
Biological and Ecological Aspects of the Blackmouth Catshark (Galeus melastomus Rafinesque, 1810) in the Southern Tyrrhenian Sea

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
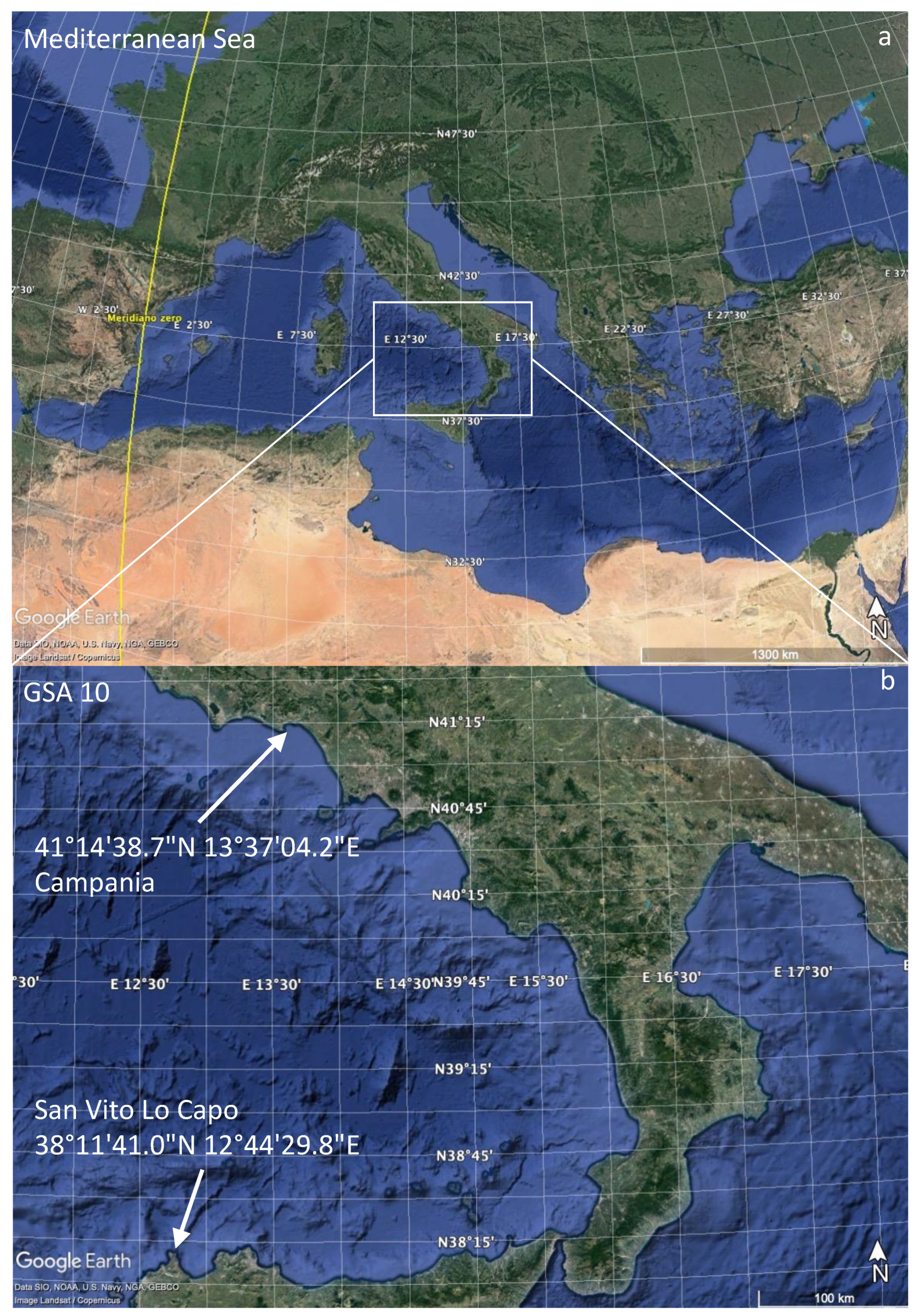
2. Materials and Methods
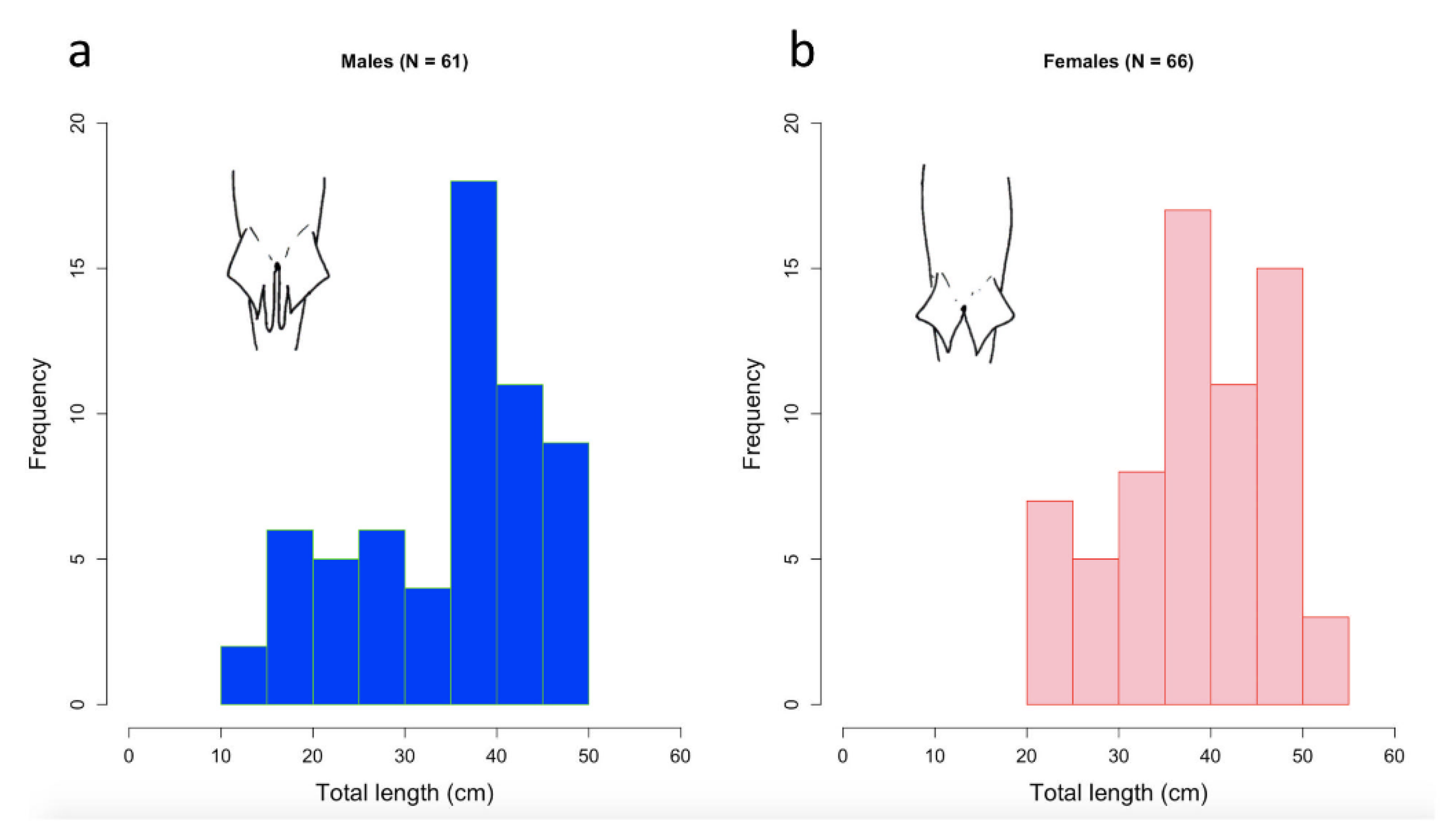
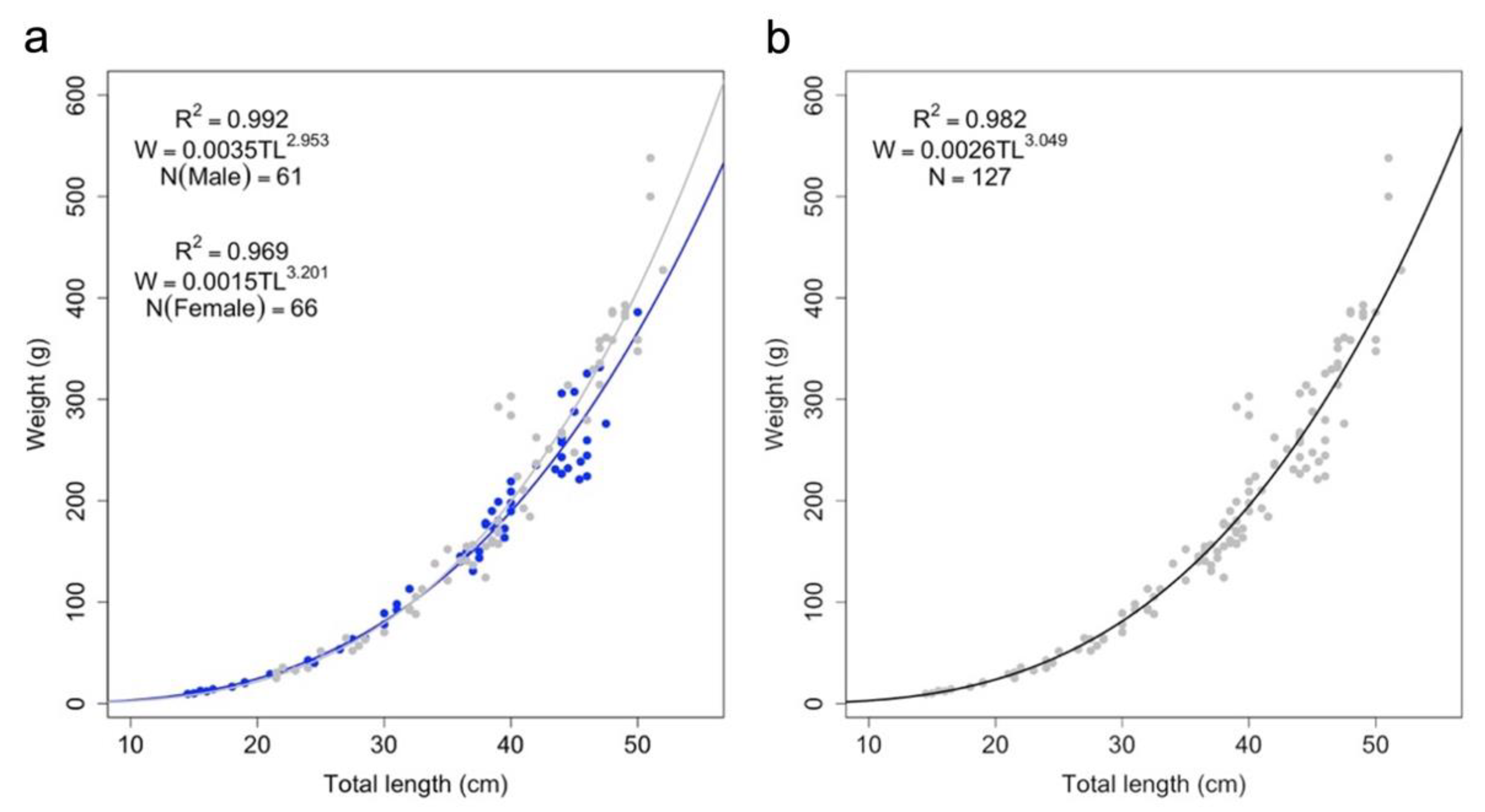
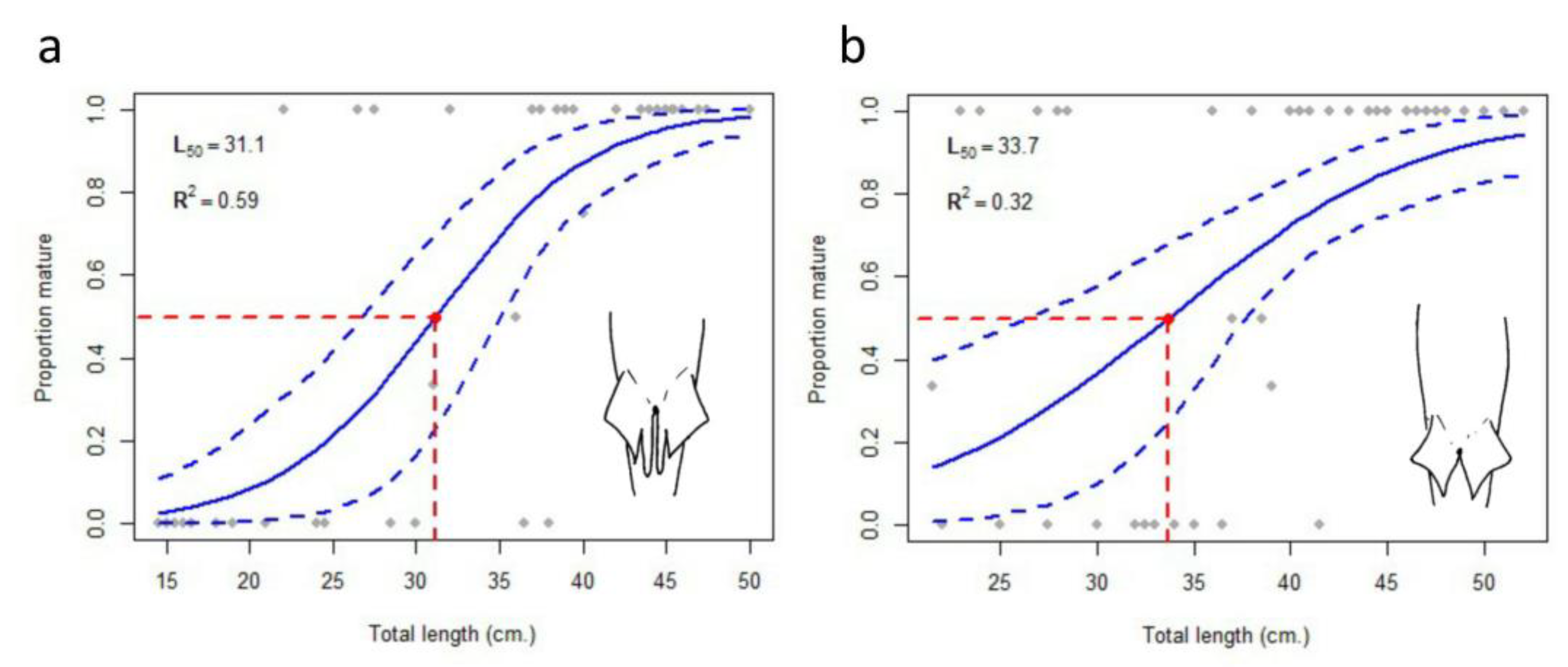
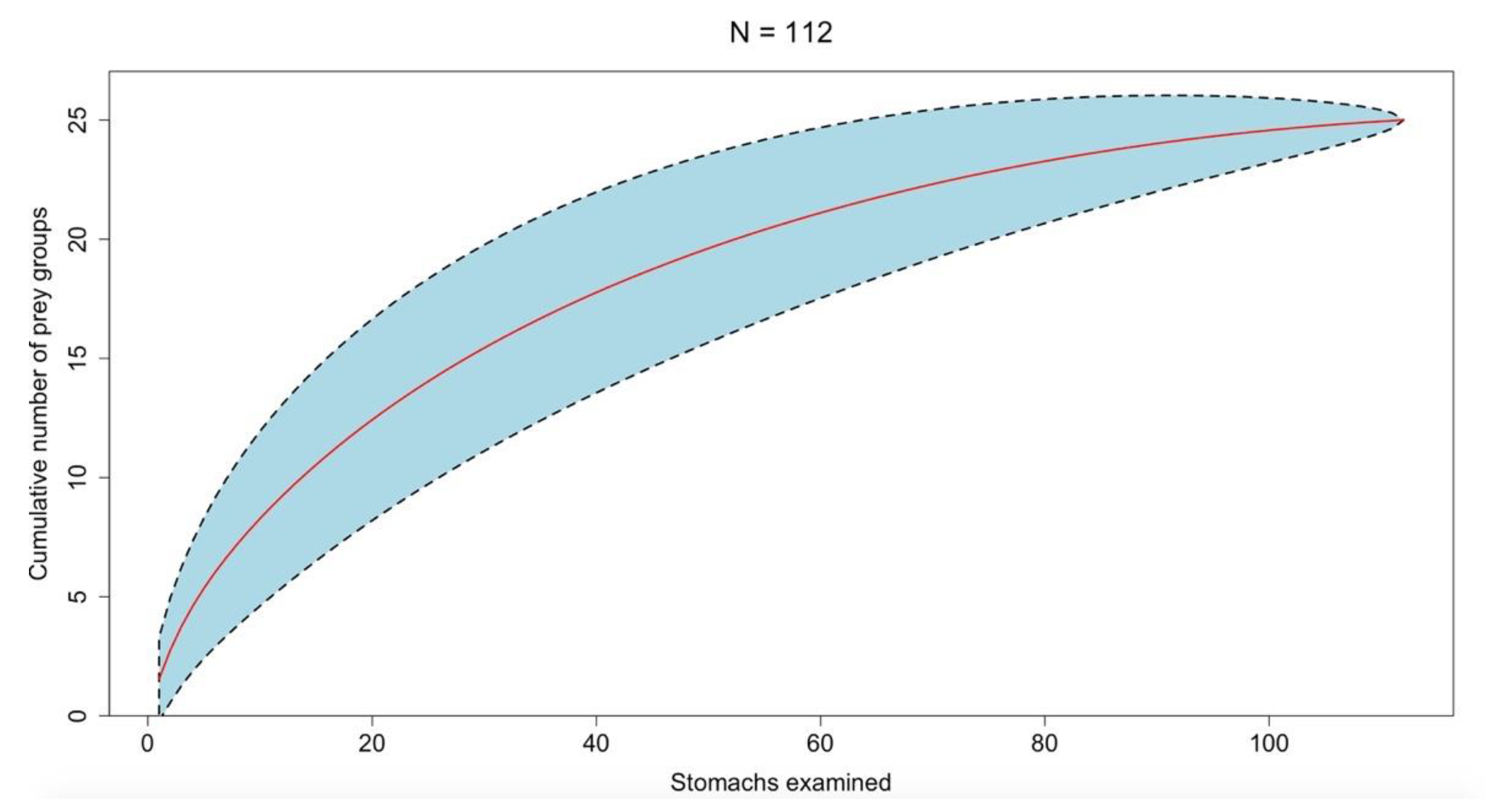
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nowicki, R.J.; Thomson, J.A.; Wirsing, A.J.; Heithaus, M.R.; Fourqurean, J.W. Loss of predation risk from apex predators can exacerbate marine tropicalization caused by extreme climatic events. J. Anim. Ecol. 2021, 1–12. [Google Scholar] [CrossRef]
- Tiralongo, F.; Messina, G.; Lombardo, B.M. Biological Aspects of Juveniles of the Common Stingray, Dasyatis pastinaca (Linnaeus, 1758) (Elasmobranchii, Dasyatidae), from the Central Mediterranean Sea. J. Mar. Sci. Eng. 2020, 8, 269. [Google Scholar] [CrossRef] [Green Version]
- Tiralongo, F.; Messina, G.; Gatti, R.C.; Tibullo, D.; Lombardo, B.M. Some biological aspects of juveniles of the rough ray, Raja radula Delaroche, 1809 in Eastern Sicily (central Mediterranean Sea). J. Sea Res. 2018, 142, 174–179. [Google Scholar] [CrossRef]
- Tiralongo, F.; Messina, G.; Brundo, M.V.; Lombardo, B.M. Biological aspects of the common torpedo, Torpedo torpedo (Linnaeus, 1758) (Elasmobranchii: Torpedinidae), in the central Mediterranean Sea (Sicily, Ionian Sea). Eur. Zool. J. 2019, 86, 488–496. [Google Scholar] [CrossRef]
- Pacoureau, N.; Rigby, C.L.; Kyne, P.M.; Sherley, R.B.; Winker, H.; Carlson, J.K.; Fordham, S.V.; Barreto, R.; Fernando, D.; Francis, M.P.; et al. Half a century of global decline in oceanic sharks and rays. Nature 2021, 589. [Google Scholar] [CrossRef]
- Tiralongo, F.; Messina, G.; Lombardo, B.M. Discards of elasmobranchs in a trammel net fishery targeting cuttlefish, Sepia officinalis Linnaeus, 1758, along the coast of Sicily (central Mediterranean Sea). Reg. Stud. Mar. Sci. 2018, 20, 60–63. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Allen, D.J.; Ralph, G.M.; Walls, R.H.L. The Conservation Status of Sharks, Rays and Chimaeras in the Mediterranean Sea; IUCN: Malaga, Spain, 2016. [Google Scholar]
