
Homeostasis of Mitochondrial Ca2+ Stores Is Critical for Signal Amplification in Drosophila melanogaster Olfactory Sensory Neurons


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Fly Rearing and Antennal Preparation
2.3. Calcium Imaging
2.4. Statistical Analysis
3. Results
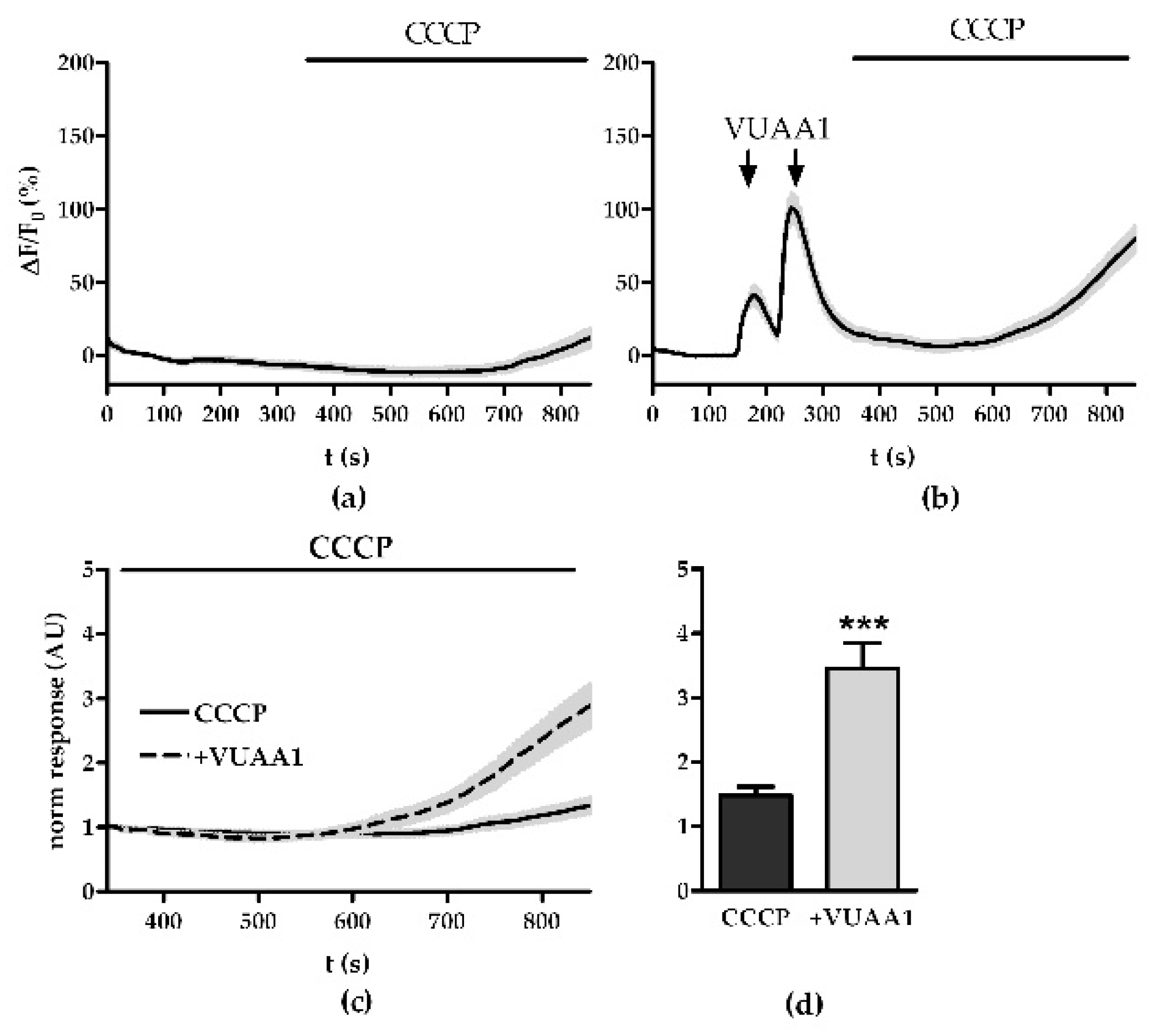
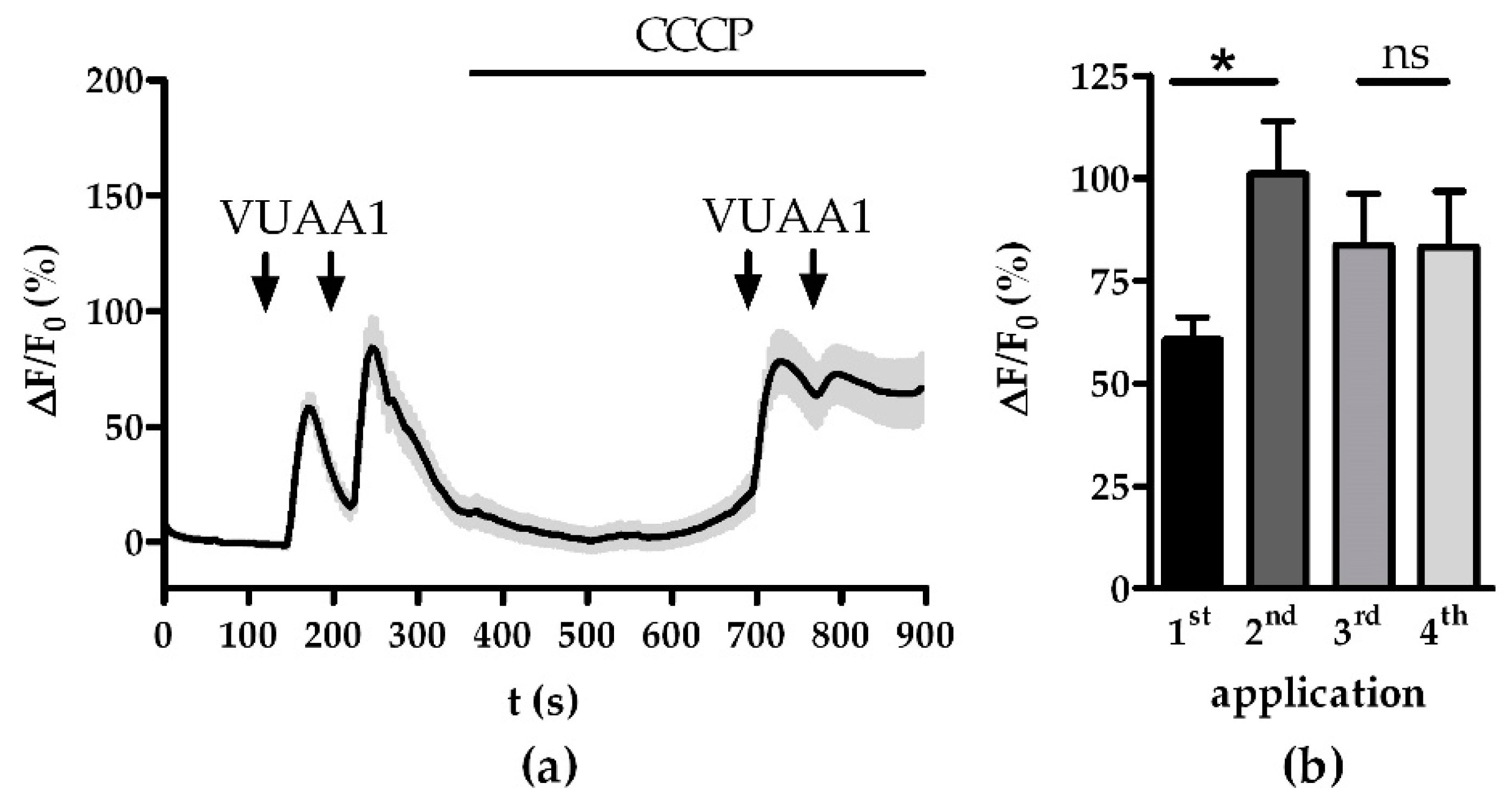
3.1. Modulation of Ca2+ Homeostasis
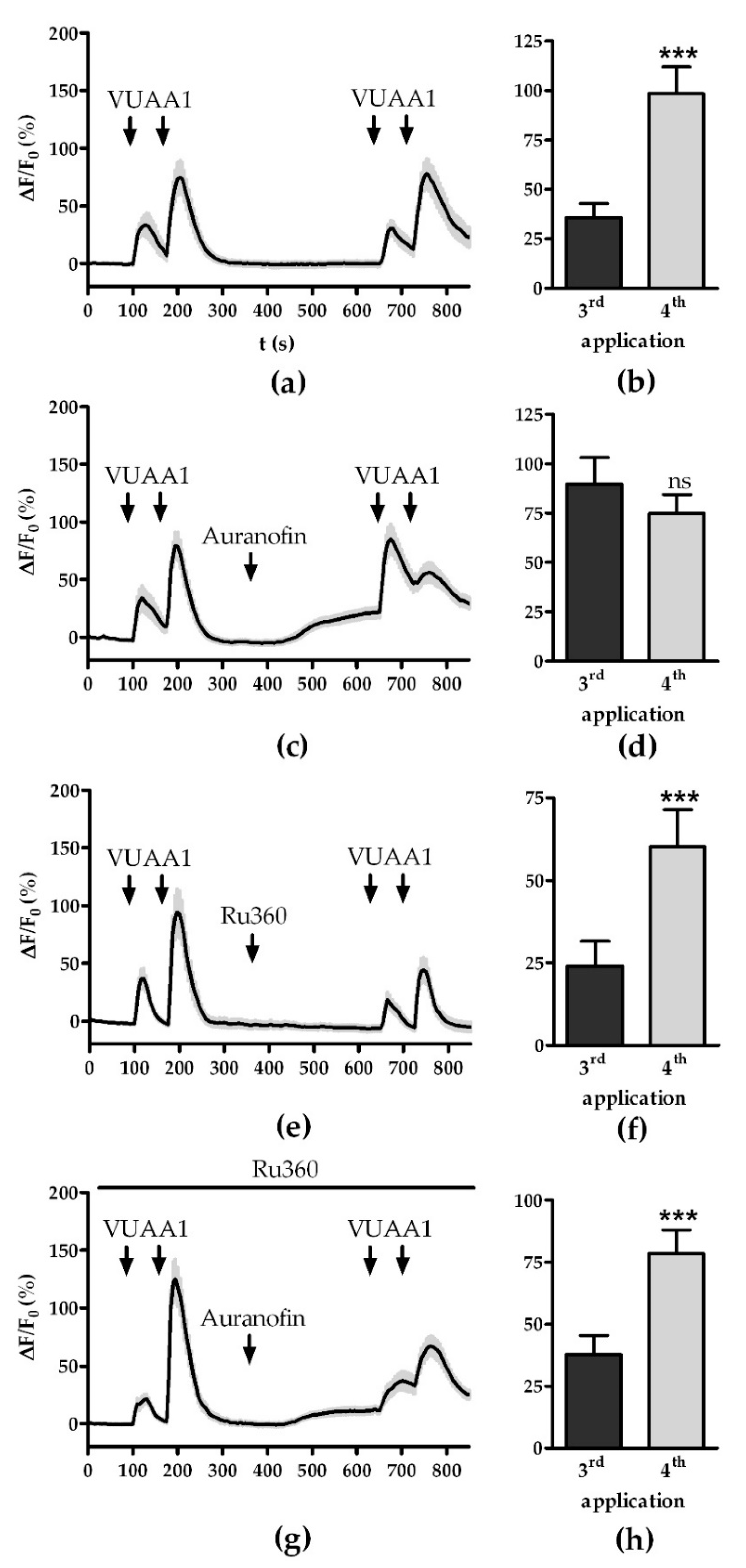
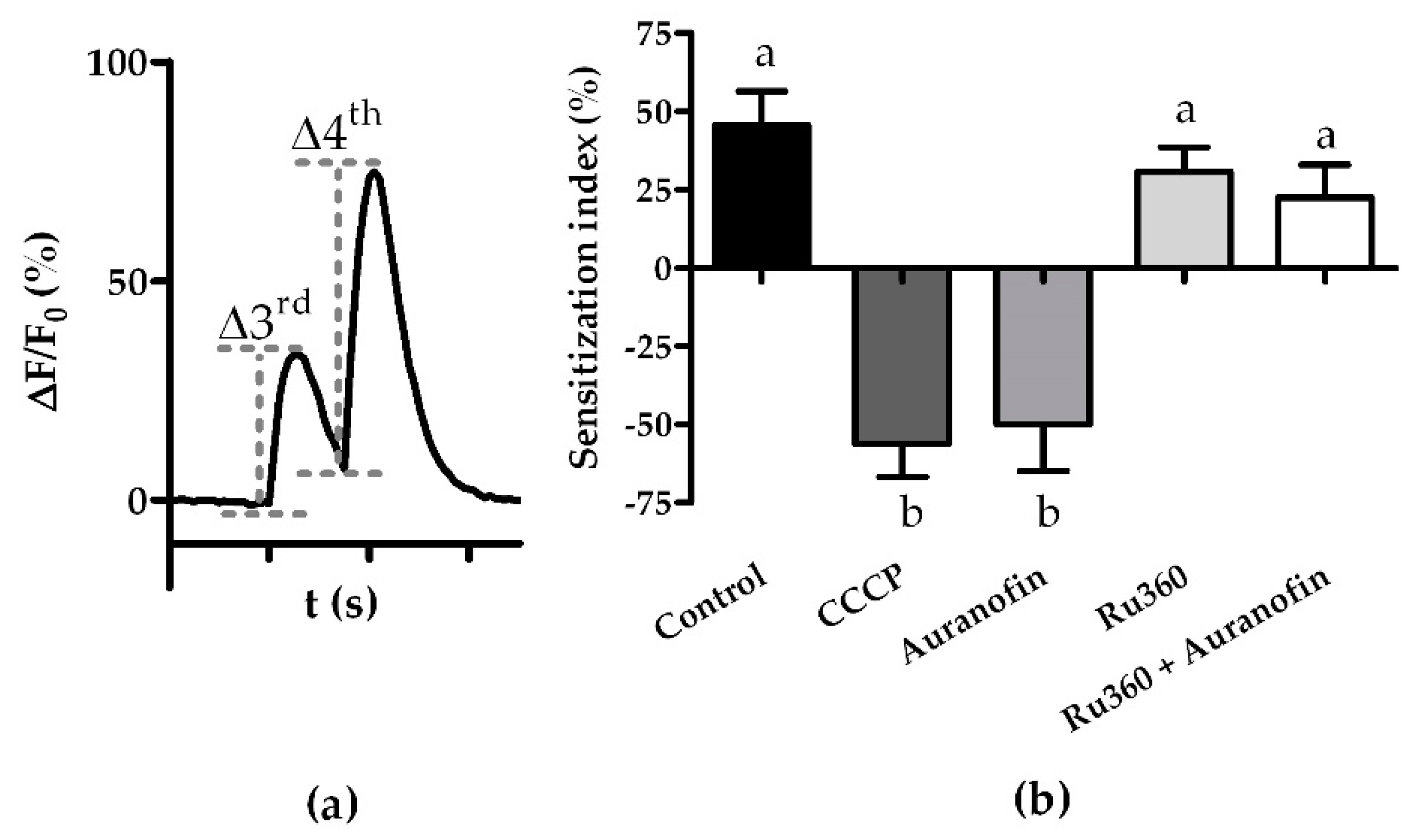
3.2. Modulation of the Mitochondrial Ca2+ Export and Import
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hansson, B.S.; Stensmyr, M.C. Evolution of Insect Olfaction. Neuron 2011, 72, 698–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wicher, D. Tuning Insect Odorant Receptors. Front. Cell. Neurosci. 2018, 12, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Getahun, M.N.; Thoma, M.; Lavista-Llanos, S.; Keesey, I.; Fandino, R.A.; Knaden, M.; Wicher, D.; Olsson, S.B.; Hansson, B.S. Intracellular regulation of the insect chemoreceptor complex impacts odour localization in flying insects. J. Exp. Biol. 2016, 219 Pt 21, 3428–3438. [Google Scholar] [PubMed] [Green Version]
- Missbach, C.; Dweck, H.K.M.; Vogel, H.; Vilcinskas, A.; Stensmyr, M.C.; Hansson, B.S.; Gosse-Wilde, E. Evolution of insect olfactory receptors. eLife 2014, 3, e02115. [Google Scholar] [CrossRef] [PubMed]
- Koehl, M. The Fluid Mechanics of Arthropod Sniffing in Turbulent Odor Plumes. Chem. Sens. 2005, 31, 93–105. [Google Scholar] [CrossRef] [PubMed]
