
Neural Correlates of Aberrant Salience and Source Monitoring in Schizophrenia and At-Risk Mental States—A Systematic Review of fMRI Studies


Abstract
:1. Introduction
1.1. Role of Aberrant Salience in Psychosis
1.2. Role of Source Monitoring in Psychosis
1.3. Possible Integration of Aberrant Salience and Source Monitoring Deficits and Aims of this Review
2. Materials and Methods
2.1. Literature Search Strategy
2.2. Literature Selection
2.3. Computation, Interpretation of Results, and Bias Assessment
3. Results
3.1. Results of Systematic Review
3.2. Aberrant Salience
3.2.1. Behavioral Results for Aberrant Salience
3.2.2. Behavioral Results for Adaptive Salience
3.2.3. Neuroimaging Results for Group Differences in Salience and Reward Prediction
3.2.4. Additional Neuroimaging Results

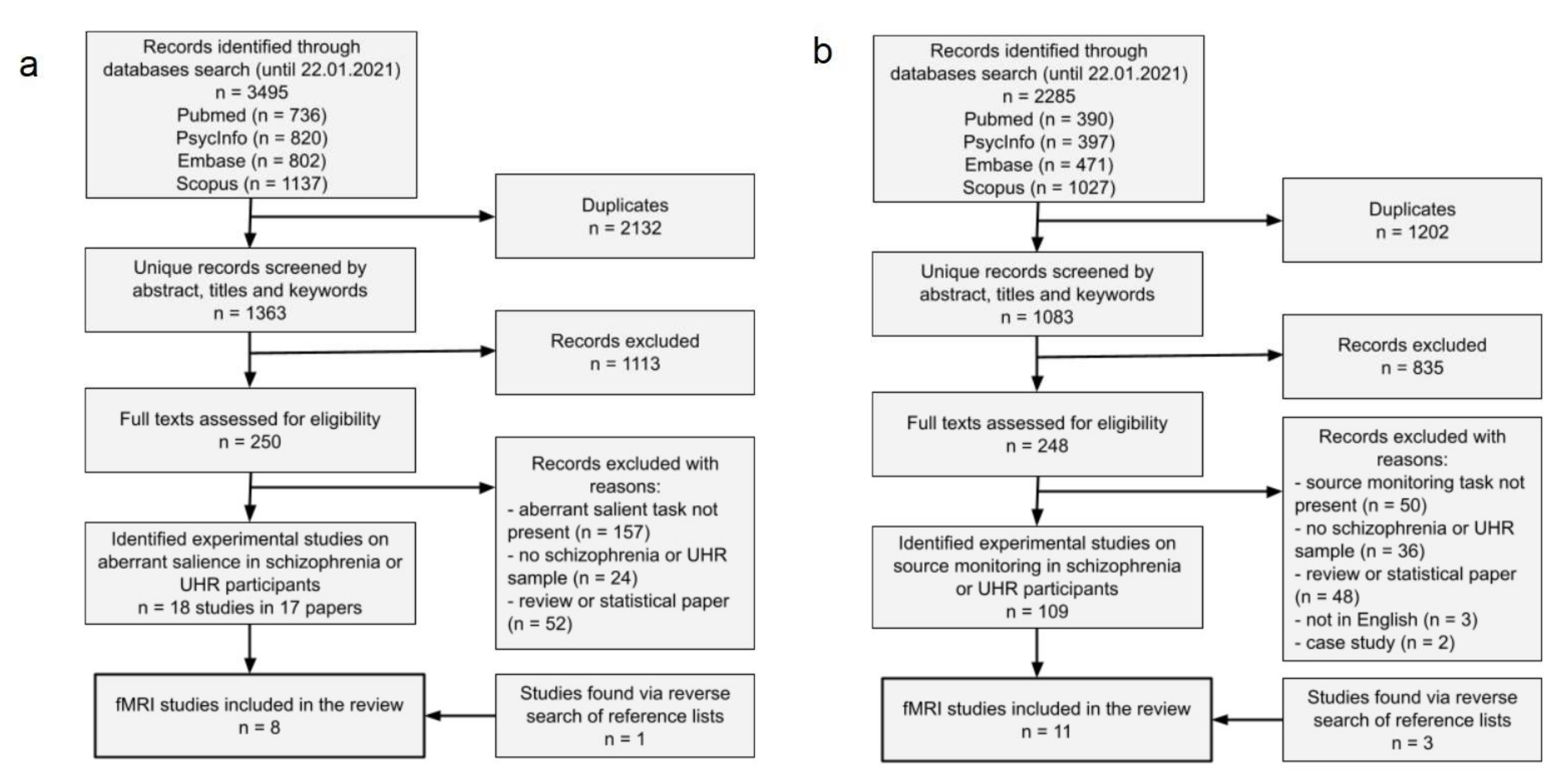{kind=link}
{kind=link}
| Study | Study Type | Sample Sizes | Clinical Sample(s) Characteristics | Experimental Task | Aberrant Salience Behavioural Results | Main Neuroimaging Results—Group Comparisons | Additional Neuroimaging Results |
|---|---|---|---|---|---|---|---|
| Esslinger et al., 2012 [59] | A comparative study of FEP and HC; an fMRI study | FEP = 27 HC = 27 | Convenience sample from an admissions centre for a mental health hospital; never medicated | Implicit Salience Attribution Task (famous and non-famous faces) |
|
|
|
| Katthagen et al., 2018 [55] | A comparative study of SCH and HC; an fMRI study | SCH = 42 HC = 42 | Convenience sample from inpatient and outpatient units; all participants had antipsychotic medication | Implicit Salience Paradigm |
| Not applicable |
|
| Pankow et al., 2016 [31] | A comparative study of SCH, subclinical delusions and HC; an fMRI study | SCH = 29 subclinical delusions = 24 HC = 50 | SCH: a convenience sample from a hospital department of psychiatry and psychotherapy. Most of the patients were medicated; subclinical delusions: people with results in 4Q of PDI from a large internet sample | Salience Attribution Test |
| Not applicable |
|
| Walter et al., 2016 [36] | A comparative study of SSD with lower and higher levels of positive symptoms; an fMRI study | SSD with higher positive symptoms = 21 SSD with lower positive symptoms = 21 | A convenience sample of patients with FEP and chronic SCH patients with a history of violent offense; most of the patients were medicated | Salience Attribution Test |
|
|
|
| Smieskova et al., 2015 [56] | A comparative study of ARMS, FEP and HC; an fMRI study | FEP with medication = 12 FEP without medication = 17 ARMS = 34 HC = 19 | A convenience sample of patients with FEP (medicated and unmedicated) and ARMS patients | Salience Attribution Test |
|
|
|
| Roiser et al., 2013 [33] | Comparative study of UHR and HC; an fMRI and PET study | UHR = 18 HC = 18 | A convenience sample; 2 participants with antipsychotic medication (authors checked that data from these participants did not alter the results) | Salience Attribution Test |
|
|
|
| Schmidt et al., 2017 [34] | A longitudinal study of UHR and HC; an fMRI study | UHR = 23 HC = 13 | A convenience sample from a clinical service for people at high risk of psychosis | Salience Attribution Test |
|
|
|
| Winton-Brown et al., 2017 [27] | A comparative study of UHR and HC; an fMRI study | UHR = 29 HC = 32 | A convenience sample from a clinical service for people at high risk of psychosis | Salience Integration Task |
|
|
|
3.3. Source Monitoring
3.3.1. Behavioral Results for Source Monitoring
| Study | Study Type | Sample Size | Sample Characteristics | Experimental Task | Behavioural Results | Main Neuroimaging Results—Group Comparison | Additional Neuroimaging Results |
|---|---|---|---|---|---|---|---|
| Allen et al., 2007 [42] | Comparative study of SCH sample and HC, an fMRI study | SCH = 20 (division on AVH = 10; non-AVH = 10) HC = 11 | A convenience sample of psychiatric patients recruited through the South London and Maudsley National Health Service Trust | Source monitoring task (discrimination between self/researcher and distorted/undistorted speech ) |
|
| - |
| Garrison et al., 2017 [62] | Comparative study of SCH sample and HC, an fMRI study | SCH = 20 HC = 20 | A convenience sample | Reality monitoring task (discrimination between perceived/imagined and self/researcher word-pairs) |
|
|
|
| Kambeitz-Ilankovic et al., 2013 [69] | Comparative study of FEP and HC; an fMRI study | FEP = 20 HC = 20 | A convenience sample of psychiatric patients recruited through Maudsley National Health Service Trust | Source attribution task (discrimination between self/other speech, ambiguity: undistorted/distorted speech and validity: valid/invalid cues) |
|
|
|
| Kumari et al., 2008 [63] | Comparative study of SCH and HC; an fMRI study | SCH = 63 HC = 20 | A convenience sample | Self-monitoring task (discrimination between self/other speech and undistorted/distorted speech) |
|
|
|
| Mechelli et al., 2007 [61] | Comparative study of SCH and HC; an fMRI study | SCH = 20 (division on AVH = 11; non-AVH = 10) HC = 10 | A convenience sample of psychiatric patients recruited via the South London and Maudsley National Healthy Trust (SLAM) | Source monitoring task (discrimination between self/researcher and disorted/undistorted speech ) |
