
Tracking Fungal Growth: Establishment of Arp1 as a Marker for Polarity Establishment and Active Hyphal Growth in Filamentous Ascomycetes

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media and Growth Conditions
2.2. Multiple Sequence Alignment of Fungal Arp1
2.3. Construction of Plasmids
2.4. Generation of the Smarp1 Deletion Strain ∆Smarp1

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Strain | Genotype | Reference |
|---|---|---|
| Sordaria macrospora | ||
| DSM997 | S. macrospora wild-type (wt) | DSMZ |
| S23442 | mutation in fus1-1 gene, brownish ascospores, fertile | [64] |
| fus::RH2B | ectopic integration of pRH2B into S23442; hygR, pt, Pgpd::hh2b::tdTomato::TtrpC | [65] |
| wt::TagRFP-T | ectopic integration of ptRFP_nat into DSM997; natR, ssi, Pccg1::TagRFP-T::TtrpC | [61] |
| wt::GG-C-F-mNG | ectopic integration of pGG-C-F-mNG in DSM997; natR, ssi, Pgpd::mNG::3xFLAG::TtrpC | [59] |
| ∆Smku70 | ∆Smku70::natR, fertile | [66] |
| Sm::Smarp1-TagRFP-T | ectopic integration of pSmarp1-TagRFP-T in DSM997; hygR, ssi, Smarp1P::Smarp1::TagRFP-T::TtrpC | this study |
| Sm::Smarp1-mNG | ectopic integration of pSmarp1-mNG in DSM997; natR, ssi, Smarp1P::Smarp1::mNG::TtrpC | this study |
| ∆Smarp1 | ∆Smarp1::hygR, sterile | this study |
| ∆Smarp1::Smarp1 | ectopic integration of pSmarp1 in ∆Smarp1; hygR, natR, ssi, Smarp1P::Smarp1::Smarp1T | this study |
| ∆Smarp1::Smarp1-TagRFP-T | ectopic integration of pSmarp1-TagRFP-T in ∆Smarp1; hygR, natR, ssi, Smarp1P::Smarp1::TagRFP-T::TtrpC | this study |
| Colletotrichum graminicola | ||
| CgM2 | C. graminicola wild-type (wt); also referred to as M1.001 | [67] |
| CgM2::Smarp1-mNG | ectopic integration of pSmarp1-mNG in CgM2; natR, ssi, Smarp1P::Smarp1::mNG::TtrpC | this study |
| CgM2::Cgarp1-TagRFP-T | ectopic integration of pCgarp1-TagRFP-T in CgM2; genR, ssi, Cgarp1P::Cgarp1::TagRFP-T::TtrpC | this study |
2.5. Generation of S. macrospora Strains for Fluorescence Microscopy
2.6. Generation of C. graminicola Strains
2.7. Fluorescence Microscopy
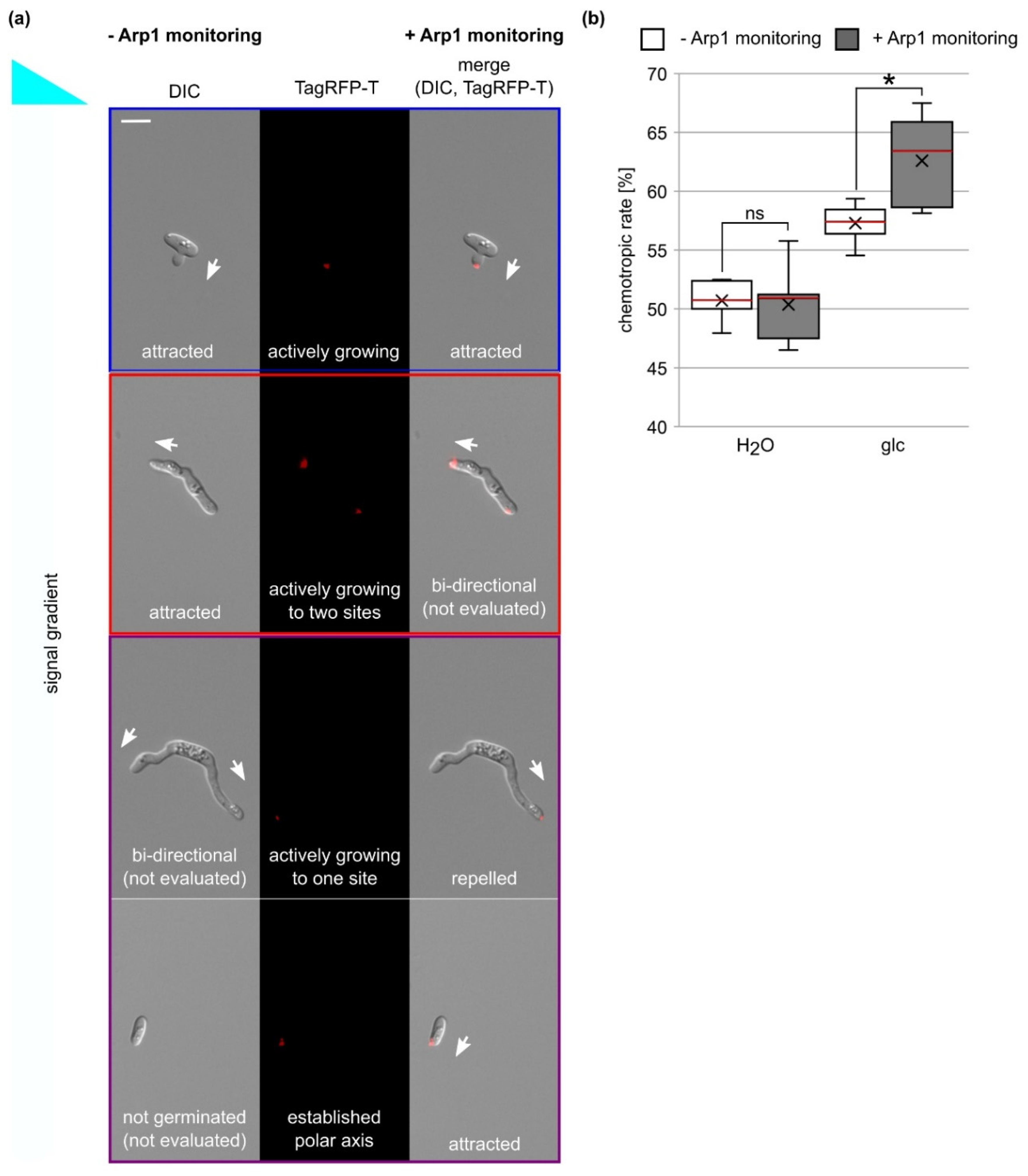
2.8. Analysis of Chemotropic Growth Using the Arp1-TagRFP-T Marker
2.9. Protein Sample Preparation and Western Blot Hybridization
2.10. Phenotypic Analysis
3. Results
3.1. Arp1 Deletion in S. macrospora Results in a Severe Growth Defect That Can Only Partially Be Complemented
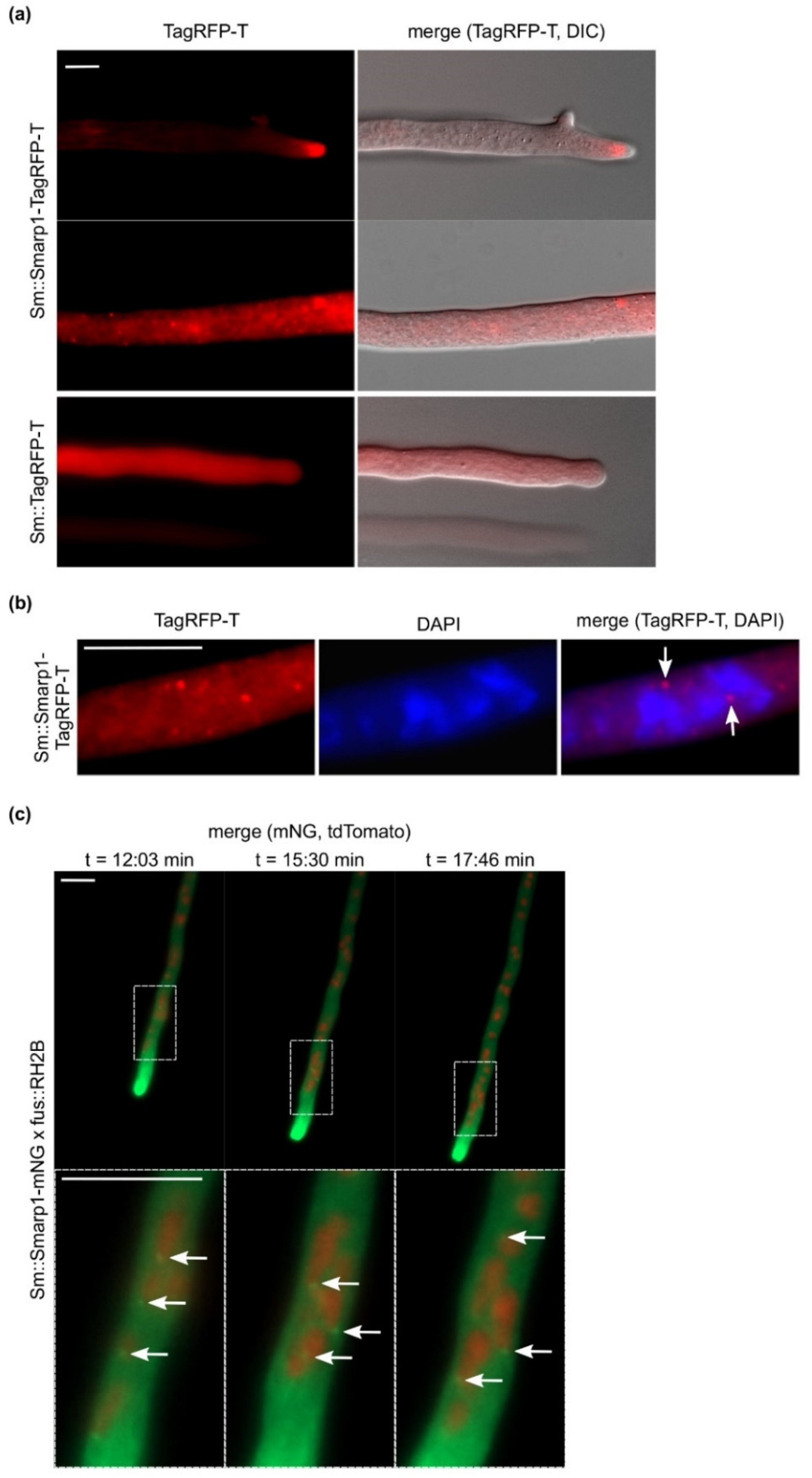
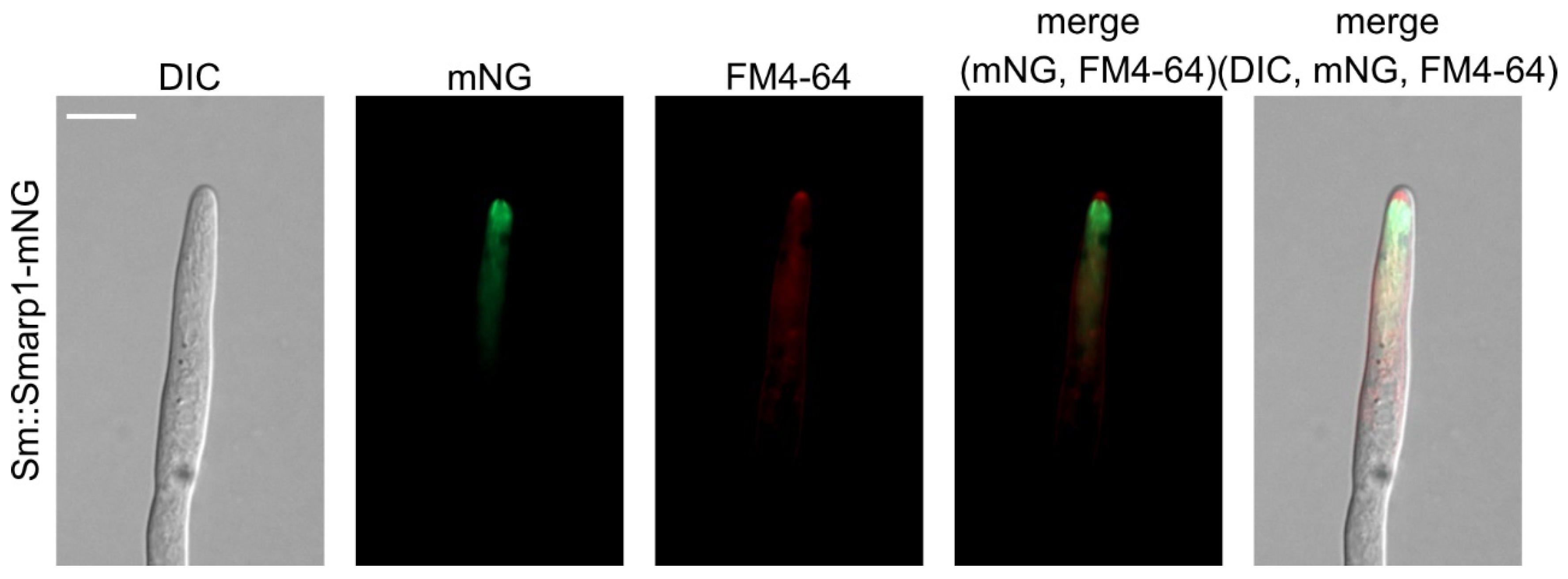
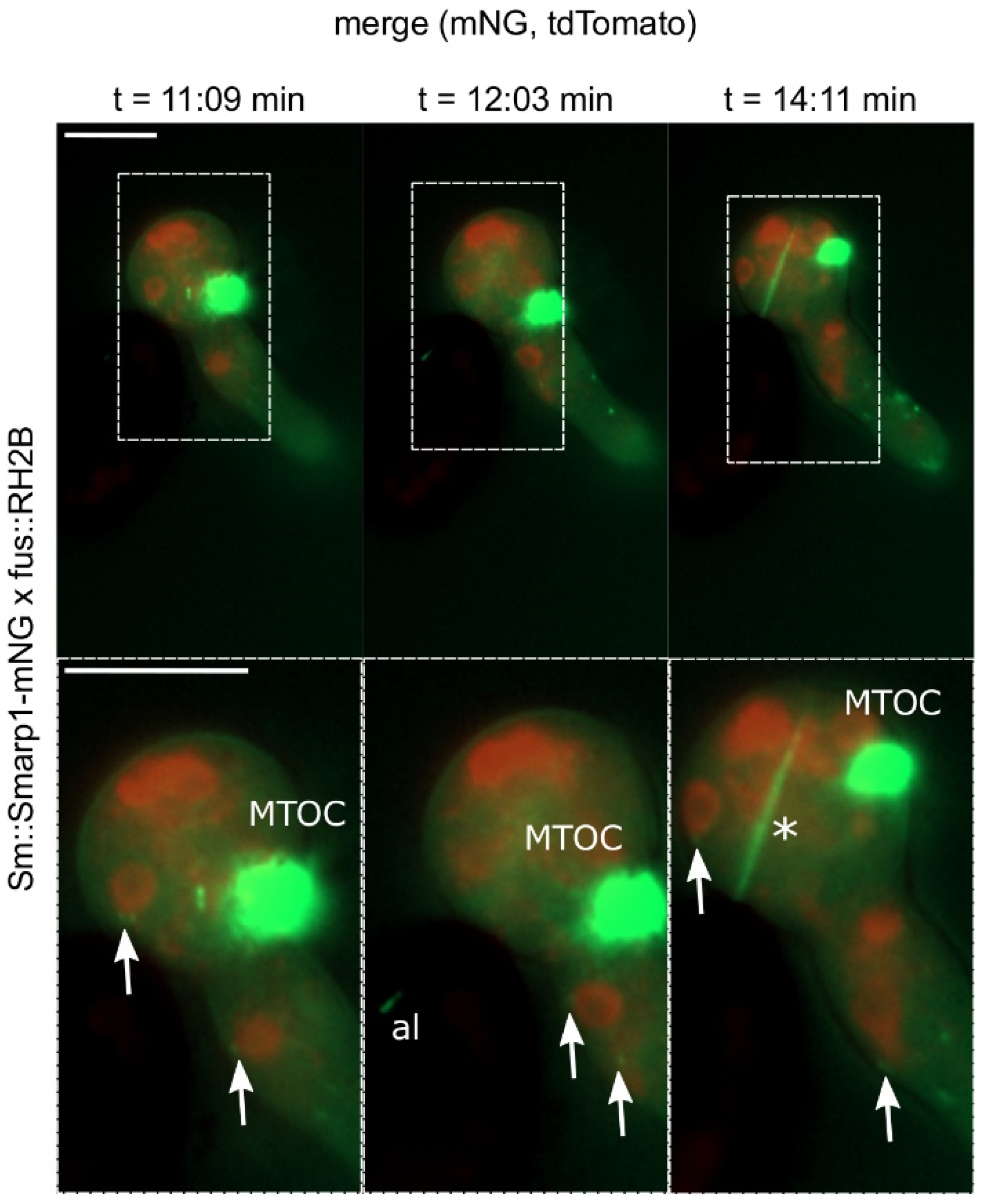
3.2. SmArp1 Localization Is a Dynamic Process at Growing Hyphal Tips and in Proximity to the Nucleus
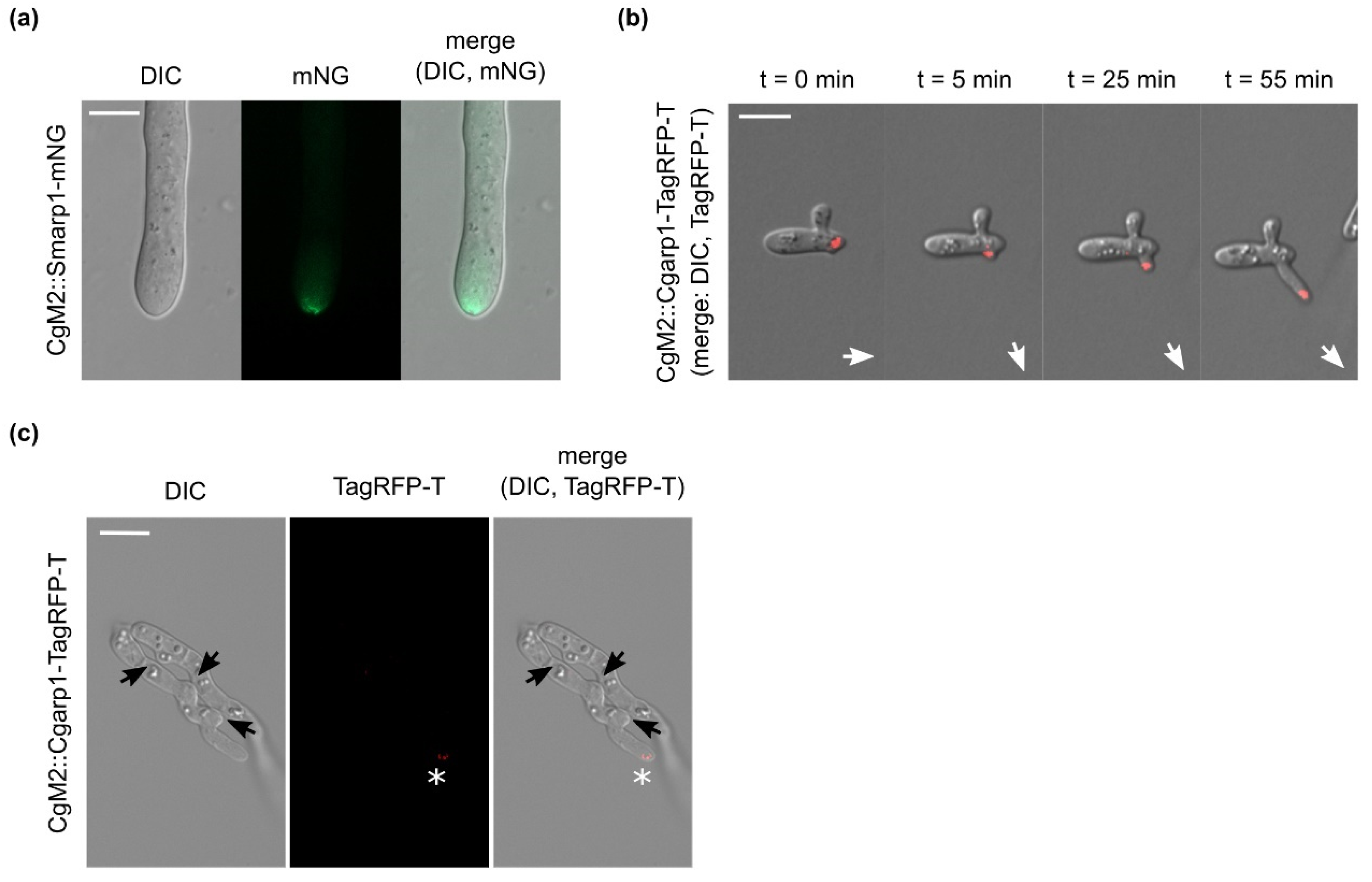
3.3. Dynamic Localization of Arp1 in C. graminicola
3.4. Absence of CgArp1 localization to C. graminicola future fusion points
3.5. Arp1 Localizes Subapical to SPK
3.6. S. macrospora Arp1 Shows Dynamic Localization during Ascospore Germination
3.7. Analysis of Chemotropic Growth Is Improved by the Usage of the Arp1-TagRFP-T Marker
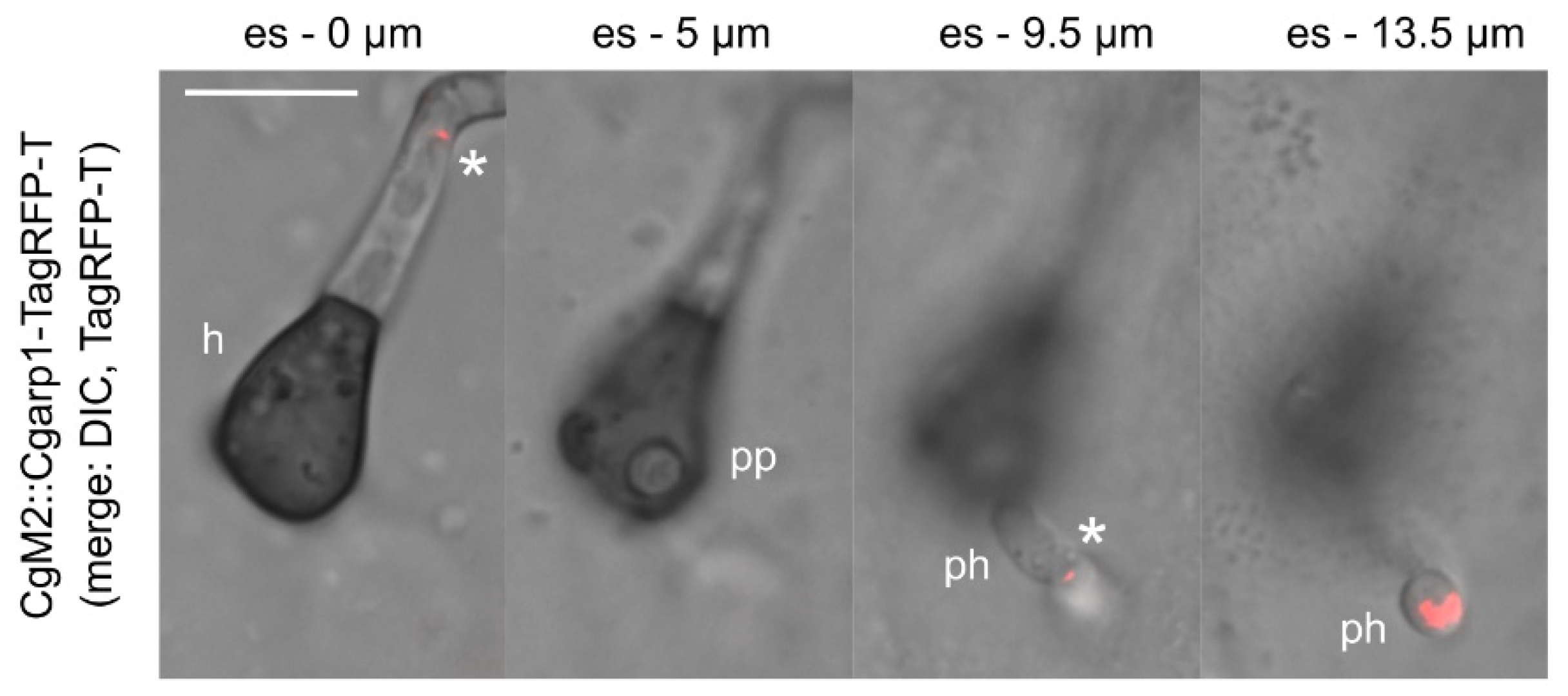
3.8. Tracking of Penetration Hyphae into Plant Tissue by Monitoring of Arp1 Localization in C. graminicola
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Riquelme, M. Tip growth in filamentous fungi: A road trip to the apex. Annu. Rev. Microbiol. 2013, 67, 587–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, R.; Zekert, N.; Takeshita, N. Polarized growth in fungi-interplay between the cytoskeleton, positional markers and membrane domains. Mol. Microbiol. 2008, 68, 813–826. [Google Scholar] [CrossRef]
