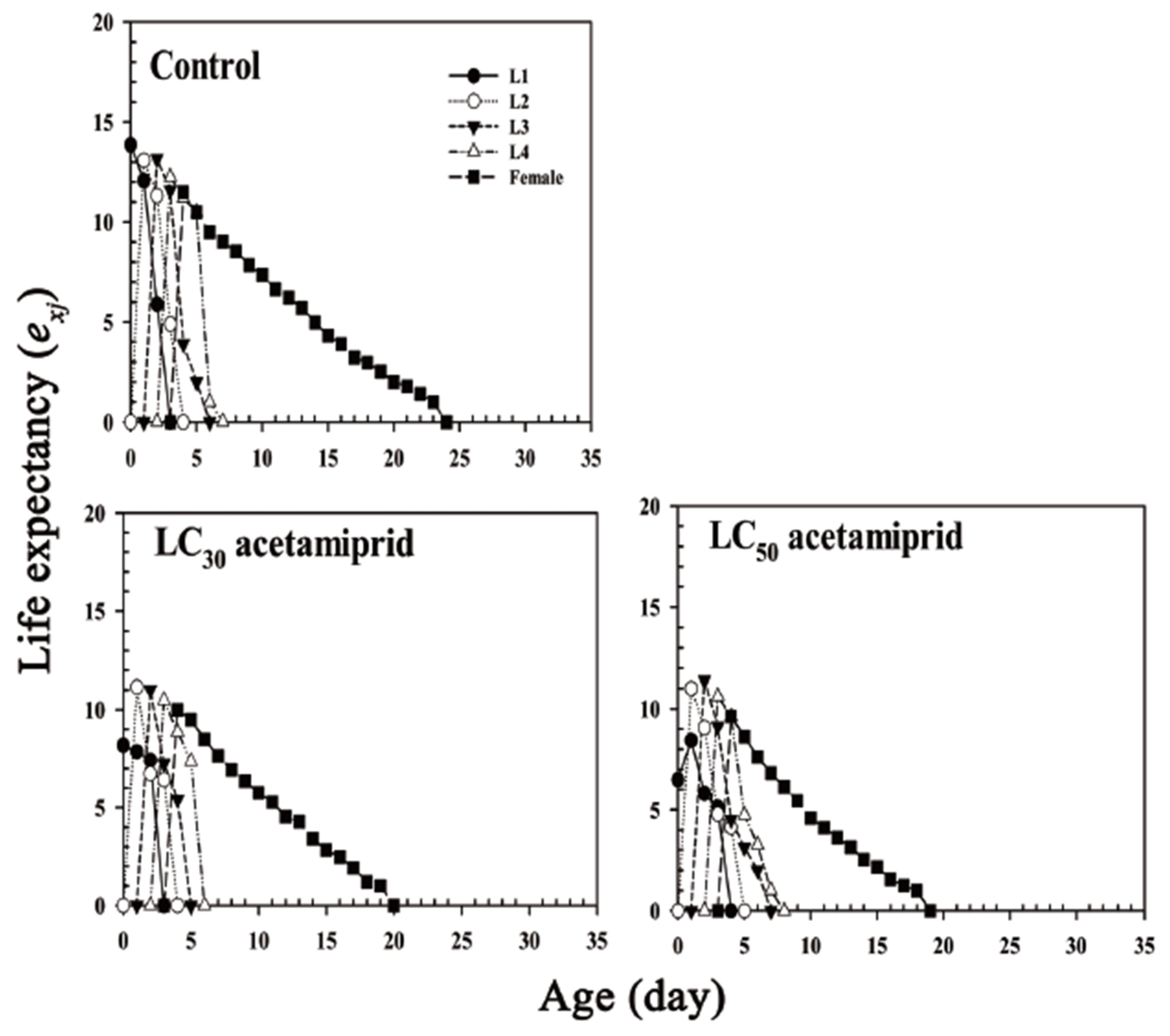
3.3. Life Table and Fertility Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hopper, K.R.; Lanier, K.; Rhoades, J.H.; Hoelmer, K.A.; Meikle, W.G.; Heimpel, G.E.; O’Neil, R.J.; Voegtlin, D.G.; Woolley, J.B. Host specificity of Aphelinus species collected from soybean aphid in Asia. Biol. Control 2017, 115, 55–73. [Google Scholar] [CrossRef]
- Ragsdale, D.W.; Voegtlin, D.J.; O’Neil, R.J. Soybean aphid biology in North America. Ann. Entomol. Soc. Am. 2004, 97, 204–208. [Google Scholar] [CrossRef]
- Ragsdale, D.W.; Landis, D.A.; Brodeur, J.; Heimpel, G.E.; Desneux, N. Ecology and management of the soybean aphid in North America. Annu. Rev. Entomol. 2011, 56, 375–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mokbel, E. Resistance risk assessment: Realized heritability, cross resistance and resistance stability of acetamiprid in the cotton aphid, Aphis gossypii Glover (Homoptera: Aphididae). J. Plant Prot. Res. 2019, 58, 328–334. [Google Scholar]
- Aseperi, A.K.; Busquets, R.; Hooda, P.S.; Cheung, P.C.W.; Barker, J. Behaviour of neonicotinoids in contrasting soils. J. Environ. Manag. 2020, 276, 111329. [Google Scholar] [CrossRef]
- Lu, Z.; Ye, W.; Feng, P.; Dai, M.; Bian, D.; Ren, Y.; Zhu, Q.; Mao, T.; Su, W.; Li, F.; et al. Low concentration acetamiprid-induced oxidative stress hinders the growth and development of silkworm posterior silk glands. Pestic. Biochem. Phys. 2021, 12, 104824. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Fauvergue, X.; Dechaume-Moncharmont, F.X.; Kerhoas, L.; Ballanger, Y.; Kaiser, L. Diaeretiella rapae limits Myzus persicae populations after applications of deltamethrin in oilseed rape. J. Econ. Entomol. 2005, 98, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Fenner, K.; Canonica, S.; Wackett, L.P.; Elsner, M. Evaluating pesticide degradation in the environment: Blind spots and emerging opportunities. Science 2013, 341, 752–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, F.; Gul, H.; Tariq, K.; Desneux, N.; Gao, X.; Song, D. Functional analysis of cytochrome P450 genes linked with acetamiprid resistance in melon aphid, Aphis gossypii. Pestic. Biochem. Phys. 2020, 175, 104687. [Google Scholar] [CrossRef] [PubMed]