- D’Iglio, C.; Savoca, S.; Rinelli, P.; Spanò, N. Diet of the Deep-Sea Shark Galeus melastomus Rafinesque, 1810, in the Mediterranean Sea: What We Know and What We Should Know. Sustainability 2021, 13, 3962. [Google Scholar] [CrossRef]
- Capapé, C.; Reynaud, C.; Vergne, Y.; Quignard, J.P. Biological observations on the smallspotted catshark scyliorhinus canicula (chondrichthyes: Scyliorhinidae) off the languedocian coast (Southern France, Northern mediterranean). Panam. J. Aquat. Sci. 2008, 3, 282–289. [Google Scholar]
- Tursi, A.; D’Onghia, G.; Matarrese, A.; Piscitelli, G. Observations on population biology of the blackmouth catshark Galeus melastomus (Chondrichthyes, Scyliorhinidae) in the Ionian Sea. Cybium 1993, 17, 187–196. [Google Scholar]
- Ramírez-Amaro, S.; Ordines, F.; Terrasa, B.; Esteban, A.; García, C.; Guijarro, B.; Massutí, E. Demersal chondrichthyans in the western Mediterranean: Assemblages and biological parameters of their main species. Mar. Freshw. Res. 2016, 67, 636–652. [Google Scholar] [CrossRef] [Green Version]
- Camhi, M.D.; Fowler, S.L.; Musick, J.A.; Bräutigam, A.; Fordham, S.V.; Brautigam, A. Sharks and Their Relatives: Ecology and Conservation; IUCN: Gland, Switzerland, 1998; Volume 3, ISBN 283170460X. [Google Scholar]
- Rey, J.; de Sola, L.G.; Massutí, E. Distribution and biology of the blackmouth catshark Galeus melastomus in the Alboran Sea (Southwestern Mediterranean). J. Northwest Atl. Fish. Sci. 2005, 35, 215–223. [Google Scholar] [CrossRef]
- Lauriano, E.R.; Pergolizzi, S.; Aragona, M.; Montalbano, G.; Guerrera, M.C.; Crupi, R.; Faggio, C.; Capillo, G. Fish and Shellfish Immunology Intestinal immunity of dogfish Scyliorhinus canicula spiral valve: A histochemical, immunohistochemical and confocal study. Fish Shellfish Immunol. 2019, 87, 490–498. [Google Scholar] [CrossRef]
- Pergolizzi, E.R.L.S.; Kuciel, J.G.M.; Faggio, M.A.C. Immunohistochemical colocalization of G protein alpha subunits and 5-HT in the rectal gland of the cartilaginous fish Scyliorhinus canicula. Microsc. Res. Tech. 2017. [Google Scholar] [CrossRef]
- Ordines, F.; Massutí, E.; Moranta, J.; Quetglas, A.; Guijarro, B.; Fliti, K. Islas baleares vs argelia: Dos poblaciones cercanas de elasmobranquios en el mediterráneo occidental con distintas condiciones oceanográficas e historias pesqueras. Sci. Mar. 2011, 75, 707–717. [Google Scholar] [CrossRef]
