3.2. Taxonomy
MycoBank number: MB 825096.
Type—Uganda. Western Region, Kibale National Park, Kabarole District, Makere University, N 0°30′, E 30°24′, on dead wood, 20 April 2002, L. Ryvarden, Ryvarden 44929 (K 133591, holotype).
Etymology—Africana (Lat.): refers to the distribution of the species in Africa.
Basidiomata—Gelatinous when rehydrated, greyish brown to fuscous, caespitose, resupinate to effused-reflexed; pileus sometimes observed, free lobed, margin undulate, projecting up to 1 cm, 1.5–2 mm thick, 0.2–0.4 mm thick when dry; upper surface tomentose, concentrically zoned with canescent zones and dark bands, becoming yellowish brown to orange-brown upon drying; hymenophore surface venose with folds, becoming greyish brown to fuscous upon drying.
Internal features—Medulla absent; crystals present, usually scattered in the hymenium; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, apical tips acute or obtuse, tufted, 300–500 × 1–2 µm; hyphae with clamp connections and simple septa, slightly inflated with a lumen in KOH, up to 4.5 µm in diam; basidia clavate, transversely 3-septate, with oil guttules, 50–80 × 4.5–6 µm, sterigmata sometimes observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, (11.5–)12–14(–14.2) × (4.8–)4.9–5.4(–5.5) µm, L = 12.54 µm, W = 5.12 µm, Q = 2.45 (n = 30/1).
Distribution—Kenya and Uganda.
Notes—
Auricularia africana belongs to the
A. mesenterica complex and is characterized by resupinate to effused-reflexed basidiomata with concentric zones on the upper surface. It is macro-morphologically similar to
A. pusio Berk. (
Figure 3g), but differs from the later by shorter abhymenial hairs (300–500 µm vs. 400–800 µm), slightly narrower basidia and basidiospores (4.5–6 µm vs. 5–7.5 µm and 4.9–5.4 µm vs. 5–6 µm). In the phylogeny,
A. africana is distantly related to
A. pusio and forms a single lineage with high support (
Figure 1 and
Figure 2).
Additional specimen (paratype) examined—Kenya. Central Province, Nairobi, Karura Forest, on angiosperm tree, May 1992, A. Osano, T 3 (K 34551).
(2) Auricularia americana Parmasto & I. Parmasto ex Audet, Boulet & Sirard
≡Auricularia americana Parmasto & I. Parmasto, Biblthca Mycol. 115: 137, 1987.
Basidiomata—Gelatinous when fresh, orange-brown to reddish brown, solitary, sometimes caespitose, sessile or substipitate; pileus cupulate or auriculate, sometimes with lobed margin, projecting up to 8.5 cm, 1–3 mm thick, 0.2–0.3 mm thick when dry; upper surface pilose, becoming greyish brown upon drying; hymenophore surface usually smooth, without folds, becoming fuscous to mouse-grey upon drying.
Internal features—Medulla present or absent, usually in the middle of the cross-section if present; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, apical tips acute or obtuse, tufted, 67–125(–150) × 4.5–6 µm; hyphae with clamp connections, 0.5–3.5 µm in diam. in KOH; basidia clavate, transversely 3-septate, with oil guttules, 55–71 × 4–5 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, 14–16.5(–16.8) × (4.2–)4.5–5.4 µm, L = 15.38 µm, W = 4.92 µm, Q = 2.9–3.39 (n = 60/2).
Distribution—North America and north Asia.
Notes—
Auricularia americana is one of two species that grows on gymnosperm wood. It differs from the other gymnosperm-inhabiting species,
A. tibetica Y.C. Dai & F. Wu, by smaller basidia (55–71 × 4–5 µm vs. 70–103 × 4–7 µm) and smaller basidiospores (14–16.5 × 4.5–5.4 µm vs. 15–18.5 × 5.8–6.2 µm).
Auricularia americana was originally described from Canada [
13], and recently reported from China [
5]. Samples from North America and north Asia have a few differences in molecular data and show intraspecific genetic variations in our phylogenies (
Figure 1).
Specimens examined—China. Heilongjiang Province, Dailing, Liangshui, on dead tree of Picea, 26 August 2014, B.K. Cui, Cui 11657 (BJFC 016841); Ningan County, Jingbohu Underground Forest Park, on fallen trunk of Abies, 5 September 2013, Y.C. Dai, Dai 13461 (BJFC 014922), Dai 13463 (BJFC 014924); Tangyuan County, Daliangzihe National Park, on fallen trunk of Picea, 25 August 2014, B.K. Cui, Cui 11509 (BJFC 016751); Yichun, Fenglin Nature Reserve, on fallen trunk of Picea, 2 August 2011, B.K. Cui, Cui 9887 (BJFC 010780). Inner Mongolia Autonomous Region, Hulun Buir, on fallen trunk of Larix, 12 August 2013, Y.C. Dai, Dai 13636 (BJFC 015102). Jilin Province, Antu County, Changbaishan Nature Reserve, on fallen trunk of Abies, 7 September 2013, Y.C. Dai, Dai 13476 (BJFC 014937), Dai 13480A (BJFC 014942), Dai 13488 (BJFC 014949). Russia. Amur Region, Khingansky Reserve, 10 October 1991, Mikulin (VLA M-11352); Primorye Territory, Sikhote-Alin Nature Reserve, on fallen branch of Pinus, 16 August 2012, Gromyko (LE 296428). USA. Minnesota, Bohall Trail, Itasca State Park, on Abies, 6 August 1981, Burdsall, HHB-11370 (CFMR), HHB-11374 (CFMR); Wisconsin, Treehaven, Tomahawk, on fallen trunk of Abies, 7 September 1991, Burdsall, HHB-14337 (CFMR).
(3)
Auricularia angiospermarum Y.C. Dai, F. Wu & D.W. Li, sp. nov.
Figure 5b and
Figure 7.
MycoBank number: MB 840864.
=Auricularia angiospermarum Y.C. Dai, F. Wu & D.W. Li, in Wu et al., Mycol. Prog. 14(95): 8, 2015, nom. inval., Art. F.5.6 (San Juan).
Type—USA. Connecticut, Windsor, N 41°51′, W 72°38′, on fallen trunk of Quercus, 4 December 2014, B.K. Cui, Cui 12360 (BJFC 017274, holotype).
Basidiomata—Gelatinous when fresh, fawn to vinaceous brown, solitary, sometimes caespitose, sessile or substipitate; pileus cupulate or auriculate, sometimes with lobed margin, projecting up to 8 cm, 1–3 mm thick, 0.2–0.36 mm thick when dry; upper surface pilose, sometimes with a few folds, becoming greyish brown upon drying; hymenophore surface usually smooth, without folds, becoming vinaceous grey to fuscous upon drying.
Internal features—Medulla absent; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, sometimes separated, apical tips acute or obtuse, single or tufted, 80–140 × 5–6 µm; hyphae with clamp connections, 0.5–5 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 47–63 × 3.5–5 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one to three large guttules, IKI–, CB–, (12.5–)13–15 × (4.5–)4.8–5.5(–6) µm, L = 13.72 µm, W = 4.97 µm, Q = 2.75–2.86 (n = 40/2).
Distribution—USA.
Notes—
Auricularia angiospermarum was described as a new species by Wu et al. [
23], but the name was invalid because the registration identifier cited in the protologue, ‘MycoBank no.: MB 812851′, was not issued for the name published. Therefore, the name is validated here.
The species was segregated from
A. americana. Morphologically, there is no distinct differences between the two species and they were considered as a single species by Looney et al. [
20]. However, these two species are clustered in two distinct lineages with strong support in the phylogenies (
Figure 1 and
Figure 2), and
A. angiospermarum grows on angiosperm rather than on gymnosperm wood.
Auricularia angiospermarum has so far been reported in North America only, while
A. americana occurs both in North America and north Asia. The description taken from Wu et al. [
23].
Specimens examined—USA. Arizona, Sycamore Canyon, on Juglans, 7 July 1980, Burdsall, HHB-11037 (CFMR). Illinois, Pomona Natural Bridge, Shawnee National Forest, on angiosperm wood, 29 May 1986, Nakasone, FP-102097 (CFMR). Wisconsin, Wyalusing State Park, on living tree of Carya, 31 May 1993, Volk, TJV-93-12-Sp (CFMR).
Basidiomata—Gelatinous when fresh, fawn to greyish brown or buff to cream, caespitose, resupinate to effused-reflexed; pileus free lobed, margin undulate, projecting up to 5 cm, 1–3 mm thick, 0.12–0.16 mm thick when dry; upper surface villose, distinctly and concentrically zoned with wide whitish zones and thin black bands in the central pileal surface, becoming clay pink upon drying; hymenophore surface venose with folds, becoming dark greyish blue upon drying.
Internal features—Medulla absent; crystals present, usually scattered in hymenium; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, apical tips acute or obtuse, tufted, 800–1200 × 1.5–3 µm; hyphae with clamp connections, obviously inflated with a lumen in KOH, up to 11 µm in diam; basidia clavate, transversely 3-septate, with some small oil guttules, 40–52 × 3–6 µm, sterigmata usually observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, (11–)11.2–12.3(–12.6) × (4.1–)4.5–5.2(–5.5) µm, L = 11.83 µm, W = 4.93 µm, Q = 2.4 (n = 30/1).
Distribution—Asia.
Notes—
Auricularia asiatica was recently described from Asia by Bandara et al. [
26]. It belongs to the
A. mesenterica complex and is very similar to
A. brasiliana Y.C. Dai & F. Wu by sharing resupinate to effused-reflexed, basidiomata with a free lobed pileus and similar-sized basidiospores, but the latter is distinctly and concentrically zoned with wide whitish zones and thin black bands throughout the pileal surface [
25]. Phylogenetically, the species is distant from
A. brasiliana and closely related to a new species,
A. srilankensis (
Figure 1 and
Figure 2). Differences between
A. asiatica and
A. srilankensis are discussed in the notes on
A. srilankensis.
Specimens examined—China. Hainan Province, Changjiang County, Bawangling Nature Reserve, on dead angiosperm tree, 2 July 2015, Y.C. Dai, Dai 15285 (BJFC 019396); Wuzhishan, Wuzhishan Nature Reserve, on fallen angiosperm trunk, 14 November 2015, Y.C. Dai, Dai 16149 (BJFC 020246), 15 November 2015, Y.C. Dai, Dai 16224 (BJFC 020310). Indonesia. Sumatera Barat, Padang, Limau Manis, 29 July 2009, O. Miettinen, OM 13932 (H).
≡Tremella auricula-judae Bull., Herb. Fr. (Paris) 9: Table 427, Figure 2, 1789.
=Auricularia albicans Berk., J. Linn. Soc., Bot. 13: 170, 1872.
=Auricularia auricula (L.) Underw., Mem. Torrey bot. Club12: 15, 1902.
=Auricularia sambuci Pers., Mycol. eur. (Erlanga) 1: 97, 1822.
=Auricularia auricularis (Gray) G.W. Martin, Am. Midl. Nat. 30: 81, 1943.
=Auricularia lactea (Quél.) Bigeard & H. Guill., Fl. Champ. Supér. France (Chalon-sur-Saône) 2: 491, 1913.
Basidiomata—Gelatinous when fresh, reddish brown to fuscous or white, solitary, sometimes caespitose, sessile or substipitate; pileus cupulate or auriculate, sometimes with lobed margin, projecting up to 9 cm, 1–3 mm thick, 0.3–0.48 mm thick when dry; upper surface pilose, becoming yellowish brown or pinkish buff upon drying; hymenophore surface usually smooth, without folds, becoming fuscous to black or cinnamon buff upon drying.
Internal features—Medulla present or absent, usually near hymenium if present; crystals present, usually scattered in hymenium; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, apical tips acute or obtuse, tufted, 100–150(–250) × 5–7.5 µm; hyphae with clamp connections, 1–5 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 65–85 × 4–5.5 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, (14–)15–22(–23) × (4–)5–7(–8) µm, L = 19.03 µm, W = 6.16 µm, Q = 2.81–3.33 (n = 150/5).
Distribution—Europe.
Notes—
Auricularia auricula-judae is characterized by thick basidiomata, dense and tufted hairs, and large basidia and basidiospores. The species was reported to have a wide distribution worldwide [
12]. However, the so-called “
A. auricula-judae” previously reported in the USA and Asia is confirmed as
A. americana and
A. heimuer, respectively, based on morphological characters and molecular analyses [
5,
20].
Auricularia auricula-judae can be distinguished from
A. americana and
A. heimuer by its larger basidia (55–71 × 4–5 µm in
A. americana, 40–67 × 3–6.5 µm in
A. heimuer) and basidiospores (14–16.5 × 4.5–5.4 µm in
A. americana, 11–13 × 4–5 µm in
A. heimuer). Phylogenetically, the species is distantly related to
A. americana and
A. heimuer and forms a single lineage with high support (
Figure 1 and
Figure 2).
Specimens examined—Czech Republic. Brno, on fallen trunk of Carpinus, 26 March 2014, M. Tomšovský, MT 4/2014 (duplicate, BJFC 018550). Rousinov, on dead branch of Sambucus, 11 April 2014, M. Tomšovský, MT 7/2014 (duplicate, BJFC 018561). Denmark. Nordjylland, Tranum, 16 September 1998, K. Jaederfeldt, F 6772 (S). France. Lyon, Botanical Garden, on living tree of Platanus, 26 November 2012, Y.C. Dai, Dai 13210 (BJFC 014074), on fallen trunk of Robinia, 3 November 2015, Y.C. Dai, Dai 16353 (BJFC 020440). Orliénas, on fallen trunk of Cornus, 5 October 2013, B. Rivoire, Dai 13549 (duplicate, BJFC 015011); on angiosperm wood, 11 May 2014, B. Rivoire, LYBR 5404 (duplicate, BJFC 018555). Germany. Munich, Botanical Garden, on Ulmus, November 1963, E. Albertshofer, Albertshofer (PDD 62991). Russia. Western Caucasus, Karachaevo-Cherkesia, Teberda State Biosphere Reserve, on fallen branch of Fraxinus, 22 August 2009, V. Malysheva, (LE 254030); on fallen trunk of Euonymus, 10 August 2009, V. Malysheva, (LE 254071). UK. Scotland, Fife, Kilconquhar Castle, 16 January 2014, JT 04 (MFLU); Fife, Balbuthie Wood, on Sambucus, 5 Jan 2014, JT 06 (MFLU); Humbie, Church Wood, on fallen branch of Acer, 5 October 2013, JT 14 (MFLU).
MycoBank number: MB 840810.
Type—Australia. New South Wales, Gulaga, Dromedary Mountain, S 36°18′, E 150°02′, on dead wood, 10 April 2012, H. T. Van der, HT 190 (MEL 2385783, holotype).
Etymology—Australiana (Lat.): refers to the species distributed in Australia.
Basidiomata—Gelatinous when rehydrated, fawn to reddish brown, solitary or caespitose, sessile or substipitate; pileus discoid or auriculate, margin entire, projecting up to 4 cm, 0.5–2 mm thick, 0.12–0.3 mm thick and dark grey when dry; upper surface pilose, sometimes with a few folds; hymenophore surface conspicuously porose-reticulate.
Internal features—Medulla absent; crystals absent; abhymenial hairs with a slightly swollen base or center, hyaline, thick-walled, with a wide or narrow septate lumen, apical tips acute or obtuse, single, 60–100 × 7–11 µm; hyphae with clamp connections and rare simple septa, 1–4 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 45–55 × 4.5–6 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one to two large guttules, IKI–, CB–, (10.5–)11–12.8(–13) × (4–)4.4–5(–5.4) µm, L = 11.57 µm, W = 4.8 µm, Q = 2.37–2.46 (n = 60/2).
Distribution—Australia.
Notes—
Auricularia australiana forms a distinct lineage and is closely related to
A. delicata (Mont. ex Fr.) Henn.,
A. sinodelicata Y.C. Dai & F. Wu,
A. lateralis Y.C. Dai & F. Wu and
A. conferta Y.C. Dai & F. Wu in our phylogenies (
Figure 1 and
Figure 2), but it differs from
A. delicata and
A. sinodelicata by its wider hairs (5–8 µm in diam in
A. delicata and 6–9 µm in diam in
A. sinodelicata), and from
A. lateralis by shorter hairs (95–250 µm long in
A. lateralis). The species is easily confused with
A. conferta also described from Australia, but
A. conferta has dense and thick folds on the hymenophore surface and wider hairs (45–95 × 8–15 µm in
A. conferta) usually with a wide and regular septate lumen.
Additional specimen (paratype) examined—Australia. Queensland, Cook, west of Mossman, Mt Lewis State Park, on wood, 10 April 2012, R. Vilgalys, HN 3213 (MEL 2238923).
Basidiomata—Gelatinous when fresh, greyish brown to vinaceous brown or pinkish buff, caespitose, resupinate to effused-reflexed; pileus free lobed, margin undulate, projecting up to 9 cm, 1.5–3 mm thick, 0.2–0.4 mm thick when dry; upper surface tomentose, distinctly and concentrically zoned with wide whitish zones and thin black bands throughout pileal surface, becoming clay pink upon drying; hymenophore surface venose with folds, becoming dark greyish blue to blackish blue upon drying.
Internal features—Medulla absent; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, apical tips acute or obtuse, tufted, 1000–1500 × 2–3.5 µm; hyphae rarely with clamp connections, obviously inflated with a lumen in KOH, up to 13 µm in diam; basidia clavate, transversely 3-septate, with oil guttules, 30–47 × 3–5 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, 11.5–12 × 4.5 µm, L = 11.8 µm, W = 4.5 µm, Q = 2.62 (n = 2/1).
Distribution—Brazil.
Notes—
Auricularia brasiliana was described from Brazil [
25], and is macro-morphologically similar to
A. asiatica, which, however, lacks whitish zones and thin black bands at margin of pileal surface [
26]. These two species form two distinct lineages with strong support in our phylogenies (
Figure 1 and
Figure 2). The morphological description of
Auricularia brasiliana taken from Wu et al. [
25].
Specimens examined—Brazil. Alagoas, RPPN São Pedro, 12 November 2016, R.L. M. Alvarenga, RLMA 344 (URM 93413). Bahia, Santa Teresinha, Serra da Jibóia, 24 January 2011, T. B. Gibertoni, TGB 24 (URM 83482). Ceará, Crato, Floresta Nacional do Araripe, 16 May 2012, C.R.S. Lira, CRSL 886 (URM 84563). Maranhão, Cidelândia, Reserva Extrativista do Ciriaco, Povoado do Ciriaco, 30 July 2013, L. Araujo-Neta, AN-MA 42 (URM 85567, holotype). Mato Grosso, Terra Indígena Sete de Setembro—Aldeia Apoena Meirelles, Rondolândia, 31 May 2009, Gomes-Silva et al. s/n (URM 80967). Pernambuco, Jaqueira, RPPN Frei Caneca, Mata do Quengo, 2012, R.S. Chikowski, RSC 359 (URM), RSC 487 (URM). Piauí, Caracol, Parque Nacional da Serra das Confusões, 1 January 2011, A.C. Gomes-Silva, ACGS 187 (URM 83468). Rondônia, Terra Indígena Sete de Setembro, Aldeia Lapetanha, Cacoal, 04 June 2009, Gomes-Silva et al. s/n (URM 80933).
(8)
Auricularia camposii Y.C. Dai & F. Wu, sp. nov.
Figure 13.
MycoBank number: MB 825097.
Type—Brazil. Paré, Maracanã, Ilha de Maiandeua, near the village of Algodoal, S 0°59′, W 47°58′, on Rhizophora mangle, July 1998, E.L. de Campos; A. Luz s.n. (URM 76905, holotype; BJFC018567, isotype).
Etymology—Camposii (Lat.): refers to the late Ezequias Lopes de Campos, mycologist and collector of the samples of the new species.
Basidiomata—Gelatinous when rehydrated, orange-brown to reddish brown, solitary or caespitose, sessile or substipitate; pileus discoid or auriculate, margin entire, projecting up to 5 cm, 1–3 mm thick, 0.2–0.45 mm thick when dry; upper surface densely tomentose, becoming greyish brown upon drying; hymenophore surface usually smooth, without folds, becoming bluish grey to greyish blue upon drying.
Internal features—Medulla present in the middle of the cross-section; crystals present, usually scattered throughout the cross-section; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a wide or narrow lumen, apical tips acute or obtuse, tufted, 120–250 × 6–7 µm; hyphae with clamp connections and simple septa, 0.5–4 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 60–75 × 4–5.5 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores reniform, hyaline, thin-walled, 12.5–15 × 5–6 µm.
Distribution—Brazil.
Notes—
Auricularia camposii is characterized by a densely tomentose upper surface, with an obvious medulla in the middle of the cross-section. It is similar to
A. cornea in macro-morphology, but has slightly shorter abhymenial hairs (120–250 µm vs. 180–425 µm). In the phylogeny,
A. camposii is very distantly related to
A. cornea and samples of these two species form two lineages (
Figure 1 and
Figure 2).
Auricularia camposii is relatively related to
A. nigricans (
Figure 1 and
Figure 2) which, however, has distinctly longer abhymenial hairs (300–600 × 7–9 µm vs. 120–250 × 6–7 µm). The type and paratype are good enough and ITS and nLSU sequences are available from them. Therefore, we venture to publish a description for
A. camposii even though we know it is inadvisable to do so based on a few basidiospores.
Additional specimen (paratype) examined—Brazil. Paraíba, Areia, Parque Estadual Mata do Pau Ferro, 2010, C.R.S. Lira, CL8 (URM 83464; duplicate, BJFC 018581).
MycoBank number: MB 840811.
Type—Australia. Queensland, Cairns, Crater Lakes National Park, S 17°14′, E 145°37′, 17 May 2018, Y.C. Dai, Dai 18825 (BJFC 027293, holotype).
Etymology—Conferta (Lat.): refers to the species having dense folds on the hymenophore surface.
Basidiomata—Gelatinous when fresh, fawn to reddish brown, solitary or caespitose, sessile; pileus auriculate, margin entire, projecting up to 6 cm, 1–3 mm thick, 0.3–0.5 mm thick and reddish brown to fuscous when dry; upper surface pilose, usually with a few folds; hymenophore surface conspicuously and densely porose-reticulate.