- Mahmoodi, L.; Mehrkhou, F.; Guz, N.; Forouzan, M.; Atlihan, R. Sublethal effects of three insecticides on fitness parameters and population projection of Brevicoryne brassicae (Hemiptera: Aphididae). J. Econ. Entomol. 2020, 113, 2713–2722. [Google Scholar] [CrossRef]
- Ullah, F.; Gul, H.; Tariq, K.; Desneux, N.; Gao, X.; Song, D. Acetamiprid resistance and fitness costs of melon aphid, Aphis gossypii: An age-stage, two-sex life table study. Pestic. Biochem. Phys. 2020, 171, 104729. [Google Scholar] [CrossRef]
- Cui, L.; Yuan, H.; Wang, Q.; Wang, Q.; Rui, C. Sublethal effects of the novel cisnitromethylene neonicotinoid cycloxaprid on the cotton aphid Aphis gossypii glover (Hemiptera: Aphididae). Sci. Rep. 2018, 8, 8915. [Google Scholar] [CrossRef]
- Ullah, F.; Gul, H.; Tariq, K.; Desneux, N.; Gao, X.; Song, D. Thiamethoxam induces transgenerational hormesis effects and alteration of genes expression in Aphis gossypii. Pestic. Biochem. Phys. 2020, 165, 104557. [Google Scholar] [CrossRef] [PubMed]
- Ullah, F.; Gul, H.; Desneux, N.; Qu, Y.; Xiao, X.; Khattak, A.M.; Gao, X.; Song, D. Acetamiprid-induced hormetic effects and vitellogenin gene (Vg) expression in the melon aphid, Aphis gossypii. Entomol. Gen. 2019, 39, 259–270. [Google Scholar] [CrossRef]
- Hanson, A.A.; Menger-Anderson, J.; Silverstein, C.; Potter, B.D.; MacRae, I.V.; Hodgson, E.W.; Koch, R.L. Evidence for soybean aphid (Hemiptera: Aphididae) resistance to pyrethroid insecticides in the upper Midwestern United States. J. Econ. Entomol. 2017, 110, 2235–2246. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, Q.; Ding, J.; Wang, Y.; Zhang, Z.; Liu, F.; Mu, W. Sublethal effects of chlorfenapyr on the life table parameters, nutritional physiology and enzymatic properties of Bradysia odoriphaga (Diptera: Sciaridae). Pestic. Biochem. Phys. 2018, 148, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Somar, R.O.; Zamani, A.A.; Alizadeh, M. Joint action toxicity of imidacloprid and pymetrozine on the melon aphid, Aphis gossypii. Crop Prot. 2019, 124, 104850. [Google Scholar] [CrossRef]
- Jan, M.T.; Abbas, N.; Shad, S.A.; Saleem, M.A. Resistance to organophosphate, pyrethroid and biorational insecticides in populations of spotted bollworm, Earias vittella (Fabricius) (Lepidoptera: Noctuidae), in Pakistan. Crop Prot. 2015, 78, 247–252. [Google Scholar] [CrossRef]