- Gouraguine, A.; Hidalgo, M.; Moranta, J.; Bailey, D.M.; Ordines, F.; Guijarro, B.; Valls, M.; Barberá, C.; De Mesa, A. Elasmobranch spatial segregation in the western Mediterranean. Sci. Mar. 2011. [Google Scholar] [CrossRef]
- Rodríguez-Cabello, C.; Sánchez, F.; Serrano, A.; Olaso, I. Effects of closed trawl fishery areas on some elasmobranch species in the Cantabrian Sea. J. Mar. Syst. 2008. [Google Scholar] [CrossRef]
- Ligas, A.; Osio, G.C.; Sartor, P.; Sbrana, M.; de Ranieri, S. Trayectoria a largo plazo de algunas especies de elasmobranquios en los mares de Toscana (Mediterráneo noroccidental) a través de 50 años de datos de captura. Sci. Mar. 2013, 77, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Heithaus, M.R.; Frid, A.; Wirsing, A.J.; Worm, B. Predicting ecological consequences of marine top predator declines. Trends Ecol. Evol. 2008, 23, 202–210. [Google Scholar] [CrossRef]
- Bradai, M.N.; Saidi, B.; Enajjar, S. Elasmobranchs of the Mediterranean and Black Sea: Status, Ecology and Biology. Bibliographic Analysis; FAO: Rome, Italy, 2012; ISBN 9789251073865. [Google Scholar]
- Psomadakis, P.N.; Giustino, S.; Vacchi, M. Mediterranean fish biodiversity: An updated inventory with focus on the Ligurian and Tyrrhenian seas. Zootaxa 2012, 3263, 1–46. [Google Scholar] [CrossRef]
- Relini, G. Checklist Della Flora e Della Fauna dei Mari Italiani (Parte II); Ministero dell’ambiente e della tutela del territorio e del mare: Rome, Italy, 2010; Volume 17. [Google Scholar]
- García-de-Vinuesa, A.; Demestre, M.; Carreño, A.; Lloret, J. The Bioactive Potential of Trawl Discard: Case Study from a Crinoid Bed Off Blanes (North-Western Mediterranean). Mar. Drugs 2021, 19, 83. [Google Scholar] [CrossRef] [PubMed]
- Rinelli, P.; Bottari, T.; Florio, G.; Romeo, T.; Giordano, D.; Greco, S. Observations on distribution and biology of Galeus melastomus (Chondrichthyes, Scyliorhinidae) in the southern Tyrrhenian Sea (central Mediterranean). Cybium 2005, 29, 41–46. [Google Scholar]
- Ragonese, S.; Di Stefano, L.; Bianchini, M.L. Catture e selettività di pesci cartilaginei nella pesca dei gamberi rossi nello Stretto di Sicilia. Biol. Mar. Medit. 2000, 7, 400–401. [Google Scholar]
