The Development of the Ascending Aortic Wall in Tricuspid and Bicuspid Aortic Valve: A Process from Maturation to Degeneration

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Patients and Tissue Samples and Ethical Approval
2.2. Sample Processing, Routine Histology, and Immunohistochemistry
2.3. Histologic Parameters and Morphometry
2.4. Statistical Analysis
3. Results
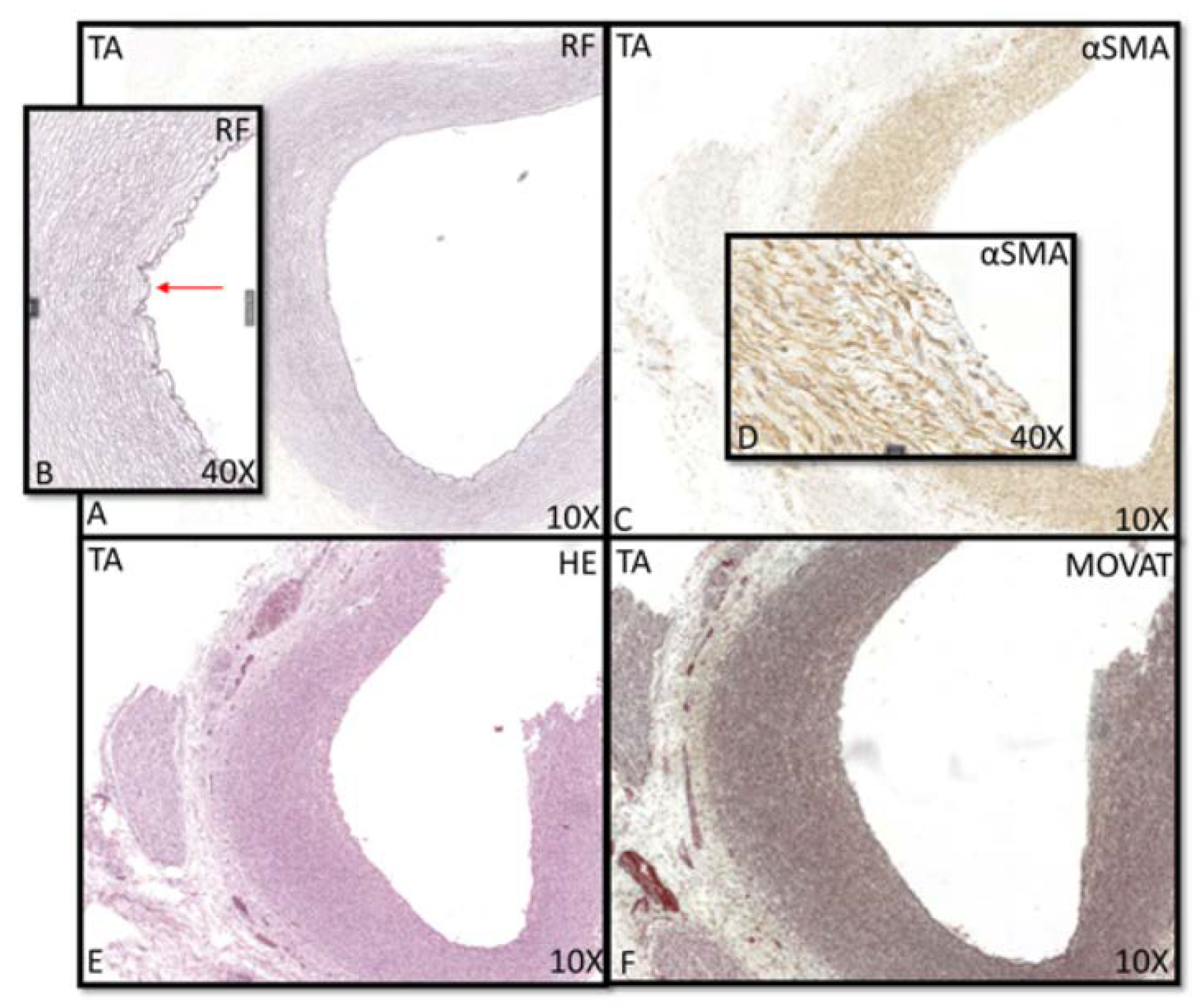
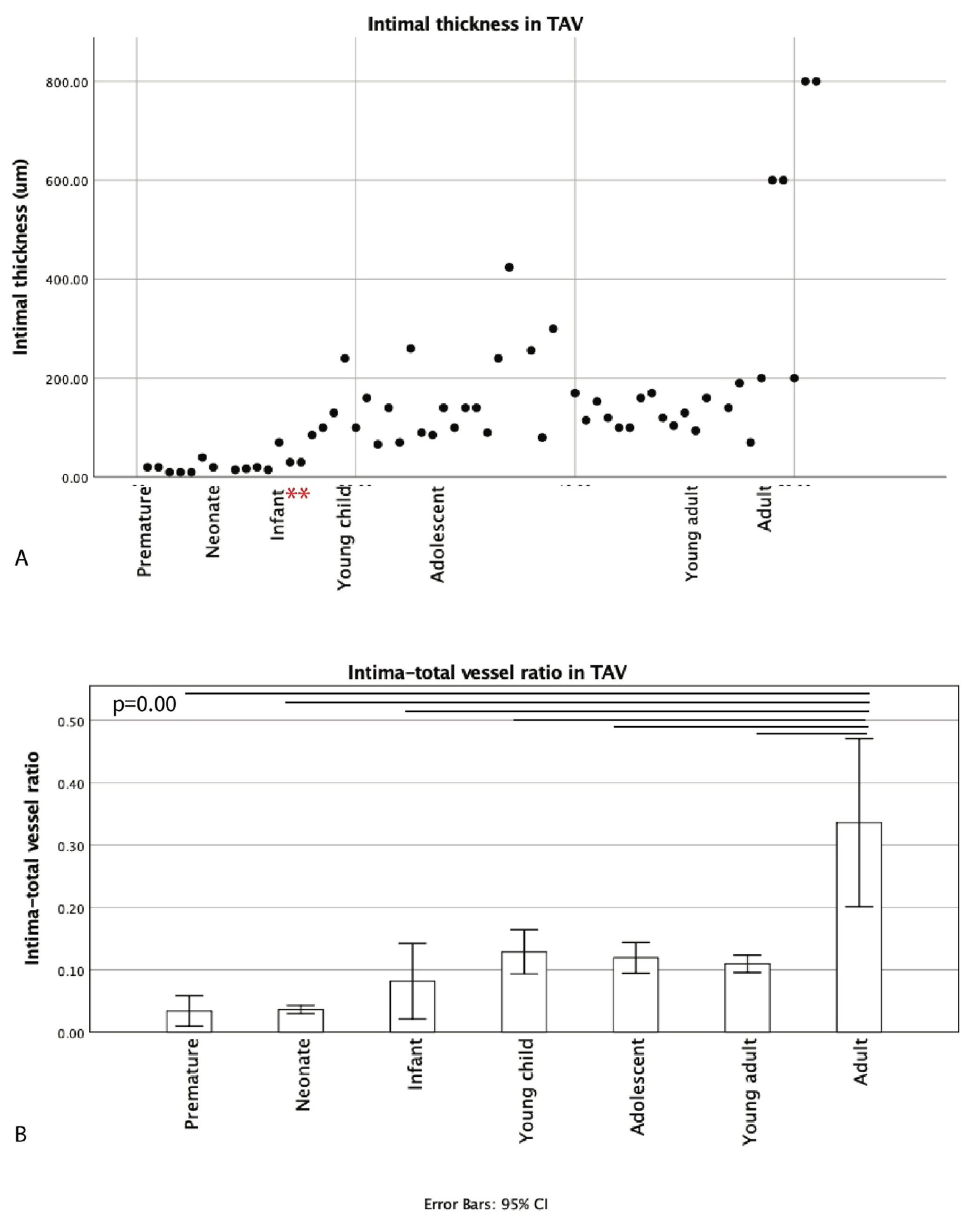
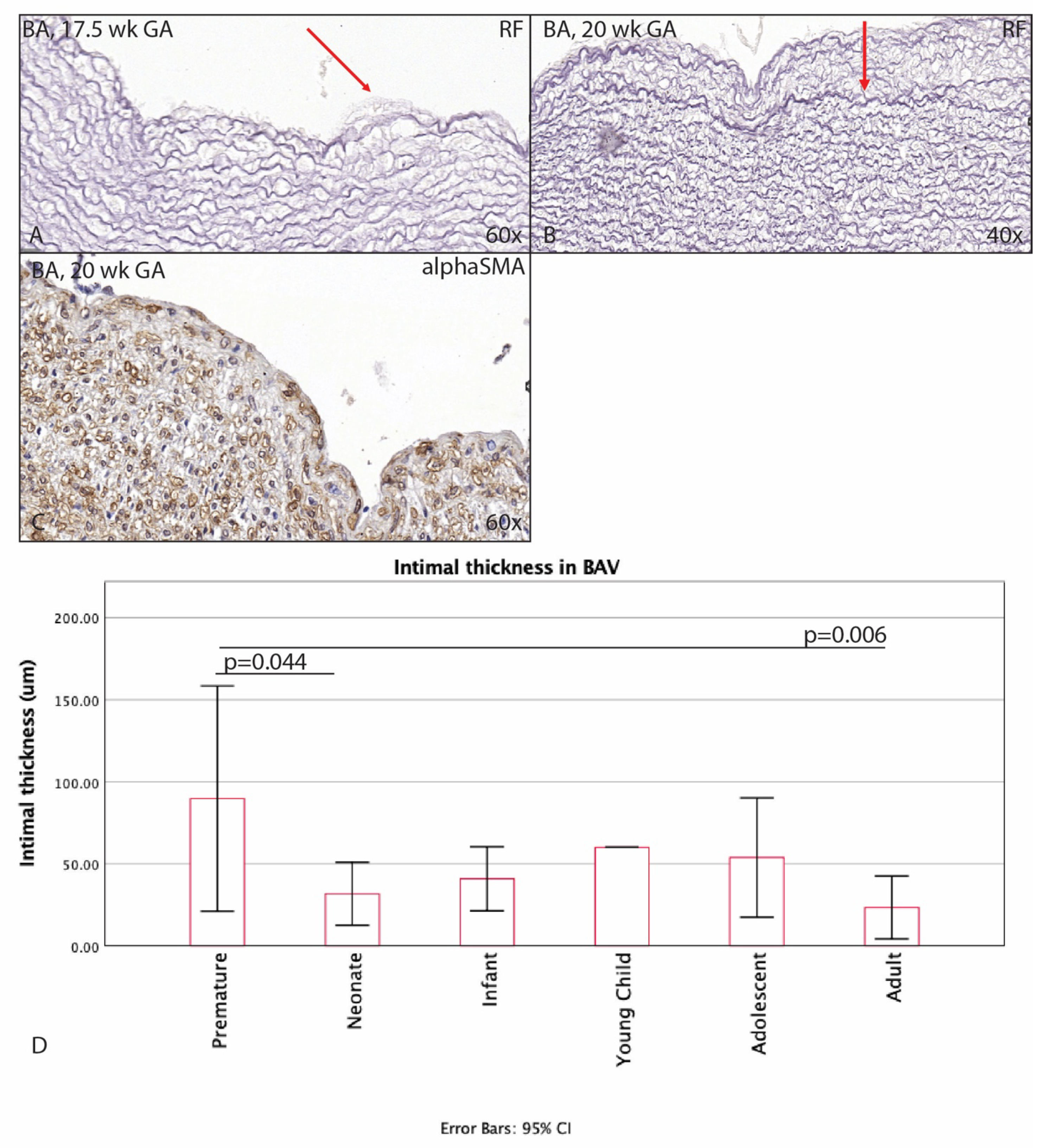
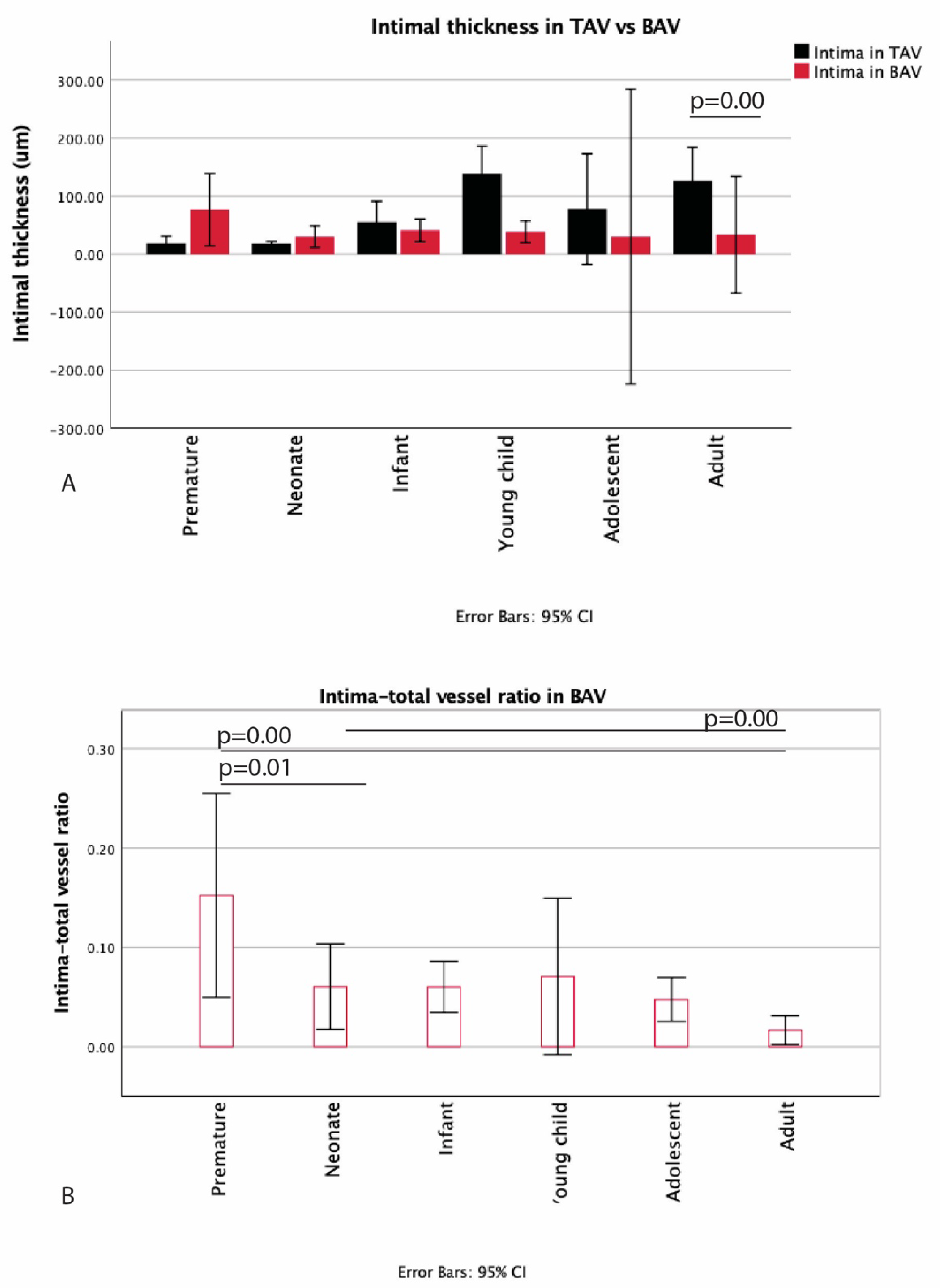
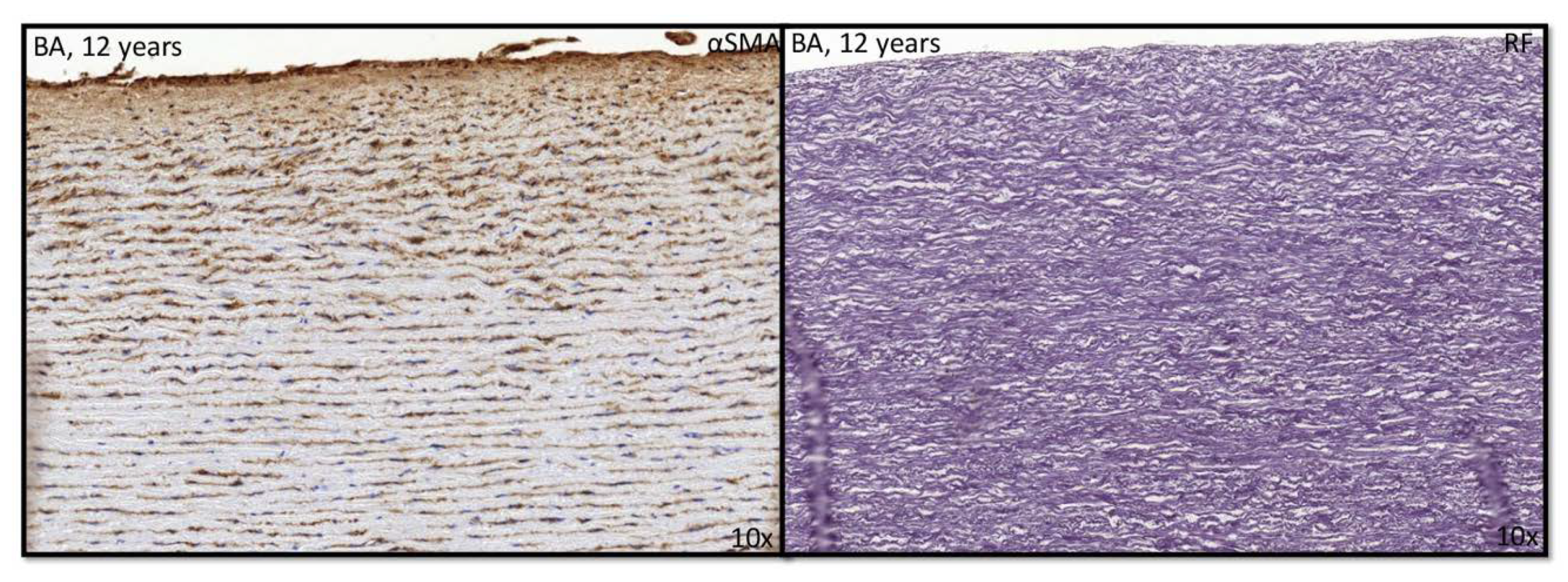
3.1. Development of the Innermost Layer of the Ascending Aorta
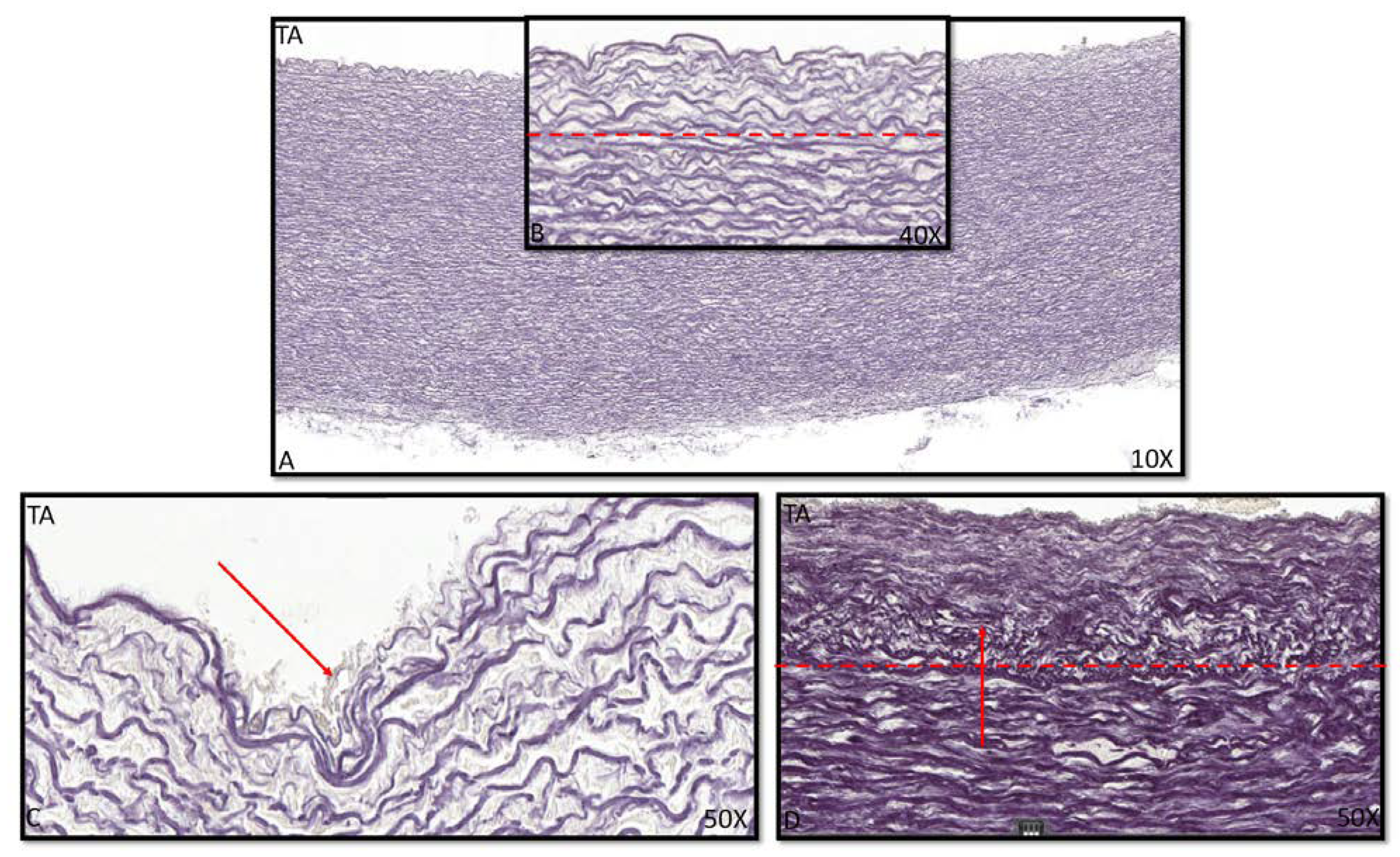
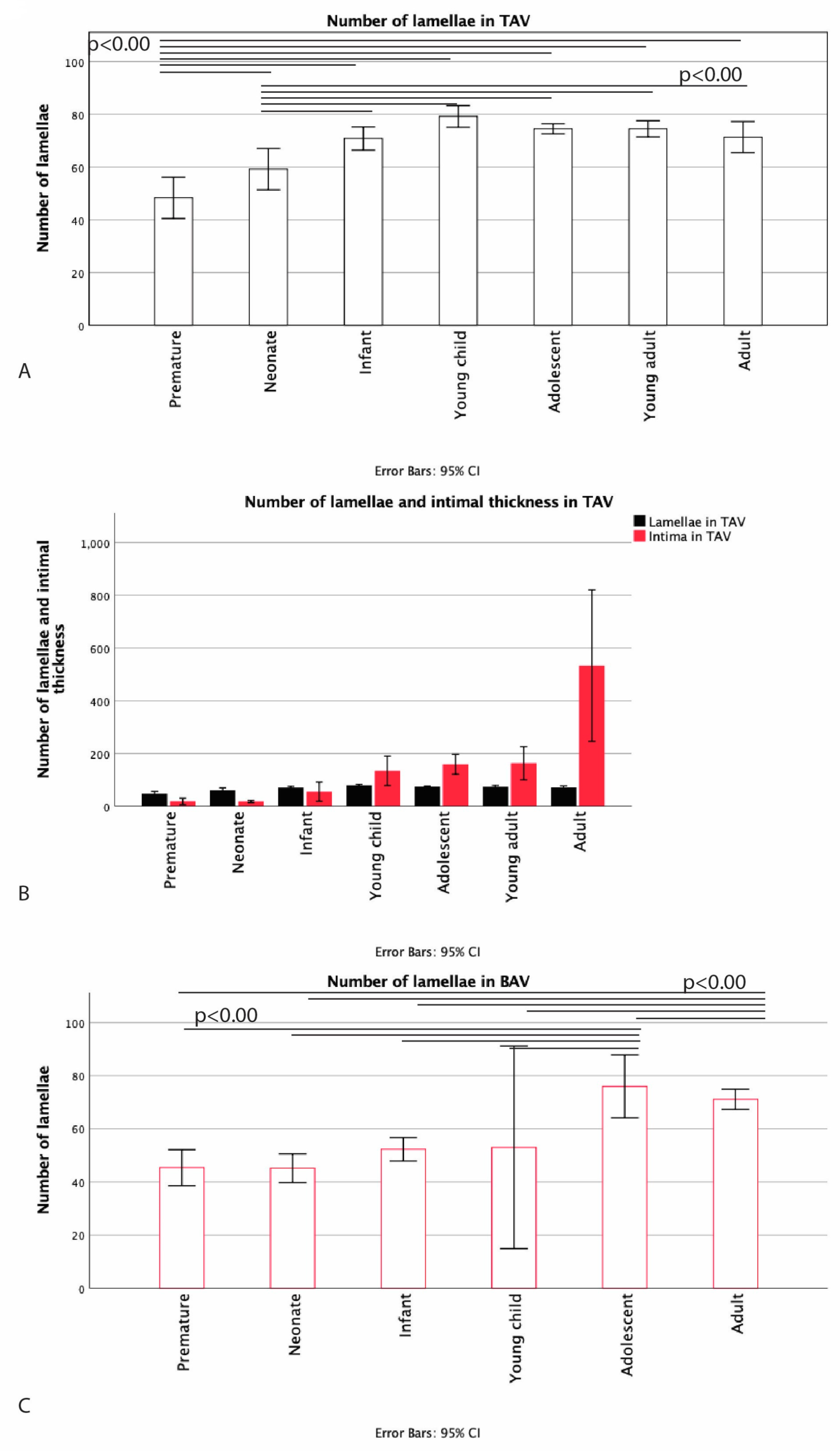
3.2. Development of the Middle Layer of the Ascending Aorta
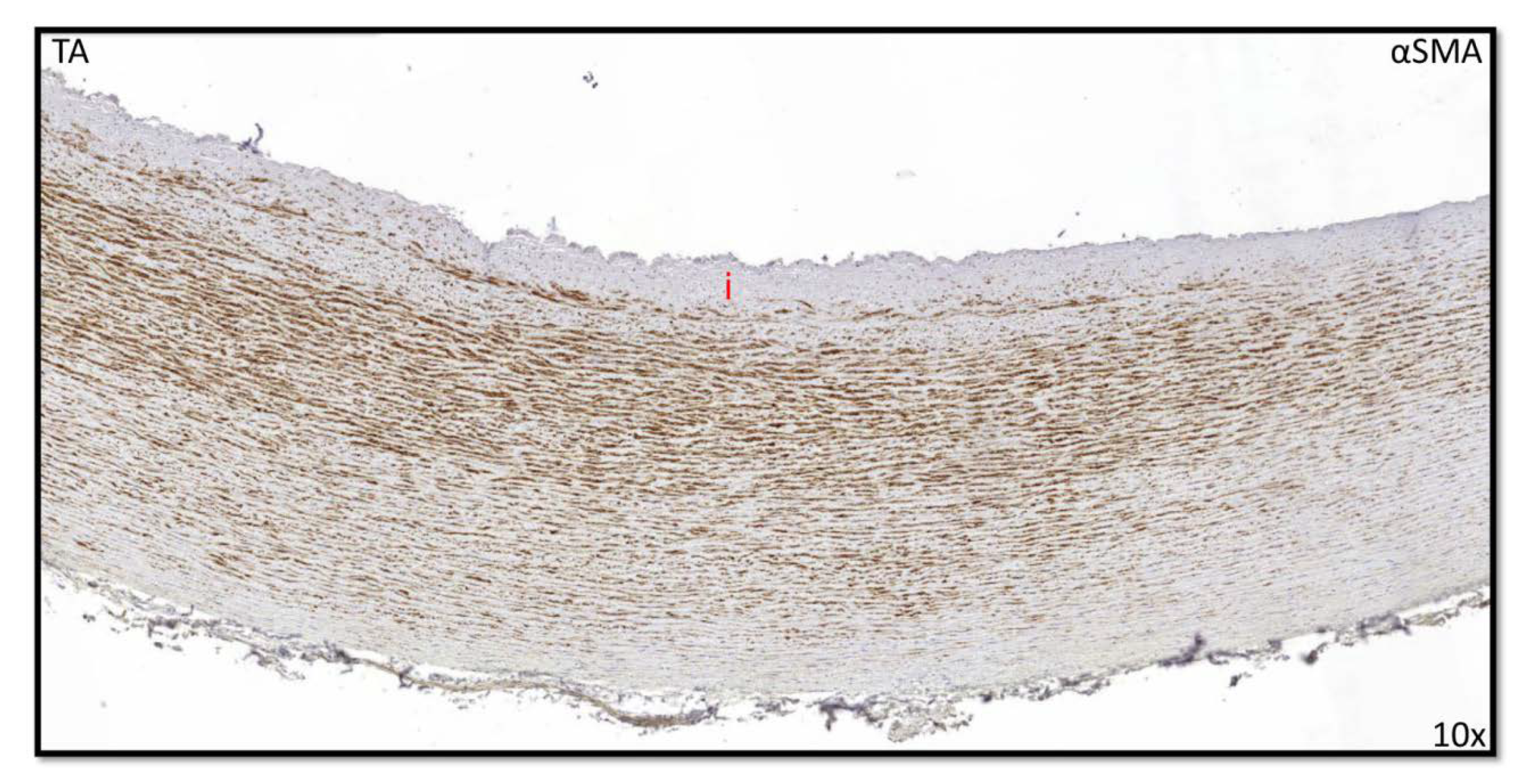
3.3. Development of the Outer Most Layer of the Ascending Aorta
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- DeRuiter, M.C.; Poelmann, R.E.; VanMunsteren, J.C.; Mironov, V.; Markwald, R.R.; Gittenberger-de Groot, A.C. Embryonic endothelial cells transdifferentiate into mesenchymal cells expressing smooth muscle Actins in vivo and in vitro. Circ. Res. 1997, 80, 444–451. [Google Scholar] [CrossRef]
- Gittenberger-de Groot, A.C.; Winter, E.M.; Bartelings, M.M.; Goumans, M.J.; DeRuiter, M.C.; Poelmann, R.E. The arterial and cardiac epicardium in development, disease and repair. Differ. Res. Biol. Divers. 2012, 84, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.G.; Brown, N.A.; Buckingham, M.E. The arterial pole of the mouse heart forms from Fgf10-expressing cells in pharyngeal mesoderm. Dev. Cell 2001, 1, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Waldo, K.L.; Hutson, M.R.; Stadt, H.A.; Zdanowicz, M.; Zdanowicz, J.; Kirby, M.L. Cardiac neural crest is necessary for normal addition of the myocardium to the arterial pole from the secondary heart field. Dev. Biol. 2005, 281, 66–77. [Google Scholar] [CrossRef] [Green Version]