Internal features—Medulla present indistinctly near the hymenium; crystals absent; abhymenial hairs with a slightly swollen base or center, hyaline, thick-walled, usually with a wide and regular septate lumen, apical tips acute or obtuse, single, 45–95 × 8–15 µm; hyphae with clamp connections, rarely simple septa, 1–4.5 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 44–58 × 4.5–6.5 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one to two large guttules, IKI–, CB–, (10.8–)11–13(–13.5) × (4–)4.5–5.2(–5.5) µm, L = 11.91 µm, W = 4.9 µm, Q = 2.43 (n = 30/1).
Distribution—Australia.
Notes—Auricularia conferta is macro-morphologically similar to A. sinodelicata because A. sinodelicata sometimes also has dense and thick folds on the hymenophore surface. However, A. conferta has larger basidia (44–58 × 4.5–6.5 µm vs. 30–45 × 4–5.5 µm in A. sinodelicata) and mean basidiospores length (L = 11.91 µm vs. L = 10.87 µm in A. sinodelicata).
Additional specimen (paratype) examined—Australia. Queensland, Cairns, Crater Lakes National Park, 17 May 2018, Y.C. Dai, Dai 18826 (BJFC 027294).
≡Exidia cornea (Ehrenb.) Fr., Syst. mycol. (Lundae) 2(1): 222, 1822.
=Auricularia polytricha var. argentea D.Z. Zhao & Chao J. Wang, Acta Mycol. Sin., 10(2): 108, 1991.
=Auricularia reticulata L.J. Li, Acta Mycol. Sin. 4(3): 151, 1985.
=Auricularia leucochroma Kobayasi, Bull. cent. nat. Mus. Manchoukuo 4: 26, 1942.
Basidiomata—Gelatinous when fresh, fawn to reddish brown or buff to white, solitary or caespitose, sessile or substipitate; pileus discoid or auriculate, sometimes with lobed margin, projecting up to 9 cm, 1–2 mm thick, 0.2–0.48 mm thick when dry; upper surface densely pilose, becoming light vinaceous grey or cream upon drying; hymenophore surface usually smooth, without folds, becoming mouse-grey to black or buff-yellow upon drying.
Internal features—Medulla present in the middle of the cross-section or near the abhymenium; crystals present, usually scattered in the hymenium; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a wide or narrow lumen, apical tips acute or obtuse, tufted, 180–425 × 6–9 µm; hyphae with clamp connections and simple septa, 0.5–3 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 60–75 × 4–6 µm, sterigmata sometimes observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one to three large guttules, IKI–, CB–, (13–)13.8–16.5(–17.6) × (4.2–)4.5–6(–6.4) µm, L = 14.89 µm, W = 5.39 µm, Q = 2.42–3.06 (n = 270/9).
Distribution—Africa, North and South America, Asia and Europe.
Notes—
Auricularia cornea is characterized macroscopically by the variability in color of fresh basidiomata, dense hairs on the upper surface, and microscopically by the presence of an obvious medulla. It has a wide distribution, being recorded almost all over the world, and is very common in both natural and managed forests of subtropical and tropical areas. It is cultivated in China as “Maomuer” and was first reported as
A. polytricha by Patouillard and Olivier [
19], a name used in almost all Chinese reports until 2015 and in other Asian and Pacific areas [
12,
17,
42,
43,
44,
45,
46,
47,
48,
49,
50,
51]. However,
A. polytricha is a synonym of
A. nigricans, which is distributed throughout America. Zhao & Wang described
A. polytricha var.
argentea D.Z. Zhao & Chao J. Wang based on its white basidiomata [
52]. According to our study,
A. polytricha var.
argentea [basidia measuring 50–65 × 4–5.5 µm; basidiospores measuring (12.7–)13–15(–15.2) × (4.7–)4.9–5.4(–5.6) µm, L = 13.91 µm, W = 5.02 µm, Q = 2.77 (n = 30/1), from type material] is a synonym of
A. cornea. In fact, we found that
A. cornea produced both brown and white basidiomata in nature (
Figure 15h). In our phylogenies (
Figure 1 and
Figure 2), all samples of
A. cornea, wild
A. polytricha var.
argentea (Dai 14876) and cultivated
A. polytricha var.
argentea (Wu 07) form a lineage although with some variations. The white basidiomata of
A. cornea is called “Yumuer” and is now cultivated in China (
Figure 15g).
In addition,
A. reticulata L.J. Li was reported as a new species from China due to its reticulate sterile surface [
53]. However,
A. cornea sometimes has a reticulate sterile surface, too, e.g., Cui 7517 (
Figure 15f). We examined the type of
A. reticulata, and found it has an obvious medulla and the abhymenial hairs are almost the same as those of
A. cornea. We did not find basidiospores from the type, but the spores were reported as 14.5 × 5–6 µm in the original description [
53], which fit the dimensions of those of
A. cornea. For these reasons, we treat
A. reticulata as a synonym of
A. cornea. Another species,
A. leucochroma, was also described from Asia [
16], and treated as a synonym of
A. nigricans [
20]. We rejected this opinion because we examined the type material of
A. leucochroma, and treated it as a synonym of
A. cornea. Because of the variation of morphology
Figure 15a–h) and of the molecular data (
Figure 1 and
Figure 2), in the present study all these samples with variable morphology are treated as
A. cornea for the time being.
Specimens examined—Benin. On fallen angiosperm trunk, 2 July 2015, Y.C. Dai, Dai 15447 (BJFC 019552), Dai 15448 (BJFC 019553). Brazil. Acre, Sena Madureira, Floresta Nacional de São Francisco, 28 January 2016, A Meiras-Ottoni, AMO 573 (URM 93416). Maranhão, Reserva Extrativista do Ciriaco, Cidelândia, 26 July 2013, L. Araujo-Neta, (URM 85561), L. Araujo-Neta, (URM 85563); Ceará, Tianguá, Trilha do Riacho, 2012, C.R.S. Lira, Lira 663 (URM 83696); Pernambuco, Tamandaré, Reserva Biológica de Saltinho, 11 March 2016, R.L.M. Alvarenga, RLMA 310 (URM 93419). Paraíba, Areia, Parque Estadual Mata do Pau Ferro, 16 April 2012, C.R.S. Lira, CRSL 663 (URM 83696). São Paulo, São Paulo, on dead angiosperm tree, 18 June 2015, S.s.n. Visnadi, AG 6 (BJFC 020594), 7 June 2015, A.M. Gugliotta, Y. Gafforov, AG 1546 (BJFC 020595), A.M. Gugliotta, Y. Gafforov, AG 1547 (BJFC 020596); 7 August 2015, A.M. Gugliotta, Y. Gafforov, AG 1559 (BJFC 020601), A.M. Gugliotta, Y. Gafforov, AG 1560 (BJFC 020601); 16 July 2015, A.M. Gugliotta, Y. Gafforov, AG 1554 (BJFC 020600); Jardim Botânico de São Paulo, April 2017, R.L.M. Alvarenga, RLMA 467 (URM 93423), RLMA 464 (URM 93422). China. Fujian Province, Nanjing County, Huboliao Nature Reserve, on fallen angiosperm trunk, 26 October 2013, B.K. Cui, Cui 11333 (BJFC 015449), Xiamen, Botanical Garden, on rotten angiosperm wood, 27 October 2013, B.K. Cui, Cui 11346 (BJFC 015462). Guangdong Province, Shixing County, Chebaling Nature Reserve, on rotten wood of Idesia polycarpa, 17 September 2009, B.K. Cui, Cui 7517 (BJFC 006005). Guangxi Autonomous Region, Nanning, Campus of Guangxi University, on fallen angiosperm trunk, 9 December 2013, Y.C. Dai, Dai 13640 (BJFC 015103), Dai 13641 (BJFC 015104), Qingxiu Mountain, on fallen angiosperm branch, 7 June 2015, Y.C. Dai, Dai 15368 (BJFC 019479); Shangsi County, Shiwandashan Nature Reserve, on fallen angiosperm trunk, 6 June 2015, Y.C. Dai, Dai 15336 (BJFC 019447), on fallen angiosperm branch, 6 June 2015, Y.C. Dai, Dai 15338 (BJFC 019449), Dai 15346 (BJFC 019457). Guizhou Province, Suiyang County, Kuankuoshui Nature Reserve, on rotten wood of Fagus, 25 November 2014, Y.C. Dai, Dai 15016 (BJFC 018129), 26 November 2014, Y.C. Dai, Dai 15070 (BJFC 018182). Hainan Province, Changjiang County, Bawangling National Forest Park, on rotten angiosperm wood, 16 June 2014, Y.C. Dai, Dai 13679 (BJFC 017419); Danzhou County, Botanical Garden, on fallen angiosperm trunk, 19 October 2013, Y.C. Dai, Dai 13547 (BJFC 015009), Dai 13548 (BJFC 015010), 15 June 2014, Y.C. Dai, Dai 13659 (BJFC 017398); Ledong County, Jianfengling Nature Reserve, on rotten angiosperm wood, 11 May 2009, Y.C. Dai, Dai 10845 (BJFC 005087). Hebei Province, Laishui County, Yesanpo Grand Canyon, on dead Acer, 18 November 2014, Y.C. Dai, Dai 14876 (BJFC 017989). Henan Province, Neixiang County, Baotianman Nature Reserve, on fallen angiosperm trunk, 21 June 2015, Y.C. Dai, Dai 15379 (BJFC 019490). Jiangxi Province, Fuzhou, on rotten wood of Albizia, 1 July 2013, Y.C. Dai, Dai 13449 (BJFC 014910). Jiangsu Province, Taizhou, on rotten wood of Salix, 30 October 2013, Y.C. Dai, Dai 13621 (BJFC 015084). Jilin Province, Antu County, Changbaishan Nature Reserve, Cultivated, 9 August 2017, F. Wu, Wu 07 (BJFC 026013). Shanxi Province, Mei County, Taibai Mounty, Honghegu Forest Park, on fallen angiosperm trunk, 9 September 2013, B.K. Cui, Cui 11162 (BJFC 015277). Yunnan Province, Jinghong, Xishuangbanna Tropical Botanical Garden, on fallen angiosperm trunk, 21 July 2014, Y.C. Dai, Dai 13750 (BJFC 017481), Dai 13753 (BJFC 017484), Dai 13754 (BJFC 017485); Puer, Taiyanghe National Forest Park, on fallen angiosperm trunk, 8 July 2013, B.K. Cui, Cui 11026 (BJFC 015142); Yingjiang County, Tongbiguan Reserve, on fallen trunk of Prunus, 30 October 2012, Y.C. Dai, Dai 13124 (BJFC 013341); Yongde County, Daxueshan Nature Reserve, on rotten angiosperm stump, 27 August 2015, Y.C. Dai, Dai 15668 (BJFC 019772). Germany. On fallen angiosperm wood, 7 August 2015, Y. Gofforov, YG-Dr1 (duplicate, BJFC 020603). Ghana. Bunso Arboretum Garden, 8 November 2016, Y.C. Dai, Dai 17352 (BJFC 024110). Indonesia. Papua, on fallen angiosperm trunk, 2 March 2007, O. Miettinen, OM 11587 (H). Japan. Bonin Islands, 1936, Okabe, (F193009, type of A. leucochroma). Singapore. Bukit Timah Nature Reserve, on fallen angiosperm trunk, 20 July 2017, Y.C. Dai, Dai 17865 (BJFC 025397). South Africa. Durban, on fallen angiosperm trunk, 1 October 2011, Y.C. Dai, Dai 12579 (BJFC 012167), Dai 12583 (BJFC 012169), Dai 12592 (BJFC 012178); on fallen trunk of Ficus, 1 October 2011, Dai 12587 (BJFC 012173). Sri Lanka. Mitirigala Nissarana Vanaga Forest, on fallen angiosperm trunk, 4 March 2019, Y.C. Dai, Dai 19650 (BJFC 031327). Tanzania. Southern Highland Province, on fallen angiosperm trunk, 16 December 1990, TS 649 (H), TS 655 (H). USA. Puerto Rico, on fallen angiosperm trunk, 24 January 2014, O. Miettinen, OM 18106 (H). Vietnam. Ho Chi Minh City, Saigon Zoo & Botanical Garden, on dead angiosperm tree, 13 October 2017, Y.C. Dai, Dai 18315 (BJFC 025838).
≡Laschia delicata Mont. ex Fr., Linnaea 5: 533, 1830.
Epitype (designated here):—Cameroon. Nsimalen Airport, near Yaounde, on fallen angiosperm trunk, 27 July 1996, D. Mossebo, P 14 (K 43873, epitype).
Basidiomata—Gelatinous when rehydrated, dark brownish to vinaceous brown, solitary or caespitose, sessile or substipitate; pileus orbricular, sometimes with lobed margin, projecting up to 8 cm, 1.5–2.5 mm thick, 0.18–0.28 mm thick and dark reddish brown to fuscous when dry; upper surface pilose, sometimes with several folds; hymenophore surface conspicuously porose-reticulate.
Internal features—Medulla absent or indistinctly present near the hymenium; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a wide or narrow septate lumen, apical tips acute or obtuse, single, 60–100 × 5–8 µm; hyphae with clamp connections, 0.5–6 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 48–65 × 4–6 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one large guttule, IKI–, CB–, (9.5–)10–11.5(–11.8) × (4.3–)4.5–5(–5.5) µm, L = 10.83 µm, W = 4.93 µm, Q = 2.08 (n = 30/1).
Distribution—Western Africa.
Notes—
Auricularia delicata was originally described from Equatorial Guinea in West Africa and it was reported throughout tropical America, Africa, Asia, Australia and South Pacific [
12,
17].
Auricularia delicata was found to be a species complex, and two new species belonging to the complex,
A. scissa and
A. subglabra, were described because they have schizomedulla, which is distinctly different from the original description of the medulla of
A. delicata by Looney et al. [
20]. In addition, samples from Australia and Mexico form two other lineages in the phylogeny, which were treated, respectively, as
A. delicata clade I and
A. delicata clade II by Looney et al. [
20], because of the shortage of samples from the type locality of
A. delicata.
In the present study, one collection from Cameroon (K 43873) was morphologically examined and phylogenetically analyzed, and its morphology fits the description of
A. delicata. In addition, Cameroon is very close to Equatorial Guinea, so we consider the specimen (K 43873) represents the real
A. delicata, and K 43873 is treated as epitype of
A. delicata. The
A. delicata clade I and
A. delicata clade II are, respectively, treated as
A. australiana and
A. tremellosa (Fr.) Pat. in our study (
Figure 1).
Auricularia tremellosa was originally described from Mexico [
53], and more samples were morphological and phylogenetically studied. These samples form a distinct lineage (
Figure 1 and
Figure 2) and their morphology fits the description of
A. tremellosa.
Auricularia delicata is characterized by a conspicuously porose-reticulate hymenophore, short hairs and relatively small basidiospores. It is closely related to A. sinodelicata Y.C. Dai & F. Wu, but A. sinodelicata has smaller basidia (30–45 µm × 4–5.5 µm), slightly shorter hairs (30–80 µm × 6–9 µm) and a distribution in Asia.
Basidiomata—Gelatinous when rehydrated, cream to honey-yellow, solitary or caespitose, sessile or substipitate; pileus discoid or auriculate, sometimes with lobed margin, projecting up to 4.8 cm, 0.6–0.8 mm thick, 0.2–0.26 mm thick when dry; upper surface densely pilose, becoming cinnamon-buff upon drying; hymenophore surface usually with several folds, becoming honey-yellow upon drying.
Internal features—Medulla present in the middle of the cross-section or near the abhymenium; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a wide or narrow septate lumen, apical tips acute or obtuse, tufted, 280–380 × 5–6.5 µm; hyphae with clamp connections, 0.5–2.5 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 50–65 × 4–5 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores not observed in the type, but reported as (14.5–)15.6–18 × 5.5–7.5 µm by Li and Liu [
54].
Distribution—China.
Notes—Auricularia eburnea is characterized by cream to honey-yellow fresh basidiomata. According to our observations of the type, the species is macro-morphologically similar to A. cornea, but A. eburnea has obviously larger basidiospores than those in A. cornea (15.6–18 × 5.5–7.5 µm vs 13.8–16.5 × 4.5–6 µm). We failed to extract DNA, because the type material is in poor condition. For the time being we accept A. eburnea as an independent species based on its original description.
Specimen examined—China. Hainan Province, Sansha, East of Xisha Islands, on rotten angiosperm wood, July 1983, L.J. Li, B. Liu, HPNHW 1449 (BJM, holotype).
=Auricularia squamosa Pat. & Har., Bull. Soc. mycol. Fr. 9: 210, 1893.
Basidiomata—Gelatinous when rehydrated, reddish brown to vinaceous brown, solitary or caespitose, sessile; pileus discoid or auriculate, margin entire, projecting up to 12 cm, 0.6–0.9 mm thick, 0.2–0.4 mm thick when dry; upper surface densely pilose, becoming vinaceous grey upon drying; hymenophore surface usually smooth, without folds, becoming fuscous upon drying.
Internal features—Medulla present in the middle of the cross-section or near the abhymenium; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with narrow lumen, apical tips acute or obtuse, tufted, up to 1 cm long and 4–6 µm broad; hyphae with clamp connections and simple septa, 0.5–4.5 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 40–60 × 4–6 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores not observed in the studied specimens, but reported as 12–14 × 4–5 µm by Lowy [
17].
Distribution—Africa.
Notes—
Auricularia eminii is characterized by the longest hairs on the upper surface in the genus and is distributed in Africa only. Lowy indicated that
A. squamosa was a synonym of
A. eminii [
17]. In the present study, we confirm this synonym because the morphological differences between the two type specimens are too small to justify the segregation of these two species.
Specimens examined—Congo. 1893, F 19526 (type of Auricularia eminii, S), 1893, FH (type of Auricularia squamosa). Ethiopia. Bonga, 1984, D. Abate, (K 27093). Tanzania. Flora, 1966, R.F. Cain, (K 89904).
Basidiomata—Soft gelatinous when fresh, fawn to reddish brown, solitary or caespitose, sessile or substipitate; pileus discoid or auriculate, margin entire, projecting up to 6 cm, 0.35–0.5 mm thick, 0.04–0.2 mm thick and fawn to fuscous when dry; upper surface scantly pilose; hymenophore surface usually with folds.
Internal features—Medulla indistinctly present in the middle of the cross-section; crystals present, very small, scattered throughout the cross-section; abhymenial hairs with an obviously swollen base, hyaline, thick-walled, with a wide septate lumen, apical tips acute or obtuse, single, 60–100 × 10–20 µm; hyphae with clamp connections, 1–3 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 41–57 × 4–6 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, (10.7–)11–14(–14.5) × 4–5(–5.6) µm, L = 12.39 µm, W = 4.47 µm, Q = 2.45–3.03 (n = 150/5).
Distribution—Africa, Asia and Oceania.
Notes—
Auricularia fibrillifera was originally described from Papua New Guinea [
55], and the type specimen (F 234519) was studied.
Auricularia thailandica Bandara & K.D. Hyde was recently described from Thailand by Bandara et al. [
24], and it is very close to
A. fibrillifera both in morphology and phylogeny. Both species occur in Southeast Asia and it is very difficult to distinguish them by morphology, especially the dry basidiomata. However, these two species clustered into two distinct lineages in our phylogeny based on the concatenated ITS+nLSU dataset (
Figure 1); furthermore, the fresh basidiomata of
A. fibrillifera are softer than those of
A. thailandica. Therefore, we treat
A. fibrillifera and
A. thailandica as two independent species in the present study.
Specimens examined—China. Guizhou Province, Anshun County, Huangguoshu Nature Reserve, on fallen angiosperm trunk, 17 July 2014, Y.C. Dai, Dai 13744 (BJFC 017475). Hainan Province, Changjiang County, Bawangling National Forest Park, on fallen angiosperm trunk, 9 May 2009, B.K. Cui, Cui 6318 (BJFC 004174), Cui 6501 (BJFC 004354), Cui 6504 (BJFC 004357); Qiongzhong County, Diaoluoshan Forest Park, on fallen angiosperm trunk, 13 June 2014, Y.C. Dai, Dai 13596A (BJFC 017335), Dai 13597A (BJFC 017336), Dai 13598A (BJFC 017337); Wanning County, on fallen angiosperm trunk, 15 May 2009, B.K. Cui, Cui 6704 (BJFC 004558). Papua New Guinea. 28 December 1969, F 234519 (TNS, holotype). Zambia. Livingstone, on rotten angiosperm wood, 1 January 2018, Y.C. Dai, Dai 18486 (BJFC 026006).
≡Exidia fuscosuccinea Mont. [as ‘fusco-succinea’] Annls Sci. Nat., Bot., sér. 2 17: 125, 1842.
Basidiomata—Gelatinous when fresh, fawn to cinnamon brown, solitary or caespitose, sessile or substipitate; pileus discoid or auriculate, margin entire, projecting up to 6.5 cm, 1–3 mm thick, 0.15–0.3 mm thick and cinnamon brown to vinaceous brown when dry; upper surface pilose, sometimes with folds; hymenophore surface smooth, sometimes with folds.
Internal features—Medulla present near the abhymenium; crystals present, scattered in the hymenium; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a wide or narrow lumen, sometimes with a septate lumen in acute or obtuse apical tips, single or tufted, 75–165 × 5–8 µm; hyphae with clamp connections and simple septa, 1–6 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 42–66 × 4–7 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, (11.8–)12–14(–14.5) × (4–)4.3–5.2(–5.5) µm, L = 13 µm, W = 4.92 µm, Q = 2.62–2.66 (n = 60/2).
Distribution—Tropical and subtropical America.
Notes—
Auricularia fuscosuccinea is characterized by macroscopically cinnamon brown basidiomata when dry and microscopically by a medulla distinctly present near the abhymenium. It is similar to
A. fibrillifera and
A. thailandica but differs by having slightly thicker basidiomata and hyphae with simple septa. In the phylogeny,
A. fuscosuccinea is distantly related to
A. fibrillifera and
A. thailandica (
Figure 1 and
Figure 2). The species was previously reported in China [
44]. Vouchers of Chinese materials were studied, and they are actually
A. fibrillifera or
A. thailandica. Because the three species share similar morphology,
A. fuscosuccinea is temporarily considered as a species complex even if the phylogenetic relations are distant (
Figure 1 and
Figure 2).
