Adverse Childhood Experiences and Methylation of the FKBP5 Gene in Patients with Psychotic Disorders

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Clinical Assessment
2.3. Sampling of Biological Material
2.4. Assessment of DNA Methylation
2.5. Statistics
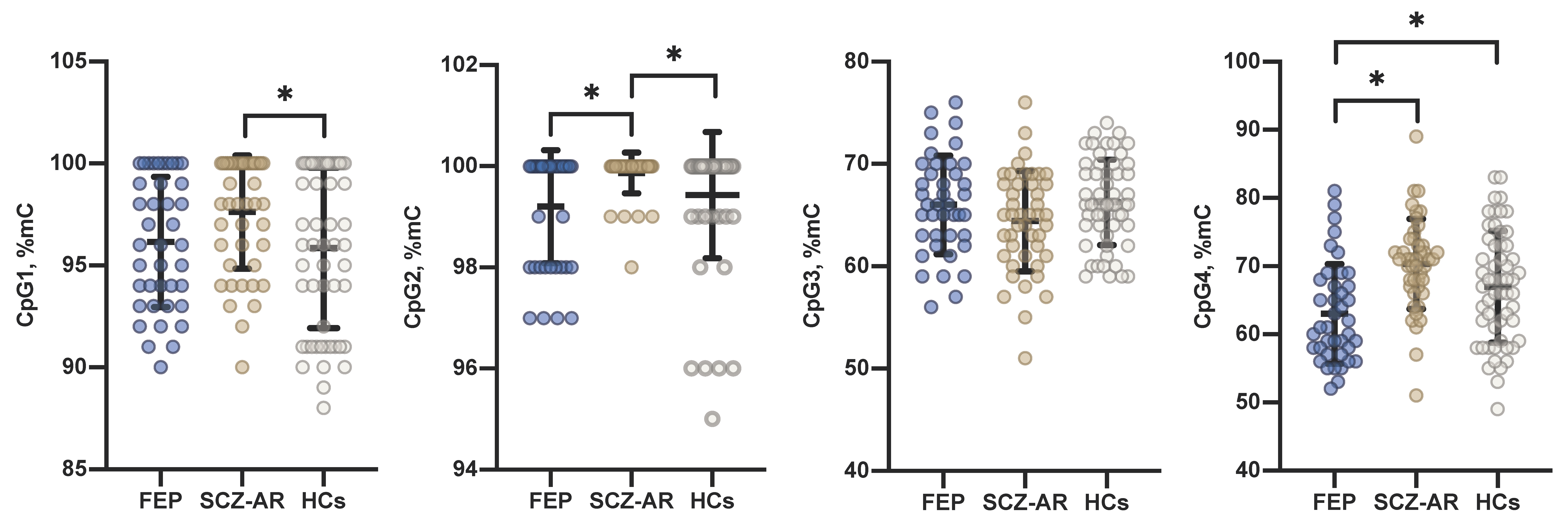
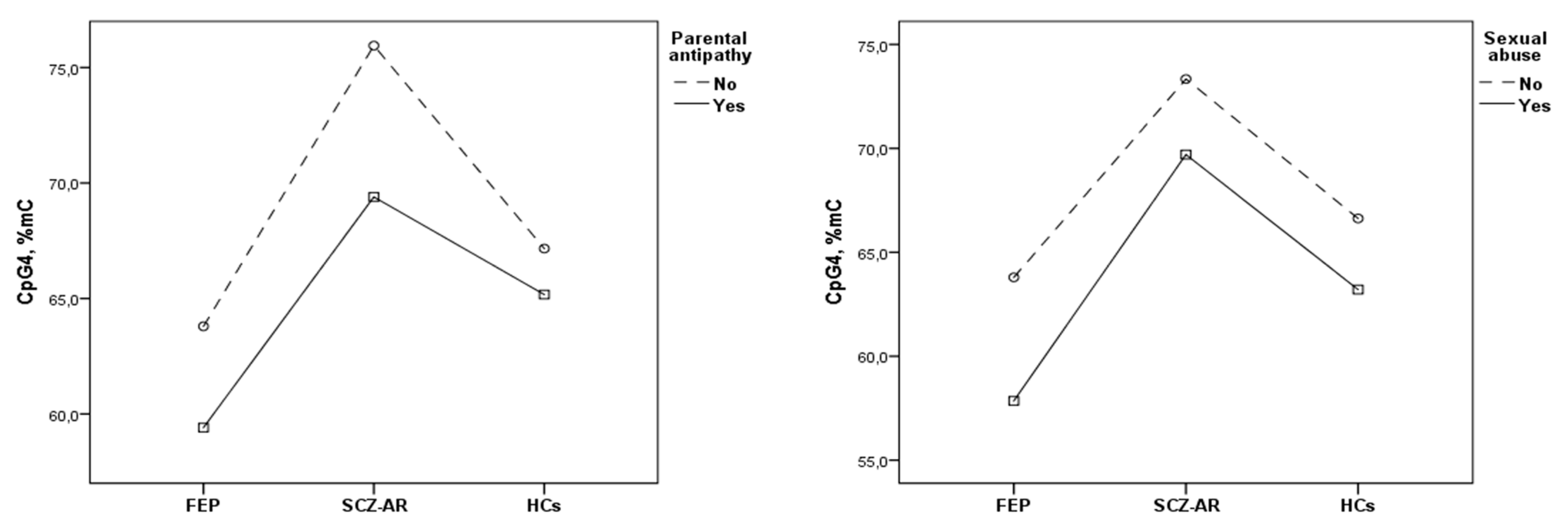
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| CpG1 | CpG2 | CpG3 | CpG4 | |
|---|---|---|---|---|
| Age | r = 0.072, p = 0.396 | r = 0.167, p = 0.048 | r = −0.092, p = 0.277 | r = 0.156, p = 0.064 |
| Sex | Males vs. females: 96.6 ± 3.6 vs. 96.4 ± 3.4, p = 0.617 | Males vs. females: 99.3 ± 1.2 vs. 99.7 ± 0.9, p = 0.048 | Males vs. females: 63.9 ± 4.2 vs. 67.1 ± 4.5, p < 0.001 | Males vs. females: 64.6 ± 6.8 vs. 69.1 ± 8.3, p = 0.002 |
| Cigarette smoking | Non−smokers vs. smokers: 96.3 ± 3.6 vs. 96.7 ± 3.0, p = 0.684 | Non−smokers vs. smokers: 99.6 ± 1.0 vs. 99.3 ± 1.2 p = 0.107 | Non−smokers vs. smokers: 66.1 ± 4.4 vs. 64.2 ± 5.0, p = 0.035 | Non−smokers vs. smokers: 67.1 ± 7.9 vs. 65.9 ± 8.0, p = 0.330 |
| BMI | r = 0.171, p = 0.047 | r = 0.070, p = 0.421 | r = 0.212, p = 0.013 | r = 0.212, p = 0.013 |
| CPZeq | r = 0.139, p = 0.227 | r = 0.166, p = 0.149 | r = 0.068, p = 0.232 | r = 0.262, p = 0.022 |
| Cortisol | r = 0.167, p = 0.070 | r = 0.267, p = 0.003 | r = −0.198, p = 0.031 | r = 0.086, p = 0.355 |
References
- Janssen, I.; Krabbendam, L.; Bak, M.; Hanssen, M.; Vollebergh, W.; De Graaf, R.; Van Os, J. Childhood abuse as a risk factor for psychotic experiences. Acta Psychiatr. Scand. 2004, 109, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Kessler, R.C.; McLaughlin, K.A.; Green, J.G.; Gruber, M.J.; Sampson, N.A.; Zaslavsky, A.M.; Aguilar-Gaxiola, S.; Alhamzawi, A.O.; Alonso, J.; Angermeyer, M.; et al. Childhood adversities and adult psychopathology in the WHO world mental health surveys. Br. J. Psychiatry 2010, 197, 378–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varese, F.; Smeets, F.; Drukker, M.; Lieverse, R.; Lataster, T.; Viechtbauer, W.; Read, J.; Van Os, J.; Bentall, R.P. Childhood adversities increase the risk of psychosis: A meta-analysis of patient-control, prospective-and cross-sectional cohort studies. Schizophr. Bull. 2012, 38, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Muenzenmaier, K.H.; Seixas, A.A.; Schneeberger, A.R.; Castille, D.M.; Battaglia, J.; Link, B.G. Cumulative Effects of Stressful Childhood Experiences on Delusions and Hallucinations. J. Trauma Dissociation 2015, 16, 442–462. [Google Scholar] [CrossRef] [PubMed]
- Misiak, B.; Moustafa, A.A.; Kiejna, A.; Frydecka, D. Childhood traumatic events and types of auditory verbal hallucinations in first-episode schizophrenia patients. Compr. Psychiatry 2016, 66, 17–22. [Google Scholar] [CrossRef]
- Vargas, T.; Lam, P.H.; Azis, M.; Osborne, K.J.; Lieberman, A.; Mittal, V.A. Childhood trauma and neurocognition in adults with psychotic disorders: A systematic review and meta-analysis. Schizophr. Bull. 2019, 45, 1195–1208. [Google Scholar] [CrossRef]
- Aas, M.; Andreassen, O.A.; Aminoff, S.R.; Færden, A.; Romm, K.L.; Nesvåg, R.; Berg, A.O.; Simonsen, C.; Agartz, I.; Melle, I. A history of childhood trauma is associated with slower improvement rates: Findings from a one-year follow-up study of patients with a first-episode psychosis. BMC Psychiatry 2016, 16, 126. [Google Scholar] [CrossRef] [Green Version]
