Alkaloid Concentrations of Lolium perenne Infected with Epichloë festucae var. lolii with Different Detection Methods—A Re-Evaluation of Intoxication Risk in Germany?

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Dataset 1: Field Study in Germany
2.2. Dataset 2: Alkaloid Concentrations throughout the Year in a Field Study
2.3. Dataset 3: Common Garden Study
2.4. Alkaloid Detection Methods
2.5. Statistical Analyses
2.6. Data Availability
3. Results
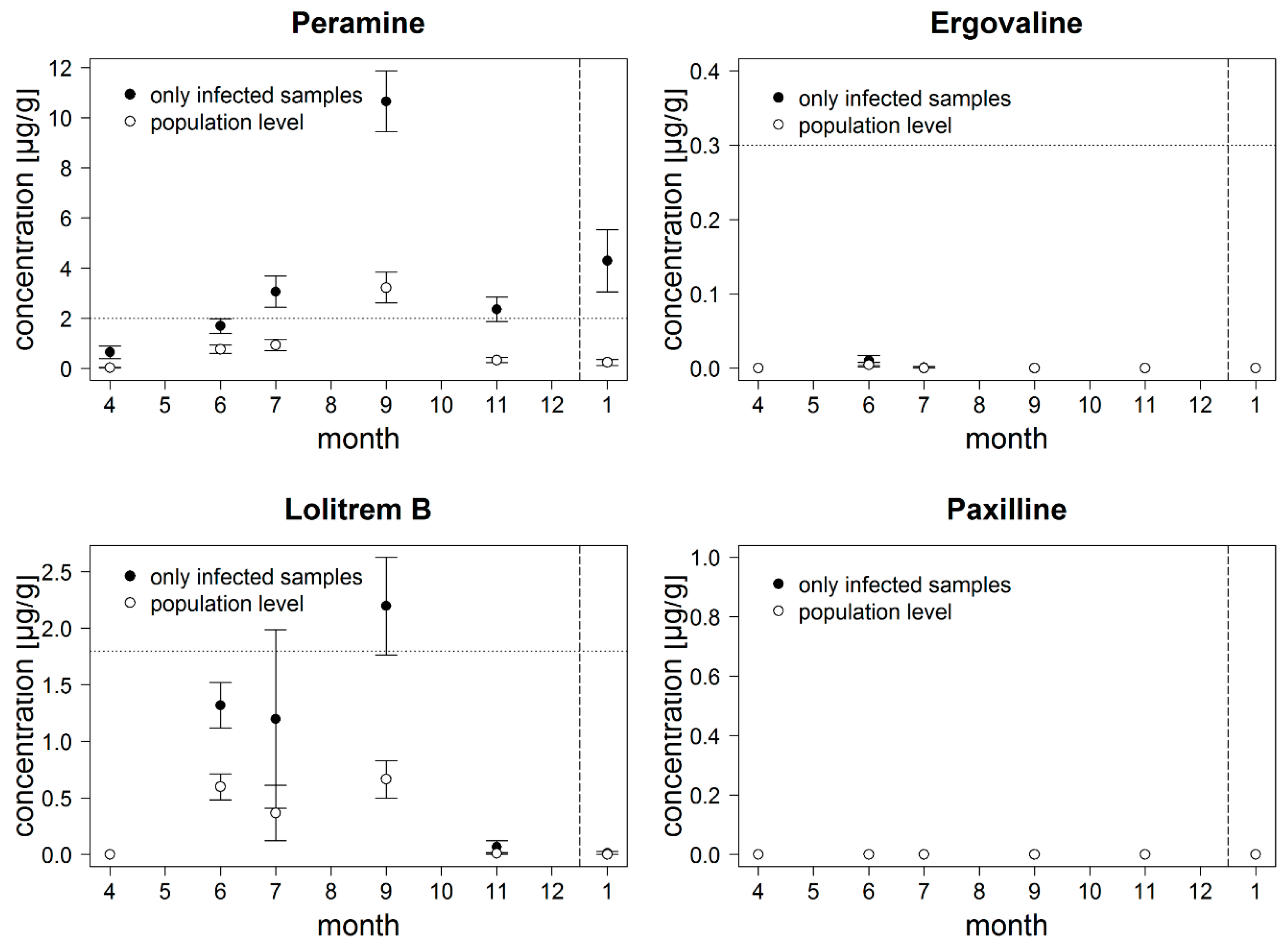
3.1. Dataset 1: Field Study in Germany
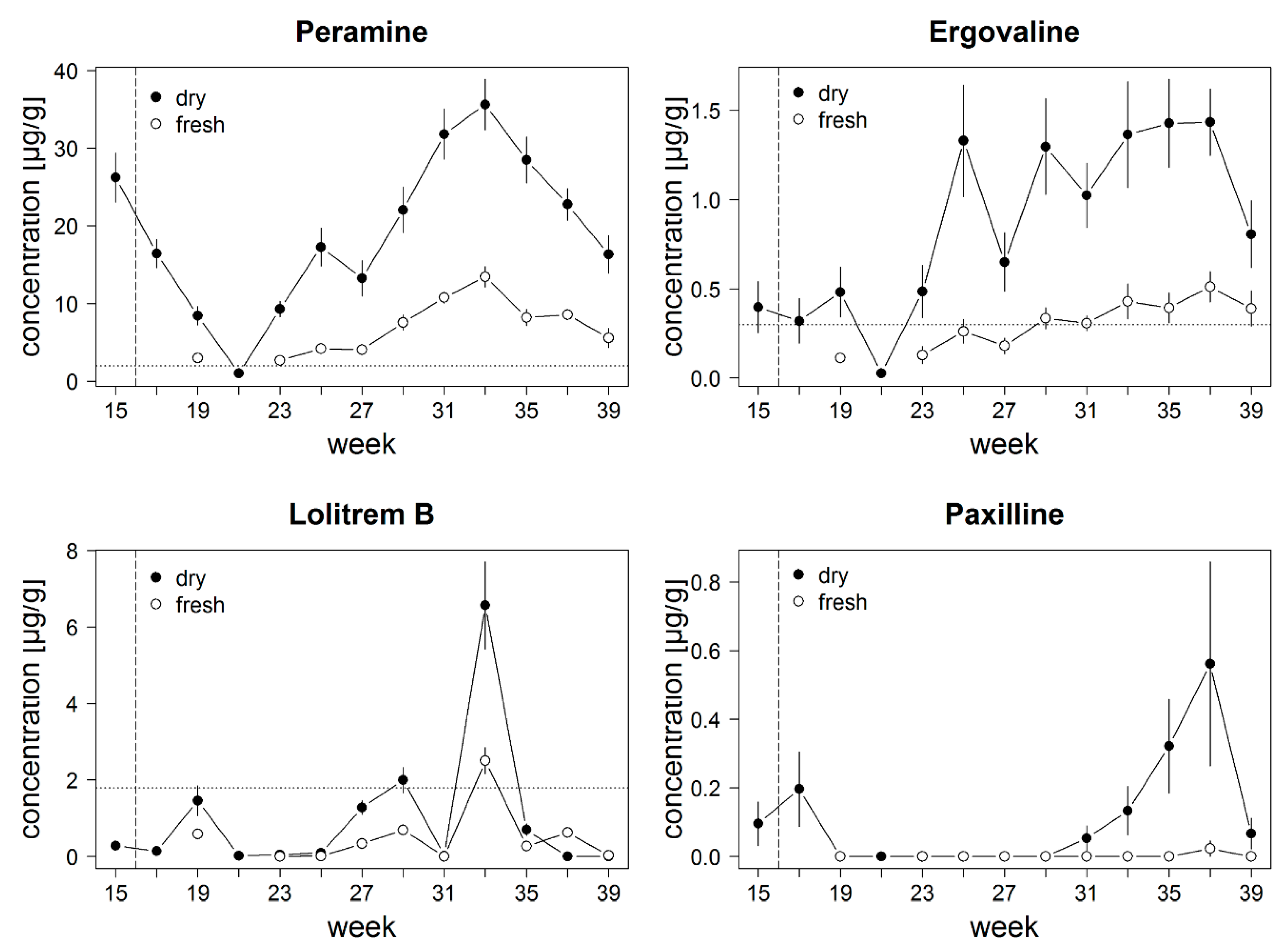
3.2. Dataset 2: Alkaloid Concentrations throughout the Year in a Field Study
3.3. Dataset 3: Common Garden Study
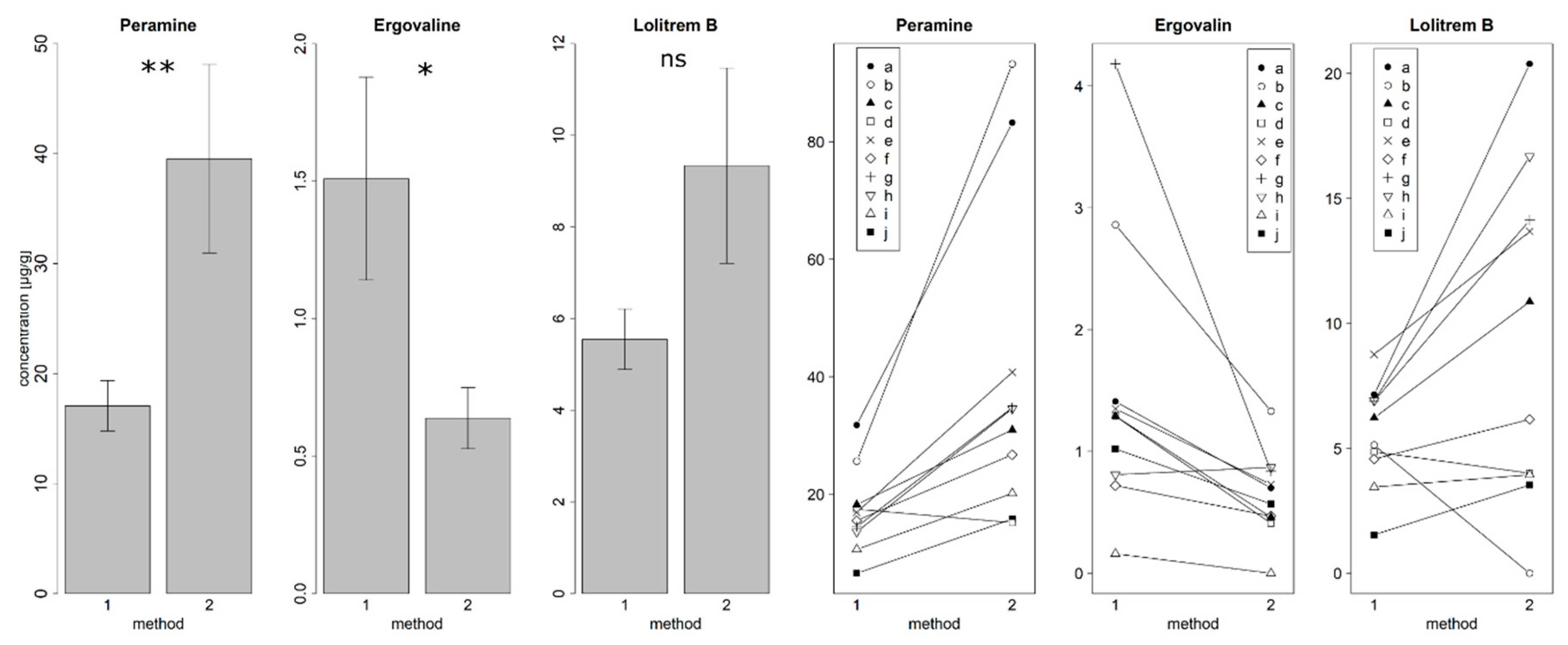
3.4. Comparison of Two Alkaloid Detection Methods
4. Discussion
4.1. Field Study in Germany
4.2. Alkaloid Concentrations throughout the Year in a Field Study
4.3. Common Garden Study
4.4. Comparison of Two Alkaloid Detection Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497. [Google Scholar] [CrossRef] [Green Version]
- Pitt, J.I. Toxigenic fungi and mycotoxins. Br. Med. Bull. 2000, 56, 184–192. [Google Scholar] [CrossRef] [Green Version]
- Sampson, K. The systemic infection of grasses by Epichloë typhina (Pers.) Tul. Trans. Br. Mycol. Soc. 1933, 18, 30–47. [Google Scholar] [CrossRef]
- Müller, C.B.; Krauss, J. Symbiosis between grasses and asexual fungal endophytes. Curr. Opin. Plant Biol. 2005, 8, 450–456. [Google Scholar] [CrossRef]
- Schardl, C.L. Epichloë species: Fungal symbionts of grasses. Annu. Rev. Phytopathol. 1996, 34, 109–130. [Google Scholar] [CrossRef]
- Bourguignon, M.; Nelson, J.A.; Carlisle, E.; Ji, H.; Dinkins, R.D.; Phillips, T.D.; McCulley, R.L. Ecophysiological responses of tall fescue genotypes to fungal endophyte infection, elevated temperature, and precipitation. Crop Sci. 2015, 55, 2895–2909. [Google Scholar] [CrossRef] [Green Version]
- Gundel, P.E.; Sun, P.; Charlton, N.D.; Young, C.A.; Miller, T.E.X.; Rudgers, J.A. Simulated folivory increases vertical transmission of fungal endophytes that deter herbivores and alter tolerance to herbivory in Poa autumnalis. Ann. Bot. 2020, 125, 981–991. [Google Scholar] [CrossRef]
- Hume, D.E.; Ryan, G.D.; Gibert, A.; Helander, M.; Mirlohi, A.; Sabzalian, M.R. Epichloë fungal endophytes for grassland ecosystems. In Sustainable Agriculture Reviews; Sustainable Agriculture Reviews; Lichtfouse, E., Ed.; Springer International Publishing: Cham, Switzerland, 2016; Volume 19, pp. 233–305. ISBN 978-3-319-26777-7. [Google Scholar]