- Bergwerff, M.; DeRuiter, M.C.; Poelmann, R.E.; Gittenberger-de Groot, A.C. Onset of elastogenesis and downregulation of smooth muscle actin as distinguishing phenomena in artery differentiation in the chick embryo. Anat. Embryol. 1996, 194, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Lalezari, S.; Hazekamp, M.G.; Bartelings, M.M.; Schoof, P.H.; Gittenberger-De Groot, A.C. Pulmonary artery remodeling in transposition of the great arteries: Relevance for neoaortic root dilatation. J. Thorac. Cardiovasc. Surg. 2003, 126, 1053–1060. [Google Scholar] [CrossRef] [Green Version]
- Poelmann, R.E.; Gittenberger-de Groot, A.C. Hemodynamics in cardiac development. J. Cardiovasc. Dev. Dis. 2018, 5, 54. [Google Scholar] [CrossRef] [Green Version]
- Harmon, A.W.; Nakano, A. Nkx2-5 lineage tracing visualizes the distribution of second heart field-derived aortic smooth muscle. Genesis 2013, 51, 862–869. [Google Scholar] [CrossRef] [Green Version]
- Peterson, J.C.; Chughtai, M.; Wisse, L.J.; Gittenberger-de Groot, A.C.; Feng, Q.; Goumans, M.T.H.; VanMunsteren, J.C.; Jongbloed, M.R.M.; DeRuiter, M.C. Bicuspid aortic valve formation: Nos3 mutation leads to abnormal lineage patterning of neural crest cells and the second heart field. Dis. Models Mech. 2018, 11. [Google Scholar] [CrossRef] [Green Version]
- Poelmann, R.E.; Lie-Venema, H.; Gittenberger-de Groot, A.C. The role of the epicardium and neural crest as extracardiac contributors to coronary vascular development. Tex. Heart Inst. J. 2002, 29, 255–261. [Google Scholar]