|
| - |
| Stephan-Otto et al., 2017 [70] | Comparative study of SCH sample and HC, an fMRI study | SCH = 23 HC = 26 | A convenience sample of psychiatric patients recruited from the Parc Sanitari Sant Joan de Deu network of mental health services in Barcelona | Reality monitoring task (discrimination between pictures and picture labels) |
|
|
|
| Subramaniam et al., 2012 [66] | An intervention study of SCH; an fMRI study | SCH (division into AT: active training and CG: computer games training groups) = 31 (15 + 14 and 2 dropouts at baseline; 13 and 12 returned 6 month later) HC = 15 | A convenience sample of psychiatric patients recruited from community mental health centres and outpatient clinics | Reality monitoring task (discrimination between self-generated or externally presented words) | Baseline assessment:
|
Baseline assessment:
|
|
| Subramaniam et al., 2017 [67] | A comparative study of SCH; an fMRI study | SCH = 20 HC = 20 | A convenience sample of psychiatric patients recruited from a double-blind randomized clinical trial of cognitive training in schizophrenia (ClinicalTrials.gov NCT02105779). | Reality monitoring task (discrimination between self-generated or externally presented words) |
|
|
|
| Thoresen et al., 2014 [64] | Comparative study of SCH sample and HC, an fMRI study | SCH = 19 HC = 20 | A convenience sample | Reality monitoring task (discrimination between presented or an imagined object/scene) |
|
|
|
| Vinogradov et al., 2008 [65] | Comparative study of SCH sample and HC, an fMRI study | SCH = 8 HC = 8 | A convenience sample | Source memory task (discrimination between the self-generated nouns and presented nouns in the sentence completion paradigm) |
|
|
|
| Wang et al., 2011 [68] | Comparative study of SCH sample and HC, an fMRI study | SCH = 23 HC = 33 | A convenience sample of psychiatric patients recruited from from psychiatric hospitals and community health agencies in and around Vancouver, BritishColumbia, Canada | Self–other source monitoring task (discrimination between self-generated and other-generated words when solving a puzzle) |
|
| - |
3.3.2. Neuroimaging Results for Group Differences in Source Monitoring
3.3.3. Additional Neuroimaging Results
4. Discussion
4.1. Behavioral Results for Aberrant Salience
4.2. Neuroimaging Results for Aberrant Salience
4.3. Behavioral Results for Source Monitoring
4.4. Neuroimaging Results for Source Monitoring
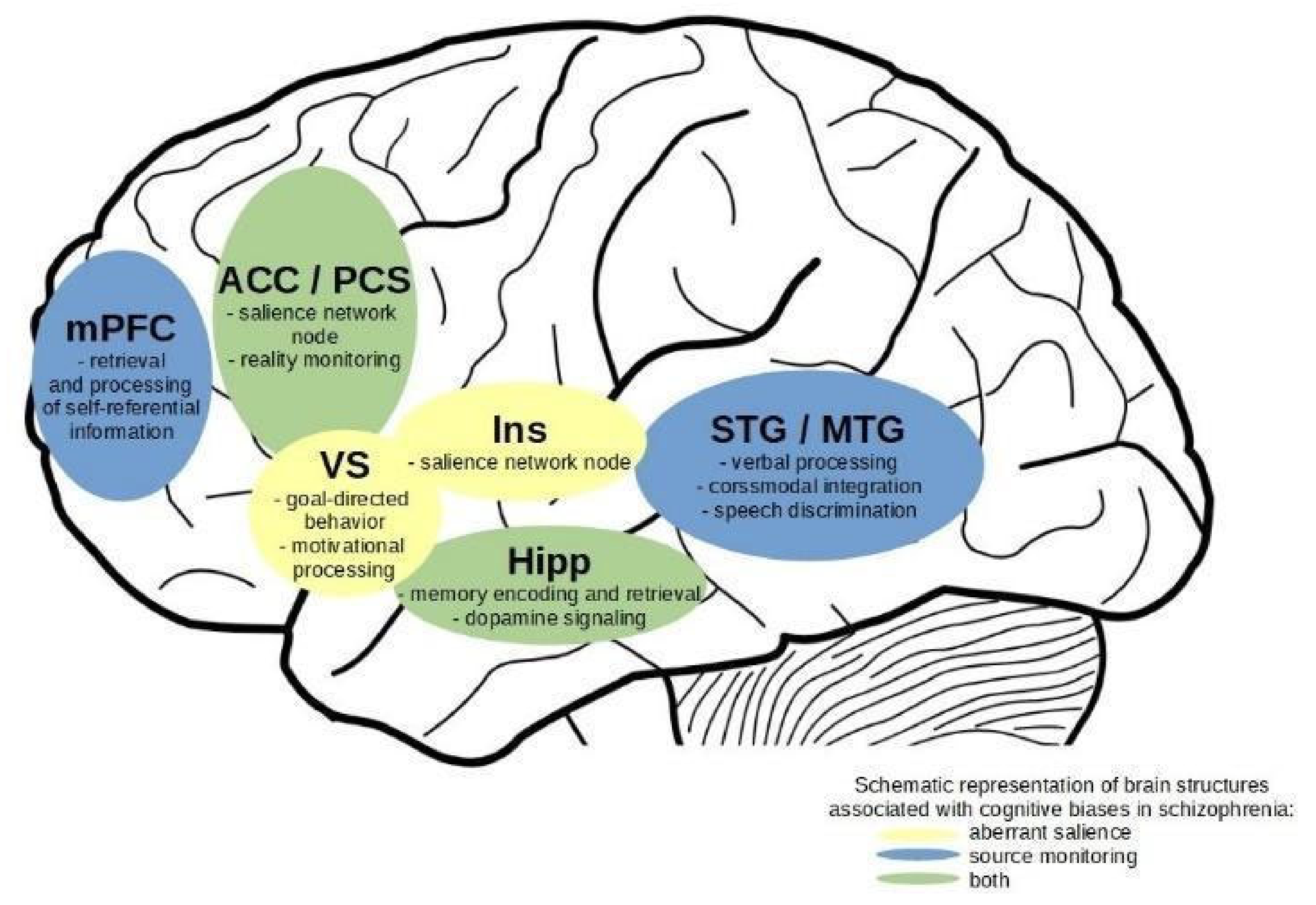
4.5. Overlaps in Neuroimaging Effects
4.6. Integration of Results on Aberrant Salience and Source Monitoring
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- McLean, B.F.; Mattiske, J.K.; Balzan, R.P. Association of the Jumping to Conclusions and Evidence Integration Biases with Delusions in Psychosis: A Detailed Meta-analysis. Schizophr. Bull. 2017, 43, 344–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livet, A.; Navarri, X.; Potvin, S.; Conrod, P. Cognitive biases in individuals with psychotic-like experiences: A systematic review and a meta-analysis. Schizophr. Res. 2020, 222, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Lavallé, L.; Dondé, C.; Gawęda, Ł.; Brunelin, J.; Mondino, M. Impaired self-recognition in individuals with no full-blown psychotic symptoms represented across the continuum of psychosis: A meta-analysis. Psychol. Med. 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Dudley, R.; Taylor, P.; Wickham, S. Psychosis, Delusions and the “jumping to Conclusions” Reasoning Bias: A Systematic Review and Meta-Analysis. Schizophr. Bull. 2016, 42, 652–665. [Google Scholar] [CrossRef]
- Brookwell, M.L.; Bentall, R.P.; Varese, F. Externalizing biases and hallucinations in source-monitoring, self-monitoring and signal detection studies: A meta-analytic review. Psychol. Med. 2013, 43, 2465–2475. [Google Scholar] [CrossRef]
- Sauvé, G.; Lavigne, K.M.; Pochiet, G.; Brodeur, M.B.; Lepage, M. Efficacy of psychological interventions targeting cognitive biases in schizophrenia: A systematic review and meta-analysis. Clin. Psychol. Rev. 2020, 78, 101854. [Google Scholar] [CrossRef] [PubMed]
- Moritz, S.; Veckenstedt, R.; Hottenrott, B.; Woodward, T.S.; Randjbar, S.; Lincoln, T.M. Different sides of the same coin? Intercorrelations of cognitive biases in schizophrenia. Cogn. Neuropsychiatry 2010, 15, 406–421. [Google Scholar] [CrossRef]
- Gawęda, Ł.; Woodward, T.S.; Moritz, S.; Kokoszka, A. Impaired action self-monitoring in schizophrenia patients with auditory hallucinations. Schizophr. Res. 2013, 144, 72–79. [Google Scholar] [CrossRef]
- Ochoa, S.; Haro, J.M.; Huerta-Ramos, E.; Cuevas-Esteban, J.; Stephan-Otto, C.; Usall, J.; Nieto, L.; Brébion, G. Relation between jumping to conclusions and cognitive functioning in people with schizophrenia in contrast with healthy participants. Schizophr. Res. 2014, 159, 211–217. [Google Scholar] [CrossRef]
- Woodward, T.S.; Menon, M.; Whitman, J.C. Source monitoring biases and auditory hallucinations. Cogn. Neuropsychiatry 2007, 12, 477–494. [Google Scholar] [CrossRef]