- Imlach, W.L.; Finch, S.C.; Dunlop, J.; Meredith, A.L.; Aldrich, R.W.; Dalziel, J.E. The molecular mechanism of “ryegrass staggers”, a neurological disorder of K+ channels. J. Pharmacol. Exp. Ther. 2008, 327, 657–664. [Google Scholar] [CrossRef] [Green Version]
- Di Menna, M.; Finch, S.C.; Popay, A.J.; Smith, B.L. A review of the Neotyphodium lolii/Lolium perenne symbiosis and its associated effects on animal and plant health, with particular emphasis on ryegrass staggers. N. Z. Vet. J. 2012, 60, 315–328. [Google Scholar] [CrossRef]
- Reddy, P.; Deseo, M.A.; Ezernieks, V.; Guthridge, K.; Spangenberg, G.; Rochfort, S. Toxic indole diterpenes from endophyte-infected perennial ryegrass Lolium perenne L.: Isolation and stability. Toxins 2019, 11, 16. [Google Scholar] [CrossRef] [Green Version]
- Rowan, D.D.; Dymock, J.J.; Brimble, M.A. Effect of fungal metabolite peramine and analogs on feeding and development of argentine stem weevil (Listronotus bonariensis). J. Chem. Ecol. 1990, 16, 1683–1695. [Google Scholar] [CrossRef]
- Siegel, M.R.; Bush, L.P. Defensive chemicals in grass-fungal endophyte associations. Recent Adv. Phytochem. 1996, 30, 55–62. [Google Scholar]
- Hewitt, K.G.; Mace, W.J.; McKenzie, C.M.; Matthew, C.; Popay, A.J. Fungal alkaloid occurrence in endophyte-infected perennial ryegrass during seedling establishment. J. Chem. Ecol. 2020, 46, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Popay, A.J.; Wyatt, R.T. Resistance to Argentine stem weevil in perennial ryegrass infected with endophytes producing different alkaloids. N. Z. Plant Prot. 1995, 48, 229–236. [Google Scholar] [CrossRef]
- Craig, A.M.; Blythe, L.L.; Duringer, J.M. The role of the Oregon State University Endophyte Service Laboratory in diagnosing clinical cases of endophyte toxicoses. J. Agric. Food Chem. 2014, 62, 7376–7381. [Google Scholar] [CrossRef]
- Hovermale, J.T.; Craig, A.M. Correlation of ergovaline and lolitrem B levels in endophyte-infected perennial ryegrass (Lolium perenne). J. Vet. Diagn. Investig. 2001, 13, 323–327. [Google Scholar] [CrossRef] [Green Version]
- Saikkonen, K.; Lehtonen, P.; Helander, M.; Koricheva, J.; Faeth, S.H. Model systems in ecology: Dissecting the endophyte-grass literature. Trends Plant Sci. 2006, 11, 428–433. [Google Scholar] [CrossRef]