- Dimech, M.; Kaiser, M.J.; Ragonese, S.; Schembri, P.J. Ecosystem effects of fishing on the continental slope in the Central Mediterranean Sea. Mar. Ecol. Prog. Ser. 2012, 449, 41–54. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.G.; Tselepides, A.; Bagley, P.M.; Collins, M.A.; Priede, I.G. Bathymetric distribution of some benthic and benthopelagic species attracted to baited cameras and traps in the deep eastern Mediterranean. Mar. Ecol. Prog. Ser. 2003, 251, 75–86. [Google Scholar] [CrossRef]
- Ungaro, N.; Marano, G. Galeus melastomus Rafinesque, 1810 (Selachii, Scyliorhinidae). Distribuzione e biologia sui fondi batiali del basso Adriatico. Atti. Rel. Acc. Pugl. Sci. 1994, 49, 195–207. [Google Scholar]
- Group, M.W. International Bottom Trawl Survey in the Mediterranean Instruction Manual Version 6. 2012. Available online: https://www.sibm.it/MEDITS%202011/docs/Medits-Handbook-2012%20versione%20definitiva.pdf (accessed on 28 August 2021).
- Ragonese, S.; Norrito, G. Le principali criticità del Piano Italiano di Raccolta Dati Alieutici dal punto di vista di un collega esperto in pesca e proveniente da un esopianeta (in vista della nuova programmazione 2021–2024). NTR-ITPP 2021, sr102, 1–49. [Google Scholar]
- Freedman, D.A.; Purves, R.A. Bayes’ Method for Bookies. Ann. Math. Stat. 1969, 40, 1177–1186. [Google Scholar] [CrossRef]
- Šmídl, V.; Quinn, A. The Variational Bayes Method in Signal Processing; Springer Science & Business Media: Berlin, Germany, 2006; ISBN 3540288201. [Google Scholar]
- Froese, R.; Thorson, J.T.; Reyes, R.B. A Bayesian approach for estimating length-weight relationships in fishes. J. Appl. Ichthyol. 2014, 30, 78–85. [Google Scholar] [CrossRef] [Green Version]
- Jisr, N.; Younes, G.; Sukhn, C.; El-Dakdouki, M.H. Length-weight relationships and relative condition factor of fish inhabiting the marine area of the Eastern Mediterranean city, Tripoli-Lebanon. Egypt. J. Aquat. Res. 2018, 44, 299–305. [Google Scholar] [CrossRef]
- Pandis, N. The chi-square test. Am. J. Orthod. Dentofac. Orthop. 2016, 150, 898–899. [Google Scholar] [CrossRef]
- Torrejon-Magallanes, J. Package sizeMat. Available online: https://cran.r-project.org/package=sizeMat (accessed on 30 August 2021).