- van Breugel, F.; Dickinson, M.H. Plume-tracking behavior of flying Drosophila emerges from a set of distinct sensory-motor reflexes. Curr. Biol. 2014, 24, 274–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanbhag, S.R.; Muller, B.; Steinbrecht, R.A. Atlas of olfactory organs of Drosophila melanogaster 2. Internal organization and cellular architecture of olfactory sensilla. Arthropod Struct. Dev. 2000, 29, 211–229. [Google Scholar] [CrossRef]
- Butterwick, J.A.; del Marmol, J.; Kim, K.H.; Kahlson, M.; Rogow, J.; Walz, T.; Ruta, V. Cryo-EM structure of the insect olfactory receptor Orco. Nature 2018, 560, 447–452. [Google Scholar] [CrossRef]
- Larsson, M.C.; Domingos, A.; Jones, W.; Chiappe, M.E.; Amrein, H.; Vosshall, L.B. Or83b Encodes a Broadly Expressed Odorant Receptor Essential for Drosophila Olfaction. Neuron 2004, 43, 703–714. [Google Scholar] [CrossRef] [Green Version]
- Benton, R.; Sachse, S.; Michnick, S.W.; Vosshall, L.B. Atypical Membrane Topology and Heteromeric Function of Drosophila Odorant Receptors In Vivo. PLoS Biol. 2006, 4, e20. [Google Scholar] [CrossRef] [Green Version]
- Wicher, D.; Schäfer, R.; Bauernfeind, R.; Stensmyr, M.; Heller, R.; Heinemann, S.H.; Hansson, B.S. Drosophila odorant receptors are both ligand-gated and cyclic-nucleotide-activated cation channels. Nature 2008, 452, 1007–1011. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Pellegrino, M.; Nakagawa, T.; Nakagawa, T.; Vosshall, L.B.; Touhara, K. Insect olfactory receptors are heteromeric ligand-gated ion channels. Nature 2008, 452, 1002–1006. [Google Scholar] [CrossRef] [PubMed]