- Wang, T.; Frandsen, H.L.; Christiansson, N.R.; Rosendal, S.E.; Pedersen, M.; Smedsgaard, J. Pyrrolizidine alkaloids in honey: Quantification with and without standards. Food Control 2019, 98, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Finch, S.C.; Thom, E.R.; Babu, J.V.; Hawkes, A.D.; Waugh, C.D. The evaluation of fungal endophyte toxin residues in milk. N. Z. Vet. J. 2013, 61, 11–17. [Google Scholar] [CrossRef]
- Finch, S.C.; Fletcher, L.R.; Babu, J.V. The evaluation of endophyte toxin residues in sheep fat. N. Z. Vet. J. 2012, 60, 56–60. [Google Scholar] [CrossRef]
- Spangenberg, G.; Wang, Z.-Y.; Potrykus, I. Biotechnology in Forage and Turf Grass Improvement; Springer Science & Business Media: Berlin, Germany, 2012; ISBN 978-3-642-72051-2. [Google Scholar]
- König, J.; Fuchs, B.; Krischke, M.; Mueller, M.J.; Krauss, J. Hide and seek—Infection rates and alkaloid concentrations of Epichloё festucae var. lolii in Lolium perenne along a land-use gradient in Germany. Grass Forage Sci. 2018, 73, 510–516. [Google Scholar] [CrossRef]
- Vikuk, V.; Young, C.A.; Lee, S.T.; Nagabhyru, P.; Krischke, M.; Mueller, M.J.; Krauss, J. Infection rates and alkaloid patterns of different grass species with systemic Epichloë endophytes. Appl. Environ. Microbiol. 2019, 85, e00465-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Easton, S.; Tapper, B. Neotyphodium research and application in New Zealand. In Neotyphodium in Cool-Season Grasses; Wiley-Blackwell—Blackwell Publishing: Oxford, UK, 2008; pp. 35–42. ISBN 978-0-470-38491-6. [Google Scholar]
- Kauppinen, M.; Saikkonen, K.; Helander, M.; Pirttilä, A.M.; Wäli, P.R. Epichloë grass endophytes in sustainable agriculture. Nat. Plants 2016, 2, 15224. [Google Scholar] [CrossRef] [PubMed]
- Malinowski, D.P.; Belesky, D.P. Adaptations of endophyte-infected cool-season grasses to environmental stresses: Mechanisms of drought and mineral stress tolerance. Crop Sci. 2000, 40, 923–940. [Google Scholar] [CrossRef]
- Gwinn, K.D.; Fribourg, H.A.; Waller, J.C.; Saxton, A.M.; Smith, M.C. Changes in Neotyphodium coenophialum infestation levels in tall fescue pastures due to different grazing pressures. Crop Sci. 1998, 38, 201–204. [Google Scholar] [CrossRef]
- Jensen, A.M.D.; Roulund, N. Occurrence of Neotyphodium endophytes in permanent grassland with perennial ryegrass (Lolium perenne) in Denmark. Agric. Ecosyst. Environ. 2004, 104, 419–427. [Google Scholar] [CrossRef]