- Mela, A.P.; Rico-Ramírez, A.M.; Glass, N.L. Syncytia in Fungi. Cells 2020, 9, 2255. [Google Scholar] [CrossRef]
- Lichius, A.; Lord, K.M. Chemoattractive mechanisms in filamentous fungi. Open Mycol. J. 2014, 8, 28–57. [Google Scholar] [CrossRef] [Green Version]
- Brand, A.; Gow, N.A. Mechanisms of hypha orientation of fungi. Curr. Opin. Microbiol. 2009, 12, 350–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turrà, D.; Nordzieke, D.; Vitale, S.; El Ghalid, M.; Di Pietro, A. Hyphal chemotropism in fungal pathogenicity. Semin. Cell Dev. Biol. 2016, 57, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.; Becker, Y.; Green, K.; Scott, B. The endophytic symbiont Epichloe festucae establishes an epiphyllous net on the surface of Lolium perenne leaves by development of an expressorium, an appressorium-like leaf exit structure. New Phytol. 2016, 211, 240–254. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.L.; Zhou, T.T.; Guo, H.S. Hyphopodium-specific VdNoxB/VdPls1-Dependent ROS-Ca2+ signaling is required for plant infection by Verticillium dahliae. PLoS Pathog. 2016, 12, e1005793. [Google Scholar] [CrossRef] [Green Version]
- Ryder, L.S.; Talbot, N.J. Regulation of appressorium development in pathogenic fungi. Curr. Opin. Plant Biol. 2015, 26, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Werner, S.; Sugui, J.A.; Steinberg, G.; Deising, H.B. A chitin synthase with a myosin-like motor domain is essential for hyphal growth, appressorium differentiation, and pathogenicity of the maize anthracnose fungus Colletotrichum graminicola. Mol. Plant Microbe Interact. 2007, 20, 1555–1567. [Google Scholar] [CrossRef] [Green Version]
- Riquelme, M.; Aguirre, J.; Bartnicki-García, S.; Braus, G.H.; Feldbrügge, M.; Fleig, U.; Hansberg, W.; Herrera-Estrella, A.; Kamper, J.; Kück, U.; et al. Fungal morphogenesis, from the polarized growth of hyphae to complex reproduction and infection structures. Microbiol. Mol. Biol. Rev. 2018, 82, e00068-17. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, Y.; Shoji, J.Y.; Arioka, M.; Kitamoto, K. Endocytosis is crucial for cell polarity and apical membrane recycling in the filamentous fungus Aspergillus oryzae. Eukaryot. Cell 2009, 8, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Momany, M. Polarity in filamentous fungi: Establishment, maintenance and new axes. Curr. Opin. Microbiol. 2002, 5, 580–585. [Google Scholar] [CrossRef]
- Riquelme, M.; Martínez-Núñez, L. Hyphal ontogeny in Neurospora crassa: A model organism for all seasons. F1000Research 2016, 5, 2801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendland, J.; Philippsen, P. Cell polarity and hyphal morphogenesis are controlled by multiple rho-protein modules in the filamentous ascomycete Ashbya gossypii. Genetics 2001, 157, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Woods, B.; Lew, D.J. Polarity establishment by Cdc42: Key roles for positive feedback and differential mobility. Small GTPases 2019, 10, 130–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, K.E.; Kang, P.J.; Park, H.-O. Regulation of Cdc42 for polarized growth in budding yeast. Microb. Cell 2020, 7, 175. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.D. Cdc42/Rho GTPases in fungi: Variations on a common theme. Mol. Microbiol. 2011, 79, 1123–1127. [Google Scholar] [CrossRef]