- Moritz, S.; Woodward, T.S. Jumping to conclusions in delusional and non-delusional schizophrenic patients. Br. J. Clin. Psychol. 2005, 44, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Gawęda, Ł.; Staszkiewicz, M.; Balzan, R.P. The relationship between cognitive biases and psychological dimensions of delusions: The importance of jumping to conclusions. J. Behav. Ther. Exp. Psychiatry 2017, 56, 51–56. [Google Scholar] [CrossRef] [PubMed]
- So, S.H.-W.; Chau, A.K.C.; Peters, E.R.; Swendsen, J.; Garety, P.A.; Kapur, S. Moment-to-moment associations between negative affect, aberrant salience, and paranoia. Cogn. Neuropsychiatry 2018, 23, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larøi, F.; Woodward, T.S. Hallucinations from a Cognitive Perspective. Harv. Rev. Psychiatry 2007, 15, 109–117. [Google Scholar] [CrossRef]
- Beck, A.T.; Rector, N.A. A Cognitive Model of Hallucinations. Cogn. Ther. Res. 2003, 27, 19–52. [Google Scholar] [CrossRef]
- Frydecka, D.; Misiak, B.; Kotowicz, K.; Pionke, R.; Krężołek, M.; Cechnicki, A.; Gawęda, Ł. The interplay between childhood trauma, cognitive biases, and cannabis use on the risk of psychosis in nonclinical young adults in Poland. Eur. Psychiatry 2020, 63, 1–9. [Google Scholar] [CrossRef]
- Gawęda, Ł.; Göritz, A.S.; Moritz, S. Mediating role of aberrant salience and self-disturbances for the relationship between childhood trauma and psychotic-like experiences in the general population. Schizophr. Res. 2019, 206, 149–156. [Google Scholar] [CrossRef]
- Kapur, S. Psychosis as a State of Aberrant Salience: A Framework Linking Biology, Phenomenology, and Pharmacology in Schizophrenia. Am. J. Psychiatry 2003, 160, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, K.J.; Johnson, M.K. Source monitoring 15 years later: What have we learned from fMRI about the neural mechanisms of source memory? Psychol. Bull. 2009, 135, 638–677. [Google Scholar] [CrossRef] [Green Version]
- Insel, T.; Cuthbert, B.; Garvey, M.; Heinssen, R.; Pine, D.S.; Quinn, K.; Sanislow, C.; Wang, P. Research Domain Criteria (RDoC): Toward a New Classification Framework for Research on Mental Disorders. Am. J. Psychiatry 2010, 167, 748–751. [Google Scholar] [CrossRef] [Green Version]
- Waters, F.; Allen, P.; Aleman, A.; Fernyhough, C.; Woodward, T.S.; Badcock, J.C.; Barkus, E.; Johns, L.; Varese, F.; Menon, M.; et al. Auditory Hallucinations in Schizophrenia and Nonschizophrenia Populations: A Review and Integrated Model of Cognitive Mechanisms. Schizophr. Bull. 2012, 38, 683–693. [Google Scholar] [CrossRef] [Green Version]
- Nelson, B.; Whitford, T.J.; Lavoie, S.; Sass, L.A. What are the neurocognitive correlates of basic self-disturbance in schizophrenia?: Integrating phenomenology and neurocognition. Part 1 (Source monitoring deficits). Schizophr. Res. 2014, 152, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Howes, O.D.; Hird, E.J.; Adams, R.A.; Corlett, P.R.; McGuire, P. Aberrant Salience, Information Processing, and Dopaminergic Signaling in People at Clinical High Risk for Psychosis. Biol. Psychiatry 2020, 88, 304–314. [Google Scholar] [CrossRef]
- Corlett, P.R.; Taylor, J.R.; Wang, X.-J.; Fletcher, P.C.; Krystal, J.H. Toward a neurobiology of delusions. Prog. Neurobiol. 2010, 92, 345–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corlett, P.R.; Horga, G.; Fletcher, P.C.; Alderson-Day, B.; Schmack, K.; Powers, A.R. Hallucinations and Strong Priors. Trends Cogn. Sci. 2019, 23, 114–127. [Google Scholar] [CrossRef] [Green Version]
- Sterzer, P.; Adams, R.A.; Fletcher, P.; Frith, C.; Lawrie, S.M.; Muckli, L.; Petrovic, P.; Uhlhaas, P.; Voss, M.; Corlett, P.R. The Predictive Coding Account of Psychosis. Biol. Psychiatry 2018, 84, 634–643. [Google Scholar] [CrossRef] [Green Version]
- Winton-Brown, T.; Schmidt, A.; Roiser, J.P.; Howes, O.D.; Egerton, A.; Fusar-Poli, P.; Bunzeck, N.; Grace, A.A.; Duzel, E.; Kapur, S.; et al. Altered activation and connectivity in a hippocampal–basal ganglia–midbrain circuit during salience processing in subjects at ultra high risk for psychosis. Transl. Psychiatry 2017, 7, e1245. [Google Scholar] [CrossRef] [Green Version]
- Cicero, D.C.; Becker, T.M.; Martin, E.A.; Docherty, A.R.; Kerns, J.G. The role of aberrant salience and self-concept clarity in psychotic-like experiences. Personal. Disord. 2013, 4, 33–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelizza, L.; Azzali, S.; Garlassi, S.; Scazza, I.; Paterlini, F.; Chiri, L.R.; Poletti, M.; Pupo, S.; Cicero, D.C.; Preti, A.; et al. Assessing aberrant salience in young community help-seekers with early psychosis: The approved Italian version of the Aberrant Salience Inventory. J. Clin. Psychol. 2021, 77, 782–803. [Google Scholar] [CrossRef]
- Roiser, J.P.; Stephan, K.E.; den Ouden, H.; Barnes, T.R.E.; Friston, K.J.; Joyce, E.M. Do patients with schizophrenia exhibit aberrant salience? Psychol. Med. 2008, 39, 199–209. [Google Scholar] [CrossRef] [Green Version]