- Shelby, R.A.; Olsovska, J.; Havlicek, V.; Flieger, M. Analysis of ergot alkaloids in endophyte-infected tall fescue by liquid chromatography electrospray ionization mass spectrometry. J. Agric. Food Chem. 1997, 45, 4674–4679. [Google Scholar] [CrossRef]
- Krauss, J.; Vikuk, V.; Young, C.A.; Krischke, M.; Mueller, M.J.; Baerenfaller, K. Epichloë endophyte infection rates and alkaloid content in commercially available grass seed mixtures in Europe. Microorganisms 2020, 8, 498. [Google Scholar] [CrossRef] [Green Version]
- Rudolph, W.; Remane, D.; Wissenbach, D.K.; Peters, F.T. Development and validation of an ultrahigh performance liquid chromatography-high resolution tandem mass spectrometry assay for nine toxic alkaloids from endophyte-infected pasture grasses in horse serum. J. Chromatogr. A 2018, 1560, 35–44. [Google Scholar] [CrossRef]
- Gallagher, R.T.; Hawkes, A.D.; Stewart, J.M. Rapid determination of the neurotoxin lolitrem B in perennial ryegrass by high-performance liquid chromatography with fluorescence detection. J. Chromatogr. A 1985, 321, 217–226. [Google Scholar] [CrossRef]
- Repussard, C.; Tardieu, D.; Alberich, M.; Guerre, P. A new method for the determination of lolitrem B in plant materials. Anim. Feed Sci. Technol. 2014, 193, 141–147. [Google Scholar] [CrossRef]
- Bauer, J.I.; Gross, M.; Cramer, B.; Wegner, S.; Hausmann, H.; Hamscher, G.; Usleber, E. Detection of the tremorgenic mycotoxin paxilline and its desoxy analog in ergot of rye and barley: A new class of mycotoxins added to an old problem. Anal. Bioanal. Chem. 2017, 409, 5101–5112. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.; Krischke, M.; Mueller, M.J.; Krauss, J. Peramine and lolitrem B from endophyte-grass associations cascade up the food chain. J. Chem. Ecol. 2013, 39, 1385–1389. [Google Scholar] [CrossRef]
- Helander, M.; Phillips, T.; Faeth, S.H.; Bush, L.P.; McCulley, R.; Saloniemi, I.; Saikkonen, K. Alkaloid quantities in endophyte-infected tall fescue are affected by the plant-fungus combination and environment. J. Chem. Ecol. 2016, 42, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Rottinghaus, G.E.; Garner, G.B.; Cornell, C.N.; Ellis, J.L. HPLC method for quantitating ergovaline in endophyte-infested tall fescue: Seasonal variation of ergovaline levels in stems with leaf sheaths, leaf blades, and seed heads. J. Agric. Food Chem. 1991, 39, 112–115. [Google Scholar] [CrossRef]