- Misiak, B.; Frydecka, D. A history of childhood trauma and response to treatment with antipsychotics in first-episode schizophrenia patients. J. Nerv. Ment. Dis. 2016, 204, 787–792. [Google Scholar] [CrossRef]
- Veling, W.; Counotte, J.; Pot-Kolder, R.; van Os, J.; van der Gaag, M. Childhood trauma, psychosis liability and social stress reactivity: A virtual reality study. Psychol. Med. 2016, 46, 3339–3348. [Google Scholar] [CrossRef]
- Aas, M.; Pizzagalli, D.A.; Laskemoen, J.F.; Reponen, E.J.; Ueland, T.; Melle, I.; Agartz, I.; Steen, N.E.; Andreassen, O.A. Elevated hair cortisol is associated with childhood maltreatment and cognitive impairment in schizophrenia and in bipolar disorders. Schizophr. Res. 2019, 213, 65–71. [Google Scholar] [CrossRef]
- Lovallo, W.R.; Farag, N.H.; Sorocco, K.H.; Cohoon, A.J.; Vincent, A.S. Lifetime adversity leads to blunted stress axis reactivity: Studies from the oklahoma family health patterns project. Biol. Psychiatry 2012, 71, 344–349. [Google Scholar] [CrossRef] [Green Version]
- Holtzman, C.W.; Trotman, H.D.; Goulding, S.M.; Ryan, A.T.; MacDonald, A.N.; Shapiro, D.I.; Brasfield, J.L.; Walker, E.F. Stress and neurodevelopmental processes in the emergence of psychosis. Neuroscience 2013, 249, 172–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marinelli, M.; Rudick, C.N.; Hu, X.-T.; White, F.J. Excitability of dopamine neurons: Modulation and physiological consequences. CNS Neurol. Disord. Drug Targets 2006, 5, 79–97. [Google Scholar] [PubMed]
- Nordholm, D.; Krogh, J.; Mondelli, V.; Dazzan, P.; Pariante, C.; Nordentoft, M. Pituitary gland volume in patients with schizophrenia, subjects at ultra high-risk of developing psychosis and healthy controls: A systematic review and meta-analysis. Psychoneuroendocrinology 2013, 38, 2394–2404. [Google Scholar] [CrossRef] [PubMed]
- Girshkin, L.; Matheson, S.L.; Shepherd, A.M.; Green, M.J. Morning cortisol levels in schizophrenia and bipolar disorder: A meta-analysis. Psychoneuroendocrinology 2014, 49, 187–206. [Google Scholar] [CrossRef] [PubMed]
- Berger, M.; Kraeuter, A.K.; Romanik, D.; Malouf, P.; Amminger, G.P.; Sarnyai, Z. Cortisol awakening response in patients with psychosis: Systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2016, 68, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Ciufolini, S.; Dazzan, P.; Kempton, M.J.; Pariante, C.; Mondelli, V. HPA axis response to social stress is attenuated in schizophrenia but normal in depression: Evidence from a meta-analysis of existing studies. Neurosci. Biobehav. Rev. 2014, 47, 359–368. [Google Scholar] [CrossRef]
- Zorn, J.V.; Schür, R.R.; Boks, M.P.; Kahn, R.S.; Joëls, M.; Vinkers, C.H. Cortisol stress reactivity across psychiatric disorders: A systematic review and meta-analysis. Psychoneuroendocrinology 2017, 77, 25–36. [Google Scholar] [CrossRef]
- Cullen, A.E.; Rai, S.; Vaghani, M.S.; Mondelli, V.; McGuire, P. Cortisol responses to naturally occurring psychosocial stressors across the psychosis spectrum: A systematic review and meta-analysis. Front. Psychiatry 2020, 11, 513. [Google Scholar] [CrossRef]
- Misiak, B.; Stramecki, F.; Gawęda, Ł.; Prochwicz, K.; Sąsiadek, M.M.; Moustafa, A.A.; Frydecka, D. Interactions between variation in candidate genes and environmental factors in the etiology of schizophrenia and bipolar disorder: A systematic review. Mol. Neurobiol. 2017, 55, 5075–5100. [Google Scholar] [CrossRef]