- Paula, D.P.; Menger, J.; Andow, D.A.; Koch, R.L. Diverse patterns of constitutive and inducible overexpression of detoxifying enzyme genes among resistant Aphis glycines populations. Pestic. Biochem. Phys. 2020, 164, 100–114. [Google Scholar] [CrossRef]
- Chi, H.; You, M.; Atlihan, R.; Smith, C.L.; Kavousi, A.; Ozgokce, M.S.; Guncan, A.; Tuan, S.J.; Fu, J.W.; Xu, Y.Y.; et al. Age- Stage, two-sex life table: An introduction to theory, data analysis, and application. Entomol. Gen. 2020, 40, 102–123. [Google Scholar] [CrossRef]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sexes and variable development rate among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- James, M.; Patrick, B.; Anitha, C.; Cole, D.; John, G.; Phillip, G.; Krista, H.; Hodgson, E.W.; Knodel, J.J.; Macrae, I.V. Implementation of a diagnostic-concentration bioassay for detection of susceptibility to pyrethroids in soybean aphid (Hemiptera: Aphididae). J. Econ. Entomol. 2020, 113, 932–939. [Google Scholar]
- Han, W.; Yang, Y.; Gao, J.; Zhao, D.; Ren, C.; Wang, S.; Zhao, S.; Zhong, Y. Chronic toxicity and biochemical response of Apis cerana cerana (Hymenoptera: Apidae) exposed to acetamiprid and propiconazole alone or combined. Ecotoxicology 2019, 28, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Goodman, D. Optimal life histories, optimal notation, and the value of reproductive value. Am. Nat. 1982, 119, 803–823. [Google Scholar] [CrossRef]
- Chi, H.; Su, H.Y. Age-stage, two-sex life tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzus persicae (Sulzer) (Homoptera: Aphididae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environ. Entomol. 2006, 35, 10–21. [Google Scholar] [CrossRef]
- Tuan, S.J.; Li, N.J.; Yeh, C.C.; Tang, L.C.; Chi, H. Effects of green manure cover crops on Spodoptera litura (Lepidoptera: Noctuidae) populations. J. Econ. Entomol. 2014, 107, 897–905. [Google Scholar] [CrossRef] [Green Version]
- Chi, H. TWOSEX-MSChart: A Computer Program for Age Stage, Two-Sex Life Table Analysis; National Chung Hsing University: Taichung, Taiwan, 2020; Available online: http://140.120.197.173/Ecology/Download/TWOSEX-MSChart.rar (accessed on 16 March 2021).