- Shelby, R.A.; Flieger, M. Improved method of analysis for ergovaline in tall fescue by high-performance liquid chromatography. J. Agric. Food Chem. 1997, 45, 1797–1800. [Google Scholar] [CrossRef]
- Spiering, M.J.; Davies, E.; Tapper, B.A.; Schmid, J.; Lane, G.A. Simplified extraction of ergovaline and peramine for analysis of tissue distribution in endophyte-infected grass tillers. J. Agric. Food Chem. 2002, 50, 5856–5862. [Google Scholar] [CrossRef]
- Baldwin, I.T. Short-term damage-induced increases in tobacco alkaloids protect plants. Oecologia 1988, 75, 367–370. [Google Scholar] [CrossRef]
- Glauser, G.; Dubugnon, L.; Mousavi, S.A.R.; Rudaz, S.; Wolfender, J.-L.; Farmer, E.E. Velocity estimates for signal propagation leading to systemic jasmonic acid accumulation in wounded Arabidopsis. J. Biol. Chem. 2009, 284, 34506–34513. [Google Scholar] [CrossRef] [Green Version]
- Vassiliadis, S.; Elkins, A.C.; Reddy, P.; Guthridge, K.M.; Spangenberg, G.C.; Rochfort, S.J. A simple LC–MS method for the quantitation of alkaloids in endophyte-infected perennial ryegrass. Toxins 2019, 11, 649. [Google Scholar] [CrossRef] [Green Version]
- Fischer, M.; Bossdorf, O.; Gockel, S.; Hänsel, F.; Hemp, A.; Hessenmoeller, D.; Korte, G.; Nieschulze, J.; Pfeiffer, S.; Prati, D.; et al. Implementing large-scale and long-term functional biodiversity research: The Biodiversity Exploratories. Basic Appl. Ecol. 2010, 11, 473–485. [Google Scholar] [CrossRef]
- Blüthgen, N.; Dormann, C.F.; Prati, D.; Klaus, V.H.; Kleinebecker, T.; Hölzel, N.; Alt, F.; Boch, S.; Gockel, S.; Hemp, A.; et al. A quantitative index of land-use intensity in grasslands: Integrating mowing, grazing and fertilization. Basic Appl. Ecol. 2012, 13, 207–220. [Google Scholar] [CrossRef]
- Krauss, J.; Härri, S.A.; Bush, L.; Husi, R.; Bigler, L.; Power, S.A.; Müller, C.B. Effects of fertilizer, fungal endophytes and plant cultivar on the performance of insect herbivores and their natural enemies. Funct. Ecol. 2007, 21, 107–116. [Google Scholar] [CrossRef]