- Drees, B.L.; Sundin, B.; Brazeau, E.; Caviston, J.P.; Chen, G.C.; Guo, W.; Kozminski, K.G.; Lau, M.W.; Moskow, J.J.; Tong, A.; et al. A protein interaction map for cell polarity development. J. Cell Biol. 2001, 154, 549–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.G.; Arkowitz, R.A. Cell polarization in budding and fission yeasts. FEMS Microbiol. Rev. 2014, 38, 228–253. [Google Scholar] [CrossRef] [Green Version]
- Sharpless, K.E.; Harris, S.D. Functional characterization and localization of the Aspergillus nidulans formin SEPA. Mol. Biol. Cell 2002, 13, 469–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichius, A.; Yáñez-Gutiérrez, M.E.; Read, N.D.; Castro-Longoria, E. Comparative live-cell imaging analyses of SPA-2, BUD-6 and BNI-1 in Neurospora crassa reveal novel features of the filamentous fungal polarisome. PLoS ONE 2012, 7, e30372. [Google Scholar] [CrossRef] [Green Version]
- Giesbert, S.; Siegmund, U.; Schumacher, J.; Kokkelink, L.; Tudzynski, P. Functional analysis of BcBem1 and its interaction partners in Botrytis cinerea: Impact on differentiation and virulence. PLoS ONE 2014, 9, e95172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, S.D.; Hamer, L.; Sharpless, K.E.; Hamer, J.E. The Aspergillus nidulans sepA gene encodes an FH1/2 protein involved in cytokinesis and the maintenance of cellular polarity. EMBO J. 1997, 16, 3474–3483. [Google Scholar] [CrossRef] [PubMed]
- Lichius, A.; Goryachev, A.B.; Fricker, M.D.; Obara, B.; Castro-Longoria, E.; Read, N.D. CDC-42 and RAC-1 regulate opposite chemotropisms in Neurospora crassa. J. Cell Sci. 2014, 127, 1953–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, P.; Nguyen, T.A.; Wong, J.Y.; Lee, M.; Nguyen, T.A.; Fan, J.S.; Yang, D.; Jedd, G. Spitzenkörper assembly mechanisms reveal conserved features of fungal and metazoan polarity scaffolds. Nat. Commun. 2020, 11, 2830. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, G. Hyphal growth: A tale of motors, lipids, and the Spitzenkörper. Eukaryot. Cell 2007, 6, 351–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, S.D.; Read, N.D.; Roberson, R.W.; Shaw, B.; Seiler, S.; Plamann, M.; Momany, M. Polarisome meets Spitzenkörper: Microscopy, genetics, and genomics converge. Eukaryot. Cell 2005, 4, 225–229. [Google Scholar] [CrossRef] [Green Version]
- Wendland, J.; Walther, A. Ashbya gossypii: A model for fungal developmental biology. Nat. Rev. Microbiol. 2005, 3, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Mouriño-Pérez, R.R.; Roberson, R.W.; Bartnicki-García, S. Microtubule dynamics and organization during hyphal growth and branching in Neurospora crassa. Fungal Genet. Biol. 2006, 43, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Konzack, S.; Rischitor, P.E.; Enke, C.; Fischer, R. The role of the kinesin motor KipA in microtubule organization and polarized growth of Aspergillus nidulans. Mol. Biol. Cell 2005, 16, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, G.; Peñalva, M.A.; Riquelme, M.; Wösten, H.A.; Harris, S.D. Cell biology of hyphal growth. Microbiol. Spectr. 2017, 5, FUNK-0034-2016. [Google Scholar] [CrossRef] [Green Version]
- Holleran, E.A.; Ligon, L.A.; Tokito, M.; Stankewich, M.C.; Morrow, J.S.; Holzbaur, E.L. βIII spectrin binds to the Arp1 subunit of dynactin. J. Biol. Chem. 2001, 276, 36598–36605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reck-Peterson, S.L.; Redwine, W.B.; Vale, R.D.; Carter, A.P. The cytoplasmic dynein transport machinery and its many cargoes. Nat. Rev. Mol. Cell Biol. 2018, 19, 382–398. [Google Scholar] [CrossRef] [PubMed]