- Follesa, M.C.; Carbonara, P. Atlas of the Maturity Stages of Mediterranean Fishery Resources. Studies and Reviews n. 99; FAO: Rome, Italy, 2019; ISBN 9789251319758. [Google Scholar]
- Euzen, O. Food habits and diet composition of some fish of Kuwait. Kuwait Bull. Mar. Sci. 1987, 9, 65–85. [Google Scholar]
- Hyslop, E.J. Stomach contents analysis—a review of methods and their application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef] [Green Version]
- Cortés, E. A critical review of methods of studying fish feeding based on analysis of stomach contents: Application to elasmobranch fishes. Can. J. Fish. Aquat. Sci. 1997, 54, 726–738. [Google Scholar] [CrossRef]
- Carrassón, M.; Matallanas, J.; Casadevall, M. Feeding strategies of deep-water morids on the western Mediterranean slope. Deep Sea Res. Part I Oceanogr. Res. Pap. 1997, 44, 1685–1699. [Google Scholar] [CrossRef]
- N’Da, K. Regime alimentaire du rouget de roche Mullus surmuletus (Mullidae) dans le Nord du Golfe de Gascogne. Cybium 1992, 16, 159–167. [Google Scholar]
- Krebs, J.C. Ecological Methodology; Harper Row: New York, NY, USA, 1989; Volume 620. [Google Scholar]
- Novakowski, G.C.; Hahn, N.S.; Fugi, R. Diet seasonality and food overlap of the fish assemblage in a pantanal pond. Neotrop. Ichthyol. 2008, 6, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Ragonese, S.; Nardone, G.; Ottonello, D.; Gancitano, S.; Giusto, G.B.; Sinacori, G. Distribution and biology of the blackmouth catshark galeus melastomus in the strait of sicily (central mediterranean sea). Mediterr. Mar. Sci. 2009, 10, 55–72. [Google Scholar] [CrossRef]
- Tserpes, G.; Maravelias, C.D.; Pantazi, M.; Peristeraki, P. Distribution of relatively rare demersal elasmobranchs in the eastern Mediterranean. Estuar. Coast. Shelf Sci. 2013. [Google Scholar] [CrossRef]
- Capapé, C.; Guélorget, O.; Vergne, Y.; Reynaud, C. Reproductive biology of the blackmouth catshark, Galeus melastomus (Chondrichthyes: Scyliorhinidae) off the Languedocian coast (southern France, northern Mediterranean). J. Mar. Biol. Assoc. UK 2008, 88, 415–421. [Google Scholar] [CrossRef]
- Metochis, C.P.; Carmona-Antoñanzas, G.; Kousteni, V.; Damalas, D.; Megalofonou, P. Population structure and aspects of the reproductive biology of the blackmouth catshark, Galeus melastomus Rafinesque, 1810 (Chondrichthyes: Scyliorhinidae) caught accidentally off the Greek coasts. J. Mar. Biol. Assoc. UK 2018, 98, 909–925. [Google Scholar] [CrossRef]
- Capapé, C.; Zaouali, J. Contribution a la biologie des Scyliorhinidae des cotes tunisiennes. VI. Galeus melastomus Rafinesque, 1810. Repartition geographique et bathymetrique, sexualite, reproduction, fecondite. Cah. Biol. Mar. 1977, 18, 449–463. [Google Scholar]
- Capape, C.; Quignard, J.P. Contribution a La Biologie Des Rajidae Des Cotes Tunisiennes. I. Raja Miraletus, Linne, 1758: Repartition Geographique Et Bathymetrique, Sexualite, Reproduction, Fecondite. Arch. Inst. Pasteur Tunis 1974, 51, 39–60. [Google Scholar]
- D’Onghia, G.; Matarrese, A.; Tursi, A.; Sion, L. Observations on the depth distribution pattern of the small-spotted catshark in the North Aegean Sea. J. Fish Biol. 1995, 47, 421–426. [Google Scholar] [CrossRef]
- Carbonara, P.; Casciaro, L.; Bitetto, I.; Zupa, W.; Gaudio, P.; Facchini, M.T.; Spedicato, M. Reproductive cycle and length at first maturity of Galeus melastomus in the central Mediterranean Sea. In Proceedings of the 16th Annual Scientific Conference of the European Elasmobranch Association (EEA), Italy, Milan, 22–25 November 2012; p. 58. [Google Scholar]
- Ramírez-Amaro, S.; Picornell, A.; Arenas, M.; Castro, J.A.; Massutí, E.; Ramon, M.M.; Terrasa, B. Contrasting evolutionary patterns in populations of demersal sharks throughout the western Mediterranean. Mar. Biol. 2018, 165, 1–16. [Google Scholar] [CrossRef]
- Roughgarden, J. A simple model for population dynamics in stochastic environments. Am. Nat. 1975, 109, 713–736. [Google Scholar] [CrossRef]
- Lindström, J.; Kokko, H. Cohort effects and population dynamics. Ecol. Lett. 2002, 5, 338–344. [Google Scholar] [CrossRef]
- Wheeler, C.R.; Gervais, C.R.; Johnson, M.S.; Vance, S.; Rosa, R.; Mandelman, J.W.; Rummer, J.L. Anthropogenic stressors influence reproduction and development in elasmobranch fishes. Rev. Fish Biol. Fish. 2020. [Google Scholar] [CrossRef]
- Savoca, S.; Bottari, T.; Fazio, E.; Bonsignore, M.; Mancuso, M.; Luna, G.M.; Romeo, T.; D’Urso, L.; Capillo, G.; Panarello, G.; et al. Plastics occurrence in juveniles of Engraulis encrasicolus and Sardina pilchardus in the Southern Tyrrhenian Sea. Sci. Total Environ. 2020, 718. [Google Scholar] [CrossRef] [PubMed]
- Savoca, S.; Capillo, G.; Mancuso, M.; Bottari, T.; Crupi, R.; Branca, C.; Romano, V.; Faggio, C.; D’Angelo, G.; Spanò, N. Microplastics occurrence in the Tyrrhenian waters and in the gastrointestinal tract of two congener species of seabreams. Environ. Toxicol. Pharmacol. 2019, 67, 35–41. [Google Scholar] [CrossRef]
- Capillo, G.; Savoca, S.; Panarello, G.; Mancuso, M.; Branca, C.; Romano, V.; D’Angelo, G.; Bottari, T.; Spanò, N. Quali-quantitative analysis of plastics and synthetic microfibers found in demersal species from Southern Tyrrhenian Sea (Central Mediterranean). Mar. Pollut. Bull. 2020, 150, 110596. [Google Scholar] [CrossRef]
- Albano, M.; Panarello, G.; Di Paola, D.; D’Angelo, G.; Granata, A.; Savoca, S.; Capillo, G. The mauve stinger Pelagia noctiluca (Cnidaria, Scyphozoa) plastics contamination, the Strait of Messina case. Int. J. Environ. Stud. 2021, 0, 1–6. [Google Scholar] [CrossRef]
- Albano, M.; Panarello, G.; Di Paola, D.; Capparucci, F.; Crupi, R.; Gugliandolo, E.; Spanò, N.; Capillo, G.; Savoca, S. The Influence of Polystyrene Microspheres Abundance on Development and Feeding Behavior of Artemia salina (Linnaeus, 1758). Appl. Sci. 2021, 11, 3352. [Google Scholar] [CrossRef]
- Fazio, F.; D’Iglio, C.; Capillo, G.; Saoca, C.; Peycheva, K.; Piccione, G.; Makedonski, L. Environmental Investigations and Tissue Bioaccumulation of Heavy Metals in Grey Mullet from the Black Sea (Bulgaria) and the Ionian Sea (Italy). Animals 2020, 10, 1739. [Google Scholar] [CrossRef]
- Fazio, F.; Saoca, C.; Sanfilippo, M.; Capillo, G.; Spanò, N.; Piccione, G. Response of vanadium bioaccumulation in tissues of Mugil cephalus (Linnaeus 1758). Sci. Total Environ. 2019, 689, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Schilling, H.T.; Smith, J.A.; Stewart, J.; Everett, J.D.; Hughes, J.M.; Suthers, I.M. Reduced exploitation is associated with an altered sex ratio and larger length at maturity in southwest Pacific (east Australian) Pomatomus saltatrix. Mar. Environ. Res. 2019, 147, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Yanti, A.; Tresnati, J.; Yasir, I.; Syafiuddin; Rahmani, P.Y.; Aprianto, R.; Tuwo, A. Size at the maturity of sea cucumber Holothuria scabra. Is it an overfishing sign in Wallacea Region? IOP Conf. Ser. Earth Environ. Sci. 2020, 473, 12056. [Google Scholar] [CrossRef]
- Froese, R. Keep it simple: Three indicators to deal with overfishing. Fish Fish. 2004, 5, 86–91. [Google Scholar] [CrossRef] [Green Version]
- Trippel, E.A. Age at maturity as a stress indicator in fisheries. Bioscience 1995, 45, 759–771. [Google Scholar] [CrossRef]
- Lister, A.L.; Van Der Kraak, G.J.; Rutherford, R.; MacLatchy, D. Fundulus heteroclitus: Ovarian reproductive physiology and the impact of environmental contaminants. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 154, 278–287. [Google Scholar] [CrossRef]
- Parisi, C.; Guerriero, G. Antioxidative defense and fertility rate in the assessment of reprotoxicity risk posed by global warming. Antioxidants 2019, 8, 622. [Google Scholar] [CrossRef] [Green Version]
- Kime, D.E. The effects of pollution on reproduction in fish. Rev. Fish Biol. Fish. 1995, 5, 52–95. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Forrest, R.E. Life histories, population dynamics, and extinction risks in chondrichthyans. Sharks and Their Relatives II: Biodiversity, Adaptive Physiology, and Conservation. 2010, pp. 639–679. Available online: https://www.researchgate.net/publication/237044390_Life_Histories_Population_Dynamics_and_Extinction_Risks_in_Chondrichthyans (accessed on 28 August 2021).
