
Leaf UV-B Irradiation and Mycorrhizal Symbionts Affect Lettuce VOC Emissions and Defence Mechanisms, but Not Aphid Feeding Preferences
 , , , , , , ,
, , , , , , , 
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Plant Material and Treatments
2.3. Analysis of AMF Root Colonization
2.4. Stomatal Density
2.5. Histochemical Detection of Hydrogen Peroxide, Lipid Peroxidation, and Callose Deposition
2.6. Plant Volatile Organic Compounds (VOCs) Analysis
2.7. Feeding Preferences of Myzus persicae
2.8. Statistical Analysis
3. Results
3.1. Mycorrhizal Status of Experimental Plants
3.2. Stomatal Density
3.3. Hydrogen Peroxide, Lipid Peroxidation, and Callose Deposition
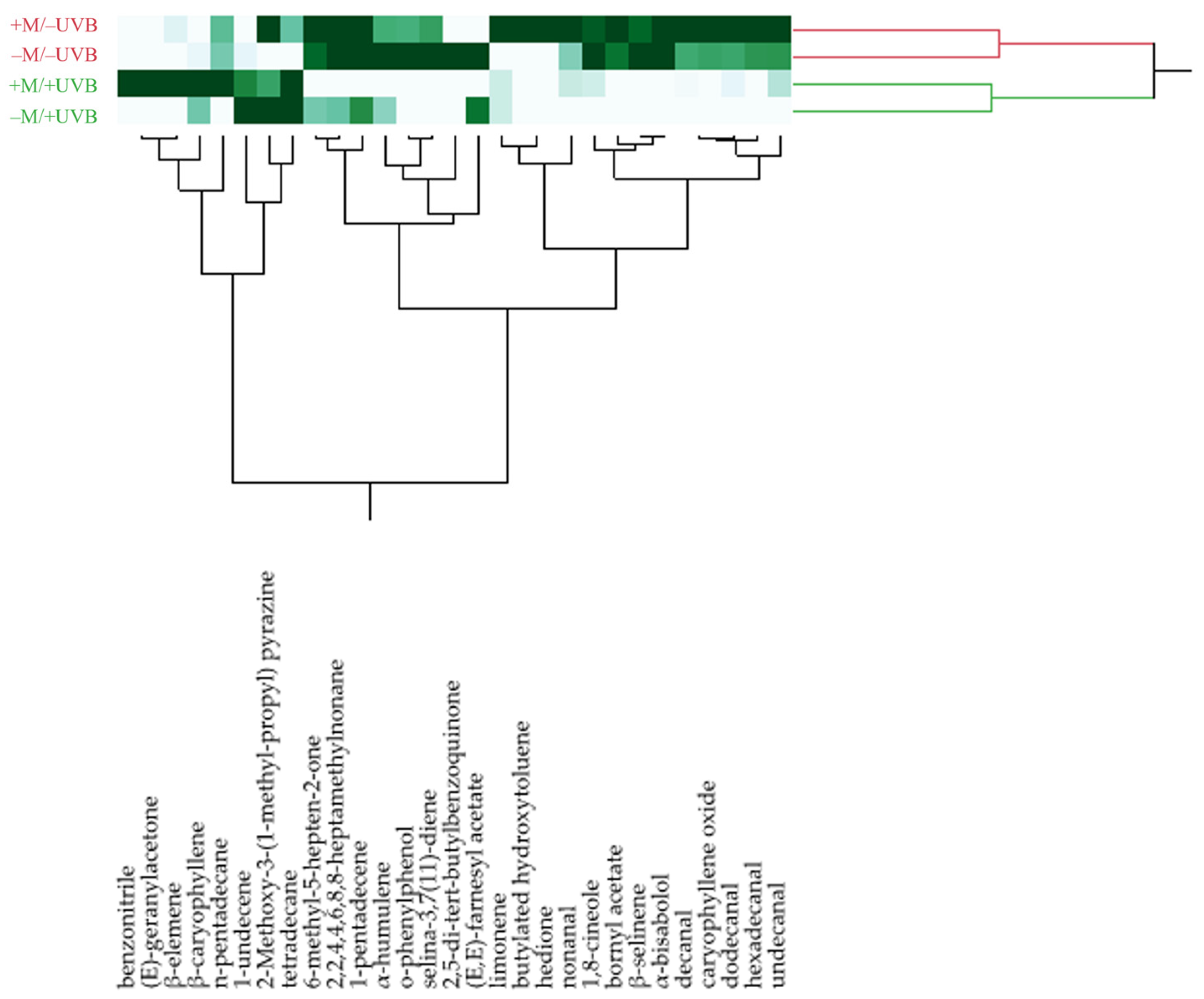
3.4. Plant Volatiles Profile
3.5. Feeding Preferences of Myzus persicae Aphids
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jenkins, G.I. Photomorphogenic responses to ultraviolet-B light. Plant Cell Environ. 2017, 40, 2544–2557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robson, T.M.; Klem, K.; Urban, O.; Jansen, M.A.K. Re-interpreting plant morphological responses to UV-B radiation. Plant Cell Environ. 2015, 38, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Bravo, R.; Klinkhamer, P.G.L.; Leiss, K.A. Interactive effects of UV-B light with abiotic factors on plant growth and chemistry, and their consequences for defense against arthropod herbivores. Front. Plant Sci. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiner, M.; Martínez-Abaigar, J.; Glaab, J.; Jansen, M. UV-B Induced Secondary Plant Metabolites. Opt. Photonik 2014, 9, 34–37. [Google Scholar] [CrossRef]
- Thomas, D.T.; Puthur, J.T. Amplification of abiotic stress tolerance potential in rice seedlings with a low dose of UV-B seed priming. Funct. Plant Biol. 2019, 46, 455–466. [Google Scholar] [CrossRef] [Green Version]
- Dhanya Thomas, T.T.; Dinakar, C.; Puthur, J.T. Effect of UV-B priming on the abiotic stress tolerance of stress-sensitive rice seedlings: Priming imprints and cross-tolerance. Plant Physiol. Biochem. 2020, 147, 21–30. [Google Scholar] [CrossRef]
- Thomas, T.T.D.; Puthur, J.T. UV-B priming enhances specific secondary metabolites in Oryza sativa (L.) empowering to encounter diverse abiotic stresses. Plant Growth Regul. 2020, 92, 169–180. [Google Scholar] [CrossRef]
- Sen, A.; Puthur, J.T. Halo- and UV-B priming-mediated drought tolerance and recovery in rice seedlings. Plant Stress 2021, 2, 100011. [Google Scholar] [CrossRef]
- Sen, A.; Challabathula, D.; Puthur, J.T. UV-B Priming of Oryza sativa Seeds Augments the Innate Tolerance Potential in a Tolerant Variety more Effectively Toward NaCl and PEG Stressors. J. Plant Growth Regul. 2021, 40, 1166–1180. [Google Scholar] [CrossRef]
- Klironomos, J.N.; Allen, M.F. UV-B-mediated changes on below-ground communities associated with the roots of Acer saccharum. Funct. Ecol. 1995, 9, 923–930. [Google Scholar] [CrossRef]
- Van de Staaij, J.; Rozema, J.; Van Beem, A.; Aerts, R. Increased solar UV-B radiation may reduce infection by arbuscular mycorrhizal fungi (AMF) in dune grassland plants: Evidence from five years of field exposure. Plant Ecol. 2001, 154, 169–177. [Google Scholar] [CrossRef]