Specimens examined—Brazil. Acre, Sena Madureira, Floresta Nacional de São Francisco, on dead angiosperm tree, 28 January 2016, A. Meiras-Ottoni, AMO 538 (URM 93429), Floresta Nacional do Macauã, 29 January 2016, A. Meiras-Ottoni, AMO 623 (URM 93435), Reserva Extrativista do Cazumbá-Iracema, 30 January 2016, A. Meiras-Ottoni, AMO 672 (URM 93430). Goiás, Floresta Nacional de Silvânia, 12 January 2016, R.L.M. Alvarenga, RLMA 395 (URM 93434); Paraíba, Areia, Parque Estadual Mata do Pau Ferro, 2013, C.R.S. Lira, CRSL 563 (URM 84766, duplicate, BJFC 018583); Pernambuco, Gravatá, RPPN Serra do Contente, on dead angiosperm tree, 18 May 2017, Y.C. Dai, Dai 17451 (BJFC 024982). São Paulo, São Paulo, on dead angiosperm tree, 7 June 2015, A.M. Gugliotta, Y. Gafforov, AG 1548 (BJFC 020597), A.M. Gugliotta, Y. Gafforov, AG 1549 (BJFC 020598), A.M. Gugliotta, Y. Gafforov, AG 1550 (BJFC020599), 2013, 12 May 2017, Y.C. Dai, Dai 17406 (BJFC 024940), Dai 17422 (BJFC 024953); Paraíba, Areia, Parque Estadual Mata do Pau Ferro, 2013, C.R.S. Lira, CRSL 563 (URM 84766, duplicate, BJFC 018583); Rondônia, Itapuã do Oeste, Floresta Nacional do Jamari, 06 February 2016, A. Meiras-Ottoni, AMO 699 (URM 93431). USA. Florida, on rotten angiosperm wood, 24 November 2013, O. Miettinen, OM 17909 (H). Louisiana, on fallen angiosperm trunk, 1990, T.J. Volk, FP-102573-SP (CFMR). Puerto Rico, on fallen angiosperm trunk, 1993, D.J. Lodge, H.H. Burdsall, Jr., & M. Boyd, PR-1378 (CFMR).
Basidiomata—Gelatinous when rehydrated, fuscous to vinaceous brown, solitary, sessile or substipitate; pileus auriculate, margin entire, projecting up to 3.5 cm, 2–3.1 mm thick, 0.6–0.7 mm thick and mouse-grey to black when dry; upper surface scantly pilose, sometimes with folds; hymenophore surface with obvious folds.
Internal features—Medulla present in the middle of the cross-section; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, sometimes septate, apical tips acute or obtuse, single, 30–80 × 4–5 µm; hyphae with clamp connections, 0.5–5 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 70–80 × 3–4 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores not observed in the type, but reported as 7.5–8.5 × 3–4 µm by Li [
18].
Distribution—China.
Notes—Auricularia hainanensis is characterized by very thick basidiomata when dried and very small basidiospores. According to the original description and our observation of the type, the species is morphologically similar to A. minor Kobayasi, but that species has thinner basidiomata (0.1–0.2 mm vs. 0.6–0.7 mm) and hymenophore surface without distinct folds. Because the type materials of both A. hainanensis and A. minor are in poor condition, and DNA extraction from the types failed, their phylogenetic relations remain uncertain.
Specimen examined—China. Hainan Province, Qiongzhong County, Limu Mountain, on fallen angiosperm trunk, 12 April 1977, L.J. Li, HPNHM 103 (BJM, holotype).
Basidiomata—Gelatinous when fresh, fawn to reddish brown, solitary or caespitose, sessile or substipitate; pileus auriculate or petaloid, margin entire, projecting up to 12 cm, 0.8–1.5 mm thick, 0.04–0.24 mm thick and greyish brown to vinaceous brown when dry; upper surface pilose, sometimes with folds; hymenophore surface smooth, sometimes with shallow folds.
Internal features—Medulla present in the middle of the cross-section; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, apical tips acute or obtuse, single or tufted, 50–150 × 4–6.5 µm; hyphae with clamp connections, 0.5–3 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 40–67 × 3–6.5 µm, sterigmata frequently observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, 11–13(–13.1) × 4–5(–5.1) µm, L = 12 µm, W = 4.71 µm, Q = 2.43–2.74 (n = 120/4).
Distribution—Temperate Asia.
Notes—Morphologically
A. heimuer is similar to
A. villosula Malysheva, but the latter has larger basidiospores (13–15.5 × 5–6.1 µm).
Auricularia heimuer has a wide distribution in temperate Asia and grows mostly on
Quercus, but basidiomata from the wild are uncommon. Previously the Asian
A. heimuer was considered the same species as the European
A. auricula-judae [
56,
57]. The cultivated and the wild
A. heimuer (
Figure 5e,f) were different from
A.
auricula-judae in both morphology (refer to the notes on
A. auricula-judae) and phylogeny, in which they clustered into two different lineages, each one with high support (
Figure 2). The morphological description of
A. heimuer taken fromWu et al. [
5].
Specimens examined—China. Heilongjiang Province, Donglin County, on fallen trunk of Quercus, 4 August 2014, Y.C. Dai, Dai 13785 (BJFC 017516), Dai 13788 (BJFC 017519); Hailin County, on fallen trunk of Quercus, 2 August 2014, Y.C. Dai, Dai 13765 (BJFC 017496, holotype); Mudanjiang, Wenchun County, on fallen trunk of Quercus, 12 August 2009, B.K. Cui, Cui 7105 (BJFC 005592); Yabuli County, Hongxing Farm, on fallen trunk of Quercus, 12 September 2013, Y.C. Dai, Dai 13503 (BJFC 014965). Jilin Province, Dunhua County, Huangnihe, cultivation, 8 August 1997, Y.C. Dai, Dai 2291 (BJFC 019121); Hunchun, Hadamen Village, on fallen trunk of Quercus, 7 August 2009, B.K. Cui, Cui 7101 (BJFC 005588); Taiyang Mountain, on fallen trunk of Quercus, 25 July 2013, Y.C. Dai, Dai 13647 (BJFC 015110). Shanxi Province, Lingchuan County, on fallen angiosperm trunk, 9 November 2009, B.K. Cui, Cui 8644 (BJFC 007133). Tianjin, Ji County, on fallen trunk of Quercus, 1 August 2009, B.K. Cui, Cui 7051 (BJFC 005538). Ningxia Autonomous Region, Liupan Mountain, on fallen angiosperm trunk, 11 July 2010, Y.C. Dai, Dai 11939 (BJFC 009040). Russia. Khankaisky District, Novo-Kachalinsk, on fallen trunk of Quercus, 26 July1996, Leley, Leley 296423 (LE). Primorsky Territory, Sikhote-Alin Nature Reserve, vicinities of Ust’-Serebryany, on fallen trunk, 4 September 1996, Morozova, (LE). Republic of Buryatia, Kabansky District, on fallen trunk of Populus, 9 August 1998, Morozova, 262806 (LE).
MycoBank number: MB 825098.
Type—China. Yunnan Province, Yongde County, Daxueshan Nature Reserve, E 99°38′, N 24°9′, on rotten angiosperm wood, 27 August 2015, Dai 15670 (BJFC 019774, holotype).
Etymology—Lateralis (Lat.): refers to the wide abhymenial hairs with slightly swollen center.
Basidiomata—Gelatinous when fresh, reddish brown to fawn, solitary or caespitose, sessile or substipitate; pileus discoid or auriculate, sometimes with lobed margin, projecting up to 5 cm, 1–3 mm thick, 0.2–0.3 mm thick and fawn to fuscous when dry; upper surface distinctly pilose, sometimes with a few folds; hymenophore surface porose-reticulate.
Internal features—Medulla present in the middle of the cross-section; crystals absent; abhymenial hairs with a slightly swollen center, hyaline, thick-walled, with a wide septate lumen, apical tips acute or obtuse, single or tufted, 95–250 × 9–14 µm; hyphae with clamp connections and simple septa, 1–6 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 50–70 × 5–6.5 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, (12.3–)12.9–14.2(–15) × (5–)5.2–6(–6.2) µm, L = 13.56 µm, W = 5.73 µm, Q = 2.37 (n = 30/1).
Distribution—China.
Notes—
Auricularia lateralis is characterized macroscopically by a conspicuously porose-reticulate hymenophore, and microscopically by wide abhymenial hairs with slightly swollen center. The species belongs to the
A. delicata complex and it is morphologically close to
A. pilosa Y.C. Dai & F. Wu sharing abhymenial hairs > 100 μm long, but
A. pilosa has shorter basidia (35–45 × 4–5.5 µm) and is distributed in Africa. Phylogenetically, the species is distantly related to
A. pilosa and forms a distinct lineage with high support (
Figure 1 and
Figure 2).
Additional specimens (paratypes) examined—China. Yunnan Province, Yongde County, Daxueshan Nature Reserve, on rotten angiosperm wood, 27 August 2015, Y.C. Dai, Dai 15669 (BJFC 019773); 5 August 2014, Y.C. Dai, Dai 16416 (BJFC 022535), Dai 16417 (BJFC 022536), Dai 16418 (BJFC 022537), Dai 16419 (BJFC 022538), Dai 16420 (BJFC 022539).
≡Helvella mesenterica Dicks., Fasc. pl. crypt. brit. (London) 1: 20, 1785.
=Auricularia tremelloides Bull., Herb. Fr. (Paris) 7: Table 290, 1787.
=Auricularia lobata Sommerf. Mag. Naturvididensk. 6: 295, 1826.
Basidiomata—Gelatinous when fresh, greyish brown to fuscous or buff to white, caespitose, resupinate to effused-reflexed; pileus free lobed, margin undulate, projecting up to 7 cm, 1.5–3 mm thick, 0.2–0.3 mm thick when dry; upper surface hispid, distinctly and concentrically zoned with canescent zones and dark bands, becoming olivaceous buff upon drying; hymenophore surface venose with obvious folds, becoming fawn to reddish brown upon drying.
Internal features—Medulla absent; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, apical tips acute or obtuse, tufted, 1000–2000 × 2–4 µm; hyphae with clamp connections, obviously inflated with a lumen in KOH, up to 14 µm in diam; basidia clavate, transversely 3-septate, with oil guttules, 48–68 × 4–6.5 µm, sterigmata rarely observed; cystidioles present, 5–9 × 2.5–5 µm.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, (13.8–)14–17(–17.6) × (4.5–)4.7–5.2(–5.3) µm, L = 15.45 µm, W = 5.01 µm, Q = 3.08–3.09 (n = 60/2).
Distribution—Europe and Uzbekistan.
Notes—
Auricularia mesenterica is a common species in Europe and grows on species of different angiosperm wood usually in summer to autumn. The species was originally described from Europe, but is also reported from the Americas and Asia [
14,
20,
43]. However,
A. mesenterica was reported as a species complex with three species (
A. asiatica,
A. brasiliana and
A. orientalis Y.C. Dai & F. Wu) [
25,
26]. In the present study, the other two new species,
A. africana and
A. submesenterica Y.C. Dai & F. Wu, are described. In addition,
A. pusio, originally described from Australia [
58], is re-studied and phylogenetically analyzed. We confirmed that it belongs to the
A. mesenterica complex and is an independent species.
Morphologically, the
A. mesenterica complex is distinct from other species in the genus by its resupinate to effused-reflexed basidiomata, the upper surface usually with distinct and concentric zones and a venose hymenophore surface with obvious folds.
Auricularia mesenterica has the largest basidiospores among the species in the complex. Phylogenetically,
A. mesenterica forms a single lineage with strong support and is closely related to
A. orientalis and
A. submesenterica (
Figure 1 and
Figure 2).
Specimens examined—Czech Republic. Bukovinka, Rakovecký les forest, on fallen angiosperm trunk, 15 September 2007, A. Vágner, (BRNM 706955, duplicate, BJFC 018552); Vranovice upon Svratka, Plačkův les Nature Reserve, on the trunk of Fraxinus, 19 November 1999, A. Vágner, (BRNM 648573, duplicate, BJFC 018554). Estonia. Viljandi District, on fallen angiosperm wood, 26 August 1989, I. Kytovuori, Kytovuori 89-333 (H). France. Péage de Roussillon, on the trunk of Populus, 15 April 2014, B. Rivoire, LYBR 5353 (BJFC 018549). Italy. Parma, La Cittadella Park, on rotten angiosperm stump, 27 February 1987, V. Härkönen, Härkönen 8045 (H). Sweden. Närke, Almby, Hjälmarsberg, on rotten wood of Ulmus, 28 September 1998, Nilsson, F 12670 (S). Switzerland. Sparse, on fallen trunk of Quercus, 1 June 2008, O. Miettinen, Miettinen 12680 (H). UK. Wales, Gwent, Abergavenny Riverside, on rotten angiosperm trunk, 5 March 1990, V. Härkönen, Härkönen 11208 (H). Uzbekistan. Profenicel garden, Taslvencf, on angiosperm trunk, 2 September 2011, Y. Gofforov, YG 029 (BJFC 020591); Xumson village, Suglens forest, on fallen trunk of Juglans, 26 September 2011, Y. Gofforov, YG 037 (BJFC 020592).
Basidiomata—Gelatinous when rehydrated, greyish brown to fawn, solitary, sessile; pileus discoid or auriculate, margin entire, projecting up to 6.5 cm, 0.65–0.8 mm thick, 0.1–0.2 mm thick when dry; upper surface scantly pilose, becoming mouse-grey upon drying; hymenophore surface smooth, sometimes with indistinct folds, becoming black upon drying.
Internal features—Medulla present in the middle of the cross-section; crystals absent; abhymenial hairs with a slightly swollen base, hyaline or clay buff, thick-walled, with a narrow lumen, sometimes septate, apical tips acute or obtuse, single or tufted, 90–120 × 6–10 µm; hyphae with clamp connections, 1–2 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 35–43 × 2–4 µm, sterigmata rarely observed; cystidioles absent.
Spores—Only one possible true basidiospore was observed in the type, and was reported as measuring 7–8 × 3–4 µm by Kobayasi [
12].
Distribution—Asia.
Notes—
Auricularia minor was originally described from tropical Asia [
12], and is characterized by its solitary and very small basidiomata. We failed to extract DNA because the type material and other samples are in poor condition, so its phylogenetic relations with other species are uncertain. Morphologically,
A. minor is very similar to
A. hainanensis described from tropical Asia, but the abhymenial hairs in
A. minor (90–120 × 6–10 µm) are longer than those in
A. hainanensis (30–80 × 4–5 µm).
Specimens examined—New Guinea. 1981, F-434 (TNS, type). Japan. 4 October 1981, F-51392 (TNS).
Basidiomata—Gelatinous when fresh, vinaceous grey to fuscous or clay buff to pinkish buff, solitary or caespitose, sessile; pileus discoid or auriculate, sometimes with lobed margin, projecting up to 4 cm, 0.5–2 mm thick, 0.04–0.2 mm thick when dry; upper surface pilose, sometimes with folds, becoming vinaceous grey upon drying; hymenophore surface smooth or with obvious folds, becoming fuscous or black upon drying.
Internal features—Medulla present in the middle of the cross-section; crystals present, usually scattered in the hymenium; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, sometimes septate, apical tips acute or obtuse, single or tufted, 50–85(–105) × 5–6 µm; hyphae with clamp connections, 1–3.5 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 46–67 × 5–8 µm, sterigmata occasionally observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, (11.8–)13–15(–16.2) × 5–6(–7) µm, L = 14.28 µm, W = 5.83 µm, Q = 2.44–2.46 (n = 80/3).
Distribution—China and Russia.
Notes—
Auricularia minutissima belongs to the
A. auricula-judae complex and is characterized by its small basidiomata. The species is similar to
A. heimuer and
A. minor, but the latter two species have smaller basidiospores (11–13 × 4–5 µm in
A. heimuer and 7–8 × 3–4 µm in
A. minor).
A. minutissima has two kinds of basidiomata (
Figure 5h,i) that are different in macro-morphology, but have the similar basidia, basidiospores and DNA data. In the phylogeny,
A. minutissima is closely related to
A. heimuer, but it forms a single lineage with strong support (
Figure 1 and
Figure 2). The morphological description of
A. minutissima taken from Wu et al. [
23].
Specimens examined—China. Gansu Province, Pingliang, Kongtongshan, on rotten branch of Acer, 3 August 2015, Y.C. Dai, Dai 15452 (BJFC 019557), Dai 15453 (BJFC 019558), Dai 15454 (BJFC 019559), Dai 15455 (BJFC 019560), Dai 15456 (BJFC 019561). Hebei Province, Laishui County, Yesanpo Nature Reserve, on fallen branch of Quercus, 18 November 2014, Y.C. Dai, Dai 14880 (BJFC 017993), Dai 14881 (BJFC 017994, holotype). Russia. Primorye Territory, Kedrovaya Pad’Reserve, 10 October 1994, Govorova, (LE 296424).
(22) Auricularia nigricans (Sw.) Birkebak, Looney & Sánchez-García
≡Peziza nigricans Sw., Fl. Ind. Occid.3: 1938, 1806.
=Exidia polytricha Mont., Voy. Indes Or., Bot. 2: 154, 1834.
Basidiomata—Gelatinous when rehydrated, reddish brown to orange-brown, solitary or caespitose, sessile or substipitate; pileus cupulate or auriculate, sometimes with lobed margin, projecting up to 6 cm, 1–3 mm thick, 0.36–0.4 mm thick when dry; upper surface usually densely hispid, becoming vinaceous grey upon drying; hymenophore surface usually smooth, without folds, becoming dark-grey upon drying.
Internal features—Medulla obviously present near the abhymenium; crystals present, usually scattered in the hymenium; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, apical tips acute or obtuse, tufted, 300–600 × 7–9 µm; hyphae with clamp connections and simple septa, 0.5–4 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 50–60 × 4–6 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores not observed in the studied samples, but reported as measuring 14.5–17 × 5–7 µm by Looney et al. [
20].
Distribution—North America.
Notes—
Auricularia polytricha is a name that has been applied to this species for a long time [
12,
17,
51], but Looney et al. recently found that
Exidia polytricha Mont. and
Peziza nigricans represent the same species [
20]. Since
P. nigricans was an earlier name and thus had the priority, they proposed the combination
A. nigricans (Sw.) Birkebak & al., and
A. polytricha is treated as a synonym of
A. nigricans.
Auricularia nigricans is characterized by the densely hispid upper surface and obvious medulla near the abhymenium. It is easily confused with
A. cornea because of the variable morphology of
A. cornea, but
A. nigricans usually has a more hispid upper surface and is distributed in North America only. Phylogenetically, both species are distantly related (
Figure 1 and
Figure 2).
Specimens examined—Costa Rica. On fallen angiosperm trunk, 4 January 1979, Ahti 36234 (H). Mexico. On fallen angiosperm trunk, 18 October 1997, T. Ahti, Ahti 55718 (H). USA. Louisiana, on fallen trunk of Quercus, 1993, T.J. Volk, TJV-93-242 (CFMR).
Mycobank number: MB 825099.
Type—New Zealand. Auckland, S 36°50′, E 174°44′, on stump, 12 May 2005, B. Dee, (PDD 83897, holotype).
Etymology—Novozealandica (Lat.): refers to the species from New Zealand.
Basidiomata—Gelatinous when rehydrated, reddish brown to fuscous, solitary or caespitose, sessile or substipitate; pileus cupulate or auriculate, sometimes with lobed margin, projecting up to 15 cm, 0.8–1.5 mm thick, 0.3–0.4 mm thick when dry; upper surface densely pilose, becoming greyish brown to clay buff upon drying; hymenophore surface usually smooth, without folds, becoming vinaceous grey upon drying.
Internal features—Medulla obviously present in the middle of the cross-section or near the abhymenium; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a wide or narrow lumen, apical tips with several septate lumens, obtuse, tufted, 100–320 × 7–9 µm; hyphae with clamp connections and simple septa, 0.5–3 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 70–86 × 5–6 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, (15–)16–19(–19.5) × (5.1–)5.3–6.1(–6.5) µm, L = 17.47 µm, W = 5.83 µm, Q = 2.94–3.05 (n = 90/3).
Distribution—New Zealand.
Notes—Auricularia novozealandica belongs to the A. cornea complex and is characterized by dense hairs on the upper surface, obvious medulla in the middle of the cross-section or near the abhymenium and very big basidiospores. It is morphologically and phylogenetically related to A. cornea, but A. cornea has smaller basidia (60–75 × 4–6 µm) and smaller basidiospores (13.8–16.5 × 4.5–6 µm).
Additional specimens (paratypes) examined—New Zealand. Auckland, 29 January 1998, R. Leschen, (PDD 70405); 14 March 2001, S.R. Whitton, C. Delleey, R. Fuller, (PDD 75110); 29 May 1999, C. Shirley, (PDD 81195); on dead wood, 8 May 2006, P. Catcheside, D. Catcheside, (PDD 88998). Nelson, 11 May 2004, D. Catcheside, (PDD 94628). Wellington, 25 March 1997, A. Freeston, R. Freeston, (PDD 94817).
Basidiomata—Gelatinous when fresh, greyish brown to fuscous, solitary or caespitose, resupinate to effused-reflexed; pileus free lobed, margin undulate, projecting up to 2 cm, 2–3 mm thick, 0.2–0.3 mm thick when dry; upper surface densely villose, sometimes concentrically zoned with canescent zones and dark bands, becoming white to olivaceous buff upon drying; hymenophore surface venose with folds, becoming greyish blue to dark greyish blue upon drying.
Internal features—Medulla absent; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, apical tips acute or obtuse, tufted, 900–1500(–1800) × 1.5–2.5 µm; hyphae with clamp connections, obviously inflated with a lumen in KOH, up to 7 µm in diam; basidia clavate, transversely 3-septate, with oil guttules, 50–72 × 3.5–6 µm, sterigmata rarely observed; cystidioles present, 6–8 × 3–3.5 µm.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one to three large guttules, IKI–, CB–, (12–)12.5–14.2(–15) × 5–6 µm, L = 13.44 µm, W = 5.48 µm, Q = 2.45 (n = 30/1).
Distribution—China.