- Pankow, A.; Katthagen, T.; Diner, S.; Deserno, L.; Boehme, R.; Kathmann, N.; Gleich, T.; Gaebler, M.; Walter, H.; Heinz, A.; et al. Aberrant Salience Is Related to Dysfunctional Self-Referential Processing in Psychosis. Schizophr. Bull. 2016, 42, 67–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katthagen, T.; Dammering, F.; Kathmann, N.; Kaminski, J.; Walter, H.; Heinz, A.; Schlagenhauf, F. Validating the construct of aberrant salience in schizophrenia—Behavioral evidence for an automatic process. Schizophr. Res. Cogn. 2016, 6, 22–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roiser, J.P.; Howes, O.D.; Chaddock, C.A.; Joyce, E.M.; McGuire, P. Neural and Behavioral Correlates of Aberrant Salience in Individuals at Risk for Psychosis. Schizophr. Bull. 2013, 39, 1328–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.; Antoniades, M.; Allen, P.; Egerton, A.; Chaddock, C.A.; Borgwardt, S.; Fusar-Poli, P.; Roiser, J.P.; Howes, O.; McGuire, P. Longitudinal alterations in motivational salience processing in ultra-high-risk subjects for psychosis. Psychol. Med. 2017, 47, 243–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tessier, C.; Sweers, K.; Frajerman, A.; Bergaoui, H.; Ferreri, F.; Delva, C.; Lapidus, N.; Lamaziere, A.; Roiser, J.P.; De Hert, M.; et al. Membrane lipidomics in schizophrenia patients: A correlational study with clinical and cognitive manifestations. Transl. Psychiatry 2016, 6, e906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, A.; Suenderhauf, C.; Smieskova, R.; Lenz, C.; Harrisberger, F.; Schmidt, A.; Vogel, T.; Lang, U.E.; Riecher-Rössler, A.; Eckert, A.; et al. Altered Insular Function during Aberrant Salience Processing in Relation to the Severity of Psychotic Symptoms. Front. Psychiatry 2016, 7, 189. [Google Scholar] [CrossRef] [Green Version]
- Spark, J.; Gawęda, Ł.; Allott, K.; Hartmann, J.A.; Jack, B.N.; Koren, D.; Lavoie, S.; Li, E.; McGorry, P.D.; Parnas, J.; et al. Distinguishing schizophrenia spectrum from non-spectrum disorders among young patients with first episode psychosis and at high clinical risk: The role of basic self-disturbance and neurocognition. Schizophr. Res. 2021, 228, 19–28. [Google Scholar] [CrossRef]
- Johnson, M.K.; Raye, C.L. Reality Monitoring. Psychol. Rev. 1981, 88, 67. [Google Scholar] [CrossRef]
- Johnson, M.K.; Hashtroudi, S.; Lindsay, D.S. Source Monitoring. Psychol. Bull. 1993, 114, 3–28. [Google Scholar] [CrossRef]
- Bentall, R.P.; Baker, G.A.; Havers, S. Reality monitoring and psychotic hallucinations. Br. J. Clin. Psychol. 1991, 30, 213–222. [Google Scholar] [CrossRef]
- Waters, F.; Woodward, T.; Allen, P.; Aleman, A.; Sommer, I. Self-recognition Deficits in Schizophrenia Patients With Auditory Hallucinations: A Meta-analysis of the Literature. Schizophr. Bull. 2012, 38, 741–750. [Google Scholar] [CrossRef] [Green Version]
- Allen, P.; Aleman, A.; McGuire, P.K. Inner speech models of auditory verbal hallucinations: Evidence from behavioural and neuroimaging studies. Int. Rev. Psychiatry 2007, 19, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Griffin, J.D.; Fletcher, P.C. Predictive Processing, Source Monitoring, and Psychosis. Annu. Rev. Clin. Psychol. 2017, 13, 265–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, P.C.; Frith, C.D. Perceiving is believing: A Bayesian approach to explaining the positive symptoms of schizophrenia. Nat. Rev. Neurosci. 2009, 10, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Nelson, B.; Whitford, T.J.; Lavoie, S.; Sass, L.A. What are the neurocognitive correlates of basic self-disturbance in schizophrenia?: Integrating phenomenology and neurocognition: Part 2 (Aberrant salience). Schizophr. Res. 2013, 152, 20–27. [Google Scholar] [CrossRef]
- Nelson, B.; Lavoie, S.; Gawęda, Ł.; Li, E.; Sass, L.A.; Koren, D.; McGorry, P.D.; Jack, B.N.; Parnas, J.; Polari, A.; et al. The neurophenomenology of early psychosis: An integrative empirical study. Conscious. Cogn. 2020, 77, 102845. [Google Scholar] [CrossRef] [PubMed]
- Nelson, B.; Sass, L.A. Towards integrating phenomenology and neurocognition: Possible neurocognitive correlates of basic self-disturbance in schizophrenia. Curr. Probl. Psychiatry 2017, 18, 184–200. [Google Scholar] [CrossRef] [Green Version]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, 1–9. [Google Scholar] [CrossRef]
- Juckel, G.; Friedel, E.; Koslowski, M.; Witthaus, H.; Özgürdal, S.; Gudlowski, Y.; Knutson, B.; Wrase, J.; Brüne, M.; Heinz, A.; et al. Ventral Striatal Activation during Reward Processing in Subjects with Ultra-High Risk for Schizophrenia. Neuropsychobiology 2012, 66, 50–56. [Google Scholar] [CrossRef]
- Wotruba, D.; Heekeren, K.; Michels, L.; Buechler, R.; Simon, J.J.; Theodoridou, A.; Kollias, S.; Rãssler, W.; Kaiser, S.; Rössler, W.; et al. Symptom dimensions are associated with reward processing in unmedicated persons at risk for psychosis. Front. Behav. Neurosci. 2014, 8, 382. [Google Scholar] [CrossRef] [Green Version]
- Shergill, S.S.; Bullmore, E.; Simmons, A.; Murray, R.; McGuire, P. Functional Anatomy of Auditory Verbal Imagery in Schizophrenic Patients With Auditory Hallucinations. Am. J. Psychiatry 2000, 157, 1691–1693. [Google Scholar] [CrossRef]
- Simons, C.J.P.; Tracy, D.K.; Sanghera, K.K.; O’Daly, O.; Gilleen, J.; Dominguez, M.-D.; Krabbendam, L.; Shergill, S.S. Functional Magnetic Resonance Imaging of Inner Speech in Schizophrenia. Biol. Psychiatry 2010, 67, 232–237. [Google Scholar] [CrossRef]
- Sawilowsky, S.S. New Effect Size Rules of Thumb. J. Mod. Appl. Stat. Methods 2009, 8, 597–599. [Google Scholar] [CrossRef]