- Eckley, D.M.; Schroer, T.A. Interactions between the evolutionarily conserved, actin-related protein, Arp11, actin, and Arp1. Mol. Biol. Cell 2003, 14, 2645–2654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christopher, L.; Fletcher, L.; Dykstra, C. Cloning and Identification of Arp1, an Actin-Related Protein from Pneumocystis carinii. J. Eukaryot. Microbiol. 1995, 42, 142–149. [Google Scholar] [CrossRef]
- Schafer, D.; Gill, S.R.; Cooper, J.; Heuser, J.; Schroer, T. Ultrastructural analysis of the dynactin complex: An actin-related protein is a component of a filament that resembles F-actin. J. Cell Biol. 1994, 126, 403–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenz, J.H.; Schuchardt, I.; Straube, A.; Steinberg, G. A dynein loading zone for retrograde endosome motility at microtubule plus-ends. EMBO J. 2006, 25, 2275–2286. [Google Scholar] [CrossRef] [Green Version]
- Egan, M.J.; Tan, K.; Reck-Peterson, S.L. Lis1 is an initiation factor for dynein-driven organelle transport. J. Cell Biol. 2012, 197, 971–982. [Google Scholar] [CrossRef]
- Zhang, J.; Li, S.; Fischer, R.; Xiang, X. Accumulation of cytoplasmic dynein and dynactin at microtubule plus ends in Aspergillus nidulans is kinesin dependent. Mol. Biol. Cell 2003, 14, 1479–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, M.; Kilaru, S.; Ashwin, P.; Lin, C.; Severs, N.J.; Steinberg, G. Controlled and stochastic retention concentrates dynein at microtubule ends to keep endosomes on track. EMBO J. 2011, 30, 652–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckley, D.M.; Gill, S.R.; Melkonian, K.A.; Bingham, J.B.; Goodson, H.V.; Heuser, J.E.; Schroer, T.A. Analysis of dynactin subcomplexes reveals a novel actin-related protein associated with the arp1 minifilament pointed end. J. Cell Biol. 1999, 147, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Plamann, M.; Minke, P.F.; Tinsley, J.H.; Bruno, K.S. Cytoplasmic dynein and actin-related protein Arp1 are required for normal nuclear distribution in filamentous fungi. J. Cell Biol. 1994, 127, 139–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Fritsch, E.; Maniatis, T. (Eds.) Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Colot, H.V.; Park, G.; Turner, G.E.; Ringelberg, C.; Crew, C.M.; Litvinkova, L.; Weiss, R.L.; Borkovich, K.A.; Dunlap, J.C. A high-throughput gene knockout procedure for Neurospora reveals functions for multiple transcription factors. Proc. Natl. Acad. Sci. USA 2006, 103, 10352–10357. [Google Scholar] [CrossRef] [Green Version]
- James, P.; Halladay, J.; Craig, E.A. Genomic libraries and a host strain designed for highly efficient two-hybrid selection in yeast. Genetics 1996, 144, 1425–1436. [Google Scholar] [CrossRef]
- Walz, M.; Kück, U. Transformation of Sordaria macrospora to hygromycin B resistance: Characterization of transformants by electrophoretic karyotyping and tetrad analysis. Curr. Genet. 1995, 29, 88–95. [Google Scholar] [CrossRef]
- Elleuche, S.; Pöggeler, S. Visualization of peroxisomes via SKL-tagged DsRed protein in Sordaria macrospora. Fungal Genet. Rep. 2008, 55, 9–12. [Google Scholar] [CrossRef] [Green Version]
- Esser, K. Cryptogams—Cyanobacteria, Algae, Fungi, Lichens; Cambridge University Press: London, UK, 1982. [Google Scholar]