Notes—
Auricularia orientalis was described as a new species in the
A. mesenterica complex [
25]. Compared to other species in this complex, only
A. orientalis and
A. mesenterica have cystidioles, but
A. orientalis has smaller basidiospores than those of
A. mesenterica (12.5–14.2 × 5–6 µm vs. 14–17 × 4.7–5.2 µm). Phylogenetically, samples of
A. orientalis clustered in a single lineage with robust support (
Figure 1 and
Figure 2). The morphological description of
A. orientalis taken from Wu et al. [
25].
Specimens examined—China. Beijing, Baihuashan Nature Reserve, on fallen angiosperm trunk, 2 November 1993, Y.C. Dai, Dai 1831 (BJFC 019131). Hebei Province, Laishui County, Yesanpo National Park, on rotten stump of Juglans, 17 November 2014, Y.C. Dai, Dai 14875 (BJFC 017988, holotype). Inner Mongolia Autonomous Region, Daqinggou Nature Reserve, on rotten angiosperm wood, 25 August 2015, Y.C. Dai, Dai 15813 (BJFC 019916).
Basidiomata—Gelatinous when rehydrated, reddish brown to cinnamon, caespitose, sessile; pileus discoid, margin entire, projecting up to 2 cm, 0.65–1 mm thick, 0.02–0.04 mm thick and yellowish brown when dry; upper surface scantly pilose, distinctly and concentrically zoned; hymenophore surface smooth, sometimes with folds.
Internal features—Medulla absent; crystals absent; abhymenial hairs with a slightly swollen base, hyaline or clay buff, thick-walled, with a narrow lumen, apical tips acute, single, 20 × 7 µm; hyphae with clamp connections, 2.8–4 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 60–70 × 8–10 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one to three small guttules, IKI–, CB–, 16–18(–18.2) × (4–)4.2–5.3(–5.5) µm, L = 17.17 µm, W = 4.86 µm, Q = 3.53 (n = 30/1).
Distribution—Japan.
Notes—Auricularia papyracea is characterized by the extremely thin and papery basidiomata when dry. It is easy to distinguish from other species in the genus by the concentric white zones on the upper surface. No fresh samples were available, and DNA extraction from the type and another sample failed. Its relation within the genus are uncertain.
Specimens examined—Japan. 17 December 1911, F-234520 (TNS, type); 7 December 1917, F-203284 (TNS).
(26)
Auricularia pilosa Y.C. Dai, L.W. Zhou & F. Wu, sp. nov.
Figure 33.
MycoBank number: MB 840812.
Type—Ethiopia. Jimma, Shebe, N 7°34′, E 36°25′, on angiosperm trunk, 21 April 2019, L.W. Zhou, LWZ20190421-7 (HMAS, holotype).
Etymology—Pilosa (Lat.): refers to the distinctly pilose abhymenial surface.
Basidiomata—Gelatinous when fresh, reddish brown to vinaceous brown, solitary or caespitose, sessile or substipitate; pileus discoid or auriculate, sometimes with lobed margin, projecting up to 4 cm, 1–2 mm thick, 0.1–0.2 mm thick and reddish brown to fuscous when dry; upper surface distinctly pilose; hymenophore surface porose-reticulate.
Internal features—Medulla absent; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a wide septate lumen, apical tips acute or obtuse, single or tufted, 90–207 × 8–16 µm; hyphae with clamp connections and simple septa, 1–5 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 35–45 × 4–5.5 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, (10.5–)11–13.8(–14.8) × (3.8–)4.2–5.8(–6.1) µm, L = 12.46 µm, W = 4.95 µm, Q = 2.52 (n = 60/1).
Distribution—Ethiopia and Tanzania.
Notes—
Auricularia pilosa is characterized by the pilose upper surface, porose-reticulate hymenophore, and wide abhymenial hairs with slightly swollen center. The species belongs to the
A. delicata complex and is easily confused with
A. lateralis because of the distinctly pilose abhymenial surface, but it differs by the slightly shorter hairs and basidia (in
A. lateralis 95–250 µm long and 50–70 µm long, respectively) and is distributed in Africa. Phylogenetically,
A. pilosa forms a single lineage distant from other species in the
A. delicata complex (
Figure 1 and
Figure 2).
Basidiomata—Gelatinous when rehydrated, fawn to greyish brown, caespitose, resupinate to effused-reflexed; pileus free lobed, margin undulate, projecting up to 3 cm, 1.5–2 mm thick, 0.2–0.4 mm thick when dry; upper surface tomentose, sometimes concentrically zoned with canescent zones and dark bands, becoming clay pink upon drying; hymenophore surface venose with folds, becoming dark greyish blue upon drying.
Internal features—Medulla absent; crystals present, usually scattered in the hymenium; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, apical tips acute or obtuse, tufted, 400–800 × 1.5–2 µm; hyphae with clamp connections and simple septa, slightly inflated with a lumen in KOH, up to 6 µm in diam; basidia clavate, transversely 3-septate, with oil guttules, 50–72 × 5–7.5 µm, sterigmata frequently observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, 12–14(–14.2) × (4.8–)5–6 µm, L = 12.97 µm, W = 5.43 µm, Q = 2.39 (n = 30/1).
Distribution—Australia and Zambia.
Notes—
Auricularia pusio was originally described from Queensland (Australia) and was considered a synonym of
A. mesenterica in the light of morphological characters (
http://www.indexfungorum.org/Names/NamesRecord.asp?RecordID=156648, accessed on 3 September 2021), but
A. mesenterica is a species complex [
25]. Basidiospores of
A. pusio are distinctly shorter and wider than those in
A. mesenterica sensu stricto (14–17 × 4.7–5.2 µm). Additionally, our phylogenies show that the two species form two distinct and distantly related lineages (
Figure 1 and
Figure 2). Therefore, we accept
A. pusio as an independent species in the
A. mesenterica complex.
Specimens examined—Australia. Western Australia, Kimberley District, Kaugaroo Pool, on dead log, 30 August 1999, B.M. Spooner, AK 547 (K 26101); Napier Range, Secret Valley, on dead standing shrub, 13 April 1988, B.M. Spooner, AK 174 (K 26100). Zambia. Mpika, North Luangwa, on 20 September 1994, D.S. Smith, Smith 18 (K 28316).
(28)
Auricularia scissa Looney, Birkebak & Matheny
Figure 35.
Basidiomata—Gelatinous when rehydrated, greyish brown to reddish brown, solitary or caespitose, sessile or substipitate; pileus discoid or auriculate, with lobed margin, projecting up to 7 cm, 1–3 mm thick, 0.1–0.2 mm thick and yellowish brown when dry; upper surface pilose; hymenophore surface conspicuously porose-reticulate.
Internal features—Schizomedulla present in the middle of the cross-section; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, apical tips acute or obtuse, single or tufted, 40–100 × 5–10 µm; hyphae with clamp connections, 0.5–5 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 40–60 × 4–5 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one to a few large guttules, IKI–, CB–, (8–)9–12 × 4–6(–6.5) µm, L = 10.73 µm, W = 5.43 µm, Q = 1.97 (n = 30/1).
Distribution—Dominican Republic and USA.
Notes—
Auricularia scissa was recently described from the Dominican Republic by Looney et al. [
20], and is characterized macroscopically by conspicuously porose-reticulate hymenophores and microscopically by the presence of schizomedulla. It is like
A. subglabra as both have porose-reticulate hymenophores and a schizomedulla, but the latter species has an almost smooth abhymenium and very short hairs (up to 45 µm). Phylogenetically,
A. scissa forms a single lineage with high support (
Figure 1 and
Figure 2) and is not closely related to
A. subglabra.
Specimens examined—Dominican Republic. 31 December 1990, T. Ahti, Ahti 49388 (H); 1998, O.P. Perdomo, DR 777 (CFMR).
MycoBank number: MB 825100.
Type—China. Chongqing, Jiangjin District, Simianshan Nature Reserve, E 104°4′, N 28°6′, on fallen angiosperm trunk, 22 July 2014, Y.C. Dai, Dai 13926 (BJFC 017656, holotype).
Etymology—Sinodelicata (Lat.): refers to the species being like A. delicata and with the distribution in China.
Basidiomata—Gelatinous when fresh, fawn to reddish brown or cinnamon to yellowish brown, solitary or caespitose, sessile or substipitate; pileus discoid or auriculate, sometimes with lobed margin, projecting up to 8 cm, 1–2.5 mm thick, 0.16–0.25 mm thick and reddish brown to black when dry; upper surface scantly pilose, sometimes with a few folds; hymenophore surface conspicuously porose-reticulate.
Internal features—Medulla indistinctly present near the hymenium; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a wide or narrow septate lumen, apical tips acute or obtuse, single, 30–80 × 6–9 µm; hyphae with clamp connections, 1–5 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 30–45 × 4–5.5 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one to three large guttules, IKI–, CB–, (9.8–)10–12(–12.2) × (4–)4.3–5.1(–5.5) µm, L = 10.87 µm, W = 4.84 µm, Q = 2.2–2.27 (n = 120/4).
Distribution—China.
Notes—Previously, Chinese samples of
A. sinodelicata were identified as
A. delicata because of the porose-reticulate hymenophore [
42,
44]). However,
A. delicata was originally described from West Africa [
12,
17], and phylogenetically
A. sinodelicata and
A. delicata cluster in two different lineages (
Figure 1). Besides,
A. delicata has longer basidia than those of
A. sinodelicata (48–65 × 4–6 µm vs. 30–45 × 4–5.5 µm).
Auricularia lateralis is described as a new species in the present study and also has a porose-reticulate hymenophore.
Auricularia lateralis and
A. sinodelicata have an overlapping distribution in southern China, but the basidiospores are distinctly larger in
A. lateralis (12.9–14.2 × 5.2–6 µm) than in
A. sinodelicata (10–12 × 4.3–5.1 µm).
Auricularia sinodelicata has variable macro-morphological characters (
Figure 10e–j) and is extremely similar to
A. tremellosa in some specimens (e.g., Cui 12236,
Figure 10e), but
A. sinodelicata usually has more or less radial folds on the hymenophore, while
A. tremellosa usually has poroid hymenophore. More importantly,
A. sinodelicata and
A. tremellosa are distantly related in phylogenies (
Figure 1 and
Figure 2), and they are distributed in Asia and the Americas, respectively.
Additional specimens (paratypes) examined—China. Chongqing, Jiangjin District, Simianshan Nature Reserve, on fallen angiosperm trunk, 22 July 2014, Y.C. Dai, Dai 13927 (BJFC 017657). Guizhou Province, Guiyang, Qianlingshan Park, on rotten angiosperm wood, 17 July 2014, Y.C. Dai, Dai 13919 (BJFC 017649), Dai 13920 (BJFC017650). Hainan Province, Lingshui County, Diaoluoshan Forest Park, on fallen angiosperm trunk, 13 June 2014, Y.C. Dai, Dai 13593A (BJFC 017332). Xizang Autonomous Region (Tibet), Motuo County, on fallen angiosperm trunk, 20 September 2014, B.K. Cui, Cui 12236 (BJFC 017150); 9 September 2014, Y.C. Dai, Dai 15083 (BJFC 018195). Yunnan Province, Jinghong, Wangtianshu Nature Reserve, on fallen angiosperm trunk, 2 November 2009, B.K. Cui, Cui 8594 (BJFC 007083), Cui 8596 (BJFC 007085), 19 July 2014, Y.C. Dai, Dai 13746 (BJFC 017477), Dai 13748 (BJFC 017479); Puer, Taiyanghe Forest Park, on fallen angiosperm trunk, 6 June 2011, Y.C. Dai, Dai 12242 (BJFC 010525), 24 July 2014, Y.C. Dai, Dai 13758 (BJFC 017489); Tengchong County, Yinghua Valley, on fallen angiosperm trunk, 28 October 2009, B.K. Cui, Cui 8315 (BJFC 006804), Cui 8316 (BJFC 006805);Yingjiang County, Tongbiguan Nature Reserve, on rotten angiosperm wood, 29 October 2010, Y.C. Dai, Dai 13112 (BJFC 013332).
Mycobank number: MB 840813.
Type—Sri Lanka. Wadduwa, South Bolgoda Lake, N 6°46′, E 79°54′, on dead angiosperm tree, 28 February 2019, Y.C. Dai, Dai 19522 (BJFC031201, holotype).
Etymology—Srilankensis (Lat.): refers to the species distribution in Sri Lanka.
Basidiomata—Gelatinous when fresh, greyish blue to vinaceous grey, caespitose, resupinate to effused-reflexed; pileus free lobed, margin undulate, projecting up to 1 cm, 1–2 mm thick, 0.08–0.16 mm thick when dry; upper surface villose, sometimes concentrically zoned with canescent zones and dark bands, becoming white to greyish violet upon drying; hymenophore surface venose with folds, becoming dark greyish blue upon drying.
Internal features—Medulla absent; crystals present; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, apical tips acute or obtuse, tufted, 400–800 × 1.5–3 µm; hyphae with clamp connections, obviously inflated with a lumen in KOH, up to 7 µm in diam; basidia clavate, transversely 3-septate, with some small oil guttules, 50–70 × 5–7 µm, sterigmata usually observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, (11–)12–14.7(–15) × (4.2–)4.5–6 µm, L = 13.18 µm, W = 5.26 µm, Q = 2.31–2.82 (n = 90/3).
Distribution—Sri Lanka.
Notes—
Auricularia srilankensis is phylogenetically closely related to
A. asiatica, but it forms one single lineage with strong support (
Figure 1 and
Figure 2) and differs from
A. asiatica by larger basidia (40–52 × 3–6 µm in
A. asiatica) and slightly longer basidiospores (11.2–12.3 µm long in
A. asiatica). Morphologically, the species is easily confused with
A. orientalis from China, but
A. orientalis has cystidioles and longer hairs (900–1500 µm).
Specimens examined—Sri Lanka. Wadduwa, South Bolgoda Lake, on dead angiosperm tree, 28 February 2019, Y.C. Dai, Dai 19519 (BJFC031198), Dai 19520 (BJFC031199); Kandy, Udawatta Kele Royal Forest Park, on fallen angiosperm trunk, 2 March 2019, Y.C. Dai, Dai 19575 (BJFC031254).
Basidiomata—Gelatinous when fresh, greyish violet to fawn, solitary or caespitose, sessile or substipitate; pileus discoid or auriculate, margin entire, projecting up to 6.5 cm, 1–2 mm thick, 0.24–0.4 mm thick and cinnamon buff to vinaceous grey when dry; upper surface scantly pilose, sometimes with a few folds; hymenophore surface slightly porose-reticulate.
Internal features—Schizomedulla present in the middle of the cross-section; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a wide or narrow septate lumen, apical tips acute or obtuse, single, 20–45 × 5–6 µm; hyphae with clamp connections and simple septa, 1–5 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 30–45 × 3.5–4.5 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, (8.5–)9–10.8(–11) × (3–)3.5–4.5 µm, L = 9.72 µm, W = 3.79 µm, Q = 2.56 (n = 30/1).
Distribution—Tropical North and South America.
Notes—
Auricularia subglabra was described from Costa Rica by Looney et al. [
20], and is characterized by slightly porose-reticulate hymenophores and the presence of schizomedulla. The species is similar to
A. scissa, which, however, has a pilose upper surface and longer abhymenial hairs (40–100 µm). Phylogenetically, both species nest in two distinct lineages with high support (
Figure 1 and
Figure 2).
Specimens examined—Brazil. Amazonas, Manaus, on rotten angiosperm wood, 12 May 2017, Y.C. Dai, Dai 17394 (BJFC 026424), Dai 17403 (BJFC 026425), Dai 17417 (BJFC 026427), F. Wu, Wu 08 (BJFC 026014), Wu 09 (BJFC 026014). French Guiana. Coralie, on rotten angiosperm trunk, 29 August 2018, J. Vlasák, JV 1808/125 (JV).
MycoBank number: MB 825101.
Type—China. Jilin Province, Antu County, Changbaishan Nature Reserve, E 127°45′, N 41°38′, on fallen angiosperm trunk, 16 July 2015, Dai 15450 (BJFC 019555, holotype).
Etymology—Submesenterica (Lat.): refers to the species being similar to Auricularia mesenterica.
Basidiomata—Gelatinous when fresh, greyish brown to fuscous, caespitose, resupinate to effused-reflexed; pileus free lobed, margin undulate, projecting up to 7 cm, 1–2 mm thick, 0.12–0.2 mm thick when dry; upper surface hispid, distinctly and concentrically zoned with canescent zones and dark bands, becoming greyish brown to deep olive upon drying; hymenophore surface venose with obvious folds, becoming dark bluish grey to dark grey upon drying.
Internal features—Medulla absent; crystals present, usually scattered in hymenium; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, apical tips acute or obtuse, tufted, 600–1000 × 2–2.5 µm; hyphae with clamp connections, obviously inflated in KOH with a wide lumen, up to 7.5 µm in diam; basidia clavate, transversely 3-septate, with oil guttules, 37–50 × 4–6.5 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, 12.5–14.8 × 5.5–6 µm, L = 13.75 µm, W = 5.72 µm, Q = 2.4 (n = 4/1).
Distribution—China.
Notes—
Auricularia submesenterica is characterized by caespitose, resupinate to effused-reflexed basidiomata with a lobed pileus, distinctly and concentrically zoned with canescent zones and dark bands on the upper surface, absence of medulla and cystidioles, and small basidia and basidiospores.
Auricularia submesenterica definitely belongs to the
A. mesenterica complex, but
A. mesenterica sensu stricto differs from
A. submesenterica by longer abhymenial hairs (1000–2000 µm), presence of cystidioles and larger basidiospores (14–17 × 4.7–5.2 µm).
Auricularia orientalis is another member in the
A. mesenterica complex and was recently described from China [
25]; it resembles
A. submesenterica by sharing similar abhymenial hairs and basidiospores, but
A. orientalis usually has resupinate basidiomata and cystidioles in the hymenium. In addition,
A. submesenterica,
A. orientalis and
A. mesenterica form three different lineages in our phylogenies (
Figure 1 and
Figure 2).
Additional specimens (paratypes) examined—China. Jilin Province, Antu County, Huangsongpu, on fallen angiosperm trunk, 28 September 1993, Y.C. Dai, Dai 792 (BJFC 019132); Changbaishan Nature Reserve, on fallen angiosperm trunk, 16 July 2015, Y.C. Dai, Dai 15449 (BJFC 019554), Dai 15451 (BJFC 019556); on fallen trunk of Quercus, 11 September 2014, Y.C. Dai, Dai 14773 (BJFC 017885). Liaoning Province, Qingyuan County, Qingyuan Ecological Station, on fallen angiosperm trunk, 26 August 2015, Y.C. Dai, Dai 15820 (BJFC 019923).
Basidiomata—Gelatinous when fresh, vinaceous to clay pink, solitary or caespitose, sessile or substipitate; pileus more or less circular, margin entire, projecting up to 6 cm, 0.4–0.7 mm thick when fresh, 0.04–0.24 mm thick and fawn to fuscous when dry; upper surface scantly pilose; hymenophore surface usually smooth, without folds, and usually with obvious white spots.
Internal features—Medulla present near the abhymenium; crystals present, very small, scattered throughout the cross-section; abhymenial hairs with a distinctly swollen base, irregular, hyaline, thick-walled with a wide septate lumen, apically acute or obtuse, single, 50–80(–100) × 6–14 µm; hyphae with clamp connections, 1–3 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 40–60 × 3–6 µm, sterigmata sometimes observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, (9–)9.5–13(–14) × 4–5.6(–6) µm, L = 11.03 µm, W = 4.69 µm, Q = 1.83–2.68 (n = 210/7).
Distribution—China and Thailand.
Notes—
Auricularia thailandica is characterized by very thin and fuscous basidiomata when dry. It is closely related to
A. fibrillifera in the phylogenies (
Figure 1 and
Figure 2). For the differences between these two species, refer to the notes on
A. fibrillifera.Specimens examined—China. Guangxi Autonomous Region, Jinxiu County, Shengtang Mountain, on rotten angiosperm wood, 17 June 2014, Y.C. Dai, Dai 15080 (BJFC 018192), Dai 15081 (BJFC 018193). Hainan Province, Baoting County, on fallen angiosperm trunk, 1 July 2010, Y.L. Wei, Wei 5513 (BJFC 009236); Changjiang County, Bawangling National Forest Park, on fallen angiosperm trunk, 16 June 2014, Y.C. Dai, Dai 13687 (BJFC 017427); Danzhou County, Danzhou Tropical Botanical Garden, on fallen angiosperm trunk, 15 June 2014, Y.C. Dai, Dai 13661 (BJFC 017401); Ledong County, Jianfengling Nature Reserve, on fallen angiosperm trunk, 17 June 2014, Y.C. Dai, Dai 13706 (BJFC 017443); Qiongzhong County, Limu Mountain, on fallen angiosperm trunk, 15 June 2014, Y.C. Dai, Dai 13629A (BJFC 017368), Dai 13635A (BJFC 017374), Dai 13654A (BJFC 017393), Dai 13655A (BJFC 017394); Tunchang County, on fallen angiosperm trunk, 23 November 2010, Y.C. Dai, Dai 11973 (BJFC 009265). Yunnan Province, Cangyuan County, on fallen angiosperm trunk, 11 July 2013, B.K. Cui, Cui 11062 (BJFC 015178); Lancang County, on fallen angiosperm trunk, 9 July 2013, B.K. Cui, Cui 11031 (BJFC 015147); Menhai County, Mangao Nature Reserve, on fallen trunk of Castanea, 8 June 2011, Y.C. Dai, Dai 12332 (BJFC 010612); Puer County, Taiyanghe National Forest Park, on fallen angiosperm trunk, 24 July 2014, Y.C. Dai, Dai 13759 (BJFC 017490).
Basidiomata—Gelatinous when fresh, orange-brown to reddish brown, usually solitary, sometimes caespitose, sessile or substipitate; pileus discoid or auriculate, usually with lobed margin, projecting up to 8.5 cm, 1–5 mm thick, 0.4–0.7 mm thick when dry; upper surface pilose, usually without folds, becoming mouse grey to greyish brown upon drying; hymenophore surface smooth, sometimes with several folds, becoming almost black upon drying.
Internal features—Medulla present in the middle of the cross-section; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, sometimes septate, apical tips acute or obtuse, single or tufted, 75–135(–150) × 5–8 µm; hyphae with clamp connections, 0.5–4.5 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 70–103 × 4–7 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one to three large guttules, IKI–, CB–, (14–)15–18.5(–20) × 5.8–6.2(–6.5) µm, L = 16.22 µm, W = 6.0 µm, Q = 2.57–2.96 (n = 90/3).