- Hayden, J.A.; Van Der Windt, D.A.; Cartwright, J.L.; Côté, P.; Bombardier, C. Assessing Bias in Studies of Prognostic Factors. Ann. Intern. Med. 2013, 158, 280–286. [Google Scholar] [CrossRef]
- Katthagen, T.; Mathys, C.; Deserno, L.; Walter, H.; Kathmann, N.; Heinz, A.; Schlagenhauf, F. Modeling subjective relevance in schizophrenia and its relation to aberrant salience. PLoS Comput. Biol. 2018, 14, e1006319. [Google Scholar] [CrossRef]
- Smieskova, R.; Roiser, J.P.; Chaddock, C.A.; Schmidt, A.; Harrisberger, F.; Bendfeldt, K.; Simon, A.; Walter, A.; Fusar-Poli, P.; McGuire, P.K.; et al. Modulation of motivational salience processing during the early stages of psychosis. Schizophr. Res. 2015, 166, 17–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roiser, J.P.; Stephan, K.E.; den Ouden, H.E.; Friston, K.J.; Joyce, E.M. Adaptive and aberrant reward prediction signals in the human brain. NeuroImage 2010, 50, 657–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vul, E.; Harris, C.; Winkielman, P.; Pashler, H. Puzzlingly High Correlations in fMRI Studies of Emotion, Personality, and Social Cognition. Perspect. Psychol. Sci. 2009, 4, 274–290. [Google Scholar] [CrossRef] [PubMed]
- Esslinger, C.; Englisch, S.; Inta, D.; Rausch, F.; Schirmbeck, F.; Mier, D.; Kirsch, P.; Meyer-Lindenberg, A.; Zink, M. Ventral striatal activation during attribution of stimulus saliency and reward anticipation is correlated in unmedicated first episode schizophrenia patients. Schizophr. Res. 2012, 140, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Allen, P.; Amaro, E.; Fu, C.H.Y.; Williams, S.C.R.; Brammer, M.J.; Johns, L.C.; McGuire, P.K. Neural correlates of the misattribution of speech in schizophrenia. Br. J. Psychiatry 2007, 190, 162–169. [Google Scholar] [CrossRef] [Green Version]
- Mechelli, A.; Allen, P.; Amaro, E.; Fu, C.H.Y.; Williams, S.C.R.; Brammer, M.J.; Johns, L.C.; McGuire, P.K. Misattribution of speech and impaired connectivity in patients with auditory verbal hallucinations. Hum. Brain Mapp. 2007, 28, 1213–1222. [Google Scholar] [CrossRef] [PubMed]
- Garrison, J.R.; Fernandez-Egea, E.; Zaman, R.; Agius, M.; Simons, J.S. Reality monitoring impairment in schizophrenia reflects specific prefrontal cortex dysfunction. NeuroImage Clin. 2017, 14, 260–268. [Google Scholar] [CrossRef]
- Kumari, V.; Fannon, D.; Ffytche, D.H.; Raveendran, V.; Antonova, E.; Premkumar, P.; Cooke, M.A.; Anilkumar, A.P.P.; Williams, S.C.R.; Andrew, C.; et al. Functional MRI of Verbal Self-monitoring in Schizophrenia: Performance and Illness-Specific Effects. Schizophr. Bull. 2008, 36, 740–755. [Google Scholar] [CrossRef] [Green Version]
- Thoresen, C.; Endestad, T.; Sigvartsen, N.P.B.; Server, A.; Bolstad, I.; Johansson, M.; Andreassen, O.A.; Jensen, J. Frontotemporal hypoactivity during a reality monitoring paradigm is associated with delusions in patients with schizophrenia spectrum disorders. Cogn. Neuropsychiatry 2014, 19, 97–115. [Google Scholar] [CrossRef] [PubMed]
- Vinogradov, S.; Luks, T.L.; Schulman, B.J.; Simpson, G.V. Deficit in a Neural Correlate of Reality Monitoring in Schizophrenia Patients. Cereb. Cortex 2008, 18, 2532–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramaniam, K.; Luks, T.L.; Fisher, M.; Simpson, G.V.; Nagarajan, S.; Vinogradov, S. Computerized Cognitive Training Restores Neural Activity within the Reality Monitoring Network in Schizophrenia. Neuron 2012, 73, 842–853. [Google Scholar] [CrossRef] [Green Version]
- Subramaniam, K.; Ranasinghe, K.G.; Mathalon, D.; Nagarajan, S.; Vinogradov, S. Neural mechanisms of mood-induced modulation of reality monitoring in schizophrenia. Cortex 2017, 91, 271–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Metzak, P.D.; Woodward, T.S. Aberrant connectivity during self–other source monitoring in schizophrenia. Schizophr. Res. 2011, 125, 136–142. [Google Scholar] [CrossRef]
- Kambeitz-Ilankovic, L.; Hennig-Fast, K.; Benetti, S.; Kambeitz, J.; Pettersson-Yeo, W.; O’Daly, O.; McGuire, P.; Allen, P. Attentional Modulation of Source Attribution in First-Episode Psychosis: A Functional Magnetic Resonance Imaging Study. Schizophr. Bull. 2013, 39, 1027–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephan-Otto, C.; Siddi, S.; Senior, C.; Cuevas-Esteban, J.; Cambra-Martí, M.R.; Ochoa, S.; Brébion, G. Remembering verbally-presented items as pictures: Brain activity underlying visual mental images in schizophrenia patients with visual hallucinations. Cortex 2017, 94, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Winton-Brown, T.T.; Fusar-Poli, P.; Ungless, M.A.; Howes, O.D. Dopaminergic basis of salience dysregulation in psychosis. Trends Neurosci. 2014, 37, 85–94. [Google Scholar] [CrossRef]
- Toga, A.W. Brain Mapping: An Encyclopedic Reference; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Limongi, R.; Jeon, P.; MacKinley, M.; Das, T.; Dempster, K.; Théberge, J.; Bartha, R.; Wong, D.; Palaniyappan, L. Glutamate and Dysconnection in the Salience Network: Neurochemical, Effective Connectivity, and Computational Evidence in Schizophrenia. Biol. Psychiatry 2020, 88, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Limongi, R.; Jeon, P.; Théberge, J.; Palaniyappan, L. Counteracting Effects of Glutathione on the Glutamate-Driven Excitation/Inhibition Imbalance in First-Episode Schizophrenia: A 7T MRS and Dynamic Causal Modeling Study. Antioxidants 2021, 10, 75. [Google Scholar] [CrossRef] [PubMed]