- Tomassi, S.; Tosato, S. Epigenetics and gene expression profile in first-episode psychosis: The role of childhood trauma. Neurosci. Biobehav. Rev. 2017, 83, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Klengel, T.; Pape, J.; Binder, E.B.; Mehta, D. The role of DNA methylation in stress-related psychiatric disorders. Neuropharmacology 2014, 80, 1115–1132. [Google Scholar] [CrossRef] [PubMed]
- Matosin, N.; Halldorsdottir, T.; Binder, E.B. Understanding the molecular mechanisms underpinning gene by environment interactions in psychiatric disorders: The FKBP5 model. Biol. Psychiatry 2018, 83, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Zannas, A.S.; Wiechmann, T.; Gassen, N.C.; Binder, E.B. Gene-stress-epigenetic regulation of FKBP5: Clinical and translational implications. Neuropsychopharmacology 2016, 41, 261–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binder, E.B. The role of FKBP5, a co-chaperone of the glucocorticoid receptor in the pathogenesis and therapy of affective and anxiety disorders. Psychoneuroendocrinology 2009, 34, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Mihaljevic, M.; Zeljic, K.; Soldatovic, I.; Andric, S.; Mirjanic, T.; Richards, A.; Mantripragada, K.; Pekmezovic, T.; Novakovic, I.; Maric, N.P. The emerging role of the FKBP5 gene polymorphisms in vulnerability–stress model of schizophrenia: Further evidence from a Serbian population. Eur. Arch. Psychiatry Clin. Neurosci. 2017, 267, 527–539. [Google Scholar] [CrossRef] [Green Version]
- Collip, D.; Myin-Germeys, I.; Wichers, M.; Jacobs, N.; Derom, C.; Thiery, E.; Lataster, T.; Simons, C.; Delespaul, P.; Marcelis, M.; et al. FKBP5 as a possible moderator of the psychosis-inducing effects of childhood trauma. Br. J. Psychiatry 2013, 202, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Green, M.J.; Raudino, A.; Cairns, M.J.; Wu, J.; Tooney, P.A.; Scott, R.J.; Carr, V.J. Do common genotypes of FK506 binding protein 5 (FKBP5) moderate the effects of childhood maltreatment on cognition in schizophrenia and healthy controls? J. Psychiatr. Res. 2015, 70, 9–17. [Google Scholar] [CrossRef]
- Weder, N.; Zhang, H.; Jensen, K.; Yang, B.Z.; Simen, A.; Jackowski, A.; Lipschitz, D.; Douglas-Palumberi, H.; Ge, M.; Perepletchikova, F.; et al. Child abuse, depression, and methylation in genes involved with stress, neural plasticity, and brain circuitry. J. Am. Acad. Child Adolesc. Psychiatry 2014, 53, 417–424. [Google Scholar] [CrossRef] [Green Version]
- Klengel, T.; Mehta, D.; Anacker, C.; Rex-Haffner, M.; Pruessner, J.C.; Pariante, C.M.; Pace, T.W.W.; Mercer, K.B.; Mayberg, H.S.; Bradley, B.; et al. Allele-specific FKBP5 DNA demethylation mediates gene-childhood trauma interactions. Nat. Neurosci. 2013, 16, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Sinclair, D.; O’Donnell, M.; Galletly, C.; Liu, D.; Weickert, C.S.; Weickert, T.W. Transcriptional changes in the stress pathway are related to symptoms in schizophrenia and to mood in schizoaffective disorder. Schizophr. Res. 2019, 213, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Misiak, B.; Kotowicz, K.; Loska, O.; Stramecki, F.; Beszłej, J.A.; Samochowiec, J.; Jabłoński, M.; Podwalski, P.; Waszczuk, K.; Wroński, M.; et al. Decreased use of active coping styles contributes to elevated allostatic load index in first-episode psychosis. Psychoneuroendocrinology 2018, 96, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Piotrowski, P.; Frydecka, D.; Kotowicz, K.; Stańczykiewicz, B.; Samochowiec, J.; Szczygieł, K.; Misiak, B. A history of childhood trauma and allostatic load in patients with psychotic disorders with respect to stress coping strategies. Psychoneuroendocrinology 2020, 115, 104645. [Google Scholar] [CrossRef] [PubMed]
