
Antennal Transcriptome Analysis and Identification of Olfactory Genes in Glenea cantor Fabricius (Cerambycidae: Lamiinae)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Sample Collection
2.2. RNA Sequencing
2.3. Transcriptome Assembly, Annotation of Unigenes, and DGE Analysis
2.4. Identification of Olfactory Genes
2.5. Sequence and Phylogenetic Analysis
2.6. Quantitative Real-Time PCR (qRT-PCR)
3. Results
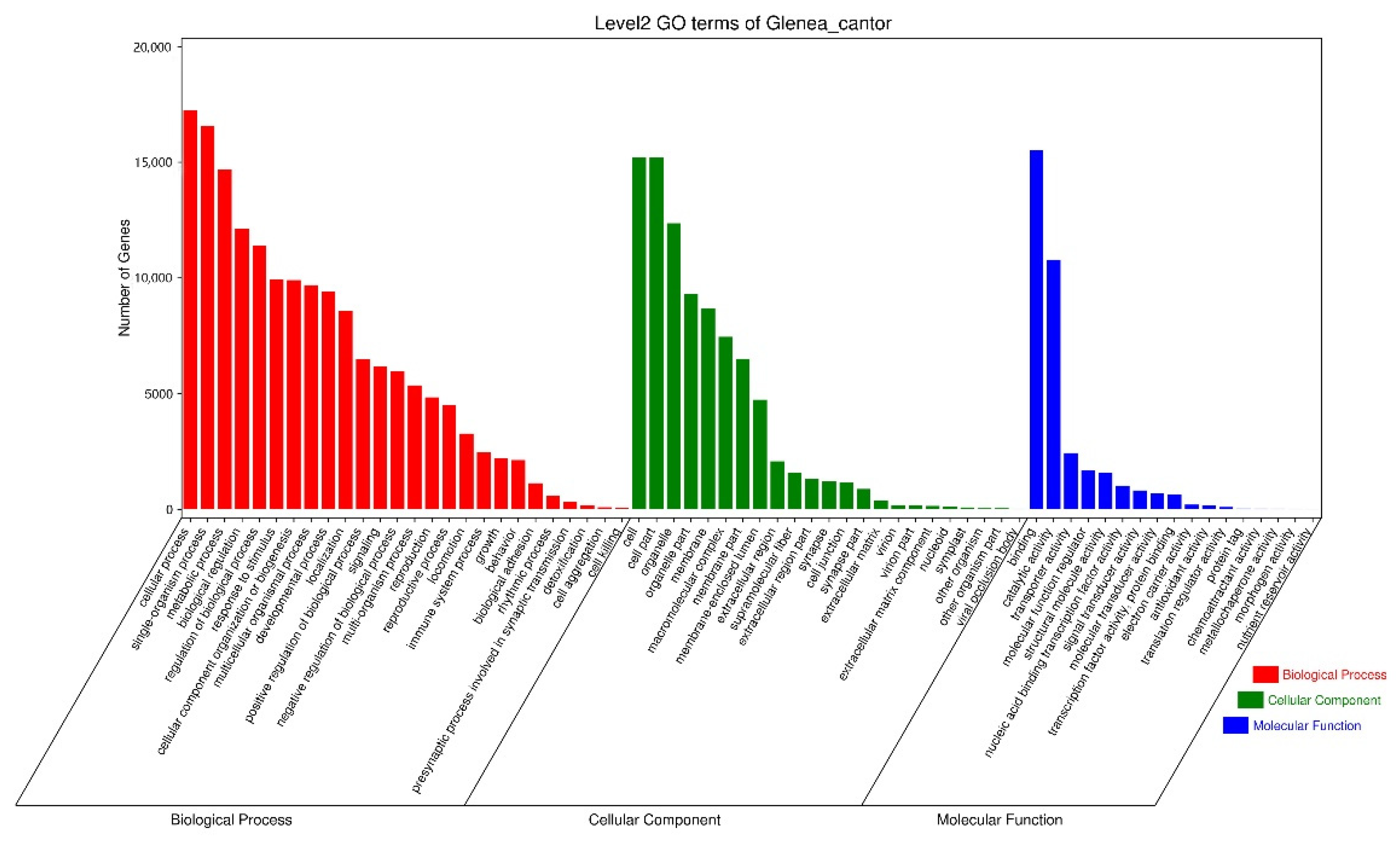
3.1. Overview of G. cantor Antennal Transcriptome
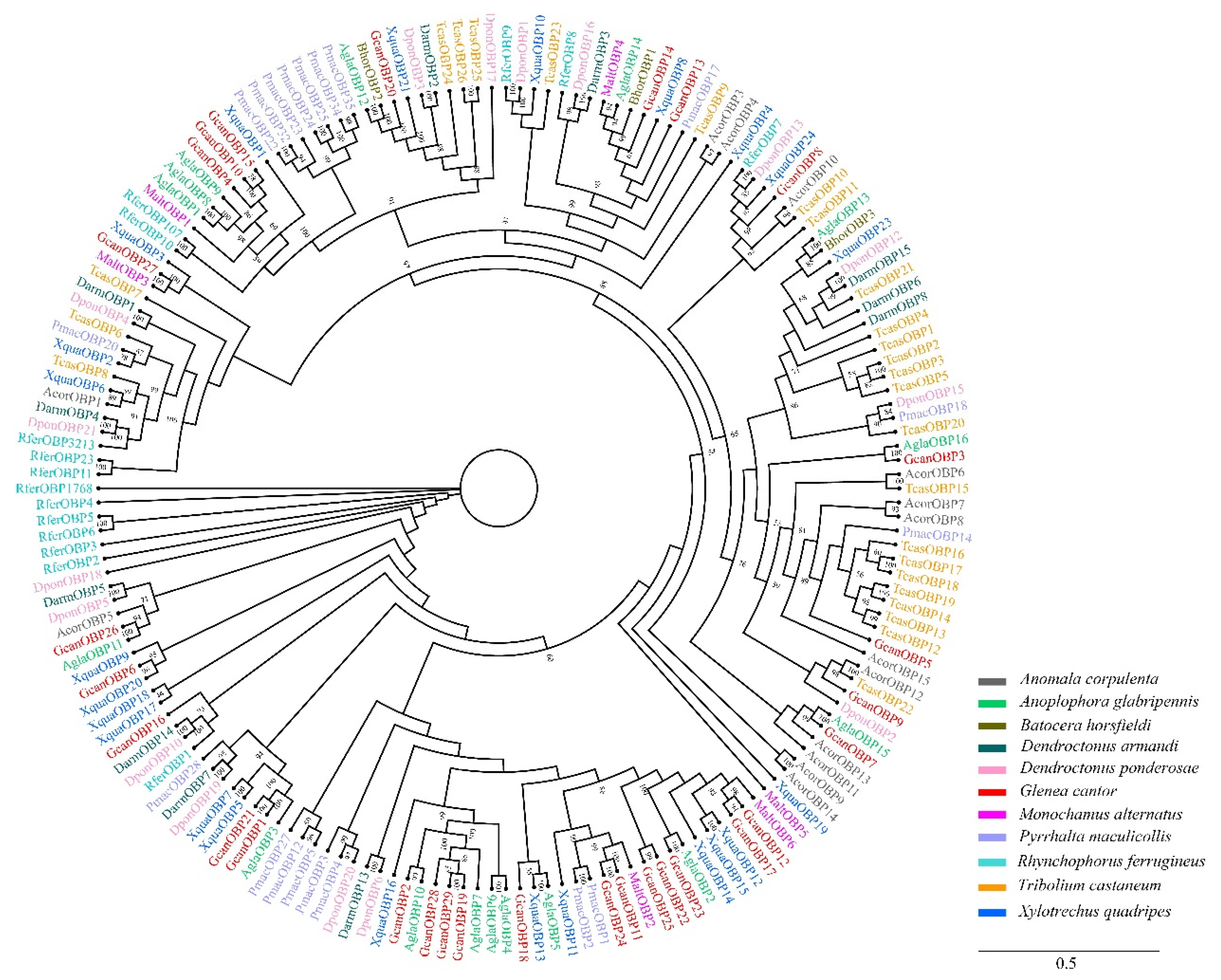
3.2. Identification of Candidate OBPs
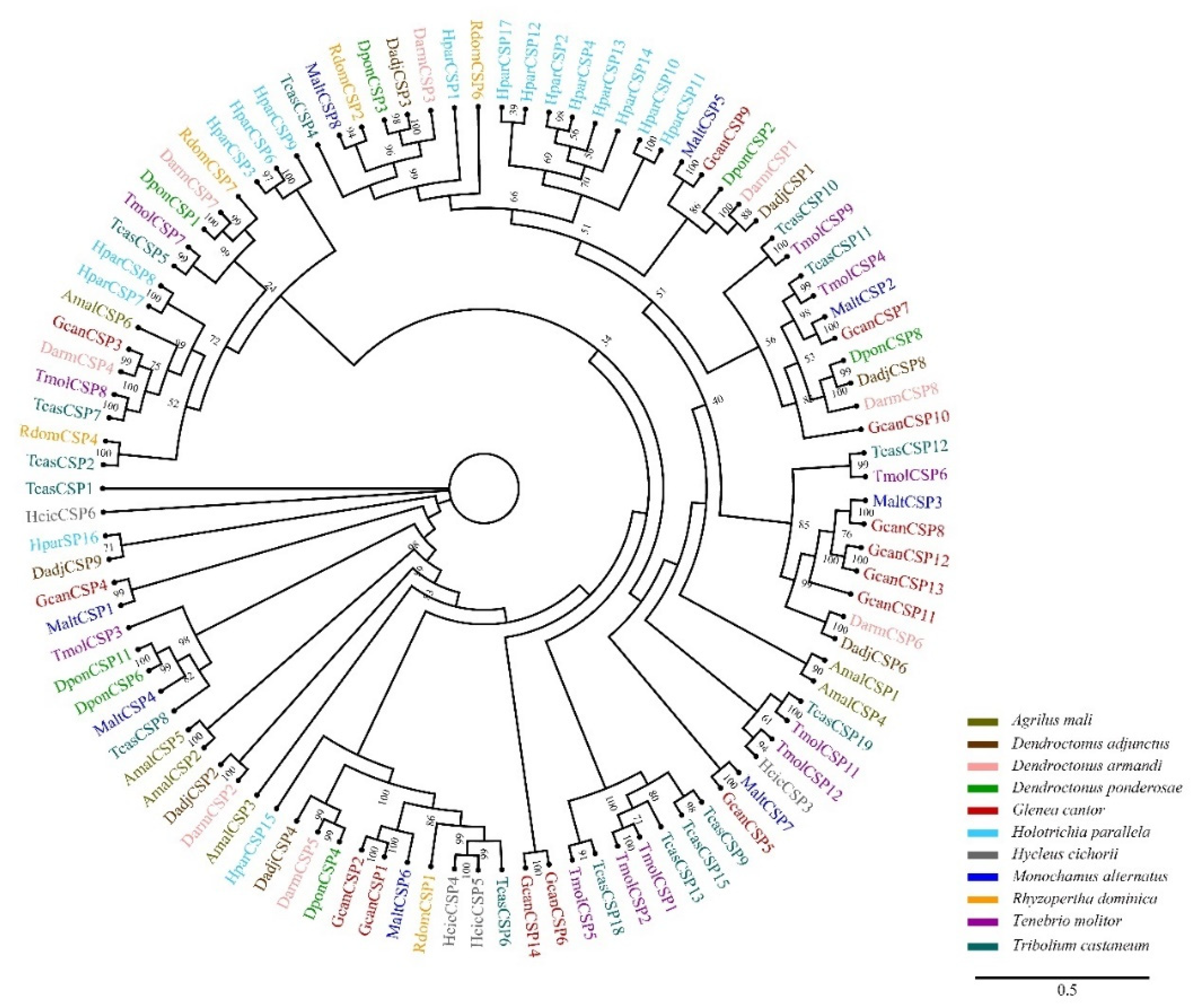
3.3. Identification of Candidate CSPs
3.4. Identification of Candidate ORs
3.5. Identification of Candidate IRs
3.6. Identification of Candidate SNMPs
3.7. Transcriptome Validation via Quantitative RT-PCR
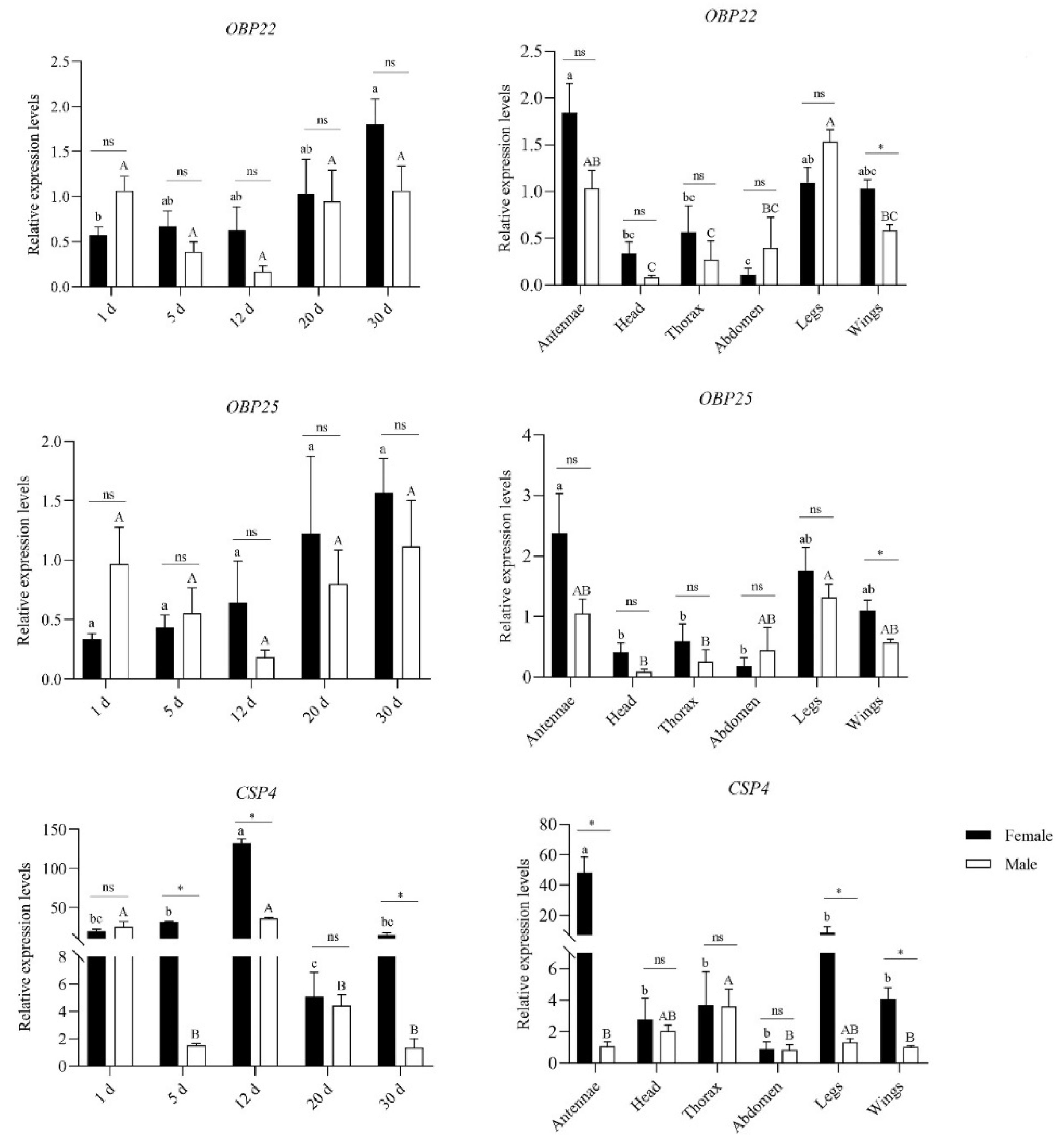
3.8. Expression Patterns of OBP and CSP Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hull, J.J.; Brent, C.S. Identification and characterization of a sex peptide receptor-like transcript from the western tarnished plant bug Lygus hesperus. Insect Mol. Biol. 2014, 23, 301–319. [Google Scholar] [PubMed]
- Zhang, Y.X.; Wang, W.L.; Li, M.Y.; Li, S.G.; Liu, S. Identification of putative carboxylesterase and aldehyde oxidase genes from the antennae of the rice leaffolder, Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). J. Asia-Pac. Entomol. 2017, 20, 907–913. [Google Scholar] [CrossRef]
- Huang, Z.Y.; Wang, X.Y.; Lu, W.; Zheng, X.L. Sensory gene identification in the transcriptome of the ectoparasitoid Quadrastichus mendeli. Sci. Rep. 2021, 11, 9726. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Hao, E.H.; Qiao, H.L.; Wang, J.Z.; Wu, W.W.; Zhou, J.J.; Lu, P.F. Antennal transcriptome analysis of olfactory genes and characterizations of odorant binding proteins in two woodwasps, Sirex noctilio and Sirex nitobei (Hymenoptera: Siricidae). BMC Genom. 2021, 22, 172. [Google Scholar] [CrossRef]
- Yi, J.K.; Yang, S.; Wang, S.; Wang, J.; Zhang, X.X.; Liu, Y.; Xi, J.H. Identification of candidate chemosensory receptors in the antennal transcriptome of the large black chafer Holotrichia parallela Motschulsky (Coleoptera: Scarabaeidae). Comp. Biochem. Physiol. D-Genom. Proteom. 2018, 28, 63–71. [Google Scholar] [CrossRef]
- Zhou, X.; Guo, J.; Zhang, M.; Bai, C.; Wang, Z.; Li, Z. Antennal transcriptome analysis and candidate olfactory genes in Crematogaster rogenhoferi. Bull. Entomol. Res. 2021, 111, 464–475. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Chen, Q.I.; Zhao, H.; Wang, J.; Wen, M.; Xi, J.; Ren, B. Identification and evolution of olfactory genes in the small poplar longhorn beetle Saperda populnea. Comp. Biochem. Physiol. D-Genom. Proteom. 2018, 26, 58–68. [Google Scholar] [CrossRef]