- Grewal, N.; DeRuiter, M.C.; Jongbloed, M.R.; Goumans, M.J.; Klautz, R.J.; Poelmann, R.E.; Gittenberger-de Groot, A.C. Normal and abnormal development of the aortic wall and valve: Correlation with clinical entities. Neth. Heart J. 2014, 22, 363–369. [Google Scholar] [CrossRef] [Green Version]
- Owens, G.K. Regulation of differentiation of vascular smooth muscle cells. Physiol. Rev. 1995, 75, 487–517. [Google Scholar] [CrossRef] [PubMed]
- Owens, G.K.; Kumar, M.S.; Wamhoff, B.R. Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiol. Rev. 2004, 84, 767–801. [Google Scholar] [CrossRef] [PubMed]
- Hungerford, J.E.; Little, C.D. Developmental biology of the vascular smooth muscle cell: Building a multilayered vessel wall. J. Vasc. Res. 1999, 36, 2–27. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.R.; Sinha, S.; Owens, G.K. Vascular smooth muscle cells in atherosclerosis. Circ. Res. 2016, 118, 692–702. [Google Scholar] [CrossRef]
- Gabbiani, G.; Schmid, E.; Winter, S.; Chaponnier, C.; de Ckhastonay, C.; Vandekerckhove, J.; Weber, K.; Franke, W.W. Vascular smooth muscle cells differ from other smooth muscle cells: Predominance of vimentin filaments and a specific alpha-type actin. Proc. Natl. Acad. Sci. USA 1981, 78, 298–302. [Google Scholar] [CrossRef] [Green Version]
- Van der Loop, F.T.; Gabbiani, G.; Kohnen, G.; Ramaekers, F.C.; van Eys, G.J. Differentiation of smooth muscle cells in human blood vessels as defined by smoothelin, a novel marker for the contractile phenotype. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Grewal, N.; Gittenberger-de Groot, A.C.; Poelmann, R.E.; Klautz, R.J.; Lindeman, J.H.; Goumans, M.J.; Palmen, M.; Mohamed, S.A.; Sievers, H.H.; Bogers, A.J.; et al. Ascending aorta dilation in association with bicuspid aortic valve: A maturation defect of the aortic wall. J. Thorac. Cardiovasc. Surg. 2014, 148, 1583–1590. [Google Scholar] [CrossRef] [Green Version]