- Spiering, M.J.; Lane, G.A.; Christensen, M.J.; Schmid, J. Distribution of the fungal endophyte Neotyphodium lolii is not a major determinant of the distribution of fungal alkaloids in Lolium perenne plants. Phytochemistry 2005, 66, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.J.; de Bonth, A.C.M.; Briggs, L.R.; Caradus, J.R.; Finch, S.C.; Fleetwood, D.J.; Fletcher, L.R.; Hume, D.E.; Johnson, R.D.; Popay, A.J.; et al. The exploitation of epichloae endophytes for agricultural benefit. Fungal Divers. 2013, 60, 171–188. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; SAGE Publications: Thousand Oaks, CA, USA, 2019; ISBN 978-1-5443-3648-0. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 26. [Google Scholar] [CrossRef] [Green Version]
- Faeth, S.H.; Fagan, W.F. Fungal endophytes: Common host plant symbionts but uncommon mutualists. Integr. Comp. Biol. 2002, 42, 360–368. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, B.; Krischke, M.; Mueller, M.J.; Krauss, J. Herbivore-specific induction of defence metabolites in a grass-endophyte association. Funct. Ecol. 2017, 31, 318–324. [Google Scholar] [CrossRef]
- Soto-Barajas, M.C.; Vázquez-de-Aldana, B.R.; Álvarez, A.; Zabalgogeazcoa, I. Sympatric Epichloë species and chemotypic profiles in natural populations of Lolium perenne. Fungal Ecol. 2019, 39, 231–241. [Google Scholar] [CrossRef]
- Fuchs, B.; Krischke, M.; Mueller, M.J.; Krauss, J. Plant age and seasonal timing determine endophyte growth and alkaloid biosynthesis. Fungal Ecol. 2017, 29, 52–58. [Google Scholar] [CrossRef]
- Jensen, A.M.D. Endophyte persistence and toxin (lolitrem b) production in a Danish seed crop of perennial ryegrass. Eur. J. Agron. 2005, 23, 68–78. [Google Scholar] [CrossRef]
- Repussard, C.; Zbib, N.; Tardieu, D.; Guerre, P. Ergovaline and lolitrem B concentrations in perennial ryegrass in field culture in southern France: Distribution in the plant and impact of climatic factors. J. Agric. Food Chem. 2014, 62, 12707–12712. [Google Scholar] [CrossRef]
- Vázquez de Aldana, B.R.; Leinonen, P.; Zabalgogeazcoa, I.; Helander, M.; Saikkonen, K. Variation and plasticity in Epichloë alkaloid content of Festuca rubra across Europe. Fungal Ecol. 2020, 47, 100942. [Google Scholar] [CrossRef]
- Kagan, I.A.; Kirch, B.H.; Thatcher, C.D.; Strickland, J.R.; Teutsch, C.D.; Elvinger, F.; Pleasant, R.S. Seasonal and Diurnal Variation in Simple Sugar and Fructan Composition of Orchardgrass Pasture and Hay in the Piedmont Region of the United States. J. Equine Vet. Sci. 2011, 31, 488–497. [Google Scholar] [CrossRef]