- Zeng, Y.; Yang, Y.T.; Wu, Q.J.; Wang, S.L.; Xie, W.; Zhang, Y.J. Genome-wide analysis of odorant-binding proteins and chemosensory proteins in the sweet potato whitefly, Bemisia tabaci. Insect Sci. 2019, 26, 620–634. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.P.; Du, L.X.; Zhu, Y.; Yang, S.Y.; Zhou, Q.; Wang, G.R.; Liu, Y. Identification and sex-biased profiles of candidate olfactory genes in the antennal transcriptome of the parasitoid wasp Cotesia vestalis. Comp. Biochem. Physiol. D-Genom. Proteom. 2020, 34, 100657. [Google Scholar] [CrossRef]
- Brito, N.F.; Moreira, M.F.; Melo, C.A. A look inside odorant-binding proteins in insect chemoreception. J. Insect Physiol. 2016, 95, 51–65. [Google Scholar] [CrossRef]
- Sun, L.; Wang, Q.; Zhang, Y.X.; Tu, X.H.; Yan, Y.T.; Wang, Q.; Dong, K.; Zhang, Y.J.; Xiao, Q. The sensilla trichodea-biased EoblPBP1 binds sex pheromones and green leaf volatiles in Ectropis obliqua Prout, a geometrid moth pest that uses Type-II sex pheromones. J. Insect Physiol. 2019, 116, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Calvello, M.; Ban, L.P. Diversity of Odorant-binding Proteins and Chemosensory Proteins in Insects. Chem. Senses 2005, 30 (Suppl. S1), i291–i292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.R.; Dani, F.R. Beyond chemoreception: Diverse tasks of soluble olfactory proteins in insects. Biol. Rev. 2018, 93, 184–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.P.; Liu, Y.; Jiang, X.C.; Wang, G.R. Cloning and functional characterization of three new pheromone receptors from the diamondback moth, Plutella xylostella. J. Insect Physiol. 2018, 107, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.L.; Dong, J.F.; Ning, C.; Ding, P.P.; Huang, L.Q.; Sun, J.G.; Wang, C.Z. An odorant receptor mediates the attractiveness of cis-jasmone to Campoletis chlorideae, the endoparasitoid of Helicoverpa armigera. Insect Mol. Biol. 2019, 28, 23–34. [Google Scholar] [CrossRef] [Green Version]
- An, X.K.; Sun, L.; Liu, H.W.; Liu, D.F.; Ding, Y.X.; Li, L.M.; Zhang, Y.J.; Guo, Y.Y. Identification and expression analysis of an olfactory receptor gene family in green plant bug Apolygus lucorum (Meyer-Dur). Sci. Rep. 2016, 6, 37870. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Gong, X.L.; Li, G.C.; Mo, B.T.; Jiang, N.J.; Huang, L.Q.; Wang, C.Z. Functional analysis of pheromone receptor repertoire in the fall armyworm, Spodoptera frugiperda. Pest Manag. Sci. 2022, 78, 2052–2064. [Google Scholar] [CrossRef]
- Koh, T.W.; He, Z.; Gorur-Shandilya, S.; Menuz, K.; Larter, N.K.; Stewart, S.; Carlson, J.R. The Drosophila IR20a clade of ionotropic receptors are candidate taste and pheromone receptors. Neuron 2014, 83, 850–865. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Zhang, W.; Nie, R.E.; Li, W.Z.; Segraves, K.A.; Yang, X.K.; Xue, H.J. Comparative transcriptome analysis of chemosensory genes in two sister leaf beetles provides insights into chemosensory speciation. Insect Biochem. Mol. Biol. 2016, 79, 108–118. [Google Scholar] [CrossRef]
- Gu, S.H.; Yang, R.N.; Guo, M.B.; Wang, G.R.; Wu, K.M.; Guo, Y.Y.; Zhou, J.J.; Zhang, Y.J. Molecular identification and differential expression of sensory neuron membrane proteins in the antennae of the black cutworm moth Agrotis ipsilon. J. Insect Physiol. 2013, 59, 430–443. [Google Scholar] [CrossRef]