- Regvar, M.; Bukovnik, U.; Likar, M.; Kreft, I. UV-B radiation affects flavonoids and fungal colonisation in Fagopyrum esculentum and F. tataricum. Cent. Eur. J. Biol. 2012, 7, 275–283. [Google Scholar] [CrossRef]
- Chen, M.; Arato, M.; Borghi, L.; Nouri, E.; Reinhardt, D. Beneficial services of arbuscular mycorrhizal fungi–from ecology to application. Front. Plant Sci. 2018, 9, 1270. [Google Scholar] [CrossRef] [PubMed]
- Welling, M.T.; Liu, L.; Rose, T.J.; Waters, D.L.E.; Benkendorff, K. Arbuscular mycorrhizal fungi: Effects on plant terpenoid accumulation. Plant Biol. 2016, 18, 552–562. [Google Scholar] [CrossRef]
- Avio, L.; Turrini, A.; Giovannetti, M.; Sbrana, C. Designing the ideotype mycorrhizal symbionts for the production of healthy food. Front. Plant Sci. 2018, 9, 1089. [Google Scholar] [CrossRef]
- Husna; Tuheteru, F.D.; Arif, A. The potential of arbuscular mycorrhizal fungi to conserve Kalappia celebica, an endangered endemic legume on gold mine tailings in Sulawesi, Indonesia. J. For. Res. 2021, 32, 675–682. [Google Scholar] [CrossRef]
- Agathokleous, E.; Kitao, M.; Komatsu, M.; Tamai, Y.; Harayama, H.; Koike, T. Single and combined effects of fertilization, ectomycorrhizal inoculation, and drought on container-grown Japanese larch seedlings. J. For. Res. 2023, in press. [Google Scholar] [CrossRef]
- Gehring, C.; Bennett, A. Mycorrhizal fungal-plant-insect interactions: The importance of a community approach. Environ. Entomol. 2009, 38, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Dicke, M.; Baldwin, I.T. TheThe evolutionary context for herbivore-induced plant volatiles: Beyond the “cry for help”. Trends Plant Sci. 2010, 15, 167–175. [Google Scholar] [CrossRef]
- Sharma, E.; Anand, G.; Kapoor, R. Terpenoids in plant and arbuscular mycorrhiza-reinforced defence against herbivorous insects. Ann. Bot. 2017, 119, 791–801. [Google Scholar] [CrossRef]
- Jaiswal, D.; Agrawal, S.B. Ultraviolet-B induced changes in physiology, phenylpropanoid pathway, and essential oil composition in two Curcuma species (C. caesia Roxb. and C. longa L.). Ecotoxicol. Environ. Saf. 2021, 208, 111739. [Google Scholar] [CrossRef] [PubMed]
- Rai, K.; Agrawal, S.B. Effect on essential oil components and wedelolactone content of a medicinal plant Eclipta alba due to modifications in the growth and morphology under different exposures of ultraviolet-B. Physiol. Mol. Biol. Plants 2020, 26, 773–792. [Google Scholar] [CrossRef] [PubMed]
- Copetta, A.; Lingua, G.; Berta, G. Effects of three AM fungi on growth, distribution of glandular hairs, and essential oil production in Ocimum basilicum L. var. Genovese. Mycorrhiza 2006, 16, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, R.; Chaudhary, V.; Bhatnagar, A.K. Effects of arbuscular mycorrhiza and phosphorus application on artemisinin concentration in Artemisia annua L. Mycorrhiza 2007, 17, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Binet, M.N.; van Tuinen, D.; Deprêtre, N.; Koszela, N.; Chambon, C.; Gianinazzi, S. Arbuscular mycorrhizal fungi associated with Artemisia umbelliformis Lam, an endangered aromatic species in Southern French Alps, influence plant P and essential oil contents. Mycorrhiza 2011, 21, 523–535. [Google Scholar] [CrossRef]