- Grewal, N.; Gittenberger-de Groot, A.C.; DeRuiter, M.C.; Klautz, R.J.; Poelmann, R.E.; Duim, S.; Lindeman, J.H.; Koenraadt, W.M.; Jongbloed, M.R.; Mohamed, S.A.; et al. Bicuspid aortic valve: Phosphorylation of c-Kit and downstream targets are prognostic for future aortopathy. Eur. J. Cardiothorac. Surg. 2014, 46, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Wilens, S.L. The nature of diffuse intimal thickening of arteries. Am. J. Pathol. 1951, 27, 825–839. [Google Scholar]
- Jores, L. Arterien. Handbuch der Speziellen Pathologischen Anatomie und Histologie; Henke-Lubarsch: Berlin, Germany, 1924; pp. 686–692. [Google Scholar]
- European Medicines Agency. Reflection Paper: Formulations of Choice for the Paediatric Population; European Medicine Agency: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Grewal, N.; Gittenberger-de Groot, A.C. Pathogenesis of aortic wall complications in Marfan syndrome. Cardiovasc. Pathol. 2018, 33, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Halushka, M.K.; Angelini, A.; Bartoloni, G.; Basso, C.; Batoroeva, L.; Bruneval, P.; Buja, L.M.; Butany, J.; d’Amati, G.; Fallon, J.T.; et al. Consensus statement on surgical pathology of the aorta from the society for cardiovascular pathology and the association for european cardiovascular pathology: II. Noninflammatory degenerative diseases—Nomenclature and diagnostic criteria. Cardiovasc. Pathol. 2016, 25, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Grewal, N.; Girdauskas, E.; DeRuiter, M.; Goumans, M.J.; Poelmann, R.E.; Klautz, R.J.M.; Gittenberger-de Groot, A.C. The role of hemodynamics in bicuspid aortopathy: A histopathologic study. Cardiovasc. Pathol. 2019, 41, 29–37. [Google Scholar] [CrossRef]