- Spanò, N.; Di Paola, D.; Albano, M.; Manganaro, A.; Sanfilippo, M.; D’Iglio, C.; Capillo, G.; Savoca, S. Growth performance and bioremediation potential of Gracilaria gracilis (Steentoft, L.M. Irvine & Farnham, 1995). Int. J. Environ. Stud. 2021, 1–13. [Google Scholar] [CrossRef]
- Savoca, S.; Lo Giudice, A.; Papale, M.; Mangano, S.; Caruso, C.; Spanò, N.; Michaud, L.; Rizzo, C. Antarctic sponges from the Terra Nova Bay (Ross Sea) host a diversified bacterial community. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Impacts, Adaptation, and Vulnerability; Field, L., Barros, C., Dokken, V., Mach, D., Mastrandrea, K., Bilir, M., Chatterjee, T., Ebi, M., Estrada, K., Genova, Y., et al., Eds.; IPCC: England, UK; New York, NY, USA, 2014. [Google Scholar]
- Di Santo, V. Intraspecific variation in physiological performance of a benthic elasmobranch challenged by ocean acidification and warming. J. Exp. Biol. 2016, 219, 1725–1733. [Google Scholar] [CrossRef] [Green Version]
- Ripley, D.M.; De Giorgio, S.; Gaffney, K.; Thomas, L.; Shiels, H.A. Ocean warming impairs the predator avoidance behaviour of elasmobranch embryos. Conserv. Physiol. 2021, 9, coab045. [Google Scholar] [CrossRef]
- Musa, S.M.; Ripley, D.M.; Moritz, T.; Shiels, H.A. Ocean warming and hypoxia affect embryonic growth, fitness and survival of small-spotted catsharks, Scyliorhinus canicula. J. Fish Biol. 2020, 97, 257–264. [Google Scholar] [CrossRef]
- Pistevos, J.C.A.; Nagelkerken, I.; Rossi, T.; Olmos, M.; Connell, S.D. Ocean acidification and global warming impair shark hunting behaviour and growth. Nat. Publ. Gr. Sci.Rep. 2015, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rosa, R.; Baptista, M.; Lopes, V.M.; Pegado, M.R.; Paula, R.; Tru, K.; Repolho, T.; Rosa, R. Early-life exposure to climate change impairs tropical shark survival Author for correspondence. Proc. R. Soc. B 2014, 281. [Google Scholar] [CrossRef] [Green Version]
- Rosa, R.; Rummer, J.L.; Munday, P.L.; Rosa, R. Biological responses of sharks to ocean acidification. Biol. Lett. 2017, 13, 201607. [Google Scholar] [CrossRef] [Green Version]
- Barragán-Méndez, C.; Sánchez-García, F.; Sobrino, I.; Mancera, J.M.; Ruiz-Jarabo, I. Air Exposure in Catshark (Scyliorhinus canicula) Modify Muscle Texture Properties: A Pilot Study. Fishes 2018, 3, 34. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.S.; Kraver, D.W.; Renshaw, G.M.C.; Rummer, J.L. Will ocean acidification affect the early ontogeny of a tropical oviparous elasmobranch (Hemiscyllium ocellatum)? Conserv. Physiol. 2016, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Schlaff, A.M.; Heupel, M.R.; Simpfendorfer, C.A. Influence of environmental factors on shark and ray movement, behaviour and habitat use: A review. Rev. Fish Biol. Fish. 2014, 24, 1089–1103. [Google Scholar] [CrossRef]
- Barnes, C.A.B.; Axwell, D.A.M.; Euman, D.A.C.R. Global patterns in predator – prey size relationships reveal size dependency of trophic transfer efficiency. Ecology 2010, 91, 222–232. [Google Scholar] [CrossRef] [Green Version]
- Perry, S.F.; Gilmour, K.M. Acid–base balance and CO 2 excretion in fish: Unanswered questions and emerging models. Respir. Physiol. Neurobiol. 2006, 154, 199–215. [Google Scholar] [CrossRef]
- Vucic-Pestic, O.; Ehnes, B.R.; Rall, B.C.; Brose, U. Warming up the system: Higher predator feeding rates but lower energetic efficiencies. Glob. Chang. Biol. 2011, 1301–1310. [Google Scholar] [CrossRef]
- Rall, B.C.; Vucic-Pestic, O.; Ehnes, B.R.; Emmerson, M.; Brose, U. Temperature, predator – prey interaction strength and population stability. Glob. Chang. Biol. 2010, 16, 2145–2157. [Google Scholar] [CrossRef]
- Danovaro, R. Climate change impacts on the biota and on vulnerable habitats of the deep Mediterranean Sea. Rend. Lincei. Sci. Fis. e Nat. 2018, 29, 525–541. [Google Scholar] [CrossRef]
- Yasuhara, M.; Danovaro, R. Temperature impacts on deep-sea biodiversity. Biol. Rev. 2016, 91, 275–287. [Google Scholar] [CrossRef]
- Sweetman, A.K.; Thurber, A.R.; Smith, C.R.; Levin, L.A.; Mora, C.; Wei, C.L.; Gooday, A.J.; Jones, D.O.B.; Rex, M.; Yasuhara, M.; et al. Major impacts of climate change on deep-sea benthic ecosystems. Elementa 2017, 5, 1–23. [Google Scholar] [CrossRef]
- Danovaro, R.; Snelgrove, P.V.R.; Tyler, P. Challenging the paradigms of deep-sea ecology. Trends Ecol. Evol. 2014, 29, 465–475. [Google Scholar] [CrossRef]
- Catarino, D.; Knutsen, H.; Verissimo, A.; Olsen, E.M.; Jorde, P.E.; Menezes, G.; SannÆs, H.; Stankovic, D.; Company, J.B.; Neat, F.; et al. The Pillars of Hercules as a bathymetric barrier to gene flow promoting isolation in a global deep-sea shark (Centroscymnus coelolepis). Mol. Ecol. 2015, 24, 6061–6079. [Google Scholar] [CrossRef]
- Henry, L.; Stehmann, M.F.W.; Clippele, L. De Seamount egg-laying grounds of the deep-water skate Bathyraja richardsoni. J. Fish Biol. 2016, 89, 1473–1481. [Google Scholar] [CrossRef] [PubMed]
- Olaso, I.; Velasco, F.; Sánchez, F.; Serrano, A.; Rodríguez-Cabello, C.; Cendrero, O. Trophic relations of lesser-spotted catshark (Scyliorhinus canicula) and blackmouth catshark (Galeus melastomus) in the Cantabrian Sea. J. Northwest Atl. Fish. Sci. 2005, 35, 481–494. [Google Scholar] [CrossRef]
- Bendiab, A.A.T.; Mouffok, S.; Boutiba, Z. Feeding Habits of Two Sharks Species Scyliorhinus canicula (Linnaeus, 1758) and Galeus melastomus (Rafinesque, 1810) in the Western Algerian coasts. J. Appl. Environ. Biol. Sci. 2016, 6, 108–116. [Google Scholar]
- Neves, A.; Figueiredo, I.; Moura, T.; Assis, C.; Gordo, L.S. Diet and feeding strategy of Galeus melastomus in the continental slope off southern Portugal. Vie Milieu 2007, 57, 165–169. [Google Scholar]
- Carrassón, M.; Cartes, J.E. Trophic relationships in a Mediterranean deep-sea fish community: Partition of food resources, dietary overlap and connections within the benthic boundary layer. Mar. Ecol. Prog. Ser. 2002, 241, 41–55. [Google Scholar] [CrossRef] [Green Version]
- Macpherson, E. Régime alimentaire de Galeus melastomus (Rafinesque, 1810), Etmopterus 8 spinax (L., 1758) et Scymnorhinus licha (Bonaterre, 1788), en Méditerranée Occidentale. 1980. Available online: https://digital.csic.es/handle/10261/161666?locale=en (accessed on 28 August 2021).