- Piotrowski, P.; Kotowicz, K.; Rymaszewska, J.; Beszłej, J.A.; Plichta, P.; Samochowiec, J.; Kalinowska, S.; Trześniowska-Drukała, B.; Misiak, B. Allostatic load index and its clinical correlates at various stages of psychosis. Schizophr. Res. 2019, 210, 73–80. [Google Scholar] [CrossRef]
- McGuffin, P.; Farmer, A.; Harvey, I. A polydiagnostic application of operational criteria in studies of psychotic illness. Arch. Gen. Psychiatry 1991, 48, 764–770. [Google Scholar] [CrossRef]
- Kay, S.R.; Fiszbein, A.; Opler, L.A. The positive and negative syndrome scale (PANSS) for schizophrenia. Schizophr. Bull. 1987, 13, 261–276. [Google Scholar] [CrossRef]
- Montgomery, S.A.; Asberg, M. A new depression scale designed to be sensitive to change. Br. J. Psychiatry 1979, 134, 382–389. [Google Scholar] [CrossRef]
- Young, R.C.; Biggs, J.T.; Ziegler, V.E.; Meyer, D.A. A rating scale for mania: Reliability, validity and sensitivity. Br. J. Psychiatry. 1978, 133, 429–435. [Google Scholar] [CrossRef]
- Hall, R.C.W. Global assessment of functioning. Psychosomatics 1995, 36, 267–275. [Google Scholar] [CrossRef]
- Randolph, C.; Tierney, M.C.; Mohr, E.; Chase, T.N. The Repeatable Battery for the Assessment of Neuropsychological Status (RBANS): Preliminary clinical validity. J. Clin. Exp. Neuropsychol. 1998, 20, 310–319. [Google Scholar] [CrossRef]
- Bifulco, A.; Bernazzani, O.; Moran, P.M.; Jacobs, C. The childhood experience of care and abuse questionnaire (CECA.Q): Validation in a community series. Br. J. Clin. Psychol. 2005, 44, 563–581. [Google Scholar] [CrossRef] [PubMed]
- Fisher, H.L.; Craig, T.K.; Fearon, P.; Morgan, K.; Dazzan, P.; Lappin, J.; Hutchinson, G.; Doody, G.A.; Jones, P.B.; McGuffin, P.; et al. Reliability and comparability of psychosis patients’ retrospective reports of childhood abuse. Schizophr. Bull. 2011, 37, 546–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiechmann, T.; Roh, S.; Sauer, S.; Czamara, D.; Arloth, J.; Kodl, M.; Beintner, M.; Knop, L.; Binder, E.B.; Provencal, N. Identification of dynamic glucocorticoid-induced methylation changes at the FKBP5 locus. Clin. Epigenet. 2019, 11, 83. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.S.; Mahon, P.B.; Zandi, P.P.; McCaul, M.E.; Yang, X.; Bali, U.; Wand, G.S. DNA methylation and sex-specific expression of FKBP5 as correlates of one-month bedtime cortisol levels in healthy individuals. Psychoneuroendocrinology 2018, 97, 164–173. [Google Scholar] [CrossRef]
- Ovenden, E.S.; McGregor, N.W.; Emsley, R.A.; Warnich, L. DNA methylation and antipsychotic treatment mechanisms in schizophrenia: Progress and future directions. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 81, 38–49. [Google Scholar] [CrossRef]
- Tyrka, A.R.; Ridout, K.K.; Parade, S.H.; Marsit, C.J.; Seifer, R. childhood maltreatment and methylation of FK506 binding protein 5 gene (FKBP5). Dev. Psychopathol. 2015, 27, 1637–1645. [Google Scholar] [CrossRef] [Green Version]
