
Triatoma guazu Lent and Wygodzinsky Is a Junior Synonym of Triatoma williami Galvão, Souza and Lima

 ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jeannel, R. Insectes Hemipteres, iii. Henicocephalidae et Reduviidae. In Voyage Alluaud et R. Jeannel en Afrique orientale (1911–1912); Résultats Scientifiques: Paris, France, 1919; Volume 3, pp. 131–314. [Google Scholar]
- Galvão, C. Taxonomia dos Vetores da Doença de Chagas da Forma à Molécula, quase três séculos de história. In Atualidades em Medicina Tropical no Brazil: Vetores, 1st ed.; Oliveira, J., Chaboli, K., Aranha, L., Meneguetti, D., Eds.; Stricto Sensu: Rio Branco, Brazil, 2020; pp. 9–37. Available online: https://sseditora.com.br/wp-content/uploads/Atualidades-em-Medicina-Tropical-no-Brasil-Vetores.pdf (accessed on 6 April 2022).
- Zhao, Y.; Galvão, C.; Cai, W. Rhodnius micki, a new species of Triatominae (Hemiptera, Reduviidae) from Bolivia. Zookeys 2021, 93, 71–93. [Google Scholar] [CrossRef] [PubMed]
- Gil-Santana, H.R.; Chavez, T.; Pita, S.; Panzera, F.; Galvão, C. Panstrongylus noireaui, a remarkable new species of Triatominae (Hemiptera, Reduviidae) from Bolivia. Zookeys 2022, in press. [Google Scholar] [CrossRef]
- Laporte, F.L. Essai d’une classification systematique de l’ordre des Hemipteres (Hemipteres Heteropteres, Latr.). Mag. Zool. 1832, 2, 51–55. [Google Scholar]
- Dale, C.; Justi, S.A.; Galvão, C. Belminus santosmalletae (Hemiptera: Heteroptera: Reduviidae): New Species from Panama, with an Updated Key for Belminus Stål, 1859 Species. Insects 2021, 12, 686. [Google Scholar] [CrossRef] [PubMed]
- Galvão, C.; Gurgel-Gonçalves, R. Vetores conhecidos no Brasil. In Vetores da Doença de Chagas no Brasil, 1st ed.; Galvão, C., Ed.; Sociedade brasileira de Zoologia: Curitiba, Brasil, 2015; Volume 1, pp. 88–170. [Google Scholar]
- Mendonça, V.J.; Alevi, K.C.C.; Pinotti, H.; Gurgel-Gonçalves, R.; Pita, S.; Guerra, A.L.; Panzera, F.; Araújo, R.F.; Azeredo-Oliveira, M.T.V.; Rosa, J.A. Revalidation of Triatoma bahiensis Sherlock & Serafim, 1967 (Hemiptera: Reduviidae) and phylogeny of the T. brasiliensis species complex. Zootaxa 2016, 2, 239–254. [Google Scholar] [CrossRef]
- Galvão, A.B.; da Silva e Souza, H.; Lima, R.R. Triatoma williami n. sp. (Hemiptera: Triatominae). Rev. Bras. Malariol. Doenças Trop. 1965, 17, 363–366. [Google Scholar]
- Martins, M.F.; Moraes, S.C.; Oliveira, J.; Santos, J.C.; Santos-Silva, L.K.; Galvão, C. Triatoma williami in intradomiciliary environments of urban areas in Mato Grosso State Brazil: Domiciliation process of a wild species? Infect. Dis. Poverty 2022, 11, 18. [Google Scholar] [CrossRef]
- Stål, C. Monographie der Gattung Conorhinus und Verwandten. Berl. Entomol. Zeitschriff 1859, 3, 99–117. [Google Scholar] [CrossRef]
- Wygodzinsky, P.; Abalos, J.W. Triatoma guasayana sp. n. (Triatominae, Reduviidae, Hemiptera) (Nota previa). Sem. Médica 1949, 56, 2. [Google Scholar]
- Lent, H.; Wygodzinsky, P. Revision of the Triatominae (Hemiptera, Reduviidae) and Their Significance as Vectors of Chagas Disease, 1st ed.; American Museum of Natural History: New York, NY, USA, 1979; pp. 1–408. [Google Scholar]
- Lent, H.; Jurberg, J.; Galvão, C. Descrição do Alótipo (Macho) de Triatoma guazu Lent & Wygodzinsky, 1979 Proveniente do Estado do Mato Grosso, Brasil (Hemiptera, Reduviidae). Memórias Inst. Oswaldo Cruz 1996, 91, 313–315. [Google Scholar]