- Pistelli, L.; Ulivieri, V.; Giovanelli, S.; Avio, L.; Giovannetti, M.; Pistelli, L. Arbuscular mycorrhizal fungi alter the content and composition of secondary metabolites in Bituminaria bituminosa L. Plant Biol. 2017, 19, 926–933. [Google Scholar] [CrossRef]
- Velásquez, A.; Valenzuela, M.; Carvajal, M.; Fiaschi, G.; Avio, L.; Giovannetti, M.; D’Onofrio, C.; Seeger, M. The arbuscular mycorrhizal fungus Funneliformis mosseae induces changes and increases the concentration of volatile organic compounds in Vitis vinifera cv. Sangiovese leaf tissue. Plant Physiol. Biochem. 2020, 155, 437–443. [Google Scholar] [CrossRef]
- Velásquez, A.; Vega-Celedón, P.; Fiaschi, G.; Agnolucci, M.; Avio, L.; Giovannetti, M.; D’Onofrio, C.; Seeger, M. Responses of Vitis vinifera cv. Cabernet Sauvignon roots to the arbuscular mycorrhizal fungus Funneliformis mosseae and the plant growth-promoting rhizobacterium Ensifer meliloti include changes in volatile organic compounds. Mycorrhiza 2020, 30, 161–170. [Google Scholar] [CrossRef]
- Ninkovic, V.; Markovic, D.; Rensing, M. Plant volatiles as cues and signals in plant communication. Plant Cell Environ. 2021, 44, 1030–1043. [Google Scholar] [CrossRef]
- Turlings, T.C.J.; Erb, M. Tritrophic interactions mediated by herbivore-induced plant volatiles: Mechanisms, ecological relevance, and application potential. Annu. Rev. Entomol. 2018, 63, 433–452. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Wadhams, L.J.; Woodcock, C.M. Insect host location: A volatile situation. Trends Plant Sci. 2005, 10, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Bruce, T.J.A.; Pickett, J.A. Phytochemistry Perception of plant volatile blends by herbivorous insects—Finding the right mix. Phytochemistry 2011, 72, 1605–1611. [Google Scholar] [CrossRef] [PubMed]
- Barriere, V.; Lecompte, F.; Lescourret, F. Efficacy of pest and pathogen control, yield and quality of winter lettuce crops managed with reduced pesticide applications. Eur. J. Agron. 2015, 71, 34–43. [Google Scholar] [CrossRef]
- Macuphe, N.; Oguntibeju, O.O. the Physiology, Growth, and Antioxidant Activities of Extracts. Plants 2021, 10, 1178. [Google Scholar] [CrossRef] [PubMed]
- Sastry, K.S.; Mandal, B.; Hammond, J.; Scott, S.W.; Briddon, R.W.; Sastry, K.S.; Mandal, B.; Hammond, J.; Scott, S.W.; Briddon, R.W. Prunus persica (Peach). Encycl. Plant Viruses Viroids 2019, 1994–2011. [Google Scholar] [CrossRef]
- Canovai, R.; Benelli, G.; Ceragioli, T.; Lucchi, A.; Canale, A. Prey selection behaviour in the multicoloured Asian ladybird, Harmonia axyridis (Coleoptera: Coccinellidae). Appl. Entomol. Zool. 2019, 54, 213–222. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Meeus, S.; Van den Bulcke, J.; Wyffels, F. From leaf to label: A robust automated workflow for stomata detection. Ecol. Evol. 2020, 10, 9178–9191. [Google Scholar] [CrossRef]
- Bogani, P.; Calistri, E.; Biricolti, S.; Castiglione, M.R.; Turrini, A.; Giovannetti, M.; Buiatti, M. The rat glucocorticoid receptor integration in Nicotiana langsdorffii genome affects plant responses to abiotic stresses and to arbuscular mycorrhizal symbiosis. Plant Biotechnol. Rep. 2015, 9, 209–220. [Google Scholar] [CrossRef]