- Macpherson, E. Resource Partitioning in a Mediterranean Demersal Fish Community. Mar. Ecol. Prog. Ser. 1981, 4, 183–193. [Google Scholar] [CrossRef]
- Tuset, V.M.; Lombarte, A.; Assis, C.A. Otolith atlas for the western Mediterranean, north and central eastern Atlantic. Sci. Mar. 2008, 72, 7–198. [Google Scholar] [CrossRef]
- D’Iglio, C.; Albano, M.; Famulari, S.; Savoca, S.; Panarello, G.; Di Paola, D.; Perdichizzi, A.; Rinelli, P.; Lanteri, G.; Spanò, N.; et al. Intra- and interspecific variability among congeneric Pagellus otoliths. Sci. Rep. 2021, 11, 16315. [Google Scholar] [CrossRef]
- Cartes, J.E.; Soler-Membrives, A.; Stefanescu, C.; Lombarte, A.; Carrassón, M. Contributions of allochthonous inputs of food to the diets of benthopelagic fish over the northwest Mediterranean slope (to 2300 m). Deep. Res. Part I Oceanogr. Res. Pap. 2016, 109, 123–136. [Google Scholar] [CrossRef]
- Anastasopoulou, A.; Mytilineou, C.; Lefkaditou, E.; Dokos, J.; Smith, C.J.; Siapatis, A.; Bekas, P.; Papadopoulou, K.N. Diet and feeding strategy of blackmouth catshark Galeus melastomus. J. Fish Biol. 2013, 83, 1637–1655. [Google Scholar] [CrossRef] [PubMed]
- Ait Darna, S.; Taleb Bendiab, A.A.; Mouffok, S.; Belmahi, A.-E.; Bouderbala, M. Observation on distribution, biology, growth, diet and feeding strategy of blackmouth catshark Galeus melastomus (Rafinesque, 1810) (Chondrichthyes: Scyliorhinidae) in western Algerian coasts. Biodivers. J. 2018, 9, 357–368. [Google Scholar] [CrossRef]
- Fanelli, E.; Rey, J.; Torres, P.; Gil De Sola, L. Feeding habits of blackmouth catshark Galeus melastomus Rafinesque, 1810 and velvet belly lantern shark Etmopterus spinax (Linnaeus, 1758) in the western Mediterranean. J. Appl. Ichthyol. 2009, 25, 83–93. [Google Scholar] [CrossRef]
- Carrassón, M.; Stefanescu, C.; Cartes, J.E. Diets and bathymetric distributions of two bathyal sharks of the Catalan deep sea (western Mediterranean). Mar. Ecol. Prog. Ser. 1992, 82, 21–30. [Google Scholar] [CrossRef]
- Bozzano, A.; Murgia, R.; Vallerga, S.; Hirano, J.; Archer, S. The photoreceptor system in the retinae of two dogfishes, Scyliorhinus canicula and Galeus melastomus: Possible relationship with depth distribution and predatory lifestyle. J. Fish Biol. 2001, 59, 1258–1278. [Google Scholar] [CrossRef]
- Bello, G. Heteroteuthis dispar (Cephalopoda: Sepiolidae) nella dieta di selaci demersali. Biol. Mar. Medit. 1995, 2, 211–215. [Google Scholar]
- Valls, M.; Quetglas, A.; Ordines, F.; Moranta, J. Feeding ecology of demersal elasmobranchs from the shelf and slope off the Balearic Sea (western Mediterranean). Sci. Mar. 2011. [Google Scholar] [CrossRef] [Green Version]
- Rodhouse, P.G.K.; Lu, C.C.; Roper, C.F.E. Malcolm Roy Clarke, FRS. J. Mar. Biol. Assoc. UK 2015, 95, 865–870. [Google Scholar] [CrossRef]
- Pierce, G.J.; Boyle, P.R. A review of methods for diet analysis in piscivorous marine mammals. Oceanogr. Mar. Biol. Annu. Rev. 1991, 29. [Google Scholar]
- Bello, G. Cephalopods from the stomach contents of demersal chondrichthyans caught in the Adriatic Sea. Vie Milieu 1997, 47, 221–227. [Google Scholar]
- Dallarés, S.; Padrós, F.; Cartes, J.E.; Solé, M.; Carrassón, M. The parasite community of the sharks Galeus melastomus, Etmopterus spinax and Centroscymnus coelolepis from the NW Mediterranean deep-sea in relation to feeding ecology and health condition of the host and environmental gradients and variables. Deep. Res. Part I Oceanogr. Res. Pap. 2017, 129, 41–58. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Length–Weight Relationship Parameters | ||||||
|---|---|---|---|---|---|---|
| a | C.I. a | b | C.I. b | R2 | p-Value | |
| Females | 0.001 | 0.001–0.002 | 3.201 | 3.058–3.343 | 0.969 | 0.027 |
| Males | 0.003 | 0.002–0.004 | 2.953 | 2.883–3.023 | 0.992 | 0.047 |
| Combined | 0.002 | 0.001–0.003 | 3.049 | 2.976–3.121 | 0.982 | 0.079 |
| %N | %W | %F | %IRI | |
|---|---|---|---|---|
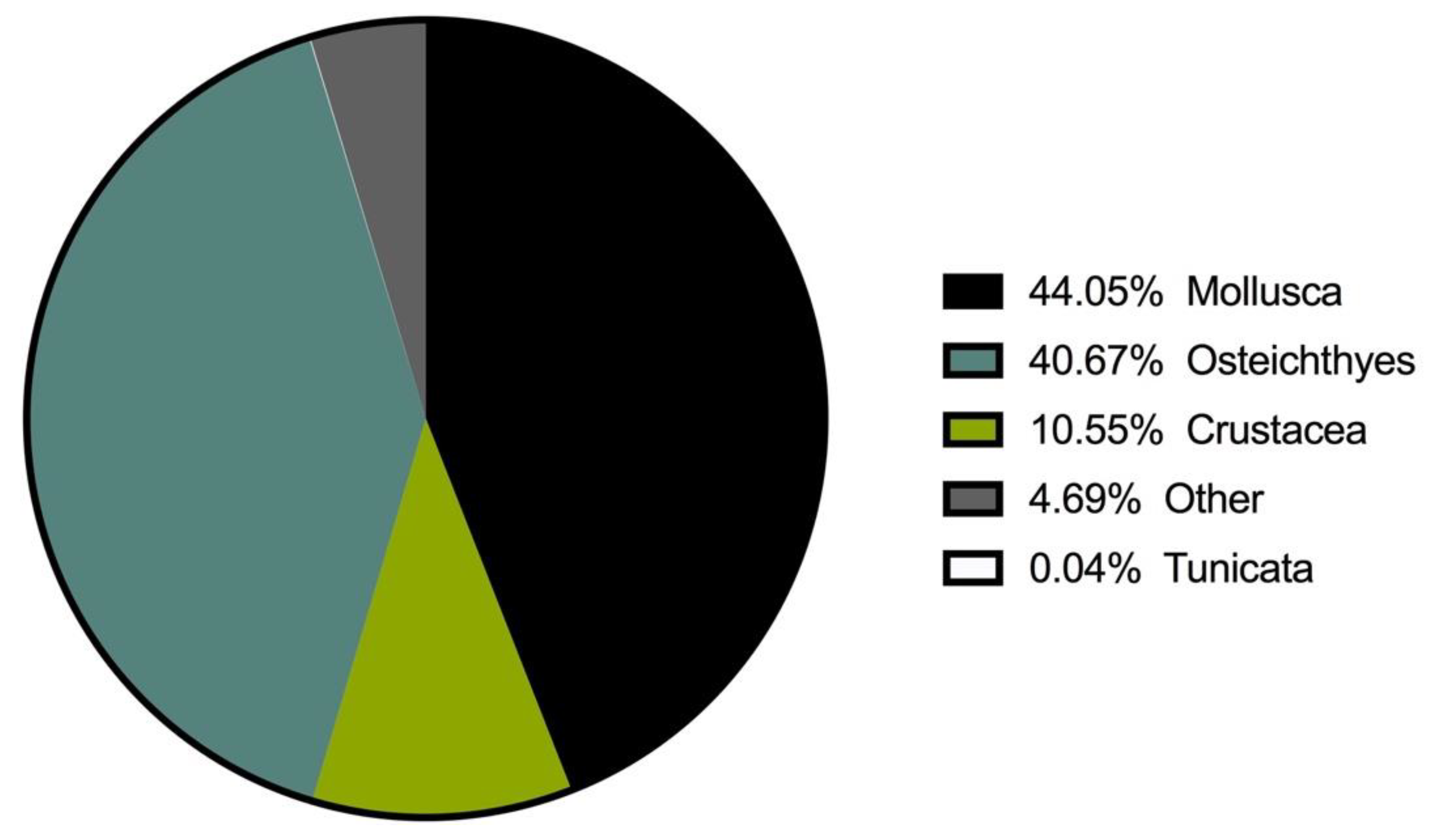
| MOLLUSCA | 29.00 | 44.05 | 56.94 | 35.57 |
| Gasteropoda | 0.28 | 0.18 | 0.76 | 0.01 |
| Gasteropoda n.i. | 0.28 | 0.18 | 0.76 | 0.01 |
| Cephalopoda | 28.72 | 43.87 | 56.18 | 35.56 |