Distribution—China.
Notes—
Auricularia tibetica is characterized morphologically by the thick basidiomata with obvious lobed margin and ecologically by the growth on gymnosperm wood.
Auricularia americana is another species in the genus growing on gymnosperm species, and it can be distinguished by shorter basidia (55–71 × 4–5 µm vs. 70–103 × 4–7 µm) and thinner basidiospores (14–16.5 × 4.5–5.4 µm vs. 15–18.5 × 5.8–6.2 µm). Phylogenetically,
A. tibetica forms a single lineage with strong support and intraspecific genetic variations exist among samples of the species (
Figure 1 and
Figure 2). The morphological description of
A. tibetica taken from Wu et al. [
25].
Specimens examined—China. Shanxi Province, Ningwu County, Qiutiangou Forest Farm, on dead tree of Picea, 7 August 2013, Y.C. Dai, Dai 13336 (BJFC 014799). Sichuan Province, Daocheng County, Sandui Village, 18 July 2015, Y.C. Dai, Dai 15604 (BJFC 019708). Xizang Autonomous Region (Tibet), Linzhi County, Kadinggou Park, on fallen gymnosperm trunk, 23 September 2014, B.K. Cui, Cui 12337 (BJFC 017251); Sejilashan, on fallen trunk of Abies, 23 September 2014, B.K. Cui, Cui 12266 (BJFC 17180), Cui 12267 (BJFC 017181, holotype), Cui 12268 (BJFC 017181). Yunnan Province, Yulong County, Laojunshan National Park, on dead tree of Abies, 31 August 2015, Y.C. Dai, Dai 15734 (BJFC 019838); 1 September 2015, Y.C. Dai, Dai 15752 (BJFC 019856).
≡Laschia tremellosa Fr., Summa vegetabilium Scandinaviae 2: 325, 1849.
Basidiomata—Gelatinous when fresh, clay pink to clay buff, solitary or caespitose, sessile or substipitate; pileus auriculate with slightly lobed margin, projecting up to 6 cm, 0.5–1.5 mm thick, 0.08–0.2 mm thick and cinnamon to clay buff when dry; upper surface pilose, sometimes with a few folds; hymenophore surface conspicuously porose-reticulate to almost poroid especially when dry.
Internal features—Medulla present or absent; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a wide or narrow septate lumen, apical tips acute or obtuse, single, 39–80 × 5–7 µm; hyphae with clamp connections and simple septa, 1–6.5 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 36–42 × 5–5.5 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, (10–)10.2–12(–12.5) × 4–5 µm, L = 11.17 µm, W = 4.39 µm, Q = 2.5–2.59 (n = 60/2).
Distribution—Mexico and tropical South America.
Notes—
Auricularia tremellosa was originally described from Mexico and then considered as synonym of