- Jiang, X.C.; Pregitzer, P.; Grosse-Wilde, E.; Breer, H.; Krieger, J. Identification and characterization of two “sensory neuron membrane proteins” (SNMPs) of the desert locust, Schistocerca gregaria (Orthoptera: Acrididae). J. Insect Sci. 2016, 16, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, W.; Wang, Q.; Tian, M.Y.; Xu, J.; Qin, A.Z. Phenology and laboratory rearing procedures of an asian longicorn beetle, Glenea cantor (Coleoptera: Cerambycidae: Lamiinae). J. Econ. Entomol. 2011, 104, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Wang, Q.; Tian, M.Y.; Xu, J.; Lv, J.; Wei, S.G.; Qin, A.Z. Reproductive traits of Glenea cantor (Coleoptera: Cerambycidae: Lamiinae). J. Econ. Entomol. 2013, 106, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Tian, M.Y.; Lai, K.P.; Wei, S.G.; Qin, A.Z. Occurrence and control of Glenea cantor. Guangxi Plant Prot. 2006, 3, 1–4. (In Chinese) [Google Scholar]
- Lei, Y.M. Glenea cantor’s damage to kapok trees and its comprehensive control. China Trop. Agric. 2015, 3, 52–54. (In Chinese) [Google Scholar]
- Dong, Z.S.; Zhang, Y.J.; Lai, K.P.; Zheng, X.L.; Wang, Q.; Lu, W. Biological characteristics of Glenea cantor Fabricius (Coleoptera: Cerambycidae). J. Environ. Entomol. 2017, 39, 1313–1318. (In Chinese) [Google Scholar]
- Lai, K.; Lu, W.; Liu, D.X.; Luo, Z.R.; Gao, P.Y. The larval instars and stadia of the longhorn beetle Glenea cantor. Chin. J. Appl. Entomol. 2008, 1, 138–140. (In Chinese) [Google Scholar]
- Dong, Z.S.; Yang, Y.B.; Dou, F.G.; Zhang, Y.J.; Huang, H.X.; Zheng, X.L.; Wang, X.Y.; Lu, W. Observations on the ultrastructure of antennal sensilla of adult Glenea cantor (Cerambycidae: Lamiinae). J. Insect Sci. 2020, 20, 7. [Google Scholar] [CrossRef]
- Huang, H.; Lu, W.; Nong, C.; Nong, Y.Y. A preliminary study on artificial diet of Glenea cantor. Guangxi Plant Prot. 2007, 1, 9–11. (In Chinese) [Google Scholar]
- Wang, X.Y.; Zheng, X.L.; Lu, W. The complete mitochondrial genome of an Asian longicorn beetle Glenea cantor (Coleoptera: Cerambycidae: Lamiinae). Mitochondrial DNA Part B 2019, 4, 2906–2907. [Google Scholar] [CrossRef] [Green Version]
- Su, R.R.; Huang, Z.Y.; Qin, C.W.; Zheng, X.L.; Lu, W.; Wang, X.Y. Evaluation of reference genes in Glenea cantor (Fabricius) by using qRT-PCR. Genes 2021, 12, 1984. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.; Haas, B.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using Real-Time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.Y.; Cao, D.P.; Wang, G.R.; Liu, Y. Identification of genes involved in chemoreception in Plutella xyllostella by antennal transcriptome analysis. Sci. Rep. 2017, 7, 11941. [Google Scholar] [CrossRef] [Green Version]
- Xing, Y.; Thanasirungkul, W.; Adeel, M.M.; Yu, J.; Aslam, A.; Chi, D.F. Identification and analysis of olfactory genes in Dioryctria abietella based on the antennal transcriptome. Comp. Biochem. Physiol. D-Genom. Proteom. 2021, 38, 100814. [Google Scholar] [CrossRef]