- Noireau, F.; Santos, S.M.; Gumiel, M.; Dujardin, J.-P.; Soares, M.S.; Carcavallo, R.U.; Galvão, C.; Jurberg, J. Phylogenetic relationships within the oliveirai complex (Hemiptera: Reduviidae: Triatominae). Infect. Genet. Evol. 2002, 2, 11–17. [Google Scholar] [CrossRef]
- Teves, S.C.; Gonçalves, T.C.M.; Freitas, S.P.C.; Lopes, C.M.; Carbajal-de-la-Fuente, A.L.; Santos-Mallet, J.R. External female genitalia of Triatoma jatai, Triatoma costalimai and Triatoma williami (Hemiptera: Reduviidae: Triatominae). Parasites Vectors 2020, 13, 538–544. [Google Scholar] [CrossRef]
- Sainz, A.C.; Mauro, L.V.; Moriyama, N.E.; García, B. Phylogeny of triatomine vectors of Trypanosoma cruzi suggested by mitochondrial DNA sequences. Genetica 2004, 121, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Hypša, V.; Tietz, D.F.; Zrzavý, J.; Rego, R.O.M.; Galvão, C.; Jurberg, J. Phylogeny and biogeography of Triatomine (Hemiptera: Reduviidae): Molecular evidence of a new world origin of the Asiatic clade. Mol. Phylogenetics Evol. 2002, 23, 447–457. [Google Scholar] [CrossRef]
- Gardim, S.; Rocha, C.S.; Almeida, C.E.; Takiya, D.M.; Silva, M.T.A.; Ambrósio, D.L.; Cicarelli, R.M.B.; Rosa, J.A. Evolutionary relationships of the Triatoma matogrossensis subcomplex, the endemic Triatoma in Central-Western Brazil, based on mitochondrial DNA sequences. Am. J. Trop. Med. Hyg. 2013, 89, 766–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Justi, S.A.; Russo, C.A.M.; Santos-Mallet, J.R.; Obara, M.T.; Galvão, C. Molecular phylogeny of Triatomini (Hemiptera: Reduviidae: Triatominae). Parasites Vectors 2014, 7, 149–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teves, S.C.; Gardim, S.; Carbajal de la Fuente, A.L.; Lopes, C.M.; Monte Gonçalves, T.C.; dos Santos Mallet, J.R.; da Rosa, J.A.; Almeida, C.E. Mitochondrial genes reveal Triatoma jatai as a sister to Triatoma costalimai (Reduviidae: Triatominae). Am. J. Trop. Med. Hyg. 2016, 94, 686–688. [Google Scholar] [CrossRef] [Green Version]
- Kieran, T.J.; Gordon, E.R.L.; Zaldívar-Riverón, A.; Ibarra-Cerdeña, C.N.; Glenn, T.; Weirauch, C. Ultraconserved elements reconstruct the evolution of Chagas disease vectoring kissing bugs (Reduviidae: Triatominae). Syst. Entomol. 2021, 46, 725–740. [Google Scholar] [CrossRef]
- Alevi, K.C.C.; Reis, Y.V.; Mendonça, V.J.; Rosa, J.A.; Azeredo Oliveira, M.T.V. Classical cytotaxonomy the Matogrossensis subcomplex (Hemiptera, Triatominae). Invertebr. Reprod. Dev. 2014, in press. [Google Scholar]
- Pita, S.; Lorite, P.; Nattero, J.; Galvão, C.; Alevi, K.C.C.; Teves, S.C.; Azeredo-Oliveira, M.T.V.; Panzera, F. New arrangements on several species subcomplexes of Triatoma genus based on the chromosomal position of ribosomal genes (Hemiptera–Triatominae). Infect. Genet. Evol. 2016, 43, 225–231. [Google Scholar] [CrossRef] [Green Version]
- Reis, Y.V.; Alevi, K.C.C.; Rosa, J.A.; Azeredo-Oliveira, M.T.V. Espermiogênese como ferramenta citotaxonômica para diferenciar Triatoma guazu e T. williami, espécies vetoras da doença de Chagas. Rev. Ciências Farm. Básica Apl. 2015, 36, 293–296. [Google Scholar]
- Rohlf, F.J. TpsDig, Version 2.1.1. Department of Ecology and Evolution, State University of New York: Stony Brook, NY, USA. Available online: http://en.freedownloadmanager.org/Windows-PC/tpsDig2.html (accessed on 16 January 2022).