- Von Düring, F.; Ristow, I.; Li, M.; Denzel, D.; Colic, L.; Demenescu, L.R.; Li, S.; Borchardt, V.; Liebe, T.; Vogel, M.; et al. Glutamate in Salience Network Predicts BOLD Response in Default Mode Network During Salience Processing. Front. Behav. Neurosci. 2019, 13, 232. [Google Scholar] [CrossRef]
- Grace, A.A.; Floresco, S.B.; Goto, Y.; Lodge, D.J. Regulation of firing of dopaminergic neurons and control of goal-directed behaviors. Trends Neurosci. 2007, 30, 220–227. [Google Scholar] [CrossRef]
- Radua, J.; Schmidt, A.; Borgwardt, S.; Heinz, A.; Schlagenhauf, F.; McGuire, P.; Fusar-Poli, P. Ventral Striatal Activation During Reward Processing in Psychosis: A Neurofunctional Meta-Analysis. JAMA Psychiatry 2015, 72, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Howes, O.D.; Kapur, S. The Dopamine Hypothesis of Schizophrenia: Version III-The Final Common Pathway. Schizophr. Bull. 2009, 35, 549–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinz, A.; Schlagenhauf, F. Dopaminergic Dysfunction in Schizophrenia: Salience Attribution Revisited. Schizophr. Bull. 2010, 36, 472–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, R.S.; Boccia, M.M.; Pedreira, M.E. The fate of memory: Reconsolidation and the case of Prediction Error. Neurosci. Biobehav. Rev. 2016, 68, 423–441. [Google Scholar] [CrossRef]
- Lodge, D.J.; Grace, A.A. Hippocampal dysregulation of dopamine system function and the pathophysiology of schizophrenia. Trends Pharmacol. Sci. 2011, 32, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Menon, V.; Uddin, L.Q. Saliency, switching, attention and control: A network model of insula function. Brain Struct. Funct. 2010, 214, 655–667. [Google Scholar] [CrossRef] [Green Version]
- Goulden, N.; Khusnulina, A.; Davis, N.J.; Bracewell, R.M.; Bokde, A.L.; McNulty, J.P.; Mullins, P.G. The salience network is responsible for switching between the default mode network and the central executive network: Replication from DCM. NeuroImage 2014, 99, 180–190. [Google Scholar] [CrossRef]
- Palaniyappan, L.; Liddle, P.F. Does the salience network play a cardinal role in psychosis? An emerging hypothesis of insular dysfunction. J. Psychiatry Neurosci. 2012, 37, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R.; Neufeld, R.W.J.; Schaefer, B.; Densmore, M.; Rajakumar, N.; Osuch, E.A.; Williamson, P.C.; Theberge, J. Functional magnetic resonance spectroscopy of glutamate in schizophrenia and major depressive disorder: Anterior cingulate activity during a color-word Stroop task. NPJ Schizophr. 2015, 1, 15028. [Google Scholar] [CrossRef]
- Wotruba, D.; Michels, L.; Buechler, R.; Metzler, S.; Theodoridou, A.; Gerstenberg, M.; Walitza, S.; Kollias, S.; Rössler, W.; Heekeren, K. Aberrant Coupling Within and Across the Default Mode, Task-Positive, and Salience Network in Subjects at Risk for Psychosis. Schizophr. Bull. 2014, 40, 1095–1104. [Google Scholar] [CrossRef] [Green Version]
- Mallikarjun, P.K.; Lalousis, P.A.; Dunne, T.F.; Heinze, K.; Reniers, R.L.; Broome, M.R.; Farmah, B.; Oyebode, F.; Wood, S.J.; Upthegrove, R. Aberrant salience network functional connectivity in auditory verbal hallucinations: A first episode psychosis sample. Transl. Psychiatry 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Manoliu, A.; Riedl, V.; Zherdin, A.; Mühlau, M.; Schwerthöffer, D.; Scherr, M.; Peters, H.; Zimmer, C.; Förstl, H.; Bäuml, J.; et al. Aberrant Dependence of Default Mode/Central Executive Network Interactions on Anterior Insular Salience Network Activity in Schizophrenia. Schizophr. Bull. 2014, 40, 428–437. [Google Scholar] [CrossRef] [Green Version]
- Bolton, T.A.W.; Wotruba, D.; Buechler, R.; Theodoridou, A.; Michels, L.; Kollias, S.; Rössler, W.; Heekeren, K.; Van De Ville, D. Triple Network Model Dynamically Revisited: Lower Salience Network State Switching in Pre-psychosis. Front. Physiol. 2020, 11, 66. [Google Scholar] [CrossRef]
- Howes, O.D.; Murray, R.M. Schizophrenia: An integrated sociodevelopmental-cognitive model. Lancet 2014, 383, 1677–1687. [Google Scholar] [CrossRef] [Green Version]
- Maia, T.V.; Frank, M.J. An Integrative Perspective on the Role of Dopamine in Schizophrenia. Biol. Psychiatry 2017, 81, 52–66. [Google Scholar] [CrossRef] [Green Version]
- McCutcheon, R.A.; Nour, M.M.; Dahoun, T.; Jauhar, S.; Pepper, F.; Expert, P.; Veronese, M.; Adams, R.A.; Turkheimer, F.; Mehta, M.A.; et al. Mesolimbic Dopamine Function Is Related to Salience Network Connectivity: An Integrative Positron Emission Tomography and Magnetic Resonance Study. Biol. Psychiatry 2019, 85, 368–378. [Google Scholar] [CrossRef] [Green Version]
- Howes, O.; McCutcheon, R.; Stone, J. Glutamate and dopamine in schizophrenia: An update for the 21st century. J. Psychopharmacol. 2015, 29, 97–115. [Google Scholar] [CrossRef] [Green Version]