- Guedes, R.N.C.; Smagghe, G.; Stark, J.D.; Desneux, N. Pesticide induced stress in arthropod pests for optimized integrated pest management programs. Annu. Rev. Entomol. 2016, 61, 43–62. [Google Scholar] [CrossRef] [Green Version]
- Sial, M.U.; Zhao, Z.; Zhang, L.; Zhang, Y.; Mao, L.; Jiang, H. Evaluation of Insecticides induced hormesis on the demographic parameters of Myzus persicae and expression changes of metabolic resistance detoxification genes. Sci. Rep. 2018, 8, 16601. [Google Scholar] [CrossRef]
- Rix, R.R.; Ayyanath, M.M.; Cutler, G.C. Sublethal concentrations of imidacloprid increase reproduction, alter expression of detoxification genes, and prime Myzus persicae for subsequent stress. J. Pest Sci. 2016, 89, 581–589. [Google Scholar] [CrossRef]
- Tang, Q.; Ma, K.; Chi, H.; Hou, Y.; Gao, X. Transgenerational hormetic effects of sublethal dose of flupyradifurone on the green peach aphid, Myzus persicae (Sulzer) (Hemiptera: Aphididae). PLoS ONE 2019, 14, e0208058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Yang, X.B.; Yang, H.; Long, G.Y.; Jin, D.C.; Bendena, B. Effects of sublethal concentrations of insecticides on the fecundity of Sogatella furcifera (Hemiptera: Delphacidae) via the regulation of vitellogenin and its receptor. J. Insect Sci. 2020, 20, 14. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.L.; Dai, R.H.; Yang, H.; Jin, D.C. Sublethal Effects of triazophos on the life table parameters of Sogatella furcifera (Hemiptera: Delphacidae). Fla. Entomol. 2016, 99, 292–296. [Google Scholar] [CrossRef]
- Ayyanath, M.M.; Cutler, G.C.; Scott-Dupree, C.D.; Sibley, P.K.; Guy, S. Transgenerational shifts in reproduction hormesis in green peach aphid exposed to low concentrations of imidacloprid. PLoS ONE 2013, 8, e74532. [Google Scholar]
- Wang, S.; Qi, Y.; Desneux, N.; Shi, X.; Biondi, A.; Gao, X. Sublethal and transgenerational effects of short-term and chronic exposures to the neonicotinoid nitenpyram on the cotton aphid Aphis gossypii. J. Pest Sci. 2017, 90, 389–396. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insecticide | LC50 (mg a.i./L) | 95% CI | LC30 (mg a.i./L) | 95% CI | Slope ± SE † | χ2 (df) |
|---|---|---|---|---|---|---|
| Acetamiprid | 6.742 | 5.133–8.629 | 3.968 | 2.676–5.203 | 2.278 ± 0.337 | 0.157 (5) |
| Stages | Control | LC30 | LC50 |
|---|---|---|---|
| Mean ± SE † | Mean ± SE | Mean ± SE | |
| L1 development time (d) | 1.23 ± 0.04 b | 1.21 ± 0.06 b | 1.62 ± 0.11 a |
| L2 development time (d) | 1.2 ± 0.04 a | 1.14 ± 0.05 a | 1.28 ± 0.06 a |
| L3 development time (d) | 1.13 ± 0.05 a | 1.12 ± 0.04 a | 1.22 ± 0.08 a |
| L4 development time (d) | 1.01 ± 0.01 a | 1.04 ± 0.03 a | 1.08 ± 0.04 a |
| Mean longevity of female adult (d) | 11.02 ± 0.55 a | 9.8 ± 0.59 ab | 8.89 ± 0.53 b |
| APOP (d) | 0.20 ± 0.04 b | 0.02 ± 0.02 c | 1.00 ± 0.16 a |
| TPOP (d) | 4.66 ± 0.07 b | 4.31 ± 0.08 c | 5.74 ± 0.16 a |
| Fecundity | 42.49 ± 1.83 a | 27.73 ± 1.88 b | 14.76 ± 1.21 c |
| Population Parameters | Control | LC30 | LC50 |
|---|---|---|---|
| Mean ± SE † | Mean ± SE | Mean ± SE | |
| Intrinsic rate of increase (r) (d−1) | 0.4387 ± 0.0080 a | 0.3580 ± 0.0171 b | 0.1859 ± 0.0171 c |
| Finite rate of increase (λ) (d−1) | 1.5507 ± 0.0124 a | 1.4304 ± 0.0244 b | 1.2043 ± 0.0205 c |
| Net reproductive rate (R0) | 36.54 ± 2.156 a | 14.14 ± 1.684 b | 5.61 ± 0.847 c |
| Mean generation time (T) (d) | 8.20 ± 0.076 b | 7.4 ± 0.106 c | 9.28 ± 0.111 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, A.; Xu, L.; Liu, Z.; Zhang, J.; Zhao, K.; Han, L. Effects of Acetamiprid at Low and Median Lethal Concentrations on the Development and Reproduction of the Soybean Aphid Aphis glycines. Insects 2022, 13, 87. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010087
Zhang A, Xu L, Liu Z, Zhang J, Zhao K, Han L. Effects of Acetamiprid at Low and Median Lethal Concentrations on the Development and Reproduction of the Soybean Aphid Aphis glycines. Insects. 2022; 13(1):87. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010087
Chicago/Turabian StyleZhang, Aonan, Ling Xu, Ziqi Liu, Jiabo Zhang, Kuijun Zhao, and Lanlan Han. 2022. "Effects of Acetamiprid at Low and Median Lethal Concentrations on the Development and Reproduction of the Soybean Aphid Aphis glycines" Insects 13, no. 1: 87. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010087