- Dujardin, J.-P. Morphometrics applied to medical entomology. Infect. Genet. Evol. 2008, 8, 875–890. [Google Scholar] [CrossRef] [PubMed]
- Rohlf, F.J. Rotational fit (Procrustes) methods. In Proceedings of the Michigan Morphometrics Workshop, 2nd ed.; Rohlf, F.J., Bookstein, F.L., Eds.; University of Michigan Museum of Zoology: Ann Arbor, MI, USA, 1990; Volume 2, pp. 227–236. [Google Scholar]
- Rohlf, F.J.; Marcus, L.F. A revolution morphometrics. Trends Ecol. Evol. 1993, 8, 129–132. [Google Scholar] [CrossRef]
- Zelditch, M.L.; Swiderski, D.L.; Sheets, H.D.; Fink, W.L. Geometric Morphometrics for Biologists: A Primer; Elsevier: Amsterdam, The Netherlands, 2004; pp. 1–443. [Google Scholar] [CrossRef]
- Forero, D.; Weirauch, C. Comparative genitalic morphology in the New World resin bugs Apiomerini (Hemiptera, Heteroptera, Reduviidae, Harpactorinae). Dtsch. Entomol. Z. 2012, 59, 5–41. [Google Scholar]
- Rédei, D.; Tsai, J.-F. The assassin bug subfamilies Centrocnemidinae and Holoptilinae in Taiwan (Hemiptera: Heteroptera: Reduviidae). Acta Entomol. Musei Natl. Pragae 2012, 51, 411–442. [Google Scholar]
- Neiva, A. Contribuição para o estudo dos hematophagos brasileiros e descrição de uma nova espécie de Triatoma. IBID 1911, 25, 421–422. [Google Scholar]
- Gonçalves, T.C.M.; Teves-Neves, S.C.; Santos-Mallet, J.R.; Carbajal-de-la-Fuente, A.L.; Lopes, C.M. Triatoma jatai sp. nov. in the state of Tocantins, Brazil (Hemiptera: Reduviidae: Triatominae). Memórias Inst. Oswaldo Cruz 2013, 108, 429–437. [Google Scholar] [CrossRef]
- GenBank. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/genbank/ (accessed on 13 January 2022).
- Thompson, D.J.; Higgins, D.G.; Gibson, T.J. CLUSTALW: Improving The Sensitivity of Progressive Multiple Sequence Alignment Through Sequence Weighting, Position-specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Revist. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Monteiro, F.A.; Perez, R.; Panzera, F.; Dujardin, J.