- Tzemos, N.; Therrien, J.; Yip, J.; Thanassoulis, G.; Tremblay, S.; Jamorski, M.T.; Webb, G.D.; Siu, S.C. Outcomes in adults with bicuspid aortic valves. JAMA 2008, 300, 1317–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, D.Y. Intimal hyperplasia in murine models. Curr. Drug Targets 2008, 9, 251–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, S.M.; deBlois, D.; O’Brien, E.R. The intima. Soil for atherosclerosis and restenosis. Circ. Res. 1995, 77, 445–465. [Google Scholar] [CrossRef] [PubMed]
- Grewal, N.; Franken, R.; Mulder, B.J.; Goumans, M.J.; Lindeman, J.H.; Jongbloed, M.R.; DeRuiter, M.C.; Klautz, R.J.; Bogers, A.J.; Poelmann, R.E.; et al. Histopathology of aortic complications in bicuspid aortic valve versus Marfan syndrome: Relevance for therapy? Heart Vessel. 2016, 31, 795–806. [Google Scholar] [CrossRef] [Green Version]
- Nabel, E.G.; Shum, L.; Pompili, V.J.; Yang, Z.Y.; San, H.; Shu, H.B.; Liptay, S.; Gold, L.; Gordon, D.; Derynck, R.; et al. Direct transfer of transforming growth factor beta 1 gene into arteries stimulates fibrocellular hyperplasia. Proc. Natl. Acad. Sci. USA 1993, 90, 10759–10763. [Google Scholar] [CrossRef] [Green Version]
- Tsai, S.; Hollenbeck, S.T.; Ryer, E.J.; Edlin, R.; Yamanouchi, D.; Kundi, R.; Wang, C.; Liu, B.; Kent, K.C. TGF-beta through Smad3 signaling stimulates vascular smooth muscle cell proliferation and neointimal formation. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H540–H549. [Google Scholar] [CrossRef] [Green Version]
- Bokenkamp, R.; DeRuiter, M.C.; van Munsteren, C.; Gittenberger-de Groot, A.C. Insights into the pathogenesis and genetic background of patency of the ductus arteriosus. Neonatology 2010, 98, 6–17. [Google Scholar] [CrossRef]