- Parade, S.H.; Parent, J.; Rabemananjara, K.; Seifer, R.; Marsit, C.J.; Yang, B.Z.; Zhang, H.; Tyrka, A.R. Change in FK506 binding protein 5 (FKBP5) methylation over time among preschoolers with adversity. Dev. Psychopathol. 2017, 29, 1627–1634. [Google Scholar] [CrossRef]
- Saito, T.; Shinozaki, G.; Koga, M.; Tanichi, M.; Takeshita, S.; Nakagawa, R.; Nagamine, M.; Cho, H.R.; Morimoto, Y.; Kobayashi, Y.; et al. Effect of interaction between a specific subtype of child abuse and the FKBP5 rs1360780 SNP on DNA methylation among patients with bipolar disorder. J. Affect. Disord. 2020, 272, 417–422. [Google Scholar] [CrossRef]
- Grasso, D.J.; Drury, S.; Briggs-Gowan, M.; Johnson, A.; Ford, J.; Lapidus, G.; Scranton, V.; Abreu, C.; Covault, J. Adverse childhood experiences, posttraumatic stress, and FKBP5 methylation patterns in postpartum women and their newborn infants. Psychoneuroendocrinology 2020, 114, 104604. [Google Scholar] [CrossRef]
- Thomassin, H.; Flavin, M.; Espinas, M.L.; Grange, T. Glucocorticoid-induced DNA demethylation and gene memory during development. EMBO J. 2001, 20, 1974–1983. [Google Scholar] [CrossRef]
- Stramecki, F.; Misiak, B.; Frydecka, D. The role of the interaction between the FKBP5 gene and stressful life events in the pathophysiology of schizophrenia: A narrative review. Arch. Psychiatry Psychother. 2020, 3, 7–16. [Google Scholar] [CrossRef]
- Alexander, N.; Kirschbaum, C.; Stalder, T.; Muehlhan, M.; Vogel, S. No association between FKBP5 gene methylation and acute and long-term cortisol output. Transl. Psychiatry 2020, 10, 175. [Google Scholar] [CrossRef] [PubMed]
- Winkler, B.K.; Lehnert, H.; Oster, H.; Kirchner, H.; Harbeck, B. FKBP5 methylation as a possible marker for cortisol state and transient cortisol exposure in healthy human subjects. Epigenomics 2017, 9, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Mak, M.; Samochowiec, J.; Frydecka, D.; Pełka-Wysiecka, J.; Szmida, E.; Karpiński, P.; Sąsiadek, M.M.; Piotrowski, P.; Samochowiec, A.; Misiak, B. First-episode schizophrenia is associated with a reduction of HERV-K methylation in peripheral blood. Psychiatry Res. 2019, 271, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Misiak, B.; Frydecka, D.; Loska, O.; Moustafa, A.A.; Samochowiec, J.; Kasznia, J.; Stańczykiewicz, B. Testosterone, DHEA and DHEA-S in patients with schizophrenia: A systematic review and meta-analysis. Psychoneuroendocrinology 2018, 89, 92–102. [Google Scholar] [CrossRef]
- Sinclair, D.; Fillman, S.G.; Webster, M.J.; Weickert, C.S. Dysregulation of glucocorticoid receptor co-factors FKBP5, BAG1 and PTGES3 in prefrontal cortex in psychotic illness. Sci. Rep. 2013, 3, 3539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darby, M.M.; Yolken, R.H.; Sabunciyan, S. Consistently altered expression of gene sets in postmortem brains of individuals with major psychiatric disorders. Transl. Psychiatry 2016, 6, e890. [Google Scholar] [CrossRef]
- Szabó, C.; Kelemen, O.; Kéri, S. Changes in FKBP5 expression and memory functions during cognitive-behavioral therapy in posttraumatic stress disorder: A preliminary study. Neurosci. Lett. 2014, 569, 116–120. [Google Scholar] [CrossRef] [Green Version]
- Levy-Gigi, E.; Szabó, C.; Kelemen, O.; Kéri, S. Association among clinical response, hippocampal volume, and FKBP5 gene expression in individuals with posttraumatic stress disorder receiving cognitive behavioral therapy. Biol. Psychiatry 2013, 74, 793–800. [Google Scholar] [CrossRef] [Green Version]