-P.; Galvão, C.; Rocha, D.; Noireau, F.; Schofield, C.; Beard, C.B. Mitochondrial DNA variation of Triatoma infestans populations and its implication on the specific status of T. melanosoma. Memórias Inst. Oswaldo Cruz 1999, 94, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Almeida, C.E.; Marcet, P.L.; Gumiel, M.; Takiya, D.M.; Cardozo-de-Almeida, M.; Pacheco, R.S.; Lopes, C.M.; Dotson, E.M.; Costa, J. Phylogenetic and phenotypic relationship among Triatoma carcavalloi (Hemiptera: Reduvidade: Triatominae) and related species collected in domiciles in Rio Grande do Sul State, Brasil. J. Vector Ecol. 2009, 34, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Gardim, S.; Almeida, C.E.; Takiya, D.M.; Oliveira, J.; Araújo, R.F.; Cicarelli, R.M.B.; Rosa, J.A. Multiple mitochondrial genes of some sylvatic Brazilian Triatoma: Non-monophyly of the T. brasiliensis subcomplex and the need for a generic revision in the Triatomini. Infect. Genet. Evol. 2014, 23, 74–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lent, H.; Jurberg, J.; Galvão, C.; Carcavallo, R.U. Triatoma melanosoma, Novo Status para Triatoma infestans melanosoma Martinez, Olmedo & Carcavallo, 1987 (Hemiptera: Reduviidae). Memórias Inst. Oswaldo Cruz 1994, 98, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Klug, F. Reise um Die Erde Ausgeführt Auf Dem … Seehandlungs-Schiffe Prinzess Louise, Commandirt von Capitain W. Wendt in … 1830, 1831 und 1832, &c; Sander: Berlin, Germany, 1834; pp. 401–440. [Google Scholar]
- Catala, S.; Torres, M. Similarity of the patterns of sensilla on the antennae of Triatoma melanosoma and Triatoma infestans. Ann. Trop. Med. Parasitol. 2001, 95, 287–295. [Google Scholar] [CrossRef]
- Blanchard, E. Insectes de I’ Amérique Méridionale. In Voyage Dans I’ Amérique Méridionale Para Alcide d’ Orbigny; Bertrand, P., Ed.; Paris and Strasbourg: Paris, France, 1837; pp. 1–22. [Google Scholar]
- Almeida, C.E.; Pacheco, R.S.; Noireau, F.; Costa, J. Triatoma rubrovaria (Blanchard, 1843) (Hemiptera: Reduviidae) I: Isoenzymatic and Chromatic Patterns of five populations from the State of Rio Grande do Sul, Brazil. Memórias Inst. Oswaldo Cruz 2002, 97, 829–834. [Google Scholar] [CrossRef] [Green Version]