- Slomp, J.; Gittenberger-de Groot, A.C.; Glukhova, M.A.; Conny van Munsteren, J.; Kockx, M.M.; Schwartz, S.M.; Koteliansky, V.E. Differentiation, dedifferentiation, and apoptosis of smooth muscle cells during the development of the human ductus arteriosus. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 1003–1009. [Google Scholar] [CrossRef]
- Harrison, O.J.; Visan, A.C.; Moorjani, N.; Modi, A.; Salhiyyah, K.; Torrens, C.; Ohri, S.; Cagampang, F.R. Defective NOTCH signaling drives increased vascular smooth muscle cell apoptosis and contractile differentiation in bicuspid aortic valve aortopathy: A review of the evidence and future directions. Trends Cardiovasc. Med. 2019, 29, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, S.A.; Misfeld, M.; Hanke, T.; Charitos, E.I.; Bullerdiek, J.; Belge, G.; Kuehnel, W.; Sievers, H.H. Inhibition of caspase-3 differentially affects vascular smooth muscle cell apoptosis in the concave versus convex aortic sites in ascending aneurysms with a bicuspid aortic valve. Ann. Anat. 2010, 192, 145–150. [Google Scholar] [CrossRef]
- Mimler, T.; Nebert, C.; Eichmair, E.; Winter, B.; Aschacher, T.; Stelzmueller, M.-E.; Andreas, M.; Ehrlich, M.; Laufer, G.; Messner, B. Extracellular matrix in ascending aortic aneurysms and dissections—What we learn from decellularization and scanning electron microscopy. PLoS ONE 2019, 14, e0213794. [Google Scholar] [CrossRef] [PubMed]
- Curci, J.A.; Liao, S.; Huffman, M.D.; Shapiro, S.D.; Thompson, R.W. Expression and localization of macrophage elastase (matrix metalloproteinase-12) in abdominal aortic aneurysms. J. Clin. Investig. 1998, 102, 1900–1910. [Google Scholar] [CrossRef] [Green Version]
- Jacob, M.P. Extracellular matrix remodeling and matrix metalloproteinases in the vascular wall during aging and in pathological conditions. Biomed. Pharm. 2003, 57, 195–202. [Google Scholar] [CrossRef]