- Moritz, S.; Silverstein, S.M.; Beblo, T.; Özaslan, Z.; Zink, M.; Gallinat, J. Much of the Neurocognitive Impairment in Schizophrenia is Due to Factors Other Than Schizophrenia Itself: Implications for Research and Treatment. Schizophr. Bull. Open 2021, 2. [Google Scholar] [CrossRef]
- Moritz, S.; Klein, J.P.; Desler, T.; Lill, H.; Gallinat, J.; Schneider, B.C. Neurocognitive deficits in schizophrenia. Are we making mountains out of molehills? Psychol. Med. 2017, 47, 2602–2612. [Google Scholar] [CrossRef]
- Gilbert, S.J.; Spengler, S.; Simons, J.S.; Steele, J.D.; Lawrie, S.M.; Frith, C.D.; Burgess, P.W. Functional Specialization within Rostral Prefrontal Cortex (Area 10): A Meta-analysis. J. Cogn. Neurosci. 2006, 18, 932–948. [Google Scholar] [CrossRef] [Green Version]
- Simons, J.S.; Garrison, J.R.; Johnson, M.K. Brain Mechanisms of Reality Monitoring. Trends Cogn. Sci. 2017, 21, 462–473. [Google Scholar] [CrossRef] [Green Version]
- Rugg, M.D.; Vilberg, K.L. Brain networks underlying episodic memory retrieval. Curr. Opin. Neurobiol. 2013, 23, 255–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moscovitch, M.; Cabeza, R.; Winocur, G.; Nadel, L. Episodic Memory and Beyond: The Hippocampus and Neocortex in Transformation. Annu. Rev. Psychol. 2016, 67, 105–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guise, K.G.; Shapiro, M.L. Medial Prefrontal Cortex Reduces Memory Interference by Modifying Hippocampal Encoding. Neuron 2017, 94, 183–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fornito, A.; Wood, S.J.; Whittle, S.; Fuller, J.; Adamson, C.; Saling, M.M.; Velakoulis, D.; Pantelis, C.; Yücel, M. Variability of the paracingulate sulcus and morphometry of the medial frontal cortex: Associations with cortical thickness, surface area, volume, and sulcal depth. Hum. Brain Mapp. 2008, 29, 222–236. [Google Scholar] [CrossRef]
- Buda, M.; Fornito, A.; Bergström, Z.M.; Simons, J.S. A Specific Brain Structural Basis for Individual Differences in Reality Monitoring. J. Neurosci. 2011, 31, 14308–14313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zmigrod, L.; Garrison, J.R.; Carr, J.; Simons, J.S. The neural mechanisms of hallucinations: A quantitative meta-analysis of neuroimaging studies. Neurosci. Biobehav. Rev. 2016, 69, 113–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisman, J.E.; Grace, A.A. The Hippocampal-VTA Loop: Controlling the Entry of Information into Long-Term Memory. Neuron 2005, 46, 703–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodge, D.J.; Grace, A.A. Aberrant Hippocampal Activity Underlies the Dopamine Dysregulation in an Animal Model of Schizophrenia. J. Neurosci. 2007, 27, 11424–11430. [Google Scholar] [CrossRef]
- Perini, I.; Gustafsson, P.A.; Hamilton, J.P.; Kämpe, R.; Zetterqvist, M.; Heilig, M. The salience of self, not social pain, is encoded by dorsal anterior cingulate and insula. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Floresco, S.B.; Todd, C.L.; Grace, A.A. Glutamatergic Afferents from the Hippocampus to the Nucleus Accumbens Regulate Activity of Ventral Tegmental Area Dopamine Neurons. J. Neurosci. 2001, 21, 4915–4922. [Google Scholar] [CrossRef] [Green Version]
- Lodge, D.J.; Grace, A.A. The Hippocampus Modulates Dopamine Neuron Responsivity by Regulating the Intensity of Phasic Neuron Activation. Neuropsychopharmacology 2006, 31, 1356–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonnenschein, S.F.; Gomes, F.V.; Grace, A.A. Dysregulation of Midbrain Dopamine System and the Pathophysiology of Schizophrenia. Front. Psychiatry 2020, 11, 613. [Google Scholar] [CrossRef]
- Howes, O.D.; Kambeitz, J.; Kim, E.; Stahl, D.; Slifstein, M.; Abi-Dargham, A.; Kapur, S. The Nature of Dopamine Dysfunction in Schizophrenia and What This Means for Treatment: Meta-Analysis of Imaging Studies. Arch. Gen. Psychiatry 2012, 69, 776–786. [Google Scholar] [CrossRef] [Green Version]
- Egerton, A.; Chaddock, C.A.; Winton-Brown, T.T.; Bloomfield, M.A.P.; Bhattacharyya, S.; Allen, P.; McGuire, P.K.; Howes, O.D. Presynaptic Striatal Dopamine Dysfunction in People at Ultra-high Risk for Psychosis: Findings in a Second Cohort. Biol. Psychiatry 2013, 74, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, S.; Matsumoto, M.; Van Erp, T.G.M. Hippocampal subregion abnormalities in schizophrenia: A systematic review of structural and physiological imaging studies. Neuropsychopharmacol. Rep. 2018, 38, 156–166. [Google Scholar] [CrossRef]
- Pantelis, C.; Velakoulis, D.; McGorry, P.D.; Wood, S.J.; Suckling, J.; Phillips, L.J.; Yung, A.R.; Bullmore, E.; Brewer, W.; Soulsby, B.; et al. Neuroanatomical abnormalities before and after onset of psychosis: A cross-sectional and longitudinal MRI comparison. Lancet 2003, 361, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.J.; Kennedy, D.; Phillips, L.J.; Seal, M.L.; Yücel, M.; Nelson, B.; Yung, A.R.; Jackson, G.; McGorry, P.D.; Velakoulis, D.; et al. Hippocampal pathology in individuals at ultra-high risk for psychosis: A multi-modal magnetic resonance study. NeuroImage 2010, 52, 62–68. [Google Scholar] [CrossRef]
- Allen, P.; Luigjes, J.; Howes, O.D.; Egerton, A.; Hirao, K.; Valli, I.; Kambeitz, J.; Fusar-Poli, P.; Broome, M.; McGuire, P. Transition to Psychosis Associated With Prefrontal and Subcortical Dysfunction in Ultra High-Risk Individuals. Schizophr. Bull. 2012, 38, 1268–1276. [Google Scholar] [CrossRef] [Green Version]