- Blair, L.J.; Criado-Marrero, M.; Zheng, D.; Wang, X.; Kamath, S.; Nordhues, B.A.; Weeber, E.J.; Dickey, C.A. The disease-associated chaperone FKBP51 impairs cognitive function by accelerating AMPA receptor recycling. eNeuro 2019, 6. [Google Scholar] [CrossRef]
- Sabbagh, J.J.; O’Leary, J.C.; Blair, L.J.; Klengel, T.; Nordhues, B.A.; Fontaine, S.N.; Binder, E.B.; Dickey, C.A. Age-associated epigenetic upregulation of the FKBP5 gene selectively impairs stress resiliency. PLoS ONE 2014, 9, e107241. [Google Scholar] [CrossRef] [PubMed]



| 1. FEP | 2. SCZ-AR | 3. HCs | p | Post-Hoc or Pairwise Comparisons | ||||
|---|---|---|---|---|---|---|---|---|
| n | Mean ± SD or n (%) | n | Mean ± SD or n (%) | n | Mean ± SD or n (%) | |||
| Age, years | 40 | 28.1 ± 7.3 | 45 | 45.2 ± 12.6 | 56 | 32.5 ± 8.2 | <0.001 | 1 < 2, 2 > 3 |
| Sex, males | 40 | 20 (50.0) | 45 | 25 (55.6) | 56 | 22 (39.3) | 0.248 | - |
| Education, years | 40 | 13.6 ± 2.5 | 38 | 12.6 ± 3.0 | 54 | 15.8 ± 2.5 | <0.001 | 1 < 3, 2 < 3 |
| Maternal education, higher | 40 | 8 (20.0) | 45 | 8 (17.8) | 53 | 18 (34.0) | 0.130 | - |
| Paternal education, higher | 40 | 8 (20.0) | 45 | 8 (17.8) | 53 | 15 (28.3) | 0.418 | - |
| BMI, kg/m2 | 40 | 23.7 ± 3.8 | 40 | 26.5 ± 5.1 | 56 | 23.8 ± 3.5 | 0.005 | 1 < 2, 2 > 3 |
| Cigarette smoking | 40 | 15 (37.5) | 40 | 22 (55.0) | 56 | 5 (8.9) | <0.001 | 1 < 2, 1 > 3, 2 > 3 |
| Parental loss | 37 | 10 (27.0) | 36 | 13 (36.1) | 54 | 12 (22.2) | 0.351 | - |
| Parental antipathy | 37 | 10 (27.0) | 36 | 18 (50.0) | 54 | 16 (57.1) | 0.071 | - |
| Parental neglect | 37 | 6 (16.2) | 36 | 13 (36.1) | 54 | 14 (25.9) | 0.153 | - |
| Physical abuse | 37 | 13 (35.1) | 36 | 17 (47.2) | 54 | 13 (24.1) | 0.074 | - |
| Sexual abuse | 37 | 5 (13.5) | 36 | 7 (19.4) | 54 | 3 (5.6) | 0.126 | - |
| RBANS–immediate memory | 40 | 42.7 ± 8.4 | 44 | 33.5 ± 11.3 | 52 | 51.9 ± 6.0 | <0.001 | 1 < 3, 1 > 2, 2 < 3 |
| RBANS–visuospatial/constructional | 40 | 34.7 ± 5.4 | 44 | 30.0 ± 8.2 | 52 | 38.1 ± 2.2 | <0.001 | 1 < 3, 1 > 2, 2 < 3 |
| RBANS–language | 40 | 28.2 ± 6.1 | 44 | 24.9 ± 6.6 | 52 | 33.7 ± 6.5 | <0.001 | 1 < 3, 2 < 3 |
| RBANS–attention | 40 | 54.2 ± 12.2 | 44 | 35.6 ± 11.8 | 52 | 68.9 ± 8.9 | <0.001 | 1 < 3, 1 > 2, 2 < 3 |
| RBANS–delayed memory | 40 | 46.9 ± 7.7 | 44 | 39.0 ± 11.3 | 52 | 56.0 ± 4.5 | <0.001 | 1 < 3, 1 > 2, 2 < 3 |
| Age of psychosis onset, years | 40 | 26.6 ± 7.3 | 45 | 24.9 ± 8.6 | - | - | 0.109 | - |
| Illness duration, weeks | 40 | 43.8 ± 87.8 | 45 | 651.7 ± 526.9 | - | - | <0.001 | - |
| Family history of psychosis | 40 | 5 (12.5) | 45 | 12 (26.7) | 56 | 0 (0) | <0.001 | 1 > 3, 2 > 3 |
| PANSS-P | 40 | 12.9 ± 5.2 | 40 | 15.2 ± 4.9 | - | - | 0.053 | - |
| PANSS-N | 40 | 18.1 ± 8.4 | 40 | 23.8 ± 9.5 | - | - | <0.001 | - |
| MADRS | 40 | 8.3 ± 8.1 | 38 | 7.8 ± 8.3 | - | - | 0.743 | - |
| YMRS | 40 | 2.1 ± 5.1 | 38 | 2.1 ± 5.0 | - | - | 0.758 | - |
| GAF | 40 | 54.2 ± 17.1 | 39 | 35.3 ± 14.0 | - | - | <0.001 | - |
| CPZeq, mg/day | 40 | 300.1 ± 169.7 | 37 | 467.7 ± 219.8 | - | - | <0.001 | - |
| Cortisol, nmol/l | 40 | 338.6 73.3 | 45 | 448.9 151.9 | 55 | 272.8 ± 87.2 | <0.001 | 1 < 2, 1 > 3, 2 > 3 |
| FKBP5, %mC | Independent Variable | Parental Loss | Parental Antipathy | Parental Neglect | Physical Abuse | Sexual Abuse |
|---|---|---|---|---|---|---|