- Nowrousian, M.; Ringelberg, C.; Dunlap, J.C.; Loros, J.J.; Kück, U. Cross-species microarray hybridization to identify developmentally regulated genes in the filamentous fungus Sordaria macrospora. Mol. Genet. Genom. 2005, 273, 137–149. [Google Scholar] [CrossRef]
- Bernhards, Y.; Pöggeler, S. The phocein homologue SmMOB3 is essential for vegetative cell fusion and sexual development in the filamentous ascomycete Sordaria macrospora. Curr. Genet. 2011, 57, 133–149. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleemann, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N.; et al. Lifestyle transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nat. Genet. 2012, 44, 1060–1065. [Google Scholar] [CrossRef]
- Nordzieke, D.E.; Sanken, A.; Antelo, L.; Raschke, A.; Deising, H.B.; Pöggeler, S. Specialized infection strategies of falcate and oval conidia of Colletotrichum graminicola. Fungal Genet. Biol. 2019, 133, 103276. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Han, M.V.; Zmasek, C.M. phyloXML: XML for evolutionary biology and comparative genomics. BMC Bioinform. 2009, 10, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Reschka, E.J.; Nordzieke, S.; Valerius, O.; Braus, G.H.; Pöggeler, S. A novel STRIPAK complex component mediates hyphal fusion and fruiting-body development in filamentous fungi. Mol. Microbiol. 2018, 110, 513–532. [Google Scholar] [CrossRef]
- Dahlmann, T.A.; Terfehr, D.; Becker, K.; Teichert, I. Golden Gate vectors for efficient gene fusion and gene deletion in diverse filamentous fungi. Curr. Genet. 2021, 67, 317–330. [Google Scholar] [CrossRef]
- Klix, V.; Nowrousian, M.; Ringelberg, C.; Loros, J.J.; Dunlap, J.C.; Pöggeler, S. Functional characterization of MAT1-1-specific mating-type genes in the homothallic ascomycete Sordaria macrospora provides new insights into essential and nonessential sexual regulators. Eukaryot. Cell 2010, 9, 894–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, A.; Otte, K.L.; Stahlhut, G.; Pöggeler, S. Establishment of the monomeric yellow-green fluorescent protein mNeonGreen for life cell imaging in mycelial fungi. AMB Express 2020, 10, 222. [Google Scholar] [CrossRef] [PubMed]
- Pöggeler, S.; Masloff, S.; Hoff, B.; Mayrhofer, S.; Kück, U. Versatile EGFP reporter plasmids for cellular localization of recombinant gene products in filamentous fungi. Curr. Genet. 2003, 43, 54–61. [Google Scholar] [CrossRef]
- Werner, A.; Otte, K.; Stahlhut, G.; Hanke, L.M.; Pöggeler, S. The glyoxysomal protease LON2 is involved in fruiting-body development, ascosporogenesis and stress resistance in Sordaria macrospora. J. Fungi 2021, 7, 82. [Google Scholar] [CrossRef]
- Bloemendal, S.; Bernhards, Y.; Bartho, K.; Dettmann, A.; Voigt, O.; Teichert, I.; Seiler, S.; Wolters, D.A.; Pöggeler, S.; Kück, U. A homologue of the human STRIPAK complex controls sexual development in fungi. Mol. Microbiol. 2012, 84, 310–323. [Google Scholar] [CrossRef] [PubMed]
- Shibayama, M.; Ooi, K.; Johnson, R.; Scott, B.; Itoh, Y. Suppression of tandem-multimer formation during genetic transformation of the mycotoxin-producing fungus Penicillium paxilli by disrupting an orthologue of Aspergillus nidulans uvsC. Curr. Genet. 2002, 42, 59–65. [Google Scholar] [CrossRef]
- Nowrousian, M.; Teichert, I.; Masloff, S.; Kück, U. Whole-genome sequencing of Sordaria macrospora mutants identifies developmental genes. G3 Genes Genomes Genet. 2012, 2, 261–270. [Google Scholar]