- Daudi, A.; O’Brien, J.A. Detection of hydrogen peroxide by DAB staining in Arabidopsis leaves. Bio-protocol 2012, 2, e263. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kobayashi, Y.; Matsumoto, H. Lipid peroxidation is an early symptom triggered by aluminum, but not the primary cause of elongation inhibition in Pea roots. Plant Physiol. 2001, 125, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kissoudis, C.; Sunarti, S.; Van De Wiel, C.; Visser, R.G.F.; Van Der Linden, C.G.; Bai, Y. Responses to combined abiotic and biotic stress in tomato are governed by stress intensity and resistance mechanism. J. Exp. Bot. 2016, 67, 5119–5132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Benedictis, M.; Brunetti, C.; Brauer, E.K.; Andreucci, A.; Popescu, S.C.; Commisso, M.; Guzzo, F.; Sofo, A.; Ruffini Castiglione, M.; Vatamaniuk, O.K.; et al. The Arabidopsis thaliana knockout mutant for phytochelatin synthase1 (cad1-3) is defective in callose deposition, bacterial pathogen defense and auxin content, but shows an increased stem lignification. Front. Plant Sci. 2018, 9, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; Volume 456, ISBN 978-1-932. [Google Scholar]
- Davies, N.W. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicon and Carbowax 20M phases. J. Chromatogr. A 1990, 503, 1–24. [Google Scholar] [CrossRef]
- Masada, Y. Analysis of Essential Oils by Gas Chromatography and Mass Spectrometry; John Wiley & Sons, Inc.: New York, NY, USA, 1976. [Google Scholar]
- Stenhagen, E.; Abrahamsson, S.; McLafferty, F.W. Registry of Mass Spectral Data; Wiley: New York, NY, USA, 1974. [Google Scholar]
- Swigar, A.A.; Silverstein, R.M. Monoterpenes, Aldrich Chemical Company; Sigma-Aldrich Corporation: Milwaukee, WI, USA, 1981; p. 130. [Google Scholar]
- Pieracci, Y.; Ascrizzi, R.; Pistelli, L.; Flamini, G. Comparison of the chemical and sensorial evaluation of dark chocolate bars. Appl. Sci. 2021, 11, 9964. [Google Scholar] [CrossRef]
- Ascrizzi, R.; Flamini, G.; Giusiani, M.; Stefanelli, F.; Deriu, V.; Chericoni, S. VOCs as fingerprints for the chemical profiling of hashish samples analyzed by HS-SPME/GC–MS and multivariate statistical tools. Forensic Toxicol. 2018, 36, 243–260. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST-palaeontological statistics, ver. 1.89. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef]
- Spinelli, F.; Cellini, A.; Marchetti, L.; Mudigere, K.; Piovene, C. Emission and function of volatile organic compounds in response to abiotic stress. Abiotic Stress Plants–Mech. Adapt. 2011, 394–397. [Google Scholar] [CrossRef]
- Eichholz, I.; Huyskens-Keil, S.; Keller, A.; Ulrich, D.; Kroh, L.W.; Rohn, S. UV-B-induced changes of volatile metabolites and phenolic compounds in blueberries (Vaccinium corymbosum L.). Food Chem. 2011, 126, 60–64. [Google Scholar] [CrossRef]
- Avio, L.; Sbrana, C.; Giovannetti, M.; Frassinetti, S. Arbuscular mycorrhizal fungi affect total phenolics content and antioxidant activity in leaves of oak leaf lettuce varieties. Sci. Hortic. 2017, 224, 265–271. [Google Scholar] [CrossRef]
- Baslam, M.; Garmendia, I.; Goicoechea, N. Arbuscular mycorrhizal fungi (AMF) improved growth and nutritional quality of greenhouse-grown Lettuce. J. Agric. Food Chem. 2011, 59, 5504–5515. [Google Scholar] [CrossRef] [PubMed]