| CpG1 | BMI | F = 3.380, p = 0.068 | F = 3.336, p = 0.070 | F = 3.561, p = 0.062 | F = 3.263, p = 0.073 | F = 3.030, p = 0.084 |
| Group | F = 1.749, p = 0.178 | F = 1.765, p = 0.176 | F = 2.702, p = 0.071 | F = 2.239, p = 0.111 | F = 0.422, p = 0.657 | |
| ACEs | F = 1.076, p = 0.302 | F = 0.007, p = 0.934 | F = 0.536, p = 0.466 | F = 0.489, p = 0.486 | F = 0.854, p = 0.357 | |
| Group × ACEs | F = 0.463, p = 0.631 | F = 0.374, p = 0.689 | F = 1.755, p = 0.177 | F = 0.710, p = 0.494 | F = 0.067, p = 0.935 | |
| CpG2 | Age | F = 0.207, p = 0.650 | F = 0.033, p = 0.856 | F = 0.465, p = 0.497 | F = 0.364, p = 0.548 | F = 0.339, p = 0.562 |
| Sex | F = 2.521, p = 0.115 | F = 2.616, p = 0.109 | F = 3.025, p = 0.085 | F = 2.780, p = 0.099 | F = 3.024, p = 0.085 | |
| Cortisol | F = 3.825, p = 0.053 | F = 3.327, p = 0.071 | F = 3.834, p = 0.052 | F = 3.601, p = 0.061 | F = 3.932, p = 0.058 | |
| Group | F = 0.875, p = 0.420 | F = 0.465, p = 0.629 | F = 1.267, p = 0.286 | F = 1.025, p = 0.362 | F = 1.305, p = 0.276 | |
| ACEs | F = 0.018, p = 0.894 | F = 0.011, p = 0.916 | F = 0.098, p = 0.755 | F = 0.091, p = 0.764 | F = 0.185, p = 0.668 | |
| Group × ACEs | F = 1.243, p = 0.293 | F = 1.882, p = 0.158 | F = 0.153, p = 0.858 | F = 1.518, p = 0.224 | F = 0.867, p = 0.423 | |
| CpG3 | Sex | F = 19.934, p < 0.001 | F = 21.481, p < 0.001 | F = 20.618, p < 0.001 | F = 22.292, p < 0.001 | F = 21.116, p < 0.001 |
| BMI | F = 1.988, p = 0.162 | F = 2.744, p = 0.101 | F = 1.601, p = 0.209 | F = 1.831, p = 0.179 | F = 2.056, p = 0.155 | |
| Cigarette smoking | F = 0.446, p = 0.506 | F = 0.528, p = 0.469 | F = 0.509, p = 0.477 | F = 0.531, p = 0.468 | F = 0.235, p = 0.629 | |
| Cortisol | F = 2.344, p = 0.129 | F = 2.523, p = 0.115 | F = 2.546, p = 0.114 | F = 2.423, p = 0.123 | F = 2.316, p = 0.131 | |
| Group | F = 0.669, p = 0.514 | F = 0.403, p = 0.670 | F = 0.384, p = 0.682 | F = 0.592, p = 0.555 | F = 0.094, p = 0.911 | |
| ACEs | F = 0.519, p = 0.473 | F = 0.734, p = 0.394 | F = 0.022, p = 0.883 | F = 0.029, p = 0.864 | F = 1.581, p = 0.211 | |
| Group × ACEs | F = 0.253, p = 0.777 | F = 0.047, p = 0.954 | F = 0.669, p = 0.514 | F = 0.935, p = 0.396 | F = 1.104, p = 0.336 | |
| CpG4 | Sex | F = 22.416, p < 0.001 | F = 25.458, p < 0.001 | F = 21.119, p < 0.001 | F = 25.114, p < 0.001 | F = 26.495, p < 0.001 |
| BMI | F = 4.020, p = 0.047 | F = 6.733,p= 0.011 | F = 3.813, p = 0.053 | F = 4.038, p = 0.047 | F = 4.556, p = 0.035 | |
| CPZeq | F = 0.004, p = 0.952 | F = 0.372, p = 0.543 | F = 0.078, p = 0.780 | F = 0.401, p = 0.528 | F = 0.009, p = 0.923 | |
| Group | F = 6.247, p = 0.003 | F = 8.302, p < 0.001 | F = 6.147, p = 0.003 | F = 6.929, p = 0.001 | F = 5.994, p = 0.003 | |
| ACEs | F = 1.040, p = 0.310 | F = 5.956, p = 0.016 | F = 0.134, p = 0.715 | F = 0.023, p = 0.880 | F = 5.470, p = 0.021 | |
| Group × ACEs | F = 1.430, p = 0.244 | F = 0.269, p = 0.765 | F = 0.195, p = 0.823 | F = 2.164, p = 0.120 | F = 0.068, p = 0.934 |
| FEP | SCZ-AR | HCs | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CpG1 | CpG2 | CpG3 | CpG4 | CpG1 | CpG2 | CpG3 | CpG4 | CpG1 | CpG2 | CpG3 | CpG4 | |
| RBANS–immediate memory | r = −0.198 p = 0.222 | r = −0.354 p = 0.025 | r = −0.039 p = 0.810 | r = −0.310 p = 0.049 | r = −0.224 p = 0.143 | r = −0.398 p = 0.008 | r = −0.029 p = 0.850 | r = −0.480 p = 0.001 | r = 0.150 p = 0.287 | r = 0.081 p = 0.570 | r = 0.037 p = 0.795 | r = 0.289 p = 0.038 |