- Nordzieke, S.; Zobel, T.; Franzel, B.; Wolters, D.A.; Kück, U.; Teichert, I. A fungal sarcolemmal membrane-associated protein (SLMAP) homolog plays a fundamental role in development and localizes to the nuclear envelope, endoplasmic reticulum, and mitochondria. Eukaryot. Cell 2015, 14, 345–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pöggeler, S.; Kück, U. Highly efficient generation of signal transduction knockout mutants using a fungal strain deficient in the mammalian ku70 ortholog. Gene 2006, 378, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Forgey, W.M.; Blanco, M.H.; Loegering, W.Q. Differences in pathological capabilities and host specificity of Colletotrichum graminicola on Zea mays. Plant Dis. Rep. 1978, 62, 573–576. [Google Scholar]
- Horbach, R.; Graf, A.; Weihmann, F.; Antelo, L.; Mathea, S.; Liermann, J.C.; Opatz, T.; Thines, E.; Aguirre, J.; Deising, H.B. Sfp-type 4′-phosphopantetheinyl transferase is indispensable for fungal pathogenicity. Plant Cell 2009, 21, 3379–3396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schunke, C.; Pöggeler, S.; Nordzieke, D.E. A 3D printed device for easy and reliable quantification of fungal chemotropic growth. Front. Microbiol. 2020, 11, 584525. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Fleissner, A.; Herzog, S. Signal exchange and integration during self-fusion in filamentous fungi. Semin. Cell Dev. Biol. 2016, 57, 76–83. [Google Scholar] [CrossRef]
- Read, N.D.; Goryachev, A.B.; Lichius, A. The mechanistic basis of self-fusion between conidial anastomosis tubes during fungal colony initiation. Fungal Biol. Rev. 2012, 26, 1–11. [Google Scholar] [CrossRef]
- Grove, S.N.; Bracker, C.E. Protoplasmic organization of hyphal tips among fungi: Vesicles and Spitzenkörper. J. Bacteriol. 1970, 104, 989–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Löpez-Franco, R.; Bracker, C.E. Diversity and dynamics of the Spitzenkörper in growing hyphal tips of higher fungi. J. Bacteriol. 1996, 195, 90–111. [Google Scholar]
- Virag, A.; Harris, S.D. The Spitzenkörper: A molecular perspective. Mycol. Res. 2006, 110, 4–13. [Google Scholar] [CrossRef]
- Riquelme, M.; Reynaga-Peña, C.G.; Gierz, G.; Bartnicki-García, S. What determines growth direction in fungal hyphae? Fungal Genet. Biol. 1998, 24, 101–109. [Google Scholar] [CrossRef]
- Peñalva, M.A. Tracing the endocytic pathway of Aspergillus nidulans with FM4-64. Fungal Genet. Biol. 2005, 42, 963–975. [Google Scholar] [CrossRef]
- Fischer-Parton, S.; Parton, R.; Hickey, P.; Dijksterhuis, J.; Atkinson, H.; Read, N. Confocal microscopy of FM4-64 as a tool for analysing endocytosis and vesicle trafficking in living fungal hyphae. J. Microsc. 2000, 198, 246–259. [Google Scholar] [CrossRef] [Green Version]
- Teichert, I. Nuclear dynamics during ascospore germination in Sordaria macrospora. Fungal Genet. Biol. 2017, 98, 20–22. [Google Scholar] [CrossRef] [PubMed]
- Turrà, D.; El Ghalid, M.; Rossi, F.; Di Pietro, A. Fungal pathogen uses sex pheromone receptor for chemotropic sensing of host plant signals. Nature 2015, 527, 521–524. [Google Scholar] [CrossRef]