A. delicata based on morphology by Lowy and Kobayasi [
12,
17]. However, in the present study, we accept
A. tremellosa as an independent species because it is different from
A. delicata sensu stricto not only in morphology but also in the phylogenies (
Figure 1 and
Figure 2). Six Brazilian specimens were studied and their morphological characters fit the description of
A. tremellosa. Auricularia tremellosa differs from
A. delicata by its poroid hymenophore, smaller basidia (36–42 × 5–5.5 µm vs. 48–65 × 4–6 µm) and distribution in Mexico and tropical South America. In the phylogeny,
A. tremellosa is very distantly related to
A. delicata (
Figure 1 and
Figure 2).
Specimens examined—Brazil. Acre, Sena Madureira, Floresta Nacional de São Francisco, on angiosperm wood, 16 January 2016, A. Meiras-Ottoni, AMO 557 (URM 93424), Floresta Nacional do Macauã, on angiosperm wood, 29 January 2016, A. Meiras-Ottoni, AMO 589 (URM 93425), Reserva Extrativista do Cazumbá-Iracema 30 January 2016, A. Meiras-Ottoni, AMO 667 (URM 93426); Amazonas, Manaus, on rotten angiosperm wood, 12 May 2017, Y.C. Dai, Dai 17415 (BJFC 026426), Dai 17419 (BJFC 026428). Goiás, Pirenópolis, RPPN Vagafogo, on rotten angiosperm wood, 31 December 2016, R.L.M. Alvarenga, RLMA 370 (URM 93427).
Basidiomata—Gelatinous when fresh, fawn to fuscous or buff yellow to orange yellow, solitary or caespitose, sessile or substipitate; pileus auriculate or petaloid, sometimes with lobed margin, projecting up to 5 cm, 1–2 mm thick, 0.08–0.3 mm thick and greyish brown to vinaceous brown when dry; upper surface pilose, sometimes with folds; hymenophore surface with obvious folds.
Internal features—Medulla absent; crystals absent; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, sometimes septate, apical tips acute or obtuse, single, 40–90 × 4.5–6 µm; hyphae with clamp connections, 1–3 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 40–61 × 4–5 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores allantoid, hyaline, thin-walled, smooth, usually with one or two large guttules, IKI–, CB–, (12.6–)13–15.5(–16) × 5–6.1(–6.3) µm, L = 14.38 µm, W = 5.57 µm, Q = 2.39–2.53 (n = 90/3).
Distribution—Asia.
Notes—
Auricularia villosula is characterized by its fawn and usually thin basidiomata when dry, and obvious folds on the hymenophore. It is easily confused with