- Endo, M. Calcium Ion as a Second Messenger with Special Reference to Excitation-Contraction Coupling. J. Pharmacol. Sci. 2006, 100, 519–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morad, M.; Soldatov, N. Calcium channel inactivation: Possible role in signal transduction and Ca2+ signaling. Cell Calcium 2005, 38, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Mukunda, L.; Miazzi, F.; Kaltofen, S.; Hansson, B.S.; Wicher, D. Calmodulin modulates insect odorant receptor function. Cell Calcium 2014, 55, 191–199. [Google Scholar] [CrossRef]
- Cao, L.-H.; Jing, B.-Y.; Yang, D.; Zeng, X. (Kevin); Shen, Y.; Tu, Y.; Luo, D.-G. Distinct signaling of Drosophila chemoreceptors in olfactory sensory neurons. Proc. Natl. Acad. Sci. USA 2016, 113, E902–E911. [Google Scholar] [CrossRef] [Green Version]
- Murmu, M.S.; Stinnakre, J.; Réal, E.; Martin, J.-R. Calcium-stores mediate adaptation in axon terminals of Olfactory Receptor Neurons in Drosophila. BMC Neurosci. 2011, 12, 105. [Google Scholar] [CrossRef] [Green Version]
- Murmu, M.S.; Martin, J.-R. Interaction between cAMP and intracellular Ca 2+ -signaling pathways during odor-perception and adaptation in Drosophila. Biochim. Biophys. Acta 2016, 1863, 2156–2174. [Google Scholar] [CrossRef]
- Guerini, D.; Coletto, L.; Carafoli, E. Exporting calcium from cells. Cell Calcium 2005, 38, 281–289. [Google Scholar] [CrossRef]
- Halty-deLeon, L.; Hansson, B.S.; Wicher, D. The Drosophila melanogaster Na+/Ca2+ Exchanger CALX Controls the Ca2+ Level in Olfactory Sensory Neurons at Rest and After Odorant Receptor Activation. Front. Cell. Neurosci. 2018, 12, 186. [Google Scholar] [CrossRef] [Green Version]