- Nataatmadja, M.; West, M.; West, J.; Summers, K.; Walker, P.; Nagata, M.; Watanabe, T. Abnormal extracellular matrix protein transport associated with increased apoptosis of vascular smooth muscle cells in marfan syndrome and bicuspid aortic valve thoracic aortic aneurysm. Circulation 2003, 108 (Suppl. S1), II329–II334. [Google Scholar] [CrossRef] [Green Version]
- Fedak, P.W.; de Sa, M.P.; Verma, S.; Nili, N.; Kazemian, P.; Butany, J.; Strauss, B.H.; Weisel, R.D.; David, T.E. Vascular matrix remodeling in patients with bicuspid aortic valve malformations: Implications for aortic dilatation. J. Thorac. Cardiovasc. Surg. 2003, 126, 797–806. [Google Scholar] [CrossRef] [Green Version]
- Boyum, J.; Fellinger, E.K.; Schmoker, J.D.; Trombley, L.; McPartland, K.; Ittleman, F.P.; Howard, A.B. Matrix metalloproteinase activity in thoracic aortic aneurysms associated with bicuspid and tricuspid aortic valves. J. Thorac. Cardiovasc. Surg. 2004, 127, 686–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikonomidis, J.S.; Jones, J.A.; Barbour, J.R.; Stroud, R.E.; Clark, L.L.; Kaplan, B.S.; Zeeshan, A.; Bavaria, J.E.; Gorman, J.H., 3rd; Spinale, F.G.; et al. Expression of matrix metalloproteinases and endogenous inhibitors within ascending aortic aneurysms of patients with bicuspid or tricuspid aortic valves. J. Thorac. Cardiovasc. Surg. 2007, 133, 1028–1036. [Google Scholar] [CrossRef] [Green Version]
- LeMaire, S.A.; Wang, X.; Wilks, J.A.; Carter, S.A.; Wen, S.; Won, T.; Leonardelli, D.; Anand, G.; Conklin, L.D.; Wang, X.L.; et al. Matrix metalloproteinases in ascending aortic aneurysms: Bicuspid versus trileaflet aortic valves. J. Surg. Res. 2005, 123, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Billaud, M.; Hill, J.C.; Richards, T.D.; Gleason, T.G.; Phillippi, J.A. Medial hypoxia and adventitial vasa vasorum remodeling in human ascending aortic aneurysm. Front. Cardiovasc. Med. 2018, 5, 124. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age Categories | Tricuspid Aortic Valve N = 60 | Bicuspid Aortic Valve N = 32 |