- Baslam, M.; Pascual, I.; Sánchez-Díaz, M.; Erro, J.; García-Mina, J.M.; Goicoechea, N. Improvement of nutritional quality of greenhouse-grown lettuce by arbuscular mycorrhizal fungi is conditioned by the source of phosphorus nutrition. J. Agric. Food Chem. 2011, 59, 11129–11140. [Google Scholar] [CrossRef]
- Baslam, M.; Garmendia, I.; Goicoechea, N. The arbuscular mycorrhizal symbiosis can overcome reductions in yield and nutritional quality in greenhouse-lettuces cultivated at inappropriate growing seasons. Sci. Hortic. 2013, 164, 145–154. [Google Scholar] [CrossRef]
- Goicoechea, N.; Garmendia, I.; Fabbrin, E.G.; Bettoni, M.M.; Palop, J.A.; Sanmartín, C. Selenium fertilization and mycorrhizal technology may interfere in enhancing bioactive compounds in edible tissues of lettuces. Sci. Hortic. 2015, 195, 163–172. [Google Scholar] [CrossRef]
- Vangelisti, A.; Natali, L.; Bernardi, R.; Sbrana, C.; Turrini, A.; Hassani-Pak, K.; Hughes, D.; Cavallini, A.; Giovannetti, M.; Giordani, T. Transcriptome changes induced by arbuscular mycorrhizal fungi in sunflower (Helianthus annuus L.) roots. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Rapparini, F.; Llusià, J.; Peñuelas, J. Effect of arbuscular mycorrhizal (AM) colonization on terpene emission and content of Artemisia annua L. Plant Biol. 2007, 9, e20–e32. [Google Scholar] [CrossRef] [Green Version]
- Shrivastava, G.; Ownley, B.H.; Augé, R.M.; Toler, H.; Dee, M.; Vu, A.; Köllner, T.G.; Chen, F. Colonization by arbuscular mycorrhizal and endophytic fungi enhanced terpene production in tomato plants and their defense against a herbivorous insect. Symbiosis 2015, 65, 65–74. [Google Scholar] [CrossRef]
- Dai, Q.; Peng, S.; Chavez, A.Q.; Vergara, B.S. Effects of UVB radiation on stomatal density and opening in rice (Oryza sativa L.). Ann. Bot. 1995, 76, 65–70. [Google Scholar] [CrossRef]
- Gitz, D.C.; Liu-Gitz, L.; Britz, S.J.; Sullivan, J.H. Ultraviolet-B effects on stomatal density, water-use efficiency, and stable carbon isotope discrimination in four glasshouse-grown soybean (Glyicine max) cultivars. Environ. Exp. Bot. 2005, 53, 343–355. [Google Scholar] [CrossRef]
- Chitarra, W.; Pagliarani, C.; Maserti, B.; Lumini, E.; Siciliano, I.; Cascone, P.; Schubert, A.; Gambino, G.; Balestrini, R.; Guerrieri, E. Insights on the impact of arbuscular mycorrhizal symbiosis on tomato tolerance to water stress. Plant Physiol. 2016, 171, 1009–1023. [Google Scholar] [CrossRef] [PubMed]
- Pantaleo, V.; Vitali, M.; Boccacci, P.; Miozzi, L.; Cuozzo, D.; Chitarra, W.; Mannini, F.; Lovisolo, C.; Gambino, G. Novel functional microRNAs from virus-free and infected Vitis vinifera plants under water stress. Sci. Rep. 2016, 6, 20167. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.W.; Michniewicz, M.; Bergmann, D.C.; Wang, Z.Y. Brassinosteroid regulates stomatal development by GSK3-mediated inhibition of a MAPK pathway. Nature 2012, 482, 419–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.N.; Wu, Q.S.; Kuča, K. Unravelling the role of arbuscular mycorrhizal fungi in mitigating the oxidative burst of plants under drought stress. Plant Biol. 2021, 23, 50–57. [Google Scholar] [CrossRef]