| RBANS–visuospatial/constructional | r = 0.120 p = 0.461 | r = −0.442 p = 0.004 | r = −0.007 p = 0.964 | r = −0.128 p = 0.430 | r = 0.218 p = 0.156 | r = 0.012 p = 0.940 | r = −0.061 p = 0.695 | r = 0.122 p = 0.431 | r = 0.023 p = 0.872 | r = 0.088 p = 0.536 | r = −0.250 p = 0.073 | r = 0.102 p = 0.473 |
| RBANS–language | r = 0.052 p = 0.748 | r = −0.259 p = 0.106 | r = 0.264 p = 0.100 | r = 0.074 p = 0.649 | r = −0.231 p = 0.132 | r = −0.013 p = 0.932 | r = −0.133 p = 0.389 | r = −0.091 p = 0.557 | r = −0.001 p = 0.993 | r = −0.101 p = 0.477 | r = 0.227 p = 0.106 | r = 0.277 p = 0.047 |
| RBANS–attention | r = 0.172 p = 0.289 | r = −0.353 p = 0.026 | r = 0.050 p = 0.758 | r = −0.348 p = 0.028 | r = −0.173 p = 0.261 | r = −0.464 p = 0.001 | r = 0.074 p = 0.634 | r = −0.452 p = 0.002 | r = −0.027 p = 0.847 | r = −0.146 p = 0.303 | r = 0.243 p = 0.083 | r = 0.170 p = 0.230 |
| RBANS–delayed memory | r = 0.050 p = 0.760 | r = −0.340 p = 0.032 | r = −0.165 p = 0.308 | r = −0.340 p = 0.032 | r = 0.006 p = 0.970 | r = −0.333 p = 0.027 | r = −0.061 p = 0.693 | r = −0.460 p = 0.002 | r = 0.149 p = 0.291 | r = 0.140 p = 0.322 | r = −0.017 p = 0.907 | r = 0.250 p = 0.074 |
| PANSS−P | r = 0.043 p = 0.791 | r = 0.147 p = 0.366 | r = −0.070 p = 0.667 | r = 0.051 p = 0.756 | r = 0.103 p = 0.526 | r = 0.262 p = 0.102 | r = 0.039 p = 0.813 | r = 0.014 p = 0.930 | − | − | − | − |
| PANSS−N | r = 0.079 p = 0.627 | r = −0.104 p = 0.525 | r = −0.160 p = 0.323 | r = −0.129 p = 0.427 | r = 0.014 p = 0.993 | r = 0.042 p = 0.799 | r = 0.087 p = 0.595 | r = −0.087 p = 0.594 | − | − | − | − |
| MADRS | r = 0.175 p = 0.280 | r = 0.057 p = 0.725 | r = −0.135 p = 0.406 | r = 0.003 p = 0.986 | r = 0.253 p = 0.126 | r = 0.256 p = 0.121 | r = 0.087 p = 0.605 | r = 0.007 p = 0.966 | − | − | − | − |
| YMRS | r = −0.019 p = 0.908 | r = 0.166 p = 0.306 | r = 0.039 p = 0.812 | r = 0.079 p = 0.626 | r = 0.265 p = 0.107 | r = 0.033 p = 0.845 | r = 0.052 p = 0.755 | r = 0.192 p = 0.248 | − | − | − | − |
| GAF | r = 0.165 p = 0.310 | r = −0.442 p = 0.004 | r = 0.053 p = 0.746 | r = −0.346 p = 0.029 | r = 0.041 p = 0.805 | r = −0.471 p = 0.003 | r = −0.175 p = 0.287 | r = −0.314 p = 0.048 | − | − | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Misiak, B.; Karpiński, P.; Szmida, E.; Grąźlewski, T.; Jabłoński, M.; Cyranka, K.; Rymaszewska, J.; Piotrowski, P.; Kotowicz, K.; Frydecka, D. Adverse Childhood Experiences and Methylation of the FKBP5 Gene in Patients with Psychotic Disorders. J. Clin. Med. 2020, 9, 3792. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm9123792
Misiak B, Karpiński P, Szmida E, Grąźlewski T, Jabłoński M, Cyranka K, Rymaszewska J, Piotrowski P, Kotowicz K, Frydecka D. Adverse Childhood Experiences and Methylation of the FKBP5 Gene in Patients with Psychotic Disorders. Journal of Clinical Medicine. 2020; 9(12):3792. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm9123792
Chicago/Turabian StyleMisiak, Błażej, Paweł Karpiński, Elżbieta Szmida, Tomasz Grąźlewski, Marcin Jabłoński, Katarzyna Cyranka, Joanna Rymaszewska, Patryk Piotrowski, Kamila Kotowicz, and Dorota Frydecka. 2020. "Adverse Childhood Experiences and Methylation of the FKBP5 Gene in Patients with Psychotic Disorders" Journal of Clinical Medicine 9, no. 12: 3792. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm9123792