- Richards, S.; Gibbs, R.A.; Weinstock, G.M.; Brown, S.J.; Denell, R.; Beeman, R.W.; Gibbs, R.; Bucher, G.; Friedrich, M.; Grimmelikhuijzen, C.J.P. The genome of the model beetle and pest Tribolium castaneum. Nature 2008, 452, 949–955. [Google Scholar]
- Hu, P.; Wang, J.Z.; Cui, M.M.; Tao, J.; Luo, Y.Q. Antennal transcriptome analysis of the Asian longhorned beetle Anoplophora glabripennis. Sci. Rep. 2016, 6, 26652. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, D.Z.; Min, S.F.; Mi, F.; Zhou, S.S.; Wang, M.Q. Analysis of chemosensory gene families in the beetle Monochamus alternatus and its parasitoid Dastarcus helophoroides. Comp. Biochem. Physiol. D-Genom. Proteom. 2014, 11, 1–8. [Google Scholar] [CrossRef]
- Wang, B.; Liu, Y.; Wang, G.R. Chemosensory genes in the antennal transcriptome of two syrphid species, Episyrphus balteatus and Eupeodes corollae (Diptera: Syrphidae). BMC Genom. 2017, 18, 586. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Li, F.Q.; Zhang, W.; Zhang, X.M.; Qu, C.; Tetreau, G.; Sun, L.J.; Luo, C.; Zhou, J.J. Identification and expression profile analysis of odorant binding protein and chemosensory protein genes in Bemisia tabaci MED by head transcriptome. PLoS ONE 2017, 12, e0171739. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Wang, K.; Zeng, Y.; Guo, Z.; Zhang, Y.; Wu, Q.; Wang, S. Analysis of the antennal transcriptome and odorant-binding protein expression profiles of the parasitoid wasp Encarsia formosa. Genomics 2020, 112, 2291–2301. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Z.; Gao, P.; Luo, Y.Q.; Tao, J. Characterization and expression profiling of odorant-binding proteins in Anoplophora glabripennis Motsch. Gene 2019, 693, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Harada, E.; Nakagawa, J.; Asano, T.; Taoka, M.; Sorimachi, H.; Ito, Y.; Aigaki, T.; Matsuo, T. Functional evolution of duplicated odorant-binding protein genes, Obp57d and Obp57e, in Drosophila. PLoS ONE 2012, 7, e29710. [Google Scholar] [CrossRef]
- Song, Y.Q.; Sun, H.Z.; Du, J. Identification and tissue distribution of chemosensory protein and odorant binding protein genes in Tropidothorax elegans Distant (Hemiptera: Lygaeidae). Sci. Rep. 2018, 8, 7803. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.B.; Ding, Y.X.; Gu, S.H.; Sun, L.; Zhu, X.Q.; Liu, H.W.; Dhiloo, K.H.; Zhang, Y.J.; Guo, Y.Y. Molecular characterization and expression profiling of odorant-binding proteins in Apolygus lucorum. PLoS ONE 2015, 10, e0140562. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zhang, Y.J.; Su, H.H.; Gao, X.W.; Guo, Y.Y. Identification and expression pattern of putative odorant-binding proteins and chemosensory proteins in antennae of the Microplitis mediator (Hymenoptera: Braconidae). Chem. Senses 2009, 34, 503–512. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.J.C.; Chen, H.C.; Hong, T.L.; Yan, M.W.; Wang, J.; Shao, Z.M.; Wu, F.A.; Sheng, S.; Wang, J. Identification of chemosensory genes by antennal transcriptome analysis and expression profiles of odorant-binding proteins in parasitoid wasp Aulacocentrum confusum. Comp. Biochem. Physiol. D-Genom. Proteom. 2021, 40, 100881. [Google Scholar] [CrossRef]