- Wargent, J.J.; Jordan, B.R. From ozone depletion to agriculture: Understanding the role of UV radiation in sustainable crop production. New Phytol. 2013, 197, 1058–1076. [Google Scholar] [CrossRef]
- Czarnocka, W.; Karpiński, S. Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free Radic. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Gange, A.C. Insect-mycorrhizal interactions, patterns, processes and consequences. Ecol. Communities Plant Mediat. Indirect Interact. Webs 2007, 124–144. [Google Scholar] [CrossRef]
- Kuhlmann, F.; Müller, C. UV-B impact on aphid performance mediated by plant quality and plant changes induced by aphids. Plant Biol. 2010, 12, 676–684. [Google Scholar] [CrossRef]
- Kuhlmann, F.; Müller, C. Impacts of ultraviolet radiation on interactions between plants and herbivorous insects: A chemo-ecological perspective. Prog. Bot. 2010, 72, 305–347. [Google Scholar]
- Qi, J.; Zhang, M.; Lu, C.; Hettenhausen, C.; Tan, Q.; Cao, G.; Zhu, X.; Wu, G.; Wu, J. Ultraviolet-B enhances the resistance of multiple plant species to lepidopteran insect herbivory through the jasmonic acid pathway. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinen, R.; Biere, A.; Harvey, J.A.; Bezemer, T.M. Effects of Soil Organisms on Aboveground Plant-Insect Interactions in the Field: Patterns, Mechanisms and the Role of Methodology. Front. Ecol. Evol. 2018, 6, 106. [Google Scholar] [CrossRef]
- Kaur, J.; Chavana, J.; Soti, P.; Racelis, A.; Kariyat, R. Arbuscular mycorrhizal fungi (AMF) influences growth and insect community dynamics in Sorghum-sudangrass (Sorghum x drummondii). Arthropod. Plant. Interact. 2020, 14, 301–315. [Google Scholar] [CrossRef]
- Kuśnierczyk, A.; Winge, P.; Midelfart, H.; Armbruster, W.S.; Rossiter, J.T.; Bones, A.M. Transcriptional responses of Arabidopsis thaliana ecotypes with different glucosinolate profiles after attack by polyphagous Myzus persicae and oligophagous Brevicoryne brassicae. J. Exp. Bot. 2007, 58, 2537–2552. [Google Scholar] [CrossRef] [PubMed]
- Verheggen, F.J.; Fagel, Q.; Heuskin, S.; Lognay, G.; Francis, F.; Haubruge, E. Electrophysiological and behavioral responses of the multicolored Asian lady beetle, Harmonia axyridis pallas, to sesquiterpene semiochemicals. J. Chem. Ecol. 2007, 33, 2148–2155. [Google Scholar] [CrossRef]
- Verheggen, F.J.; Arnaud, L.; Bartram, S.; Gohy, M.; Haubruge, E. Aphid and plant volatiles induce oviposition in an aphidophagous hoverfly. J. Chem. Ecol. 2008, 34, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Vandermoten, S.; Mescher, M.C.; Francis, F.; Haubruge, E.; Verheggen, F.J. Aphid alarm pheromone: An overview of current knowledge on biosynthesis and functions. Insect Biochem. Mol. Biol. 2012, 42, 155–163. [Google Scholar] [CrossRef]
- Ahmed, N.; Darshanee, H.L.C.; Khan, I.A.; Zhang, Z.F.; Liu, T.X. Host selection behavior of the green peach aphid, Myzus persicae, in response to volatile organic compounds and nitrogen contents of cabbage cultivars. Front. Plant Sci. 2019, 10, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, F.; Vandermoten, S.; Verheggen, F.; Lognay, G.; Haubruge, E. Is the (E)-β-farnesene only volatile terpenoid in aphids? J. Appl. Entomol. 2005, 129, 6–11. [Google Scholar] [CrossRef]