- Dale, C.; Braga, M.V.; Galvão, C. Taxonomia integrada aplicada ao subcomplexo Triatoma rubrovaria no Estado do Rio grande do Sul, Brasil. In Atualidades em Medicina Tropical na America do Sul: Vetores; Oliveira, J., Alevi, K.C.C., Camargo, L.M.A., Meneguetii, D.U.O., Eds.; Stricto Sensu Editora: Rio Branco, Brasil, 2021; pp. 38–68. [Google Scholar]
- Succi, M.; Alevi, K.C.C.; Mendonça, O.O.; Bardella, V.B.; Rosa, J.A.; Azeredo-Oliveira, M.T.V. Spermatogenesis in Triatoma williami Galvão, Souza and Lima (1965) (Hemiptera, Triatominae). Invertebr. Reprod. Dev. 2013, 58, 124–127. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
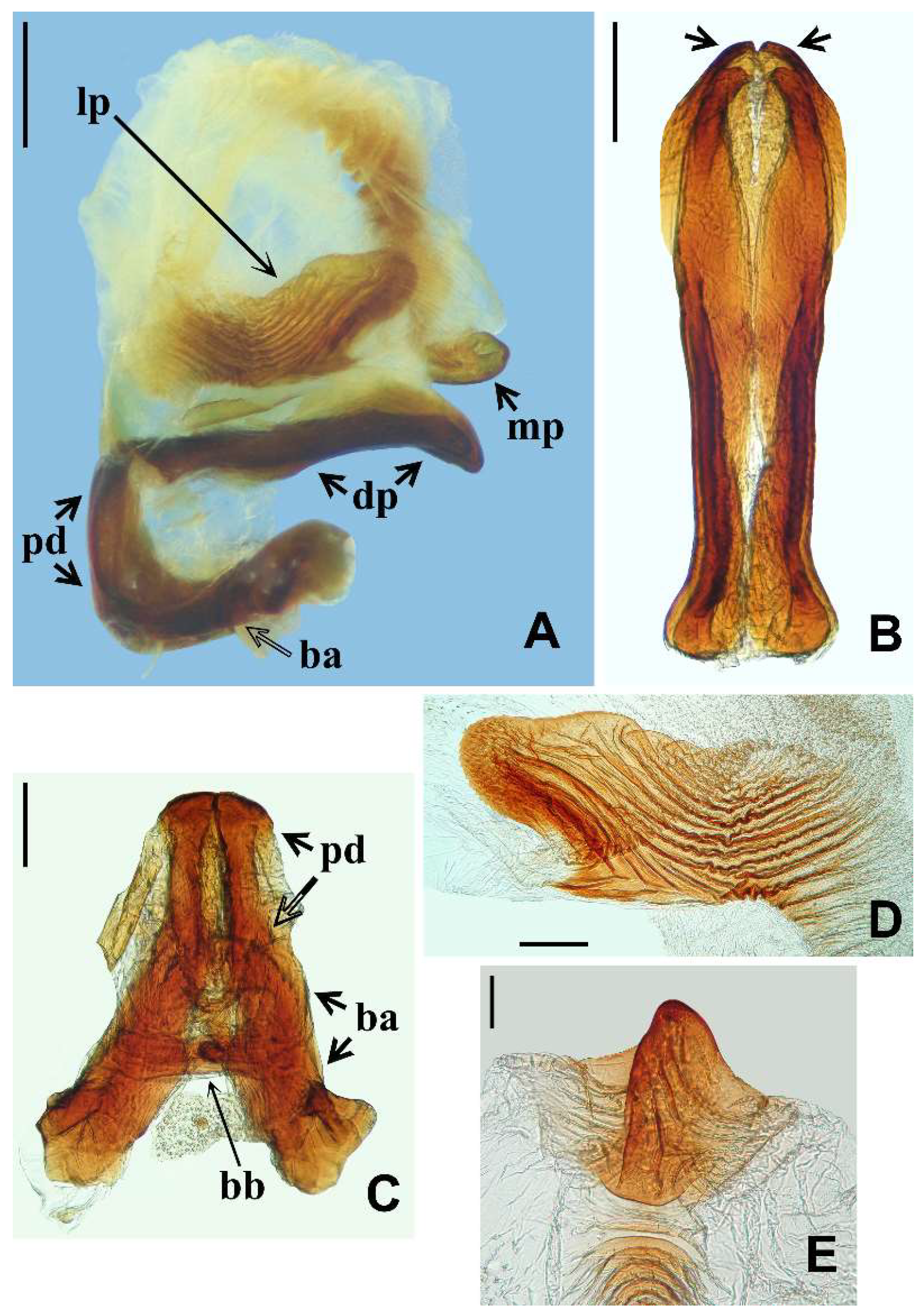{kind=link}
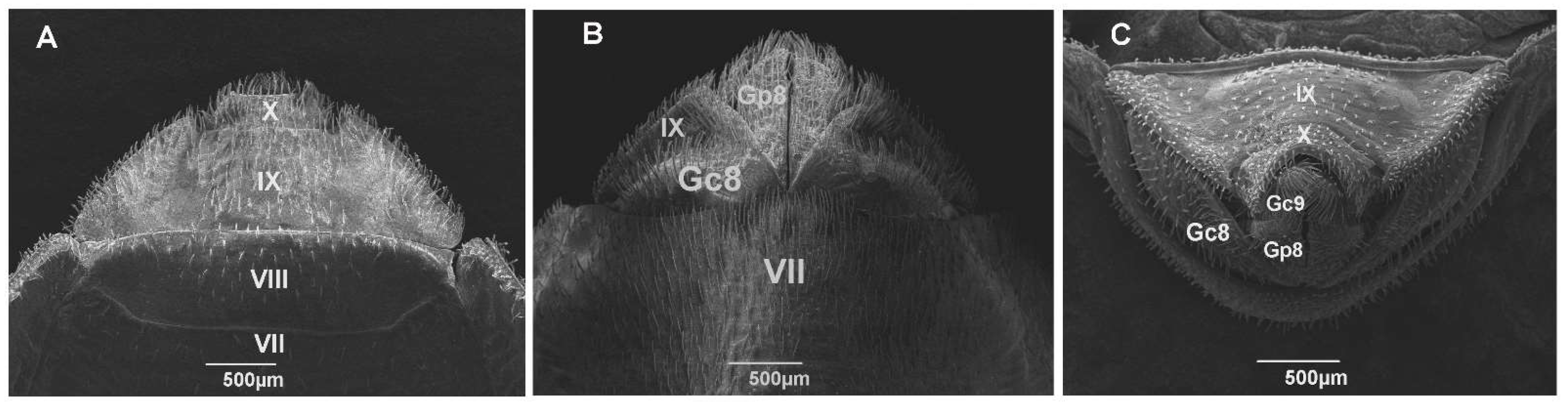{kind=link}
| Species | ID | Geographic Origin | Markers | |||
|---|---|---|---|---|---|---|
| 16S rRNA | COI | COII | Cytb | |||
| Triatoma brasiliensis | 40A | Curaçá, BA, Brazil | KC248986 | KC249319 | - | KC249240 |
| 40B | Curaçá, BA, Brazil | - | KC249320 | - | - | |
| - | - | - | ||||
| 172 | Tauá, CE, Brazil | - | KC249318 | - | - | |
| 41 | Sobral, CE, Brazil | KC248987 | - | - | KC249241 | |
| 174 | Tauá, CE, Brazil | KC248985 | - | KC249413 | KC249239 | |
| Triatoma guazu | - | - | - | - | - | |
| Barra do Garças, MT, Brazil | - | - | KC249440 | - | ||
| Barra do Garças, MT, Brazil | - | - | - | - | ||
| Barra do Garças, MT, Brazil | - | - | - | - | ||
| 29 | Barra do Garças, MT, Brazil | KC249013 | - | - | - | |
| Barra do Garças, MT, Brazil | - | KC608984 | - | KC608976 | ||
| Triatoma jatai | 03 | Paranã, TO, Brazil | KT601153 | KT601162 | - | - |
| 05 | Paranã, TO, Brazil | KT601154 | KT601163 | - | - | |
| 16 | Paranã, TO, Brazil | KT601155 | KT601164 | - | - | |
| Triatoma juazeirensis | 209 | Uiabi, BA, Brazil | KC249026 | KF826892 | - | KC249263 |
| CTA207 | Juazeiro, BA, Brazil | KF769453 | - | - | - | |
| Triatoma matogrossensis | - | AF324525 | - | - | - | |
| - | AF324526 | - | - | - | ||
| - | - | - | ||||
| Rio Verde de Mato Grosso, MS, Brazil | - | KC608985 | - | KC608972 | ||
| Rio Verde de Mato Grosso, MS, Brazil | - | - | - | KC608978 | ||
| 32 | Aquidauana, MS, Brazil | KC249039 | - | - | KC249271 | |