- Jensen, J.B.; Gonzalez, V.T.; Guevara, D.U.; Bhuvaneswari, T.V.; Wali, P.R.; Tejesvi, M.V.; Pirttila, A.M.; Bazely, D.; Vicari, M.; Brathen, K.A. Kit for detection of fungal endophytes of grasses yields inconsistent results. Methods Ecol. Evol. 2011, 2, 197–201. [Google Scholar] [CrossRef]
- Bauer, J.I.; Gross, M.; Cramer, B.; Humpf, H.-U.; Hamscher, G.; Usleber, E. Immunochemical analysis of paxilline and ergot alkaloid mycotoxins in grass seeds and plants. J. Agric. Food Chem. 2018, 66, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.; Breuer, T.; Findling, S.; Krischke, M.; Mueller, M.J.; Holzschuh, A.; Krauss, J. Enhanced aphid abundance in spring desynchronizes predator-prey and plant-microorganism interactions. Oecologia 2017, 183, 469–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köhler, G.; Perner, J.; Schumacher, J. Grasshopper population dynamics and meteorological parameters—Lessons from a case study. Ecography 1999, 22, 205–212. [Google Scholar] [CrossRef]
- Machado, S.; Paulsen, G.M. Combined effects of drought and high temperature on water relations of wheat and sorghum. Plant Soil 2001, 233, 179–187. [Google Scholar] [CrossRef]
- Schardl, C.L.; Young, C.A.; Faulkner, J.R.; Florea, S.; Pan, J. Chemotypic diversity of Epichloë, fungal symbionts of grasses. Fungal Ecol. 2012, 5, 331–344. [Google Scholar] [CrossRef]
- Hennessy, L.M.; Popay, A.J.; Finch, S.C.; Clearwater, M.J.; Cave, V.M. Temperature and plant genotype alter alkaloid concentrations in ryegrass infected with an Epichloë endophyte and this affects an insect herbivore. Front. Plant Sci. 2016, 7, 1097. [Google Scholar] [CrossRef] [PubMed]
- McCulley, R.L.; Bush, L.P.; Carlisle, A.E.; Ji, H.; Nelson, J.A. Warming reduces tall fescue abundance but stimulates toxic alkaloid concentrations in transition zone pastures of the U.S. Front. Chem. 2014, 2, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Infection Rates | Peramine | Lolitrem B | |||||||
|---|---|---|---|---|---|---|---|---|---|
| χ2 dF | χ2 | p | dF | F | p | dF | F | p | |
| Region | 2 | 17.45 | <0.001 *** | 2, 107 | 1.84 | 0.19 | 2, 75 | 2.77 | 0.11 |
| LUI | 1 | 0.53 | 0.47 | 1, 107 | 3.21 | 0.10 | 1, 75 | 0.24 | 0.64 |
| Mowing (y/n) | 1 | 0.09 | 0.76 | 1, 107 | 0.004 | 0.95 | 1, 75 | 0.01 | 0.94 |
| Mowing tot | 1 | 0.09 | 0.76 | 1, 107 | 0.10 | 0.75 | 1, 75 | 0.001 | 0.98 |
| Grazing (y/n) | 1 | 0.81 | 0.37 | 1, 107 | 0.08 | 0.79 | 1, 75 | 0.02 | 0.90 |
| Grazing tot | 1 | 0.12 | 0.73 | 1, 107 | 0.22 | 0.64 | 1, 75 | 0.01 | 0.91 |