- De Stefani, D.; Rizzuto, R.; Pozzan, T. Enjoy the Trip: Calcium in Mitochondria Back and Forth. Annu. Rev. Biochem. 2016, 85, 161–192. [Google Scholar] [CrossRef]
- Lucke, J.; Kaltofen, S.; Hansson, B.S.; Wicher, D. The role of mitochondria in shaping odor responses in Drosophila melanogaster olfactory sensory neurons. Cell Calcium 2020, 87, 102179. [Google Scholar] [CrossRef] [PubMed]
- Herrington, J.; Park, Y.B.; Babcock, D.; Hille, B. Dominant Role of Mitochondria in Clearance of Large Ca2+ Loads from Rat Adrenal Chromaffin Cells. Neuron 1996, 16, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Babcock, D.F.; Hille, B. Mitochondrial oversight of cellular Ca2+ signaling. Curr. Opin. Neurobiol. 1998, 8, 398–404. [Google Scholar] [CrossRef]
- Fluegge, D.; Moeller, L.; Cichy, A.; Gorin, M.; Weth, A.; Veitinger, S.; Cainarca, S.; Lohmer, S.; Corazza, S.; Neuhaus, E.; et al. Mitochondrial Ca2+ mobilization is a key element in olfactory signaling. Nat. Neurosci. 2012, 15, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Ignatious Raja, J.S.; Katanayeva, N.; Katanaev, V.L.; Galizia, C.G. Role of Go/i subgroup of G proteins in olfactory signaling of Drosophila melanogaster. Eur. J. Neurosci. 2014, 39, 1245–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Getahun, M.N.; Olsson, S.B.; Lavista-Llanos, S.; Hansson, B.S.; Wicher, D. Insect Odorant Response Sensitivity Is Tuned by Metabotropically Autoregulated Olfactory Receptors. PLoS ONE 2013, 8, e58889. [Google Scholar] [CrossRef] [Green Version]