| 33 | Alegria, MT, Brazil | KC249040 | - | KC249460 | - | |
| 191 | - | - | KC249359 | KC249456 | KC249269 | |
| 192 | São Gabriel D’oeste, MS, Brazil | KC249037 | KC249360 | KC249457 | KC249270 | |
| Triatoma melanica | Haplotype G | - | - | KJ580492 | - | |
| Haplotype H | - | KJ580493 | ||||
| Haplotype I | - | KJ580494 | ||||
| Haplotype J | - | KJ580495 | ||||
| Urandi, BA, Brazil | KF769454 | - | ||||
| - | KC249041 | - | KC249461 | |||
| Triatoma sherlocki | 80 | KC249068 | - | KC249378 | ||
| EU489057 | EU489058 | |||||
| Triatoma williami | 04 | Barra do Garças, MT, Brazil | KT601156 | KT601165 | - | - |
| 36 | - | KC249089 | - | KC249493 | - | |
| Barra do Garças, MT, Brazil | - | - | - | - | ||
| Barra do Garças, MT, Brazil | - | KC608981 | ||||
| - | - | - | - | - | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
|---|---|---|---|---|---|---|---|---|
| 1.Triatoma guazu | - | |||||||
| 2.Triatoma williami | 0.002 | 0.000 | ||||||
| 3.Triatoma matogrossensis | 0.066 (0.065–0.068) | 0.063 (0.062–0.065) | 0.005 (0.000–0.008) | |||||
| 4.Triatoma jatai | 0.053 | 0.049 | 0.072 (0.071–0.074) | 0.000 | ||||
| 5.Triatoma brasiliensis | 0.054 (0.052–0.055) | 0.054 (0.052–0.055) | 0.084 (0.080–0.090) | 0.064 (0.061–0.068) | 0.003 (0.028–0.005) | |||
| 6.Triatoma juazeirensis | 0.063 (0.062–0.065) | 0.060 (0.059–0.062) | 0.082 (0.077–0.090) | 0.060 (0.058–0.061) | 0.021 (0.019–0.025) | 0.002 | ||
| 7.Triatoma melanica | 0.057 (0.055–0.059) | 0.054 (0.052–0.055) | 0.086 (0.083–0.090) | 0.054 (0.052–0.055) | 0.026 (0.022–0.028) | 0.033 (0.031–0.034) | 0.002 | |
| 8. Triatoma sherlocki | 0.057 (0.056–0.059) | 0.054 (0.053–0.056) | 0.084 (0.080–0.089) | 0.051 (0.049–0.052) | 0.022 (0.019–0.024) | 0.030 (0.028–0.031) | 0.018 (0.017–0.019) | 0.000 |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
|---|---|---|---|---|---|---|---|
| 1. Triatoma guazu | - | ||||||
| 2. Triatoma williami | 0.015 (0.000–0.015) | 0.015 | |||||
| 3. Triatoma matogrossensis | 0.123 (0.108–0.138) | 0.126 (0.108–0.145) | 0.020 (0.000–0.030) | ||||
| 4. Triatoma jatai | 0.090 | 0.088 (0.085–0.090) | 0.147 (0.137–0.157) | 0.000 | |||