- Rojas, V.; Jimenez, H.; Palma-Millanao, R.; Gonzalez-Gonzalez, A.; Machuca, J.; Godoy, R.; Ceballos, R.; Mutis, A.; Venthur, H. Analysis of the grapevine moth Lobesia botrana antennal transcriptome and expression of odorant-binding and chemosensory proteins. Comp. Biochem. Physiol. D-Genom. Proteom. 2018, 27, 1–12. [Google Scholar] [CrossRef]
- Liu, G.X.; Ma, H.M.; Xie, H.Y.; Xuan, N.; Guo, X.; Fan, Z.X.; Rajashekar, B.; Arnaud, P.; Offmann, B.; Picimbon, J.F. Biotype characterization, developmental profiling, insecticide response and binding property of Bemisia tabaci chemosensory proteins: Role of CSP in insect defense. PLoS ONE 2016, 11, e0154706. [Google Scholar] [CrossRef] [Green Version]
- Gu, S.H.; Wang, S.Y.; Zhang, X.Y.; Ji, P.; Liu, J.T.; Wang, G.R.; Wu, K.M.; Guo, Y.Y.; Zhou, J.J.; Zhang, Y.J. Functional characterizations of chemosensory proteins of the alfalfa plant bug Adelphocoris lineolatus indicate their involvement in host recognition. PLoS ONE 2012, 7, e42871. [Google Scholar] [CrossRef]
- Zhang, Y.N.; Ye, Z.F.; Yang, K.; Dong, S.L. Antenna-predominant and male-biased CSP19 of Sesamia inferens is able to bind the female sex pheromones and host plant volatiles. Gene 2014, 536, 279–286. [Google Scholar] [CrossRef]
- Lu, W.; Wang, Q.; Tian, M.Y.; Xu, J.; Qin, A.Z.; He, L.; Jia, B.; Cai, J.J. Host selection and colonization strategies with evidence for a female-produced oviposition attractant in a longhorn beetle. Environ. Entomol. 2011, 40, 1487–1493. [Google Scholar] [CrossRef] [PubMed]
- Leal, W.S. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.N.; Grosse-Wilde, E.; Keeling, C.I.; Bengtsson, J.M.; Yuen, M.M.S.; Li, M.; Hillbur, Y.; Bohlmann, J.; Hansson, B.S.; Schlyter, F. Antennal transcriptome analysis of the chemosensory gene families in the tree killing bark beetles, Ips typographus and Dendroctonus ponderosae (Coleoptera: Curculionidae: Scolytinae). BMC Genom. 2013, 14, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Ju, Q.; Jie, W.; Li, F.; Jiang, X.; Jiang, X.J.; Hu, J.J.; Qu, M.J. Correction: Chemosensory gene families in adult antennae of Anomala corpulenta Motschulsky (Coleoptera: Scarabaeidae: Rutelinae). PLoS ONE 2015, 10, e0144214. [Google Scholar] [CrossRef]
- Wang, S.N.; Peng, Y.; Lu, Z.Y.; Dhiloo, K.H.; Gu, S.H.; Li, R.J.; Zhou, J.J.; Zhang, Y.J.; Guo, Y.Y. Identification and expression analysis of putative chemosensory receptor genes in Microplitis mediator by antennal transcriptome screening. Int. J. Biol. Sci. 2015, 11, 737–751. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, R.F.; Hughes, D.T.; Luetje, C.W.; Millar, J.G.; Soriano-Agatón, F.; Hanks, L.M.; Robertson, H.M. Sequencing and characterizing odorant receptors of the cerambycid beetle Megacyllene caryae. Insect Biochem. Mol. Biol. 2012, 42, 499–505. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Buhl, E.; Xu, M.; Croset, V.; Rees, J.S.; Lilley, K.S.; Benton, R.; Hodge, J.J.L.; Stanewsky, R. Drosophila ionotropic receptor 25a mediates circadian clock resetting by temperature. Nature 2015, 527, 516–520. [Google Scholar] [CrossRef]