- Sargsyan, V.; Getahun, M.N.; Llanos, S.L.; Olsson, S.B.; Hansson, B.S.; Wicher, D. Phosphorylation via PKC Regulates the Function of the Drosophila Odorant Co-Receptor. Front. Cell. Neurosci. 2011, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Miazzi, F.; Hansson, B.S.; Wicher, D. Odor induced cAMP production in Drosophila melanogaster olfactory sensory neurons. J. Exp. Biol. 2016, 219 Pt 12, 1798–1803. [Google Scholar] [CrossRef] [Green Version]
- Mukunda, L.; Miazzi, F.; Sargsyan, V.; Hansson, B.S.; Wicher, D. Calmodulin Affects Sensitization of Drosophila melanogaster Odorant Receptors. Front. Cell. Neurosci. 2016, 10, 28. [Google Scholar] [CrossRef] [Green Version]
- Prelic, S.; Mahadevan, V.P.; Venkateswaran, V.; Lavista-Llanos, S.; Hansson, B.S.; Wicher, D. Functional Interaction Between Drosophila Olfactory Sensory Neurons and Their Support Cells. Front. Cell. Neurosci. 2022, 15, 789086. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.B. GAL4 system in Drosophila: A fly geneticist’s Swiss army knife. Genesis 2002, 34, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.-W.; Wardill, T.J.; Sun, Y.; Pulver, S.R.; Renninger, S.L.; Baohan, A.; Schreiter, E.R.; Kerr, R.A.; Orger, M.B.; Jayaraman, V.; et al. Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature 2013, 499, 295–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardi, P. Mitochondrial Transport of Cations: Channels, Exchangers, and Permeability Transition. Physiol. Rev. 1999, 79, 1127–1155. [Google Scholar] [CrossRef] [PubMed]