| Abraliopsis morisii | 0.28 | 0.50 | 0.76 | 0.02 |
| Alloteuthis media | 0.56 | 0.54 | 1.52 | 0.06 |
| Eledone cirrhosa | 0.28 | 0.85 | 0.76 | 0.03 |
| Eledone sp. | 0.56 | 11.22 | 1.52 | 0.68 |
| Heteroteuthis dispar | 1.41 | 0.61 | 2.27 | 0.18 |
| Histioteuthis reversa | 0.85 | 0.86 | 2.27 | 0.15 |
| Histioteuthis sp. | 0.28 | 6.96 | 0.76 | 0.21 |
| Loligo sp. | 0.56 | 0.35 | 1.52 | 0.05 |
| Scaeurgus unicirrhus | 0.28 | 0.85 | 0.86 | 0.03 |
| Sepia orbignyana | 5.07 | 1.00 | 5.30 | 1.23 |
| Sepietta oweniana | 0.28 | 0.13 | 0.76 | 0.01 |
| Sepiola intermedia | 0.56 | 0.09 | 0.76 | 0.02 |
| Todarodes sagittatus | 3.10 | 9.66 | 5.30 | 2.59 |
| Cephalopoda n.i. | 14.65 | 10.25 | 31.82 | 30.30 |
| CRUSTACEA | 38.86 | 10.55 | 39.44 | 11.63 |
| Amphipoda | 0.28 | 0.04 | 0.76 | 0.01 |
| Amphipoda n.i. | 0.28 | 0.04 | 0.76 | 0.01 |
| Decapoda | 10.97 | 9.13 | 25.79 | 5.01 |
| Eusergestes arcticus | 0.28 | 0.09 | 0.76 | 0.01 |
| Parapenaeus longirostris | 0.28 | 0.26 | 0.76 | 0.02 |
| Pasiphaea multidentata | 0.56 | 2.00 | 0.76 | 0.07 |
| Pasiphea sivado | 1.41 | 0.78 | 3.79 | 0.32 |
| Pasiphea sp. | 0.56 | 0.46 | 1.52 | 0.06 |
| Plesionika giglioli | 0.28 | 1.07 | 0.76 | 0.04 |
| Robustosergia robusta | 0.28 | 0.45 | 0.76 | 0.02 |
| Solenocera membranacea | 0.28 | 0.56 | 0.76 | 0.02 |
| Brachyura n.i. | 0.28 | 0.13 | 0.76 | 0.01 |
| Decapoda n.i. | 0.56 | 0.26 | 1.52 | 0.05 |
| Dendrobranchiata n.i. | 5.92 | 2.98 | 12.88 | 4.38 |
| Sergestidae n.i. | 0.28 | 0.09 | 0.76 | 0.01 |
| Euphausiacea | 27.05 | 1.27 | 11.37 | 6.57 |
| Meganyctiphanes norvegica | 3.10 | 0.35 | 3.03 | 0.40 |
| Nematoscelis atlantica | 1.41 | 0.16 | 1.52 | 0.09 |
| Euphausiacea n.i. | 22.54 | 0.76 | 6.82 | 6.08 |
| Isopoda | 0.56 | 0.11 | 1.52 | 0.04 |
| Isopoda n.i. | 0.56 | 0.11 | 1.52 | 0.04 |
| OSTEICHTHYES | 27.02 | 40.67 | 56.11 | 48.56 |
| Ceratoscopelus maderensis | 2.82 | 3.57 | 5.30 | 1.30 |
| Chauliodus sloani | 0.28 | 0.04 | 0.76 | 0.01 |
| Diaphus holti | 0.28 | 0.89 | 0.76 | 0.03 |
| Diaphus sp. | 0.85 | 0.18 | 1.52 | 0.06 |
| Electrona risso | 1.41 | 3.16 | 2.27 | 0.40 |
| Gadiculus argenteus | 0.28 | 0.11 | 0.76 | 0.01 |
| Hygophum benoiti | 0.28 | 0.13 | 0.76 | 0.01 |
| Hygophum hygomii | 0.28 | 0.32 | 0.76 | 0.02 |
| Hygophum sp. | 0.28 | 0.04 | 0.76 | 0.01 |
| Macroramphosus scolopax | 0.28 | 0.15 | 0.76 | 0.01 |
| Myctophum punctatum | 0.28 | 0.29 | 0.76 | 0.02 |
| Ophidion rochei | 0.28 | 0.37 | 0.76 | 0.02 |
| Peristedion cataphractum | 0.28 | 0.09 | 0.76 | 0.01 |
| Sardina pilchardus | 0.56 | 3.77 | 1.52 | 0.25 |
| Sardinella aurita | 0.56 | 5.74 | 1.52 | 0.37 |
| Stomias boa | 0.28 | 0.06 | 0.76 | 0.01 |
| Synchiropus phaeton | 0.28 | 0.39 | 0.76 | 0.02 |
| Congridae n.i. | 0.28 | 0.08 | 0.76 | 0.01 |
| Myctophidae n.i. | 0.56 | 1.07 | 1.52 | 0.09 |
| Osteichthyes n.i. | 16.62 | 20.22 | 32.58 | 45.90 |
| TUNICATA | 0.28 | 0.04 | 0.76 | 0.01 |
| Ascidiacea n.i. | 0.28 | 0.04 | 0.76 | 0.01 |
| OTHER | 4.79 | 4.69 | 12.88 | 4.22 |
| Digested | 4.51 | 4.57 | 12.12 | 4.21 |
| Feathers (Mergus merganser) | 0.28 | 0.12 | 0.76 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Iglio, C.; Albano, M.; Tiralongo, F.; Famulari, S.; Rinelli, P.; Savoca, S.; Spanò, N.; Capillo, G. Biological and Ecological Aspects of the Blackmouth Catshark (Galeus melastomus Rafinesque, 1810) in the Southern Tyrrhenian Sea. J. Mar. Sci. Eng. 2021, 9, 967. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9090967
D’Iglio C, Albano M, Tiralongo F, Famulari S, Rinelli P, Savoca S, Spanò N, Capillo G. Biological and Ecological Aspects of the Blackmouth Catshark (Galeus melastomus Rafinesque, 1810) in the Southern Tyrrhenian Sea. Journal of Marine Science and Engineering. 2021; 9(9):967. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9090967
Chicago/Turabian StyleD’Iglio, Claudio, Marco Albano, Francesco Tiralongo, Sergio Famulari, Paola Rinelli, Serena Savoca, Nunziacarla Spanò, and Gioele Capillo. 2021. "Biological and Ecological Aspects of the Blackmouth Catshark (Galeus melastomus Rafinesque, 1810) in the Southern Tyrrhenian Sea" Journal of Marine Science and Engineering 9, no. 9: 967. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9090967