|---|---|---|
| Premature (<38 weeks gestational age) | N = 6
| N = 5
|
| Neonate (0 < 30 days) | N = 5
| N = 6
|
| Infant (1 month < 2 years) | N = 6
| N = 6
|
| Young child (2 < 6 years) | N = 9
| N = 2
|
| Child (6 < 12 years) | N = 1
| N = 0 |
| Adolescent (12 < 18 years) | N = 22
| N = 4
|
| Young adult (18 < 21 years) | N = 5
| N = 0 |
| Adult (> 21 years) | N = 6
| N = 9
|
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grewal, N.; Gittenberger-de Groot, A.C.; von der Thusen, J.; Wisse, L.J.; Bartelings, M.M.; DeRuiter, M.C.; Klautz, R.J.M.; Poelmann, R.E. The Development of the Ascending Aortic Wall in Tricuspid and Bicuspid Aortic Valve: A Process from Maturation to Degeneration. J. Clin. Med. 2020, 9, 908. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm9040908
Grewal N, Gittenberger-de Groot AC, von der Thusen J, Wisse LJ, Bartelings MM, DeRuiter MC, Klautz RJM, Poelmann RE. The Development of the Ascending Aortic Wall in Tricuspid and Bicuspid Aortic Valve: A Process from Maturation to Degeneration. Journal of Clinical Medicine. 2020; 9(4):908. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm9040908
Chicago/Turabian StyleGrewal, Nimrat, Adriana C. Gittenberger-de Groot, Jan von der Thusen, Lambertus J. Wisse, Margot M. Bartelings, Marco C. DeRuiter, Robert J.M. Klautz, and Robert E. Poelmann. 2020. "The Development of the Ascending Aortic Wall in Tricuspid and Bicuspid Aortic Valve: A Process from Maturation to Degeneration" Journal of Clinical Medicine 9, no. 4: 908. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm9040908