A. heimuer in the field, but it has larger basidiospores (13–15.5 × 5–6.1 µm vs. 11–13 × 4–5 µm) and is only distantly related to
A. heimuer in the phylogenies (
Figure 1 and
Figure 2).
Auricularia villosula has a wide distribution and is cultivated in China [
23], but it was considered as
A. heimuer in commercial markets.
Specimens examined—China. Anhui Province, Chuzhou, Shahe, on fallen trunk of Paulownia, 15 May 2009, B.K. Cui, Cui 6760 (BJFC 004614). Hunan Province, Changsha, Yuelushan, on fallen angiosperm trunk, 10 August 2015, Y.C. Dai, Dai 15994 (BJFC 020094). Inner Mongolia Autonomous Region, Chifeng, on fallen trunk of Sophora, 12 May 2014, T.Z. Liu, CFSZ 10473 (CFSZ). Jiangxi Province, Fuzhou, on living tree of Cinnamomum, 5 August 2013, Y.C. Dai, Dai 13453 (BJFC 014914); on living tree of Cercis, 21 July 2013, Y.C. Dai, Dai 13450 (BJFC 014911). Sichuan Province, Bazhong County, Bazhong Park, on dead tree of Ficus, 31 May 2010, Y.C. Dai, Dai 11633 (BJFC 009254); Yajiang County, on angiosperm trunk, 23 July 2015, Y.C. Dai, Dai 15605 (BJFC 019709). Russia. Primorye Territory, 20 June 1991, Mikulin, (LE 296422); Ussuriyskiy Nature Reserve, on fallen angiosperm trunk, 3 July 1990, Govorova, (VLA M-11348), 30 July 1992, Govorova, (VLA M-11291), 15 August 2011, V. Malysheva, (LE 262989).
Basidiomata—Gelatinous when fresh, cinnamon to reddish brown, solitary or caespitose, sessile; pileus discoid or auriculate, margin entire, projecting up to 3 cm, 0.2–0.5 mm thick, 0.14–0.16 mm thick when dry; upper surface pilose, sometimes with small folds, becoming vinaceous grey to mouse grey upon drying; hymenophore surface with obvious folds, becoming light vinaceous grey to ash-grey upon drying.
Internal features—Medulla present; crystals present, abundantly scattered throughout the cross-section; abhymenial hairs with a slightly swollen base, hyaline, thick-walled, with a narrow lumen, apical tips acute or obtuse, single, sometimes tufted, 80–120 × 4–5 µm; hyphae with clamp connections, 1–3 µm in diam in KOH; basidia clavate, transversely 3-septate, with oil guttules, 50–72 × 5–7 µm, sterigmata rarely observed; cystidioles absent.
Spores—Basidiospores not observed in the type but were reported as measuring 15–16 × 6–7 µm by Li (1984).
Distribution—China.