- Mendgen, K.; Hahn, M.; Deising, H. Morphogenesis and mechanisms of penetration by plant pathogenic fungi. Annu. Rev. Phytopathol. 1996, 34, 367–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryder, L.S.; Dagdas, Y.F.; Mentlak, T.A.; Kershaw, M.J.; Thornton, C.R.; Schuster, M.; Chen, J.; Wang, Z.; Talbot, N.J. NADPH oxidases regulate septin-mediated cytoskeletal remodeling during plant infection by the rice blast fungus. Proc. Natl. Acad. Sci. USA 2013, 110, 3179–3184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brun, S.; Malagnac, F.; Bidard, F.; Lalucque, H.; Silar, P. Functions and regulation of the Nox family in the filamentous fungus Podospora anserina: A new role in cellulose degradation. Mol. Microbiol. 2009, 74, 480–496. [Google Scholar] [CrossRef] [PubMed]
- Oakley, B.R.; Oakley, C.E.; Yoon, Y.; Jung, M.K. γ-Tubulin is a component of the spindle pole body that is essential for microtubule function in Aspergillus nidulans. Cell 1990, 61, 1289–1301. [Google Scholar] [CrossRef]
- Takeshita, N.; Manck, R.; Grün, N.; de Vega, S.H.; Fischer, R. Interdependence of the actin and the microtubule cytoskeleton during fungal growth. Curr. Opin. Microbiol. 2014, 20, 34–41. [Google Scholar] [CrossRef]
- Xiang, X.; Han, G.; Winkelmann, D.A.; Zuo, W.; Morris, N.R. Dynamics of cytoplasmic dynein in living cells and the effect of a mutation in the dynactin complex actin-related protein Arp1. Curr. Biol. 2000, 10, 603–606. [Google Scholar] [CrossRef] [Green Version]
- Baas, P.W.; Lin, S. Hooks and comets: The story of microtubule polarity orientation in the neuron. Dev. Neurobiol. 2011, 71, 403–418. [Google Scholar] [CrossRef] [Green Version]
- Dujardin, D.L.; Barnhart, L.E.; Stehman, S.A.; Gomes, E.R.; Gundersen, G.G.; Vallee, R.B. A role for cytoplasmic dynein and LIS1 in directed cell movement. J. Cell Biol. 2003, 163, 1205–1211. [Google Scholar] [CrossRef] [Green Version]
- Baas, P.W.; Vidya, N.C.; Myers, K.A. Axonal transport of microtubules: The long and short of it. Traffic 2006, 7, 490–498. [Google Scholar] [CrossRef]
- Fink, G.; Steinberg, G. Dynein-dependent motility of microtubules and nucleation sites supports polarization of the tubulin array in the fungus Ustilago maydis. Mol. Biol. Cell 2006, 17, 3242–3253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minke, P.; Lee, I.; Tinsley, J.; Plamann, M. A Neurospora crassa Arp1 mutation affecting cytoplasmic dynein and dynactin localization. Mol. Gen. Genet. MGG 2000, 264, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, N.; Fischer, R. On the role of microtubules, cell end markers, and septal microtubule organizing centres on site selection for polar growth in Aspergillus nidulans. Fungal Biol. 2011, 115, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, M.; Gierz, G.; Bartnicki-García, S. Dynein and dynactin deficiencies affect the formation and function of the Spitzenkörper and distort hyphal morphogenesis of Neurospora crassa. Microbiology 2000, 146, 1743–1752. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, G. On the move: Endosomes in fungal growth and pathogenicity. Nat. Rev. Microbiol. 2007, 5, 309–316. [Google Scholar] [CrossRef]
- Araujo-Palomares, C.L.; Castro-Longoria, E.; Riquelme, M. Ontogeny of the Spitzenkörper in germlings of Neurospora crassa. Fungal Genet. Biol. 2007, 44, 492–503. [Google Scholar] [CrossRef]
- Hickey, P.C.; Jacobson, D.; Read, N.D.; Glass, N.L. Live-cell imaging of vegetative hyphal fusion in Neurospora crassa. Fungal Genet. Biol. 2002, 37, 109–119. [Google Scholar] [CrossRef]