- Klaiber, J.; Najar-Rodriguez, A.J.; Dialer, E.; Dorn, S. Elevated carbon dioxide impairs the performance of a specialized parasitoid of an aphid host feeding on Brassica plants. Biol. Control 2013, 66, 49–55. [Google Scholar] [CrossRef]
- Masui, N.; Agathokleous, E.; Mochizuki, T.; Tani, A.; Matsuura, H.; Koike, T. Ozone disrupts the communication between plants and insects in urban and suburban areas: An updated insight on plant volatiles. J. For. Res. 2021, 32, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Masui, N.; Agathokleous, E.; Tani, A.; Matsuura, H.; Koike, T. Plant-insect communication in urban forests: Similarities of plant volatile compositions among tree species (host vs. non-host trees) for alder leaf beetle Agelastica coerulea. Environ. Res. 2022, 204, 111996. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | l.r.i. 1 | Class | Relative Abundance (%) ± SE (n = 4) | |||
|---|---|---|---|---|---|---|
| +M/−UVB | +M/+UVB | −M/−UVB | −M/+UVB | |||
| Benzonitrile | 986 | nt | 2 | 1.4 ± 0.56 | - | - |
| 6-methyl-5-hepten-2-one | 986 | nt | 1.3 ± 0.23 | - | 1.1 ± 0.22 | 0.7 ± 0.25 |
| Limonene | 1029 | mh | 2.6 ± 0.77 | 1.5 ± 0.5 | 1.4 ± 0.65 | 1.5 ± 0.44 |
| 1,8-cineole | 1032 | om | 1.8 ± 0.96 | 0.7 ± 0.25 | 1.9 ± 1.07 | 0.3 ± 0.32 |
| 1-undecene | 1091 | nt | 20.1 ± 5.55 | 48.0 ± 7.78 | 25.0 ± 4.36 | 56.6 ± 14.90 |
| Nonanal | 1105 | nt | 5.2 ± 0.71 | 2.6 ± 1.14 | 3.1 ± 0.88 | 2.0 ± 0.85 |
| 2-methoxy-3-(1-methyl-propyl) pyrazine | 1175 | pyr | 1.4 ± 0.79 | 1.2 ± 1.17 | 0.8 ± 0.84 | 1.3 ± 1.34 |
| Decanal | 1206 | nt | 27.4 ± 7.33 | 8.5 ± 3.90 | 19.2 ± 8.00 | 8.0 ± 4.87 |
| bornyl acetate | 1286 | om | 0.7 ± 0.06 | - | 0.5 ± 0.31 | - |
| Undecanal | 1305 | nt | 4.2 ± 0.27 | 2.1 ± 0.65 | 3.4 ± 0.55 | 1.2 ± 1.18 |
| 2,2,4,4,6,8,8-heptamethylnonane | 1322 | nt | 1.2 ± 0.26 | - | 1.2 ± 0.49 | 0.8 ± 0.52 |
| β-elemene | 1392 | sh | 1.5 ± 0.39 | 2.6 ± 0.78 | 1.5 ± 0.62 | 1.4 ± 0.23 |
| n-tetradecane | 1400 | nt | 1.1 ± 0.2 | 1.2 ± 0.13 | 1 ± 0.15 | 1.2 ± 0.17 |
| Dodecanal | 1409 | nt | 2.6 ± 0.18 | 1.6 ± 0.38 | 2.2 ± 0.43 | 1.5 ± 0.48 |
| β-caryophyllene | 1419 | sh | 8.6 ± 2.84 | 15.5 ± 8.97 | 9.4 ± 2.58 | 10.7 ± 5.34 |
| I-geranylacetone | 1453 | ac | - | 8.3 ± 3.24 | - | - |
| α-humulene | 1453 | sh | 11.1 ± 1.29 | - | 17.8 ± 5.75 | 7.1 ± 2.28 |
| β-selinene | 1486 | sh | 0.8 ± 0.14 | - | 0.9 ± 0.24 | - |
| 1-pentadecene | 1493 | nt | 5.0 ± 1.85 | 3.5 ± 0.68 | 5.2 ± 1.35 | 4.7 ± 0.85 |
| selina-3,7(11)-diene | 1530 | sh | 0.8 ± 0.18 | - | 1.4 ± 0.44 | - |
| caryophyllene oxide | 1582 | os | 0.5 ± 0.18 | - | 0.4 ± 0.11 | - |
| Hedione | 1646 | nt | 0.6 ± 0.31 | - | - | - |
| α-bisabolol | 1685 | os | 0.5 ± 0.16 | - | 0.5 ± 0.1 | - |
| hexadecanal | 1816 | nt | 0.6 ± 0.14 | - | 0.4 ± 0.08 | - |
| (E,E)-farnesyl acetate | 1843 | os | - | - | 1.2 ± 0.43 | 1.0 ± 0.96 |
| Chemical Classes | l.r.i. 1 | Relative Abundance (%) ± SE (n = 4) | ||||
| +M/−UVB | +M/+UVB | −M/−UVB | −M/+UVB | |||
| Monoterpene hydrocarbons (mh) | 2.6 ± 0.77 | 1.5 ± 0.5 | 1.4 ± 0.65 | 1.5 ± 0.44 | ||
| Oxygenated monoterpenes (om) | 2.5 ± 0.96 | 0.7 ± 0.25 | 2.4 ± 0.94 | 0.3 ± 0.32 | ||