- Benton, R.; Vannice, K.S.; Gomez-Diaz, C.; Vosshall, L.B. Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila. Cell 2009, 136, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Cassau, S.; Krieger, J. The role of SNMPs in insect olfaction. Cell Tissue Res. 2021, 383, 21–33. [Google Scholar] [CrossRef]
- Shan, S.; Wang, S.N.; Song, X.; Khashaveh, A.; Lu, Z.Y.; Dhiloo, K.H.; Li, R.J.; Gao, X.W.; Zhang, Y.J. Molecular characterization and expression of sensory neuron membrane proteins in the parasitoid Microplitis mediator (Hymenoptera: Braconidae). Insect Sci. 2020, 27, 425–439. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Wang, C.Y.; Lyu, Z.H.; Chen, J.X.; Tang, L.P.; Lin, T. Candidate olfactory genes identified in Heortia vitessoides (Lepidoptera: Crambidae) by antennal transcriptome analysis. Comp. Biochem. Physiol. D-Genom. Proteom. 2019, 29, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.T.; Xu, L.B.; Duan, L.Q.; Yu, L.B.; Cui, J.; Cao, Y.Y.; Zhao, Y.A. Transcriptome sequencing and identification of the Zele chlorophthalmus olfactory related protein gene. Chin. J. Appl. Entomol. 2021, 58, 846–855. (In Chinese) [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Female | Male | |
|---|---|---|
| Raw reads | 40,989,086 | 42,482,724 |
| Clean reads | 38,130,472 | 42,395,862 |
| N percentage | 0.00% | 0.00% |
| Q20 percentage | 99.78% | 99.80% |
| GC percentage | 48.82% | 48.42% |
| Assembly unigene | 24,462 | |
| GC percentage | 44.18% | |
| N50 | 8089 | |
| N50 max length (bp) | 12,058 | |
| N50 min length (bp) | 116 | |
| Gene Name | Unigene ID | Unigene Length (bp) | ORF (aa) | Complete ORF | Signal Peptide | Cysteine Number | Homology Search with Known Protein | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Name | Species | E-Value | Accession | Identity (%) | |||||||
| OBP1 | Isoform0013634 | 189 | 62 | YES | 0 | 2 | odorant binding protein 7 | Xylotrechus quadripes | 8 × 10−20 | AXO78385.1 | 62.9 |
| OBP2 | Isoform0013768 | 351 | 116 | YES | 0 | 3 | odorant binding protein 7 | Anoplophora glabripennis | 3 × 10−20 | ARH65462.1 | 50 |
| OBP3 | Isoform0018207 | 534 | 177 | YES | 22 | 10 | odorant binding protein | Anoplophora chinensis | 2 × 10−99 | AUF72969.1 | 80.34 |
| OBP4 | Isoform0021420 | 300 | 99 | YES | 20 | 4 | odorant binding protein 1 | Monochamus alternatus | 5 × 10−22 | ABR53888.1 | 55.68 |
| OBP5 | Isoform0022365 | 702 | 233 | YES | 26 | 11 | minus-C odorant binding protein 4, partial | Anoplophora glabripennis | 6 × 10−76 | ATP75519.1 | 79.5 |
| OBP6 | Isoform0022758 | 384 | 127 | YES | 20 | 5 | odorant-binding protein 18 | Monochamus alternatus | 3 × 10−18 | AIX97033.1 | 40.5 |