- Roca, M.G.; Kuo, H.C.; Lichius, A.; Freitag, M.; Read, N.D. Nuclear dynamics, mitosis, and the cytoskeleton during the early stages of colony initiation in Neurospora crassa. Eukaryot. Cell 2010, 9, 1171–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berepiki, A.; Lichius, A.; Shoji, J.Y.; Tilsner, J.; Read, N.D. F-actin dynamics in Neurospora crassa. Eukaryot. Cell 2010, 9, 547–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, M.S.; Glass, N.L. Communicate and fuse: How filamentous fungi establish and maintain an interconnected mycelial network. Front. Microbiol. 2019, 10, 619. [Google Scholar] [CrossRef] [PubMed]
- Suelmann, R.; Sievers, N.; Fischer, R. Nuclear traffic in fungal hyphae: In vivo study of nuclear migration and positioning in Aspergillus nidulans. Mol. Microbiol. 1997, 25, 757–769. [Google Scholar] [CrossRef] [PubMed]
- Veith, D.; Scherr, N.; Efimov, V.P.; Fischer, R. Role of the spindle-pole-body protein ApsB and the cortex protein ApsA in microtubule organization and nuclear migration in Aspergillus nidulans. J. Cell Sci. 2005, 118, 3705–3716. [Google Scholar] [CrossRef] [Green Version]
- Fischer, R.; Timberlake, W.E. Aspergillus nidulans apsA (anucleate primary sterigmata) encodes a coiled-coil protein required for nuclear positioning and completion of asexual development. J. Cell Biol. 1995, 128, 485–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steuer, E.R.; Wordeman, L.; Schroer, T.A.; Sheetz, M.P. Localization of cytoplasmic dynein to mitotic spindles and kinetochores. Nature 1990, 345, 266–268. [Google Scholar] [CrossRef] [PubMed]






| Sm | Nc | Cg | Fo | Mo | An | Ro | Ml | Cc | Sco | Cn | Um | Pc | Sp | Ca | Sc | |
| Sm | 100 | 100 | 93 | 92 | 88 | 78 | 74 | 74 | 70 | 73 | 71 | 74 | 74 | 51 | 56 | 50 |
| Nc | 100 | 92 | 91 | 88 | 78 | 74 | 74 | 70 | 73 | 71 | 71 | 74 | 51 | 56 | 50 | |
| Cg | 100 | 94 | 87 | 79 | 74 | 74 | 69 | 73 | 71 | 73 | 76 | 52 | 57 | 51 | ||
| Fo | 100 | 87 | 78 | 74 | 74 | 70 | 74 | 72 | 74 | 75 | 51 | 57 | 50 | |||
| Mo | 100 | 78 | 74 | 75 | 69 | 70 | 70 | 73 | 74 | 51 | 59 | 52 | ||||
| An | 100 | 73 | 74 | 68 | 72 | 71 | 71 | 71 | 50 | 58 | 51 | |||||
| Ro | 100 | 99 | 77 | 81 | 79 | 81 | 82 | 54 | 60 | 50 | ||||||
| Ml | 100 | 77 | 82 | 78 | 82 | 81 | 54 | 60 | 50 | |||||||
| Cc | 100 | 87 | 80 | 78 | 74 | 49 | 54 | 48 | ||||||||
| Sco | 100 | 83 | 82 | 77 | 52 | 58 | 51 | |||||||||
| Cn | 100 | 80 | 74 | 52 | 59 | 50 | ||||||||||
| Um | 100 | 78 | 55 | 60 | 52 | |||||||||||
| Pc | 100 | 56 | 58 | 51 | ||||||||||||
| Sp | 100 | 47 | 44 | |||||||||||||
| Ca | 100 | 53 | ||||||||||||||
| Sc | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Groth, A.; Schunke, C.; Reschka, E.J.; Pöggeler, S.; Nordzieke, D.E. Tracking Fungal Growth: Establishment of Arp1 as a Marker for Polarity Establishment and Active Hyphal Growth in Filamentous Ascomycetes. J. Fungi 2021, 7, 580. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7070580
Groth A, Schunke C, Reschka EJ, Pöggeler S, Nordzieke DE. Tracking Fungal Growth: Establishment of Arp1 as a Marker for Polarity Establishment and Active Hyphal Growth in Filamentous Ascomycetes. Journal of Fungi. 2021; 7(7):580. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7070580
Chicago/Turabian StyleGroth, Anika, Carolin Schunke, Eva Johanna Reschka, Stefanie Pöggeler, and Daniela Elisabeth Nordzieke. 2021. "Tracking Fungal Growth: Establishment of Arp1 as a Marker for Polarity Establishment and Active Hyphal Growth in Filamentous Ascomycetes" Journal of Fungi 7, no. 7: 580. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7070580