| Sesquiterpene hydrocarbons (sh) | 22.7 ± 3.75 | 18.2 ± 9.62 | 30.9 ± 6.49 | 19.2 ± 5.9 | ||
| Oxygenated sesquiterpenes (os) | 1.0 ± 0.34 | - | 2.1 ± 0.52 | 1.0 ± 0.96 | ||
| Other non-terpene derivatives (nt) | 69.9 ± 4.17 | 70.1 ± 7.48 | 62.4 ± 7.5 | 76.7 ± 7.18 | ||
| Pyrazines (pyr) | 1.4 ± 0.79 | 1.2 ± 1.17 | 0.8 ± 0.84 | 1.3 ± 1.34 | ||
| Apocarotenoids (ac) | - B | 8.3 ± 3.24 A | - B | - B | ||
| Total identified (%) | 100.0 ± 0.01 | 100.0 ± 0.00 | 100.0 ± 0.01 | 100.0 ± 0.00 | ||
| Compound | Average Dissimilarity | Individual Contribution (%) | Cumulative Contribution (%) |
|---|---|---|---|
| 1-undecene | 15.88 | 32.11 | 32.11 |
| Decanal | 8.695 | 17.58 | 49.69 |
| α-humulene | 5.582 | 11.29 | 60.98 |
| β-caryophyllene | 5.384 | 10.89 | 71.86 |
| (E)-geranyl acetone | 2.077 | 4.201 | 76.06 |
| 1-pentadecene | 1.388 | 2.806 | 78.87 |
| nonanal | 1.282 | 2.593 | 81.46 |
| undecanal | 1.263 | 2.555 | 84.02 |
| 2-methoxy-3-(1-methyl-propyl) pyrazine | 0.9025 | 1.825 | 85.84 |
| 1,8-cineole | 0.7889 | 1.595 | 87.44 |
| limonene | 0.6703 | 1.355 | 88.79 |
| β-elemene | 0.5969 | 1.207 | 90 |
| 2,2,4,4,6,8,8-heptamethylnonane | 0.5533 | 1.119 | 91.12 |
| dodecanal | 0.5377 | 1.087 | 92.2 |
| selina-3,7(11)-diene | 0.5375 | 1.087 | 93.29 |
| Compound | Average Dissimilarity | Individual Contribution (%) | Cumulative Contribution (%) |
|---|---|---|---|
| 1-undecene | 12.37 | 29.18 | 29.18 |
| decanal | 7.374 | 17.4 | 46.59 |
| β-caryophyllene | 5.208 | 12.29 | 58.88 |
| α-humulene | 4.694 | 11.08 | 69.96 |
| (E)-geranyl acetone | 2.077 | 4.903 | 74.86 |
| 1-pentadecene | 1.342 | 3.167 | 78.03 |
| nonanal | 1.187 | 2.802 | 80.83 |
| undecanal | 1.047 | 2.47 | 83.3 |
| 2-methoxy-3-(1-methyl-propyl) pyrazine | 0.8819 | 2.081 | 85.38 |
| 1,8-cineole | 0.7098 | 1.675 | 87.06 |
| limonene | 0.6746 | 1.592 | 88.65 |
| β-elemene | 0.615 | 1.451 | 90.1 |
| (E,E)-farnesyl acetate | 0.5462 | 1.289 | 91.39 |
| dodecanal | 0.4752 | 1.121 | 92.51 |
| 2,2,4,4,6,8,8-heptamethylnonane | 0.4731 | 1.117 | 93.63 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeni, V.; Grassi, A.; Santin, M.; Ricciardi, R.; Pieracci, Y.; Flamini, G.; Di Giovanni, F.; Marmugi, M.; Agnolucci, M.; Avio, L.; et al. Leaf UV-B Irradiation and Mycorrhizal Symbionts Affect Lettuce VOC Emissions and Defence Mechanisms, but Not Aphid Feeding Preferences. Insects 2023, 14, 20. https://0-doi-org.brum.beds.ac.uk/10.3390/insects14010020
Zeni V, Grassi A, Santin M, Ricciardi R, Pieracci Y, Flamini G, Di Giovanni F, Marmugi M, Agnolucci M, Avio L, et al. Leaf UV-B Irradiation and Mycorrhizal Symbionts Affect Lettuce VOC Emissions and Defence Mechanisms, but Not Aphid Feeding Preferences. Insects. 2023; 14(1):20. https://0-doi-org.brum.beds.ac.uk/10.3390/insects14010020
Chicago/Turabian StyleZeni, Valeria, Arianna Grassi, Marco Santin, Renato Ricciardi, Ylenia Pieracci, Guido Flamini, Filippo Di Giovanni, Margherita Marmugi, Monica Agnolucci, Luciano Avio, and et al. 2023. "Leaf UV-B Irradiation and Mycorrhizal Symbionts Affect Lettuce VOC Emissions and Defence Mechanisms, but Not Aphid Feeding Preferences" Insects 14, no. 1: 20. https://0-doi-org.brum.beds.ac.uk/10.3390/insects14010020