| Fertilization (y/n) | 1 | 1.70 | 0.19 | 1, 107 | 0.33 | 0.58 | 1, 75 | 0.15 | 0.70 |
| Fertilization tot | 1 | 0.36 | 0.55 | 1, 107 | 4.14 | 0.08 | 1, 75 | 0.09 | 0.78 |
| Individual Concentrations | Mean Concentrations per Study Site | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Peramine | Ergovaline | Lolitrem B | Paxilline | Peramine | Ergovaline | Lolitrem B | Paxilline | |||||||||
| Conc. (µg/g) | IND | % | IND | % | IND | % | IND | % | SITE | % | SITE | % | SITE | % | SITE | % |
| 0.0 | 4 | 3.1 | 130 | 99.2 | 54 | 41.2 | 130 | 99.2 | 38 | 57.6 | 65 | 98.5 | 42 | 63.6 | 65 | 98.5 |
| 0–0.3 | 13 | 9.9 | 1 | 0.8 | 24 | 18.30 | 1 | 0.8 | 15 | 22.7 | 1 | 1.5 | 23 | 34.9 | 1 | 1.5 |
| 0.3–1 | 52 | 39.7 | 0 | 0.0 | 32 | 24.4 | 0 | 0.0 | 9 | 13.6 | 0 | 0.0 | 1 | 1.5 | 0 | 0.0 |
| 1.0–2.0 | 36 | 27.5 | 0 | 0.0 | 10 | 7.6 | 0 | 0.0 | 4 | 6.1 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 |
| 2.0–3.0 | 10 | 7.6 | 0 | 0.0 | 4 | 3.1 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 |
| 3.0–5.0 | 10 | 7.6 | 0 | 0.0 | 3 | 2.3 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 |
| 5.0–10.0 | 4 | 3.1 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 |
| >10.0 | 2 | 1.5 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 |
| Max (µg/g) | 12.0 | 0.17 | 4.2 | 0.15 | 1.7 | 0.0 | 0.5 | 0.01 | ||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vikuk, V.; Fuchs, B.; Krischke, M.; Mueller, M.J.; Rueb, S.; Krauss, J. Alkaloid Concentrations of Lolium perenne Infected with Epichloë festucae var. lolii with Different Detection Methods—A Re-Evaluation of Intoxication Risk in Germany? J. Fungi 2020, 6, 177. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030177
Vikuk V, Fuchs B, Krischke M, Mueller MJ, Rueb S, Krauss J. Alkaloid Concentrations of Lolium perenne Infected with Epichloë festucae var. lolii with Different Detection Methods—A Re-Evaluation of Intoxication Risk in Germany? Journal of Fungi. 2020; 6(3):177. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030177
Chicago/Turabian StyleVikuk, Veronika, Benjamin Fuchs, Markus Krischke, Martin J. Mueller, Selina Rueb, and Jochen Krauss. 2020. "Alkaloid Concentrations of Lolium perenne Infected with Epichloë festucae var. lolii with Different Detection Methods—A Re-Evaluation of Intoxication Risk in Germany?" Journal of Fungi 6, no. 3: 177. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030177