- Medoff, D.R.; Holcomb, H.H.; Lahti, A.C.; Tamminga, C.A. Probing the human hippocampus using rCBF: Contrasts in schizophrenia. Hippocampus 2001, 11, 543–550. [Google Scholar] [CrossRef]
- Lahti, A.C.; Weiler, M.A.; Holcomb, H.H.; Tamminga, C.A.; Cropsey, K.L. Modulation of Limbic Circuitry Predicts Treatment Response to Antipsychotic Medication: A Functional Imaging Study in Schizophrenia. Neuropsychopharmacology 2009, 34, 2675–2690. [Google Scholar] [CrossRef] [Green Version]
- Stone, J.M.; Howes, O.D.; Egerton, A.; Kambeitz, J.; Allen, P.; Lythgoe, D.J.; O’Gorman, R.L.; McLean, M.A.; Barker, G.J.; McGuire, P. Altered Relationship Between Hippocampal Glutamate Levels and Striatal Dopamine Function in Subjects at Ultra High Risk of Psychosis. Biol. Psychiatry 2010, 68, 599–602. [Google Scholar] [CrossRef] [Green Version]
- Lodge, D.J.; Grace, A.A. Developmental pathology, dopamine, stress and schizophrenia. Int. J. Dev. Neurosci. 2011, 29, 207–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.P.; Joseph, V.; Francis, S.T.; Liddle, P.F. Aberrant salience network (bilateral insula and anterior cingulate cortex) connectivity during information processing in schizophrenia. Schizophr. Res. 2010, 123, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Tian, L.; Yan, J.; Sun, W.; Liu, Q.; Zhang, Y.-B.; Li, X.-M.; Zang, Y.-F.; Zhang, D. Functional and Anatomical Connectivity Abnormalities in Cognitive Division of Anterior Cingulate Cortex in Schizophrenia. PLoS ONE 2012, 7, e45659. [Google Scholar] [CrossRef] [PubMed]
- Rametti, G.; Junqué, C.; Bartrés-Faz, D.; Zubiaurre-Elorza, L.; Catalán, R.; Penadés, R.; Bargalló, N.; Bernardo, M. Anterior cingulate and paracingulate sulci morphology in patients with schizophrenia. Schizophr. Res. 2010, 121, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Ellison-Wright, I.; Bullmore, E. Anatomy of bipolar disorder and schizophrenia: A meta-analysis. Schizophr. Res. 2010, 117, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Goodkind, M.; Eickhoff, S.B.; Oathes, D.J.; Jiang, Y.; Chang, A.; Jones-Hagata, L.B.; Ortega, B.N.; Zaiko, Y.V.; Roach, E.L.; Korgaonkar, M.S.; et al. Identification of a Common Neurobiological Substrate for Mental Illness. JAMA Psychiatry 2015, 72, 305–315. [Google Scholar] [CrossRef]
- Sukumar, N.; Sabesan, P.; Anazodo, U.; Palaniyappan, L. Neurovascular Uncoupling in Schizophrenia: A Bimodal Meta-Analysis of Brain Perfusion and Glucose Metabolism. Front. Psychiatry 2020, 11, 754. [Google Scholar] [CrossRef] [PubMed]
- Garrison, J.R.; The Australian Schizophrenia Research Bank; Fernyhough, C.; McCarthy-Jones, S.; Haggard, M.; Simons, J.; Carr, V.; Schall, U.; Scott, R.; Jablensky, A.; et al. Paracingulate sulcus morphology is associated with hallucinations in the human brain. Nat. Commun. 2015, 6, 8956. [Google Scholar] [CrossRef] [Green Version]
- Garrison, J.R.; Fernyhough, C.; McCarthy-Jones, S.; Simons, J.S.; Sommer, I.E.C. Paracingulate Sulcus Morphology and Hallucinations in Clinical and Nonclinical Groups. Schizophr. Bull. 2019, 45, 733–741. [Google Scholar] [CrossRef]
- Sass, L.; Borda, J.P.; Madeira, L.; Pienkos, E.; Nelson, B. Varieties of Self Disorder: A Bio-Pheno-Social Model of Schizophrenia. Schizophr. Bull. 2018, 44, 720–727. [Google Scholar] [CrossRef]
- Rosen, C.; Jones, N.; Chase, K.A.; Gin, H.; Grossman, L.S.; Sharma, R.P. The intrasubjectivity of self, voices and delusions: A phenomenological analysis. Psychosis 2016, 8, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Pienkos, E.; Giersch, A.; Hansen, M.; Humpston, C.; McCarthy-Jones, S.; Mishara, A.; Nelson, B.; Park, S.; Raballo, A.; Sharma, R.; et al. Hallucinations Beyond Voices: A Conceptual Review of the Phenomenology of Altered Perception in Psychosis. Schizophr. Bull. 2019, 45, S67–S77. [Google Scholar] [CrossRef]
- Nelson, B.; Lavoie, S.; Gawęda, Ł.; Li, E.; Sass, L.A.; Koren, D.; McGorry, P.D.; Jack, B.N.; Parnas, J.; Polari, A.; et al. Testing a neurophenomenological model of basic self disturbance in early psychosis. World Psychiatry 2019, 18, 104–105. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalski, J.; Aleksandrowicz, A.; Dąbkowska, M.; Gawęda, Ł. Neural Correlates of Aberrant Salience and Source Monitoring in Schizophrenia and At-Risk Mental States—A Systematic Review of fMRI Studies. J. Clin. Med. 2021, 10, 4126. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm10184126
Kowalski J, Aleksandrowicz A, Dąbkowska M, Gawęda Ł. Neural Correlates of Aberrant Salience and Source Monitoring in Schizophrenia and At-Risk Mental States—A Systematic Review of fMRI Studies. Journal of Clinical Medicine. 2021; 10(18):4126. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm10184126
Chicago/Turabian StyleKowalski, Joachim, Adrianna Aleksandrowicz, Małgorzata Dąbkowska, and Łukasz Gawęda. 2021. "Neural Correlates of Aberrant Salience and Source Monitoring in Schizophrenia and At-Risk Mental States—A Systematic Review of fMRI Studies" Journal of Clinical Medicine 10, no. 18: 4126. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm10184126