| OBP7 | Isoform0023655 | 744 | 247 | YES | 17 | 13 | odorant-binding protein 24 | Monochamus alternatus | 2 × 10−82 | AIX97039.1 | 73.53 |
| OBP8 | Isoform0023835 | 474 | 157 | YES | 21 | 6 | odorant-binding protein 14 | Monochamus alternatus | 5 × 10−63 | AIX97029.1 | 79.49 |
| OBP9 | Isoform0023942 | 669 | 203 | YES | 19 | 10 | odorant-binding protein 19 | Monochamus alternatus | 5 × 10−86 | AIX97034.1 | 70.39 |
| OBP10 | Isoform0024133 | 432 | 143 | YES | 20 | 5 | odorant-binding protein 1 | Monochamus alternatus | 1 × 10−37 | ABR53888.1 | 50.36 |
| OBP11 | Isoform0024165 | 405 | 134 | YES | 18 | 5 | odorant-binding protein 2 | Monochamus alternatus | 2 × 10−64 | AHA39267.1 | 78.51 |
| OBP12 | Isoform0024173 | 429 | 142 | YES | 17 | 5 | odorant-binding protein 6 | Monochamus alternatus | 3 × 10−66 | AJO67868.1 | 73.33 |
| OBP13 | Isoform0024208 | 429 | 142 | YES | 20 | 8 | odorant-binding protein 4 | Monochamus alternatus | 1 × 10−69 | AHA39269.1 | 69.01 |
| OBP14 | Isoform0024216 | 429 | 142 | YES | 20 | 9 | odorant binding protein 14 | Anoplophora glabripennis | 2 × 10−71 | ARH65469.1 | 80.31 |
| OBP15 | Isoform0024236 | 432 | 143 | YES | 20 | 5 | odorant-binding protein 1 | Monochamus alternatus | 2 × 10−39 | ABR53888.1 | 51.09 |
| OBP16 | Isoform0024238 | 399 | 132 | YES | 18 | 4 | odorant-binding protein 20 | Monochamus alternatus | 2 × 10−41 | AIX97035.1 | 52.27 |
| OBP17 | Isoform0024250 | 405 | 134 | YES | 17 | 6 | odorant binding protein 6 | Monochamus alternatus | 5 × 10−73 | AJO67868.1 | 81.48 |
| OBP18 | Isoform0024256 | 402 | 133 | YES | 17 | 4 | odorant binding protein 5 | Anoplophora glabripennis | 9 × 10−28 | ARH65460.1 | 34.65 |
| OBP19 | Isoform0024263 | 420 | 139 | YES | 22 | 7 | odorant binding protein 7 | Anoplophora glabripennis | 5 × 10−44 | ARH65462.1 | 56.43 |
| OBP20 | Isoform0024283 | 414 | 137 | YES | 19 | 6 | odorant-binding protein 10 | Monochamus alternatus | 2 × 10−81 | AIX97025.1 | 83.21 |
| OBP21 | Isoform0024286 | 405 | 134 | YES | 18 | 5 | odorant binding protein 7 | Xylotrechus quadripes | 3 × 10−520 | AXO78385.1 | 60.61 |
| OBP22 | Isoform0024293 | 402 | 133 | YES | 17 | 5 | odorant-binding protein 5 | Monochamus alternatus | 3 × 10−82 | AIX97020.1 | 89.47 |
| OBP23 | Isoform0024294 | 429 | 142 | YES | 25 | 5 | odorant-binding protein 11 | Monochamus alternatus | 5 × 10−75 | AIX97026.1 | 82.48 |
| OBP24 | Isoform0024314 | 405 | 134 | YES | 18 | 5 | odorant-binding protein 2 | Monochamus alternatus | 2 × 10−64 | AHA39267.1 | 78.51 |
| OBP25 | Isoform0024319 | 405 | 134 | YES | 17 | 4 | odorant-binding protein 5 | Monochamus alternatus | 6 × 10−73 | AIX97020.1 | 85.07 |
| OBP26 | Isoform0024320 | 390 | 129 | YES | 17 | 4 | odorant-binding protein 21 | Monochamus alternatus | 4 × 10−41 | AIX97036.1 | 55.04 |
| OBP27 | Isoform0024323 | 462 | 136 | YES | 21 | 7 | odorant-binding protein 3 | Anoplophora glabripennis | 1 × 10−59 | ATO58974.1 | 61.03 |