| 5. Triatoma brasiliensis | 0.128 (0.125–0.131) | 0.137 (0.125–0.151) | 0.181 (0.161–0.201) | 0.191 (0.187–0.194) | 0.005 | ||
| 6. Triatoma melanica | 0.149 | 0.159 (0.149–0.169) | 0.191 (0.181–0.202) | 0.161 | 0.082 (0.079–0.085) | 0.000 | |
| 7. Triatoma juazeirensis | 0.126 | 0.126 | 0.133 (0.124–0.141) | 0.167 | 0.049 | 0.076 (0.042–0.087) | - |
| 1 | 2 | 3 | 4 | 5 | 6 | |
|---|---|---|---|---|---|---|
| 1.Triatoma guazu | - | |||||
| 2. Triatoma williami | 0.017 | - | ||||
| 3.Triatoma matogrossensis | 0.165 (0.162–0.167) | 0.165 (0.162–0.171) | 0.022 (0.000–0.032) | |||
| 4. Triatoma brasiliensis | 0.156 | 0.143 | 0.168 (0.168–0.187) | - | ||
| 5. Triatoma melanica | 0.166 | 0.156 | 0.245 (0.236–0.254) | 0.152 | - | |
| 6. Triatoma sherlocki | 0.164 | 0.155 | 0.206 (0.197–0.212) | 0.116 | 0.091 | - |
| 1 | 2 | 3 | 4 | 5 | 6 | |
|---|---|---|---|---|---|---|
| 1. Triatoma guazu | - | |||||
| 2. Triatoma williami | 0.009 | - | ||||
| 3. Triatoma matogrossensis | 0.153 | 0.140 | 0.006 (0–0.006) | |||
| 4. Triatoma brasiliensis | 0.198 | 0.196 | 0.181 (0.178–0.187) | 0.012 (0–0.012) | ||
| 5. Triatoma sherlocki | 0.198 | 0.204 | 0.213 | 0.141 (0.141–0.149) | - | |
| 6. Triatoma juazeirensis | 0.174 | 0.178 | 0.148 | 0.093 | 0.012 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira Correia, J.P.S.; Gil-Santana, H.R.; Dale, C.; Galvão, C. Triatoma guazu Lent and Wygodzinsky Is a Junior Synonym of Triatoma williami Galvão, Souza and Lima. Insects 2022, 13, 591. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13070591
Oliveira Correia JPS, Gil-Santana HR, Dale C, Galvão C. Triatoma guazu Lent and Wygodzinsky Is a Junior Synonym of Triatoma williami Galvão, Souza and Lima. Insects. 2022; 13(7):591. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13070591
Chicago/Turabian StyleOliveira Correia, João Paulo Sales, Hélcio Reinaldo Gil-Santana, Carolina Dale, and Cleber Galvão. 2022. "Triatoma guazu Lent and Wygodzinsky Is a Junior Synonym of Triatoma williami Galvão, Souza and Lima" Insects 13, no. 7: 591. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13070591