- Oxenoid, K.; Dong, Y.; Cao, C.; Cui, T.; Sancak, Y.; Markhard, A.L.; Grabarek, Z.; Kong, L.; Liu, Z.; Ouyang, B.; et al. Architecture of the mitochondrial calcium uniporter. Nature 2016, 533, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Ying, W.L.; Emerson, J.; Clarke, M.J.; Sanadi, D.R. Inhibition of mitochondrial calcium ion transport by an oxo-bridged dinuclear ruthenium ammine complex. Biochemistry 1991, 30, 4949–4952. [Google Scholar] [CrossRef]
- Seki, Y.; Dweck, H.K.M.; Rybak, J.; Wicher, D.; Sachse, S.; Hansson, B.S. Olfactory coding from the periphery to higher brain centers in the Drosophila brain. BMC Biol. 2017, 15, 1–20. [Google Scholar] [CrossRef]
- Knaden, M.; Strutz, A.; Ahsan, J.; Sachse, S.; Hansson, B.S. Spatial Representation of Odorant Valence in an Insect Brain. Cell Rep. 2012, 1, 392–399. [Google Scholar] [CrossRef] [Green Version]
- Ruta, V.; Datta, S.R.; Vasconcelos, M.L.; Freeland, J.; Looger, L.L.; Axel, R. A dimorphic pheromone circuit in Drosophila from sensory input to descending output. Nature 2010, 468, 686–690. [Google Scholar] [CrossRef]
- Stensmyr, M.C.; Dweck, H.K.; Farhan, A.; Ibba, I.; Strutz, A.; Mukunda, L.; Linz, J.; Grabe, V.; Steck, K.; Lavista-Llanos, S.; et al. A Conserved Dedicated Olfactory Circuit for Detecting Harmful Microbes in Drosophila. Cell 2012, 151, 1345–1357. [Google Scholar] [CrossRef] [Green Version]