| OBP28 | Isoform0024327 | 423 | 140 | YES | 20 | 5 | odorant binding protein 7 | Anoplophora glabripennis | 5 × 10−41 | ARH65462.1 | 52.71 |
| OBP29 | Isoform0024357 | 420 | 139 | YES | 23 | 6 | odorant binding protein 7 | Anoplophora glabripennis | 3 × 10−43 | ARH65462.1 | 55.22 |
| CSP1 | Isoform0017649 | 882 | 293 | YES | 21 | 4 | chemosensory protein 6 | Monochamus alternatus | 5 × 10−74 | AIX97046.1 | 82.58 |
| CSP2 | Isoform0019553 | 840 | 279 | YES | 21 | 4 | chemosensory protein 6 | Monochamus alternatus | 3 × 10−74 | AIX97046.1 | 82.58 |
| CSP3 | Isoform0023484 | 375 | 124 | YES | 20 | 4 | chemosensory protein CSP8 | Tenebrio molitor | 6 × 10−54 | AJO62214.1 | 76.92 |
| CSP4 | Isoform0023726 | 357 | 118 | YES | 24 | 5 | chemosensory protein 11 | Colaphellus bowringi | 2 × 10−49 | ALR72525.1 | 90.36 |
| CSP5 | Isoform0024206 | 378 | 125 | YES | 23 | 4 | chemosensory protein 7 | Monochamus alternatus | 1 × 10−46 | AIX97047.1 | 63.79 |
| CSP6 | Isoform0024227 | 378 | 125 | YES | 18 | 5 | CSP9 | Anoplophora glabripennis | 4 × 10−53 | ATL75742.1 | 61.6 |
| CSP7 | Isoform0024262 | 219 | 72 | YES | 22 | 0 | chemosensory protein 2 | Monochamus alternatus | 1 × 10−10 | AIX97042.1 | 89.66 |
| CSP8 | Isoform0024264 | 381 | 126 | YES | 19 | 4 | chemosensory protein 11 | Monochamus alternatus | 1 × 10−59 | AIX97086.1 | 77.06 |
| CSP9 | Isoform0024328 | 390 | 129 | YES | 19 | 5 | chemosensory protein 5 | Monochamus alternatus | 1 × 10−59 | AIX97045.1 | 80.91 |
| CSP10 | Isoform0024344 | 387 | 128 | YES | 19 | 5 | chemosensory protein 8 | Dendroctonus ponderosae | 5 × 10−50 | AGI05164.1 | 61.79 |
| CSP11 | Isoform0024345c | 381 | 126 | YES | 19 | 5 | chemosensory protein 11 | Monochamus alternatus | 1 × 10−69 | AIX97086.1 | 75.2 |
| CSP12 | Isoform0024347 | 381 | 126 | YES | 19 | 4 | chemosensory protein 11 | Monochamus alternatus | 2 × 10−75 | AIX97086.1 | 85.6 |
| CSP13 | Isoform0024362 | 381 | 126 | YES | 19 | 4 | chemosensory protein 11 | Monochamus alternatus | 5 × 10−66 | AIX97086.1 | 82.4 |
| CSP14 | Isoform0024366 | 378 | 125 | YES | 18 | 5 | CSP9 | Anoplophora glabripennis | 4 × 10−53 | ATL75742.1 | 61.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, G.; Su, R.; Ouyang, H.; Zheng, X.; Lu, W.; Wang, X. Antennal Transcriptome Analysis and Identification of Olfactory Genes in Glenea cantor Fabricius (Cerambycidae: Lamiinae). Insects 2022, 13, 553. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060553
Wu G, Su R, Ouyang H, Zheng X, Lu W, Wang X. Antennal Transcriptome Analysis and Identification of Olfactory Genes in Glenea cantor Fabricius (Cerambycidae: Lamiinae). Insects. 2022; 13(6):553. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060553
Chicago/Turabian StyleWu, Guanxin, Ranran Su, Huili Ouyang, Xialin Zheng, Wen Lu, and Xiaoyun Wang. 2022. "Antennal Transcriptome Analysis and Identification of Olfactory Genes in Glenea cantor Fabricius (Cerambycidae: Lamiinae)" Insects 13, no. 6: 553. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060553