- Wicher, D.; Miazzi, F. Functional properties of insect olfactory receptors: Ionotropic receptors and odorant receptors. Cell Tissue Res. 2021, 383, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Jain, K.; Lavista-Llanos, S.; Grabe, V.; Hansson, B.S.; Wicher, D. Calmodulin regulates the olfactory performance in Drosophila melanogaster. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, C.N.; McKaughan, Q.; Bushong, E.A.; Cauwenberghs, K.; Ng, R.; Madany, M.; Ellisman, M.H.; Su, C.-Y. Systematic morphological and morphometric analysis of identified olfactory receptor neurons in Drosophila melanogaster. eLife 2021, 10, 10. [Google Scholar] [CrossRef]
- Csordás, G.; Weaver, D.; Hajnóczky, G. Endoplasmic Reticulum–Mitochondrial Contactology: Structure and Signaling Functions. Trends Cell Biol. 2018, 28, 523–540. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiesel, E.; Kaltofen, S.; Hansson, B.S.; Wicher, D. Homeostasis of Mitochondrial Ca2+ Stores Is Critical for Signal Amplification in Drosophila melanogaster Olfactory Sensory Neurons. Insects 2022, 13, 270. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13030270
Wiesel E, Kaltofen S, Hansson BS, Wicher D. Homeostasis of Mitochondrial Ca2+ Stores Is Critical for Signal Amplification in Drosophila melanogaster Olfactory Sensory Neurons. Insects. 2022; 13(3):270. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13030270
Chicago/Turabian StyleWiesel, Eric, Sabine Kaltofen, Bill S. Hansson, and Dieter Wicher. 2022. "Homeostasis of Mitochondrial Ca2+ Stores Is Critical for Signal Amplification in Drosophila melanogaster Olfactory Sensory Neurons" Insects 13, no. 3: 270. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13030270