Notes—Auricularia xishaensis has very thin basidiomata when fresh like A. fibrillifera, but it has distinctly larger basidiospores than those of A. fibrillifera (11–14 × 4–5 µm). In addition, A. xishaensis is easily identified by obvious folds and white particles on the hymenophore surface and plenty of crystals scattered throughout the cross-section. We failed to extract DNA because the type is in poor condition, so the placement of A. xishaensis in Auricularia remain uncertain.
Specimen examined—China. Hainan Province, Sansha, East of Xisha Islands, on rotten branch of Messerschmidia, 11 April 1982, L.J. Li, HPNHM 1338 (BJM, holotype).


 ,
, 













































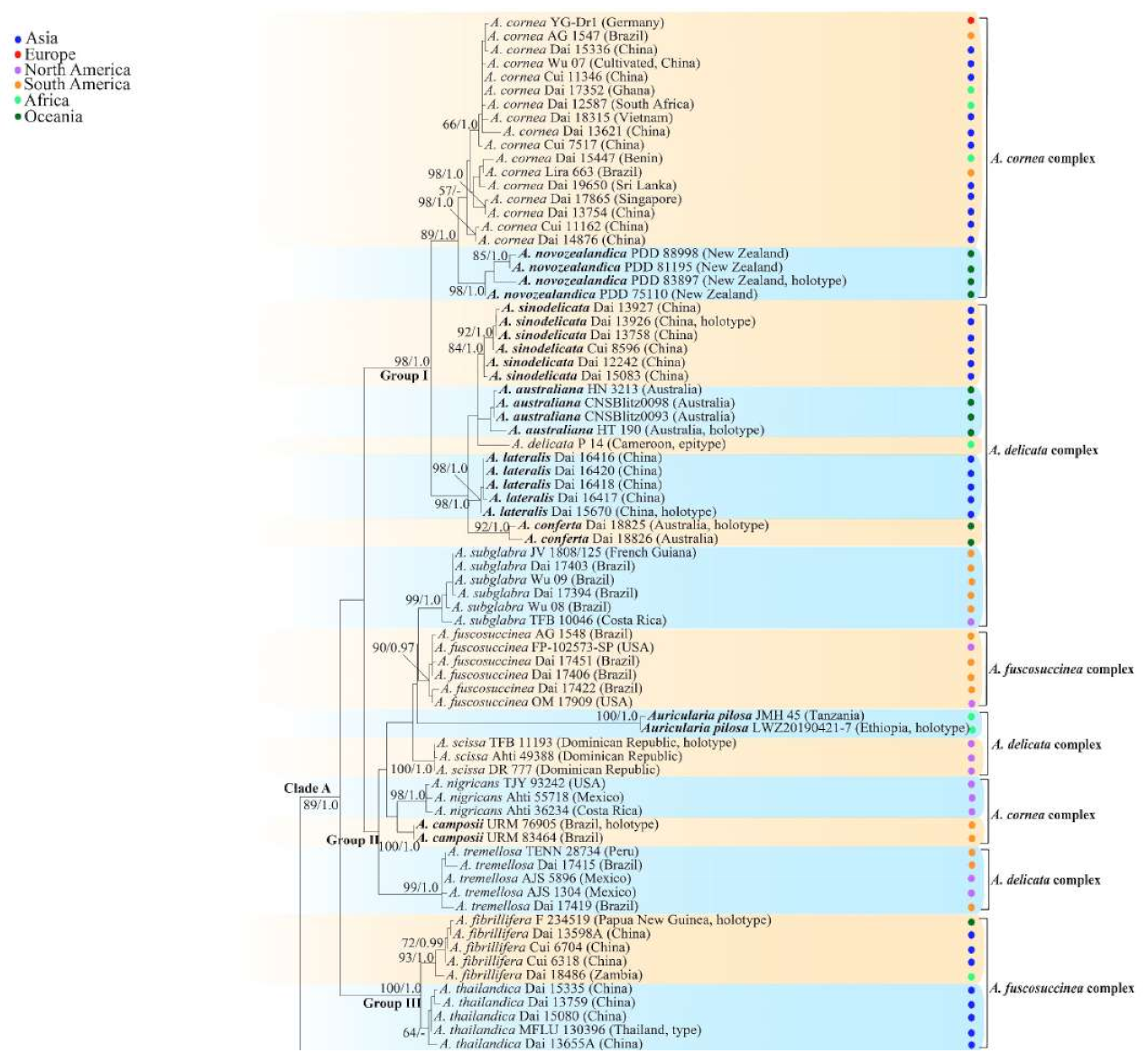{kind=link}
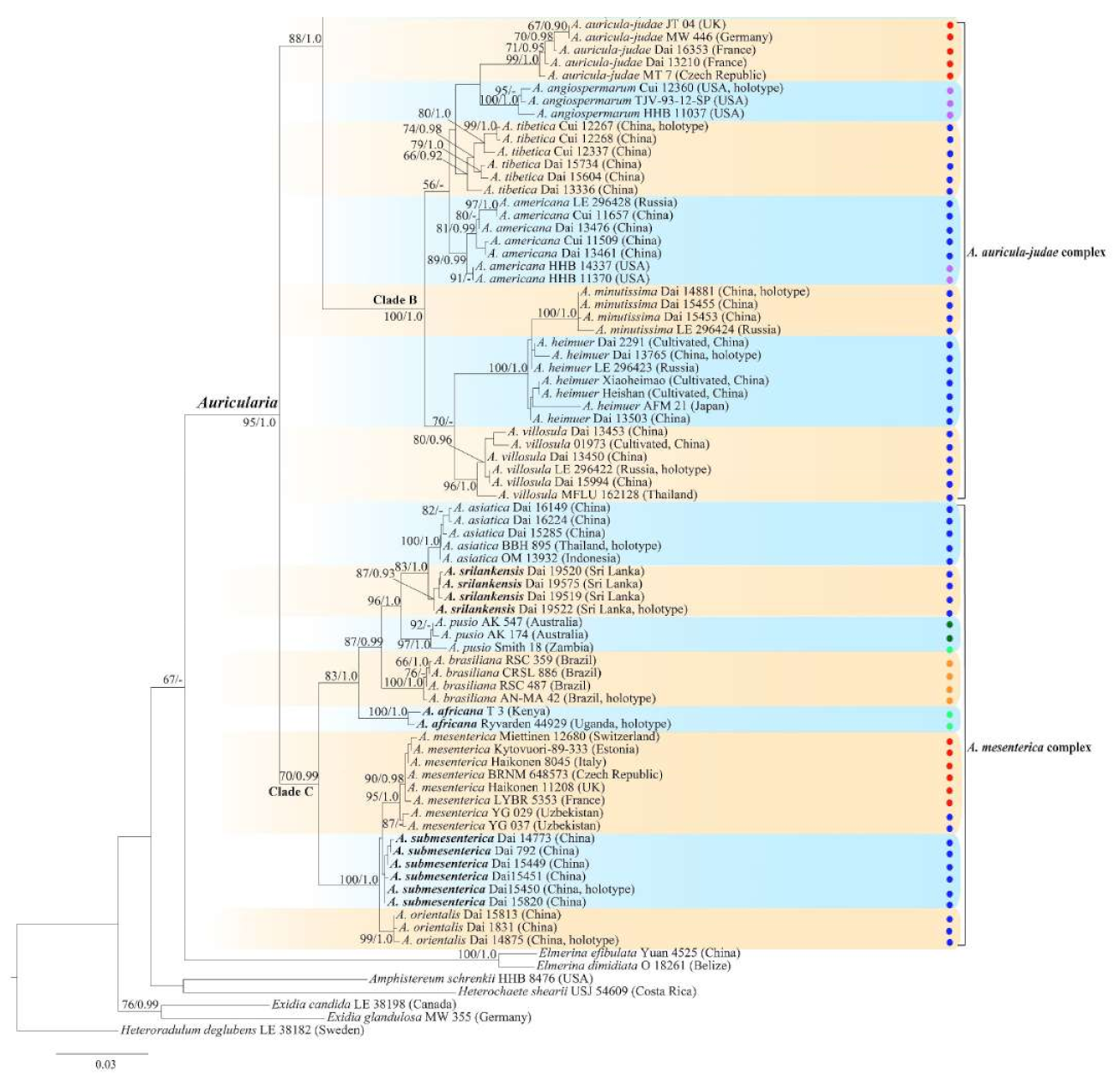{kind=link}
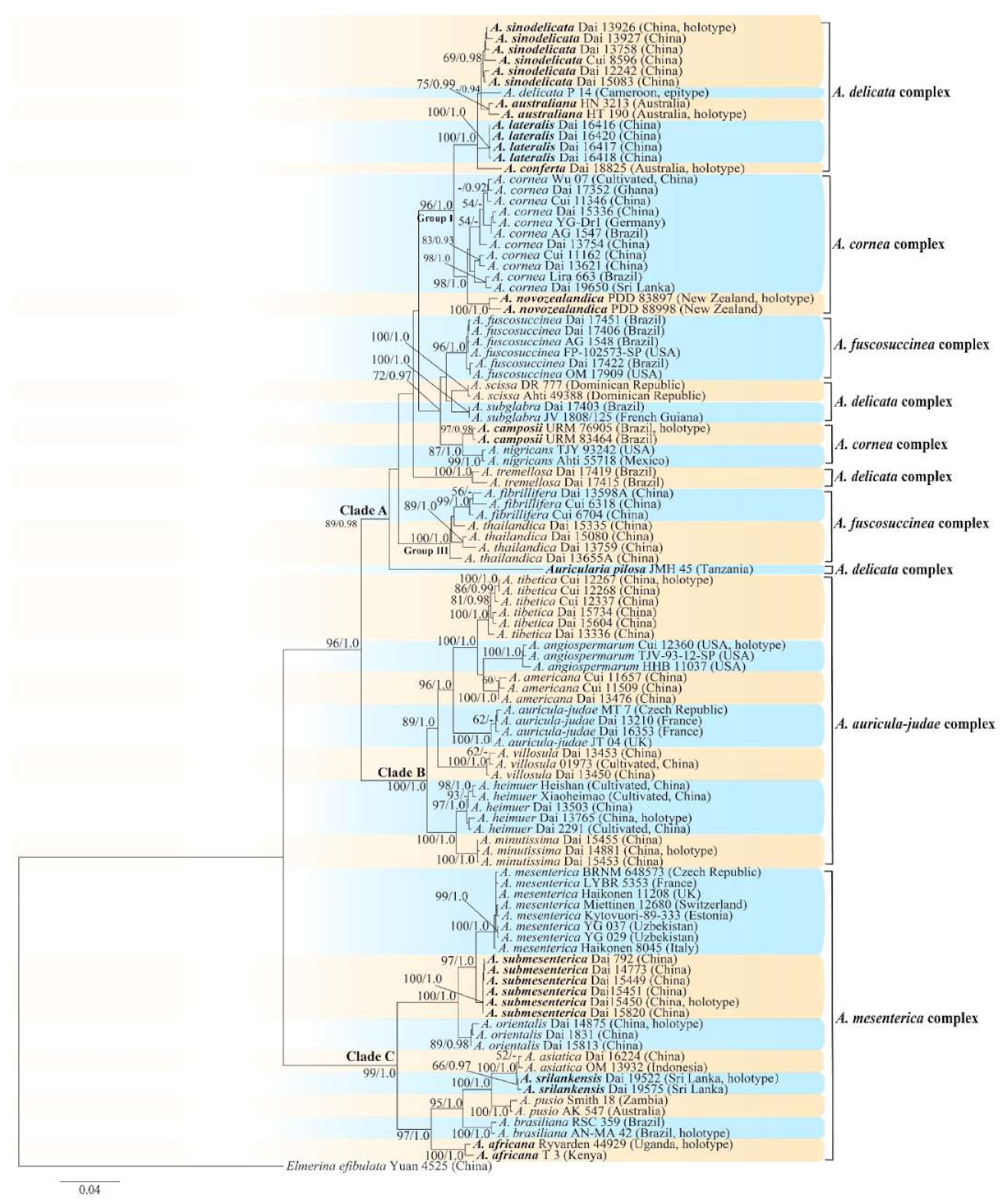{kind=link}
{kind=link}
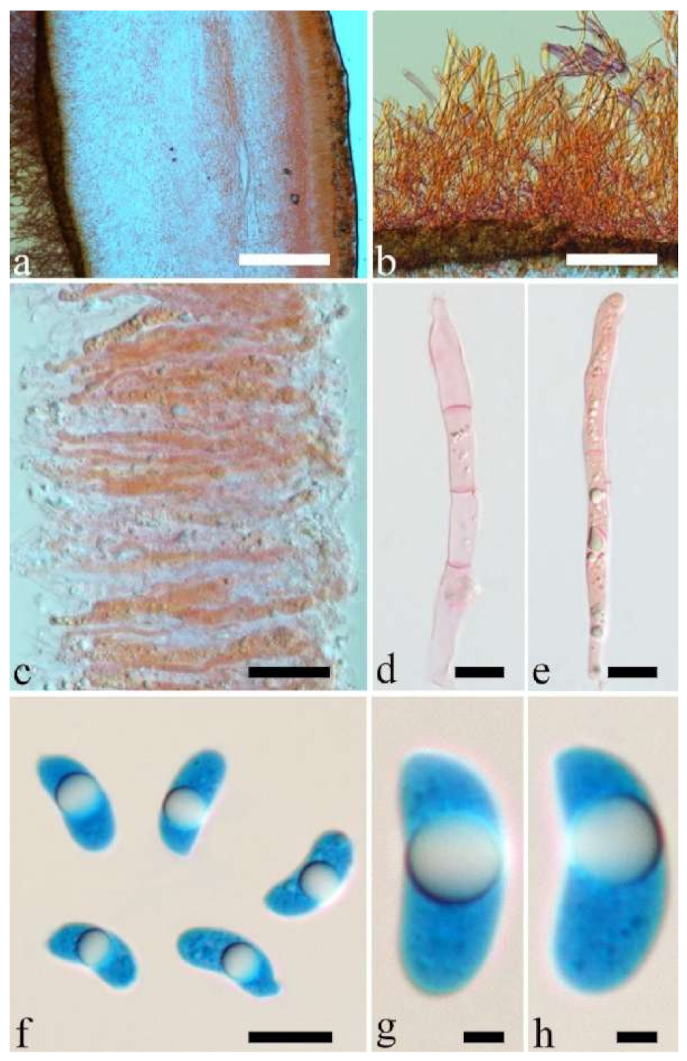{kind=link}
{kind=link}
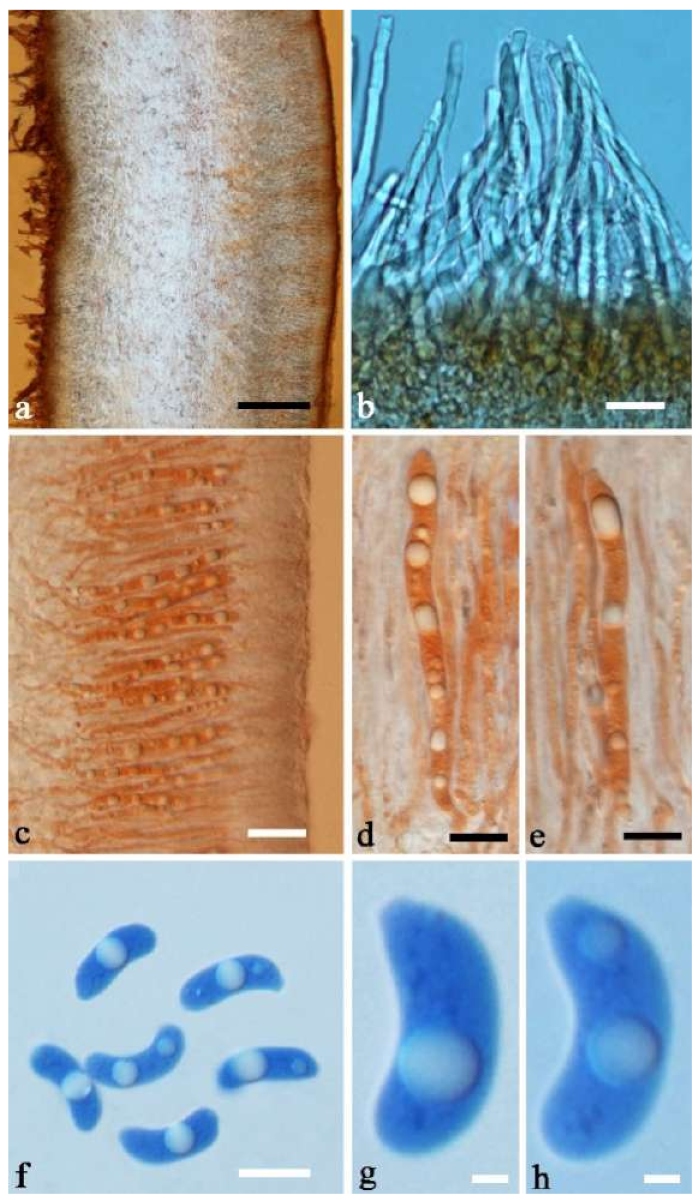{kind=link}
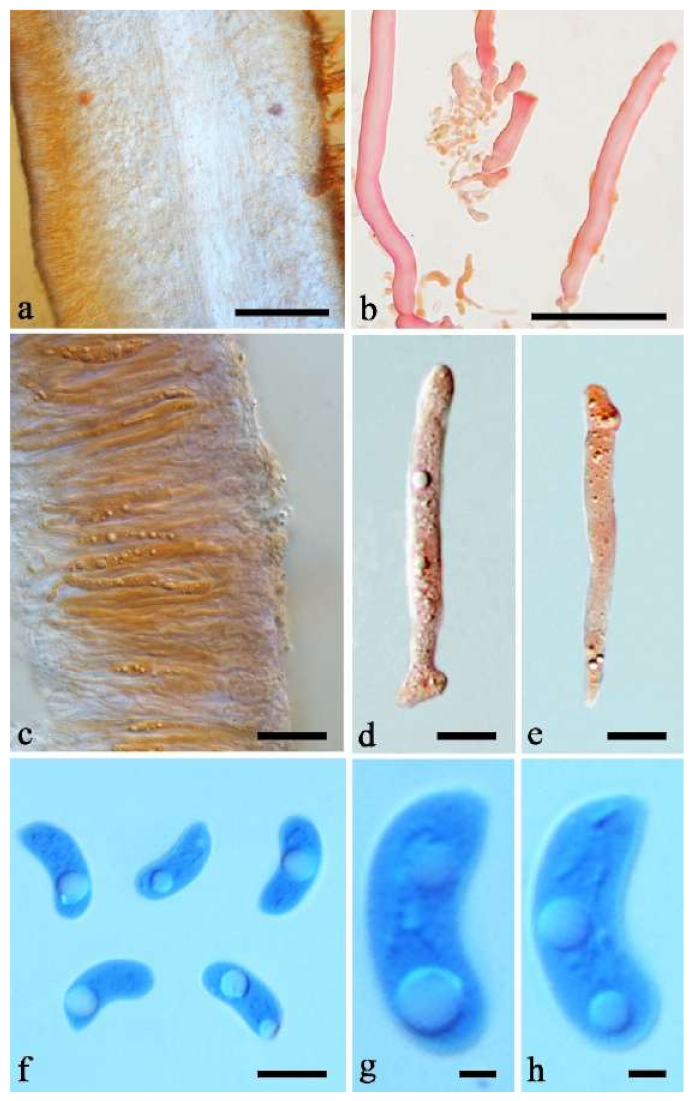{kind=link}
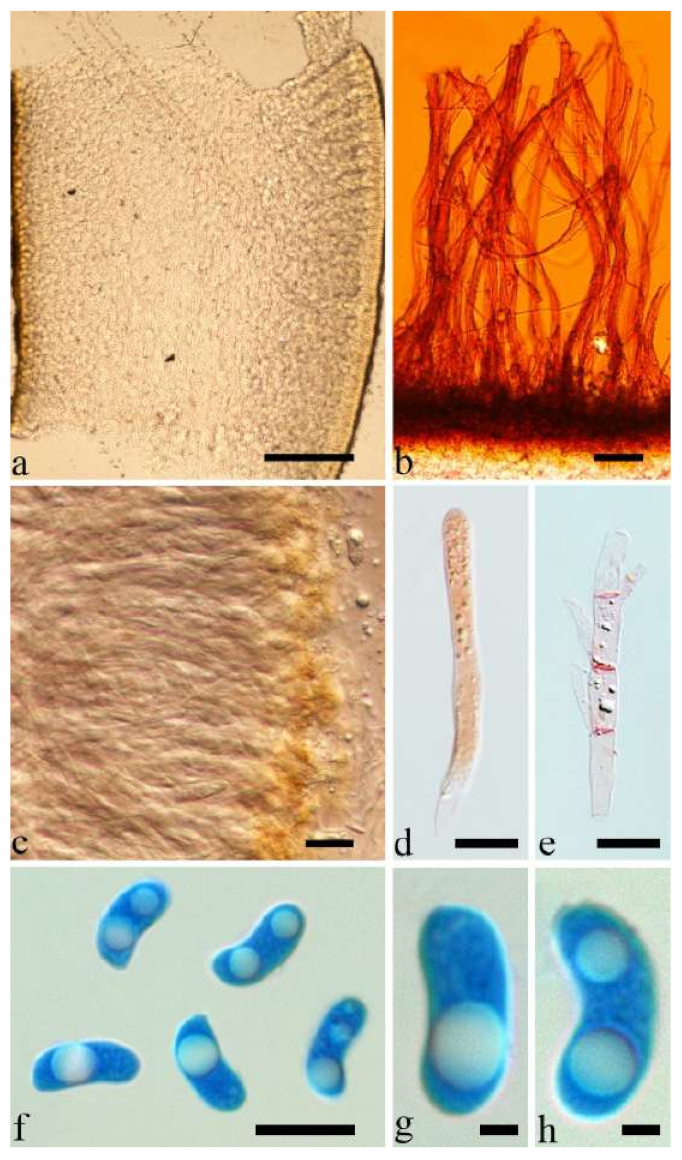{kind=link}
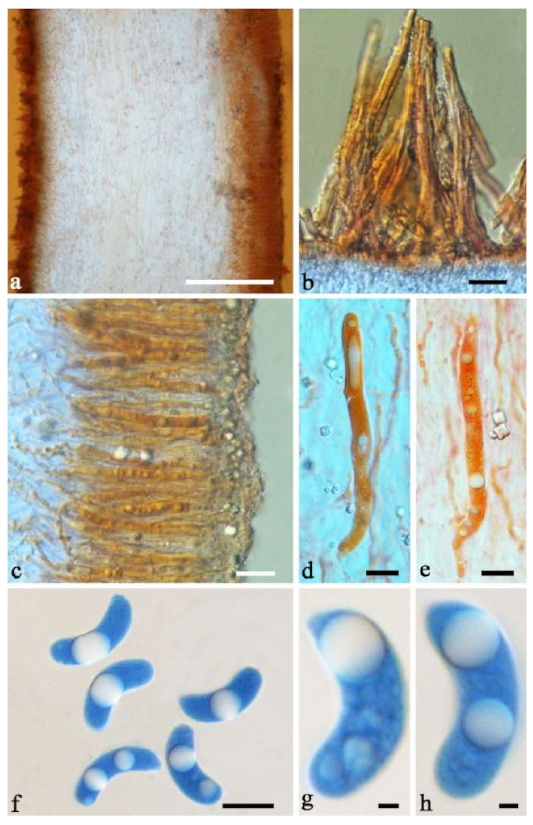{kind=link}
{kind=link}
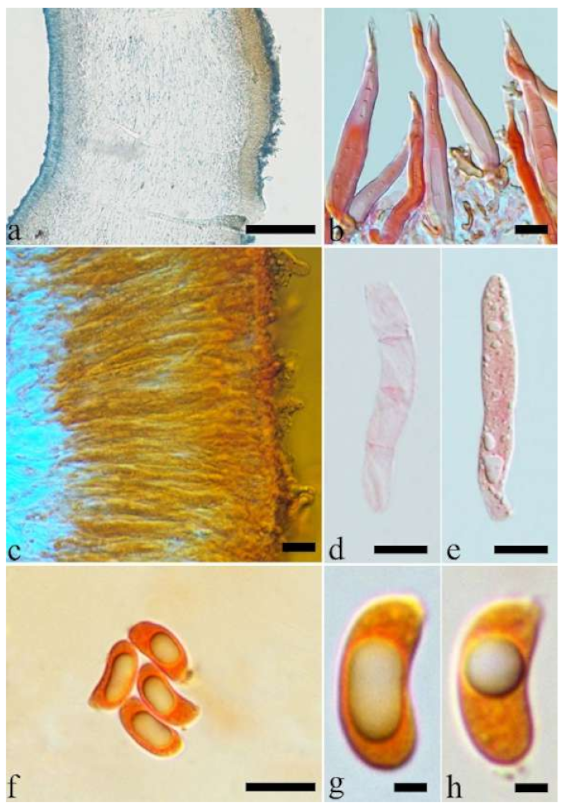{kind=link}
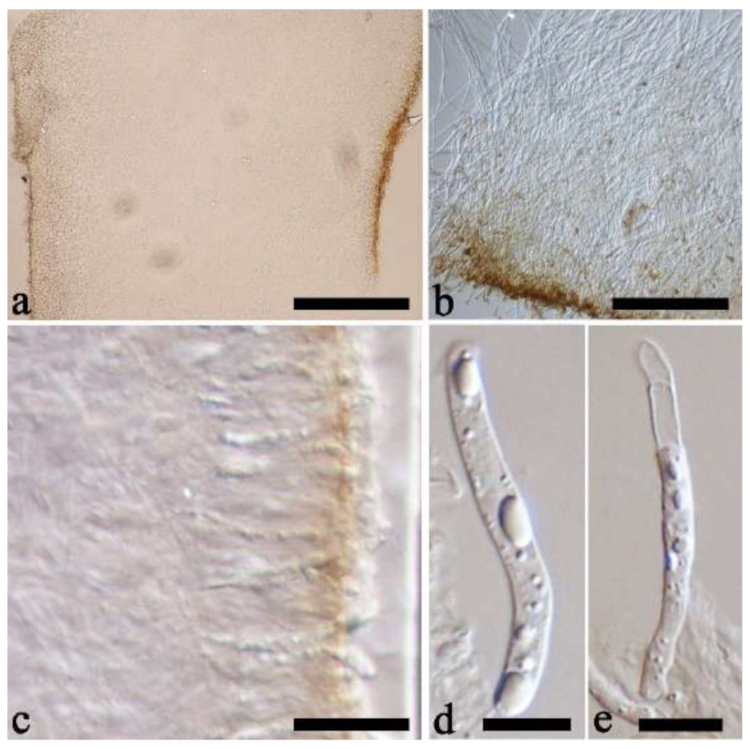{kind=link}
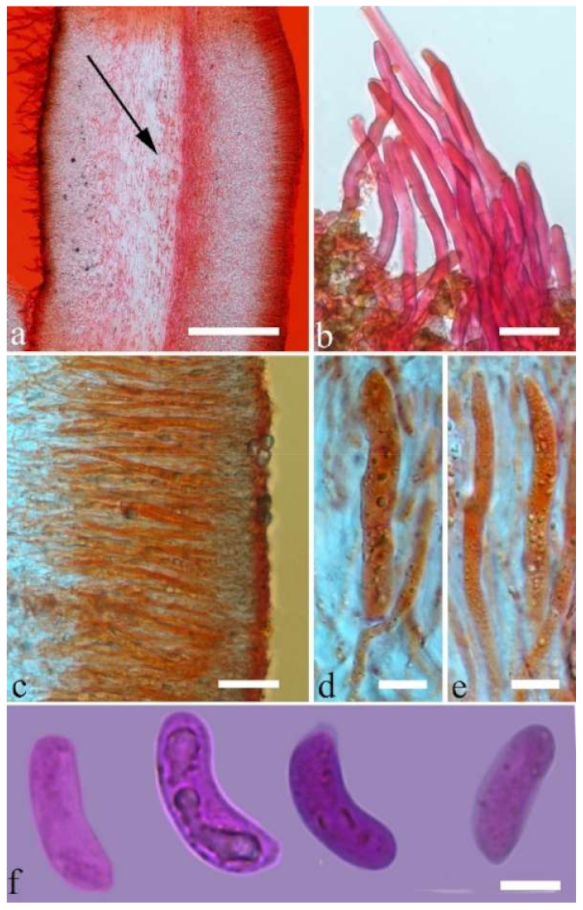{kind=link}
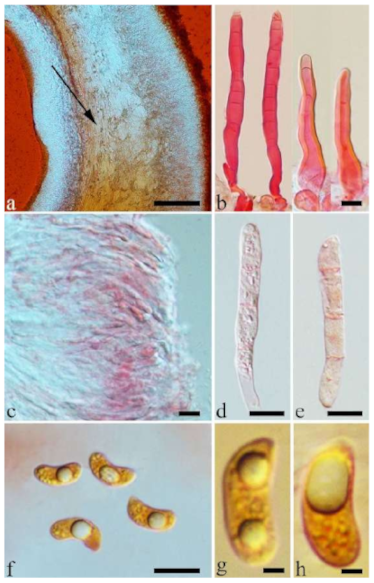{kind=link}
{kind=link}
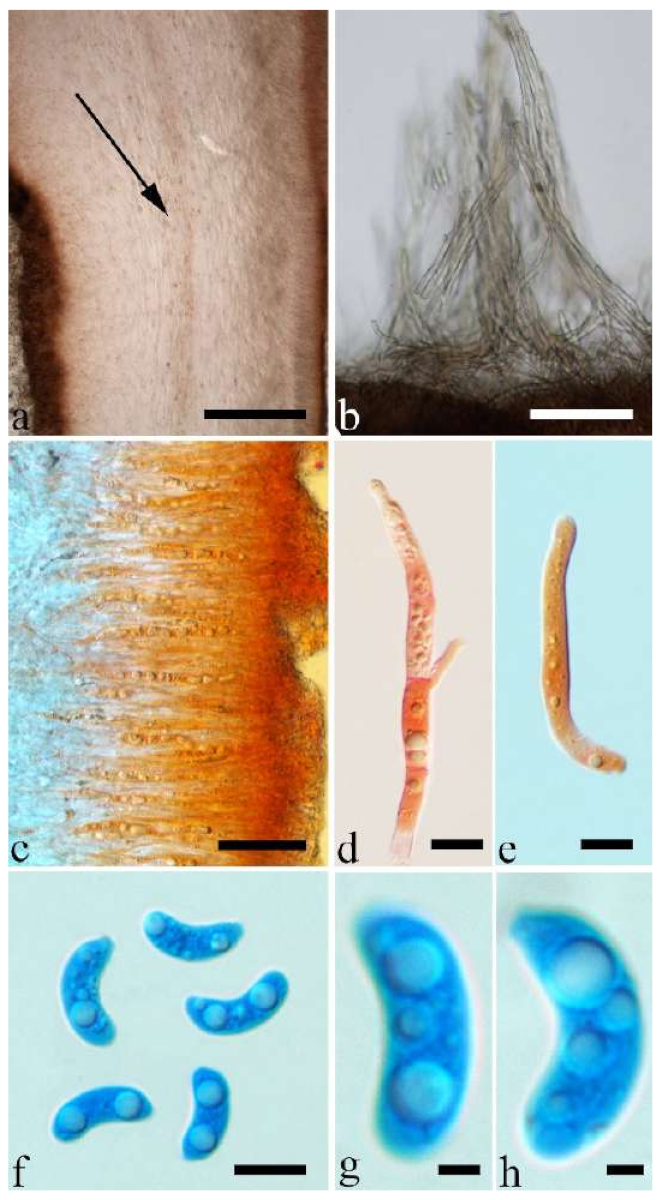{kind=link}
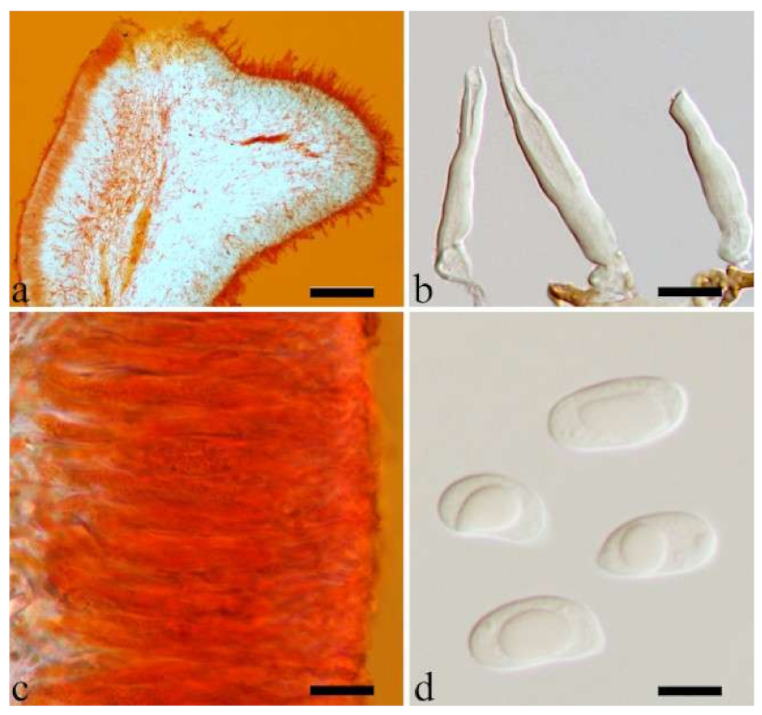{kind=link}
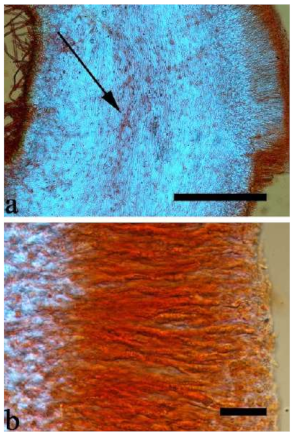{kind=link}
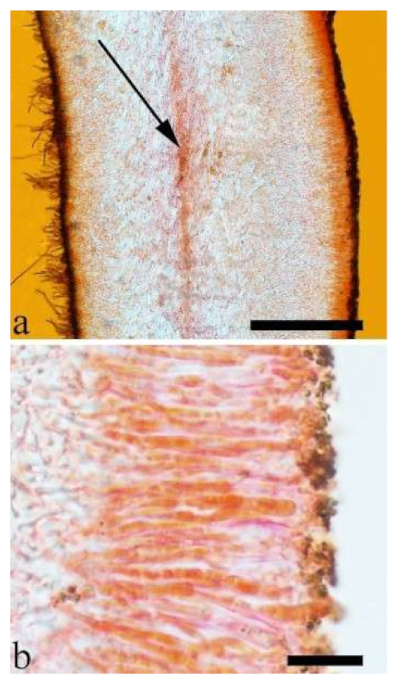{kind=link}
{kind=link}
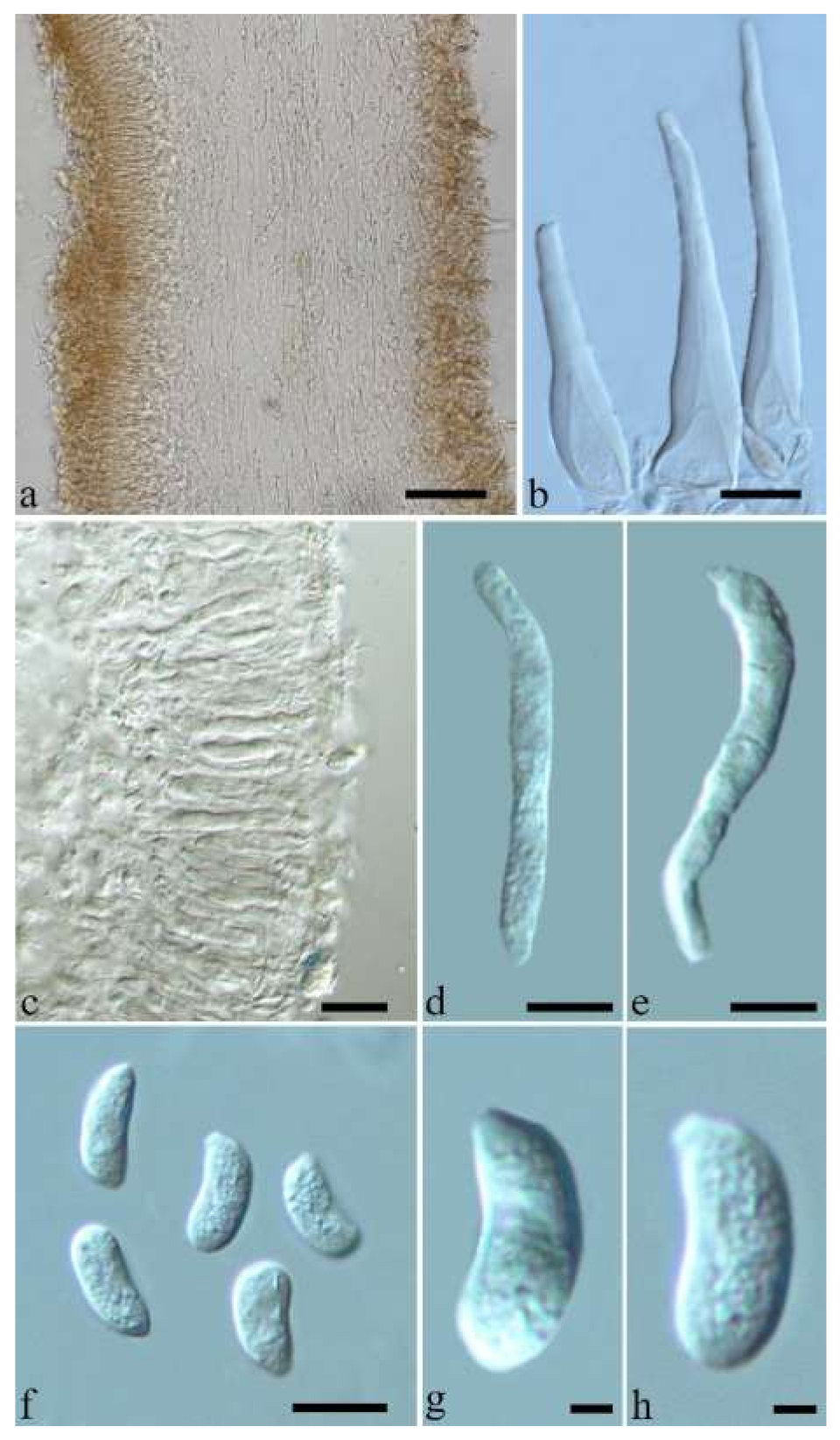{kind=link}
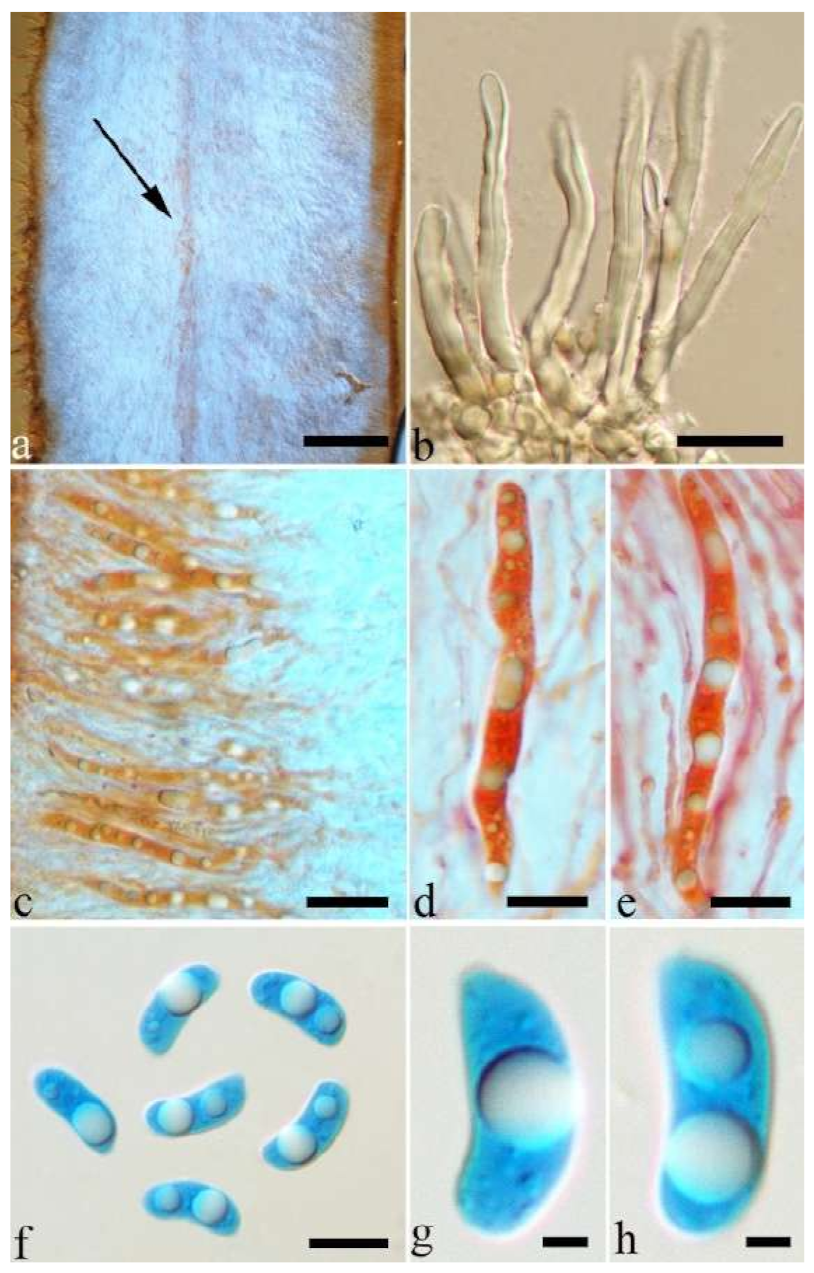{kind=link}
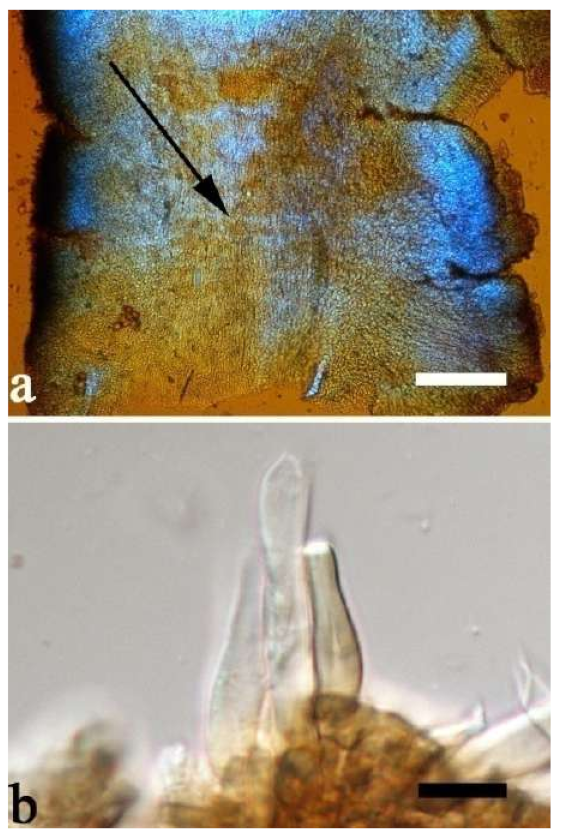{kind=link}
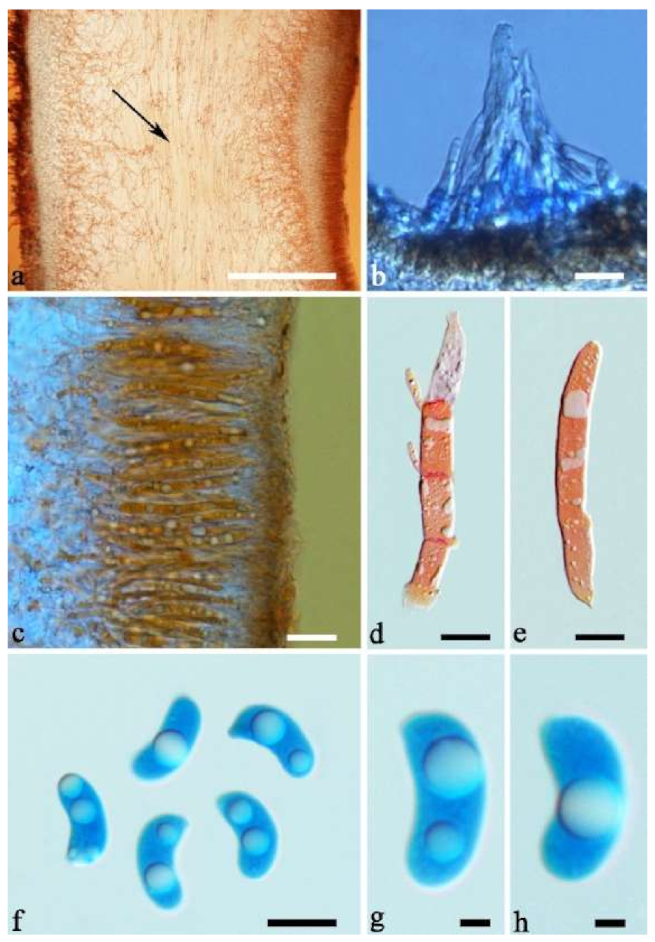{kind=link}
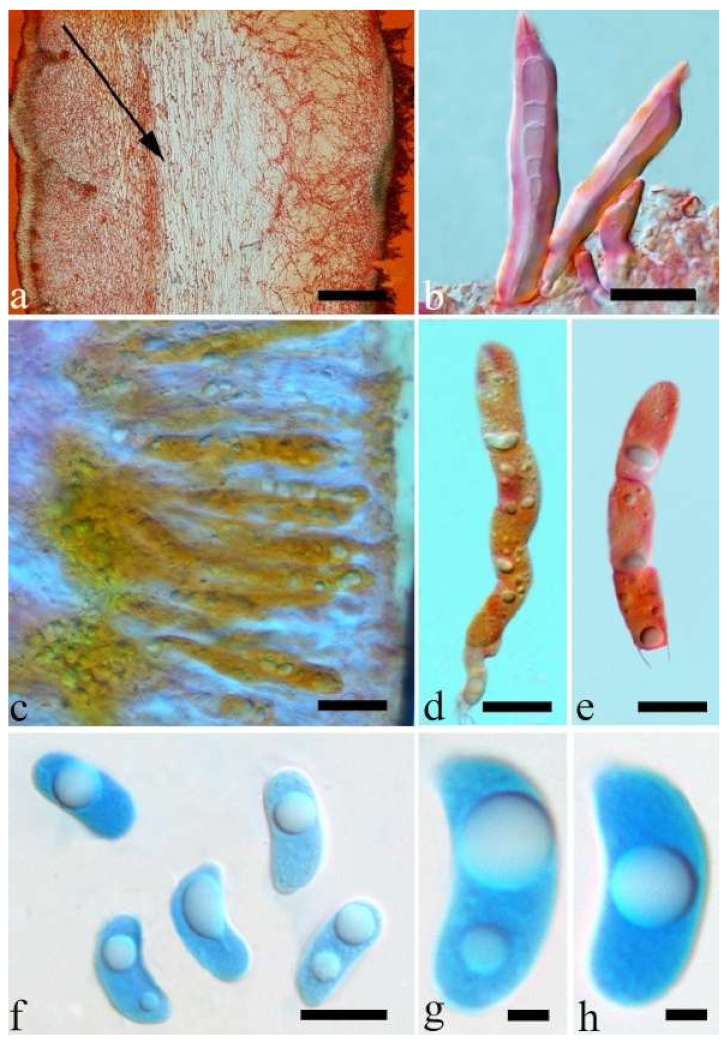{kind=link}
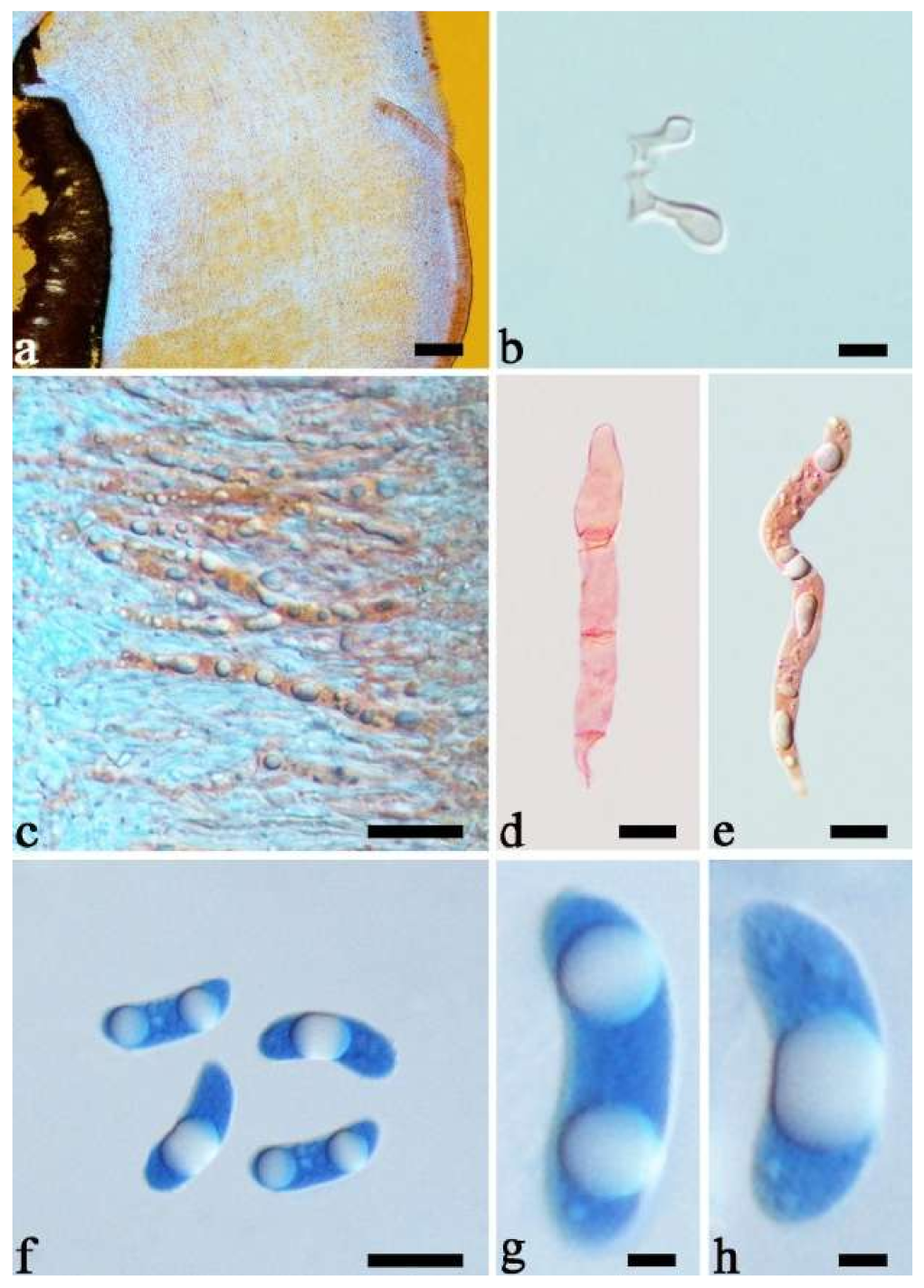{kind=link}
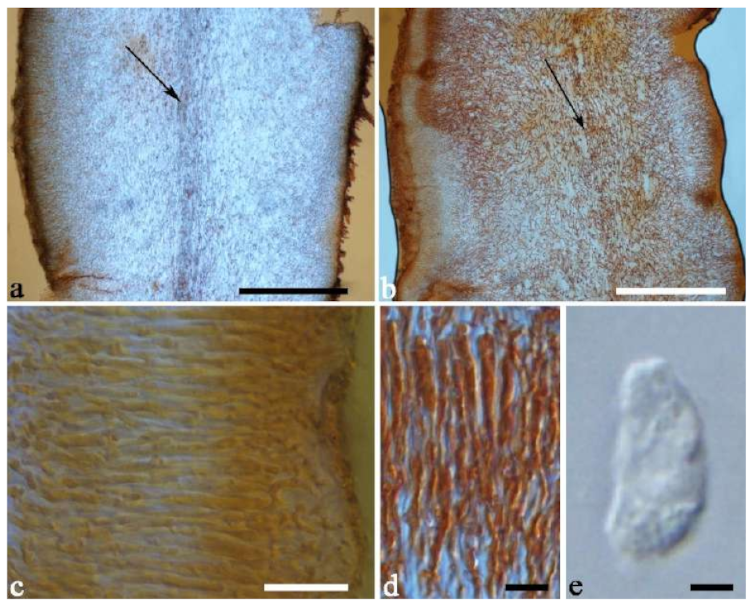{kind=link}
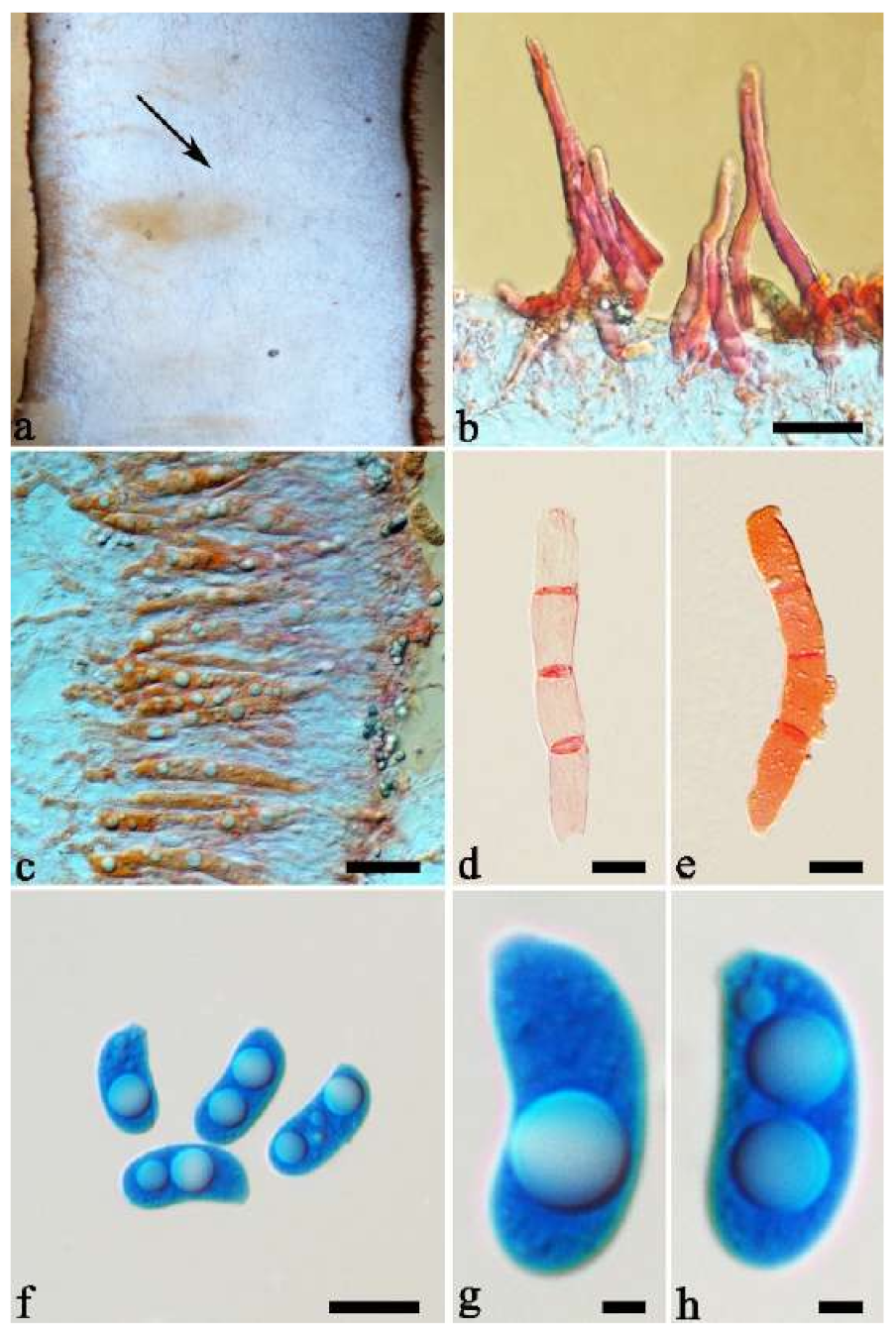{kind=link}
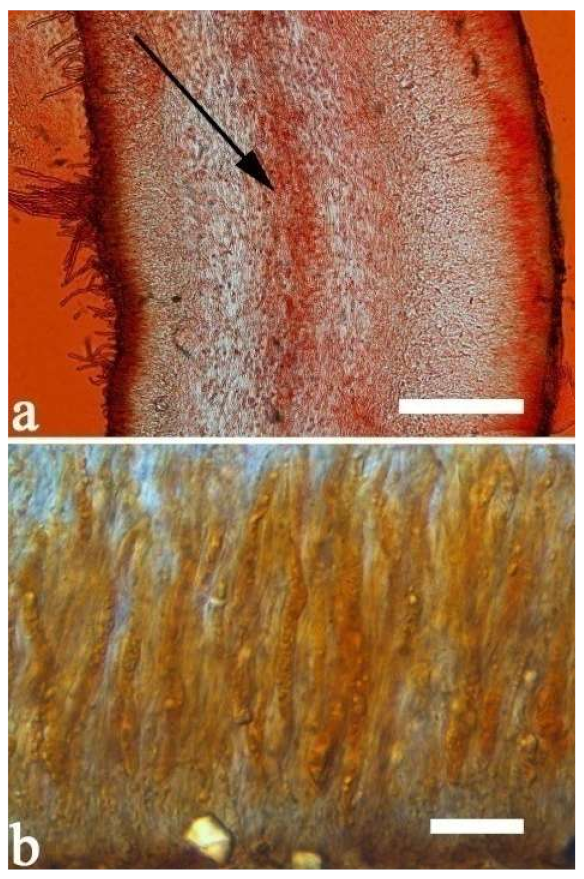{kind=link}
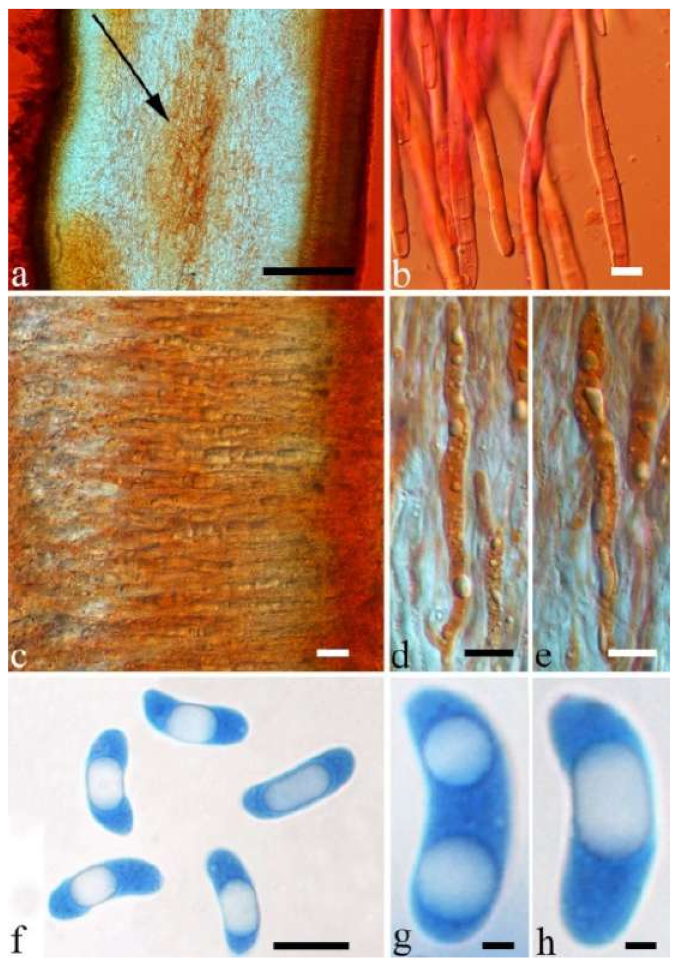{kind=link}
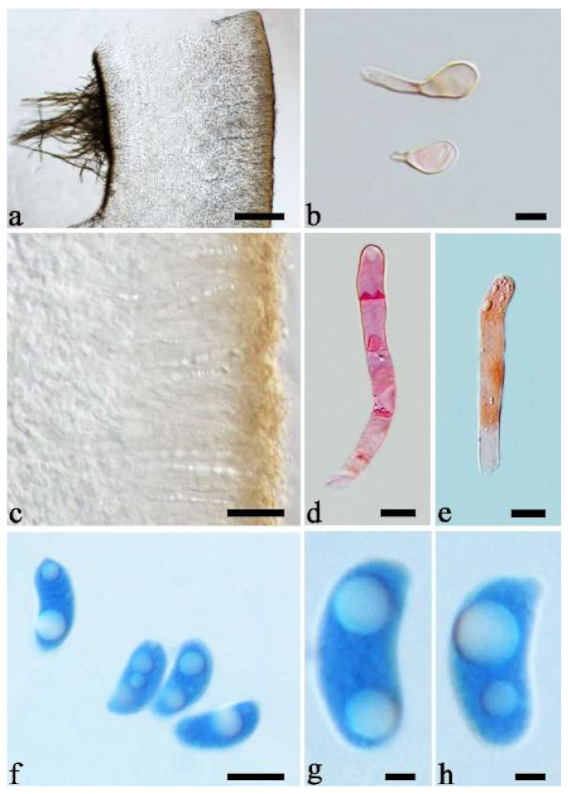{kind=link}
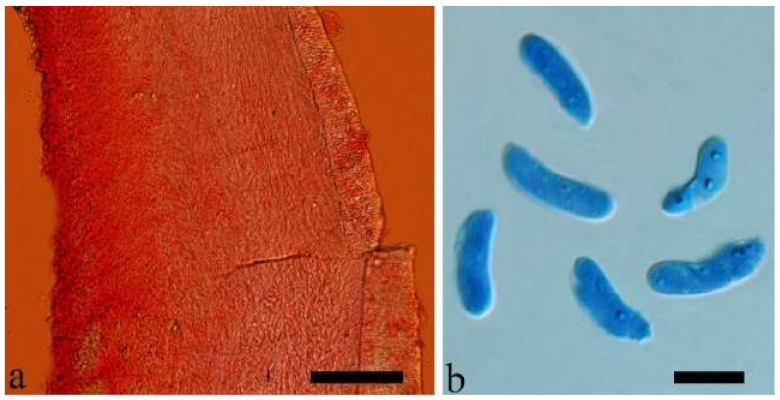{kind=link}
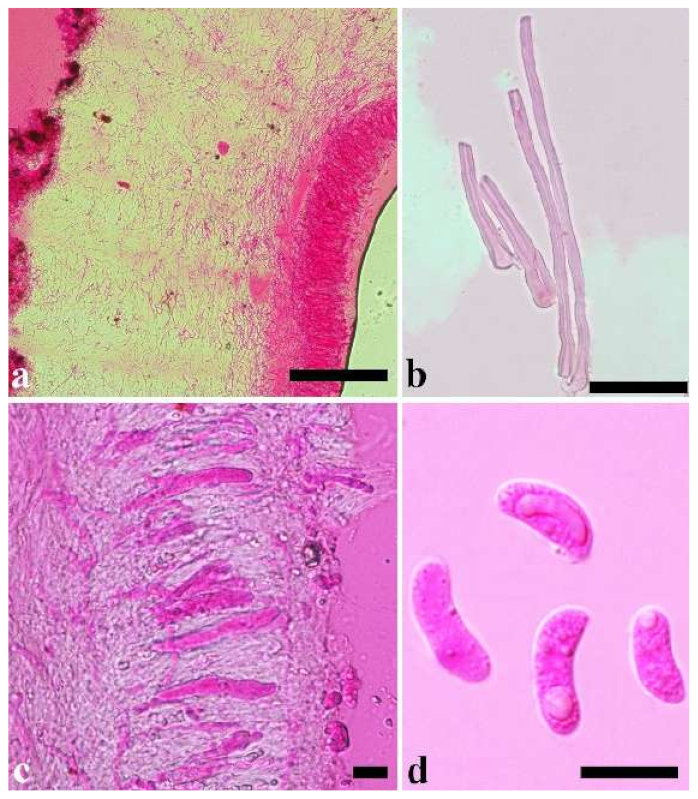{kind=link}
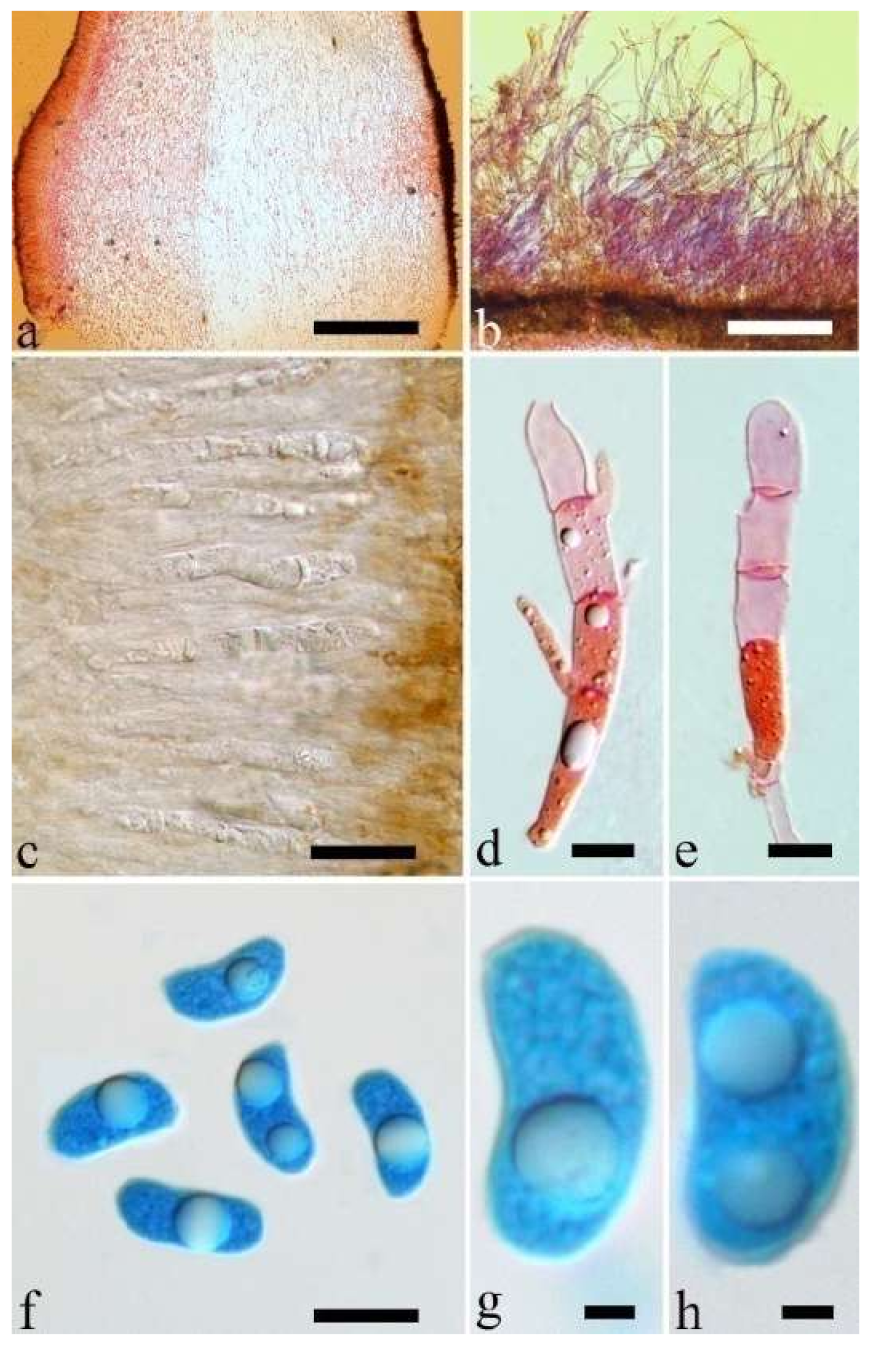{kind=link}
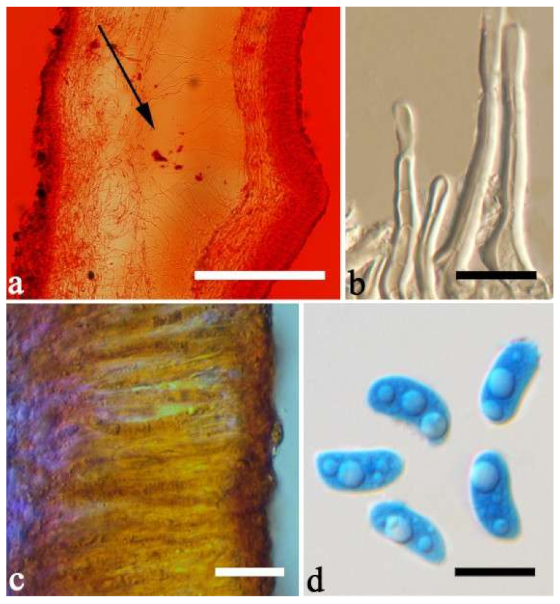{kind=link}
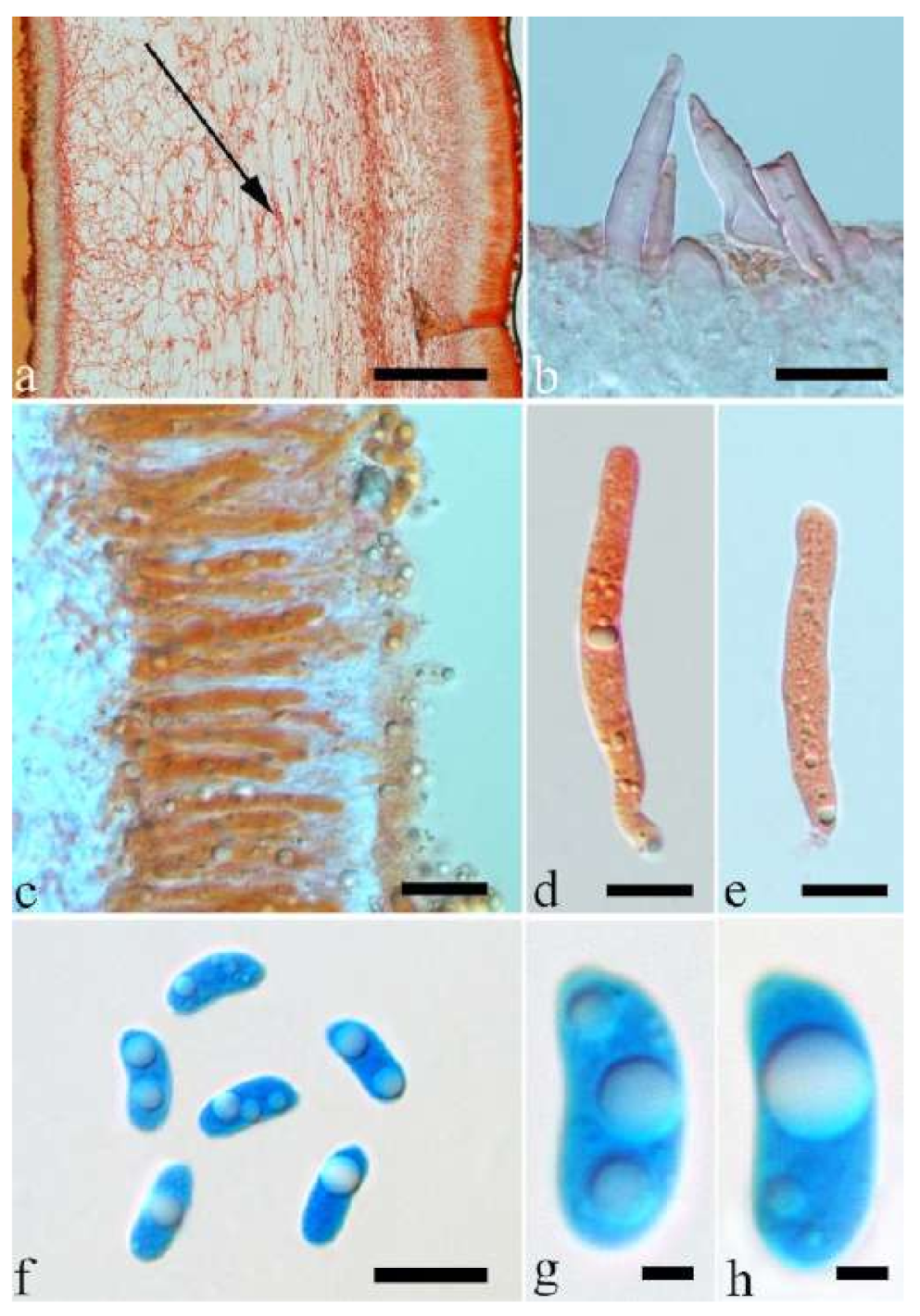{kind=link}
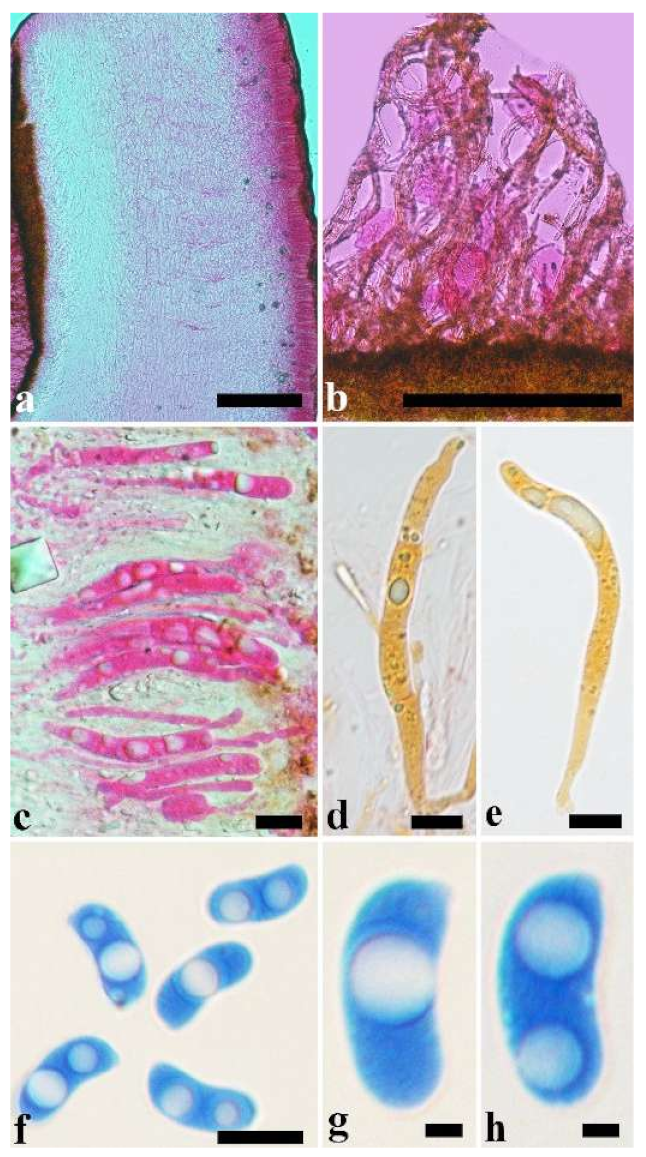{kind=link}
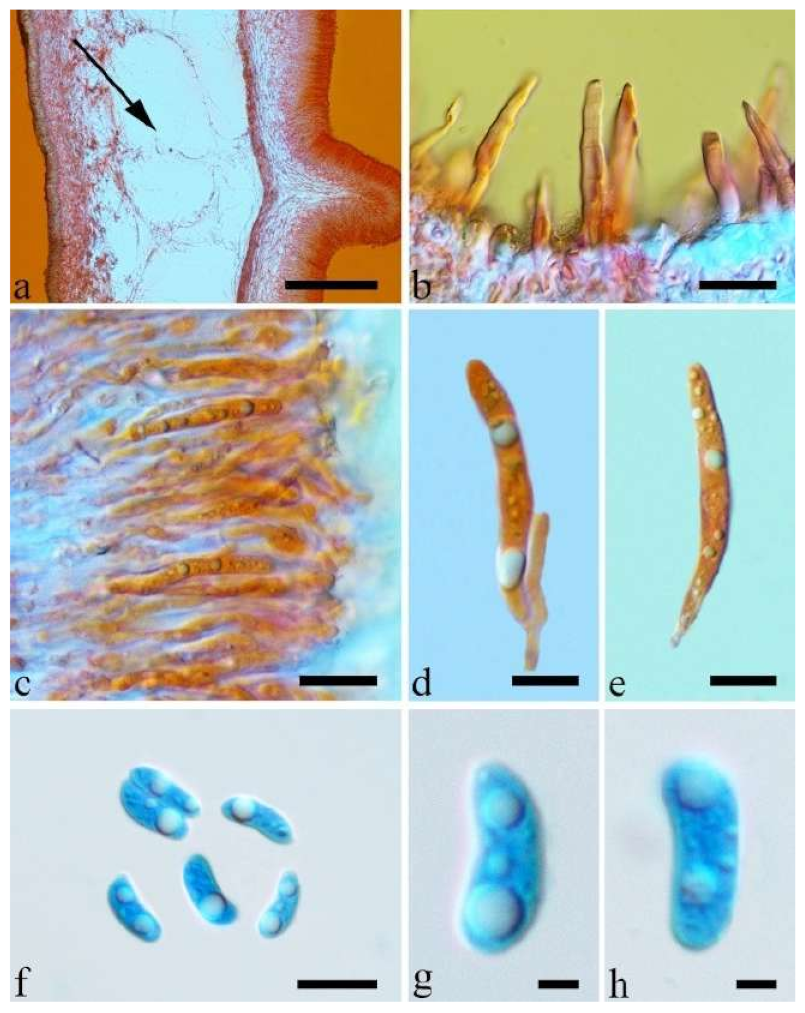{kind=link}
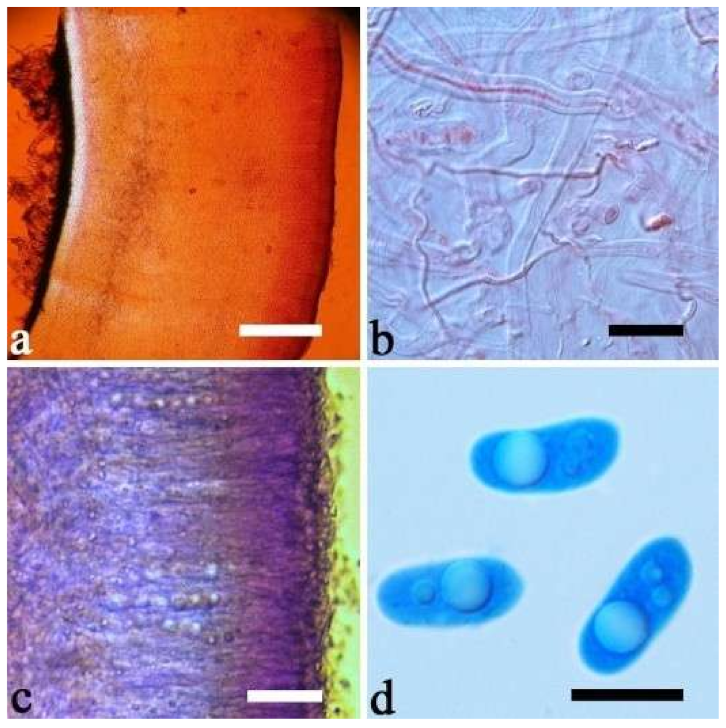{kind=link}
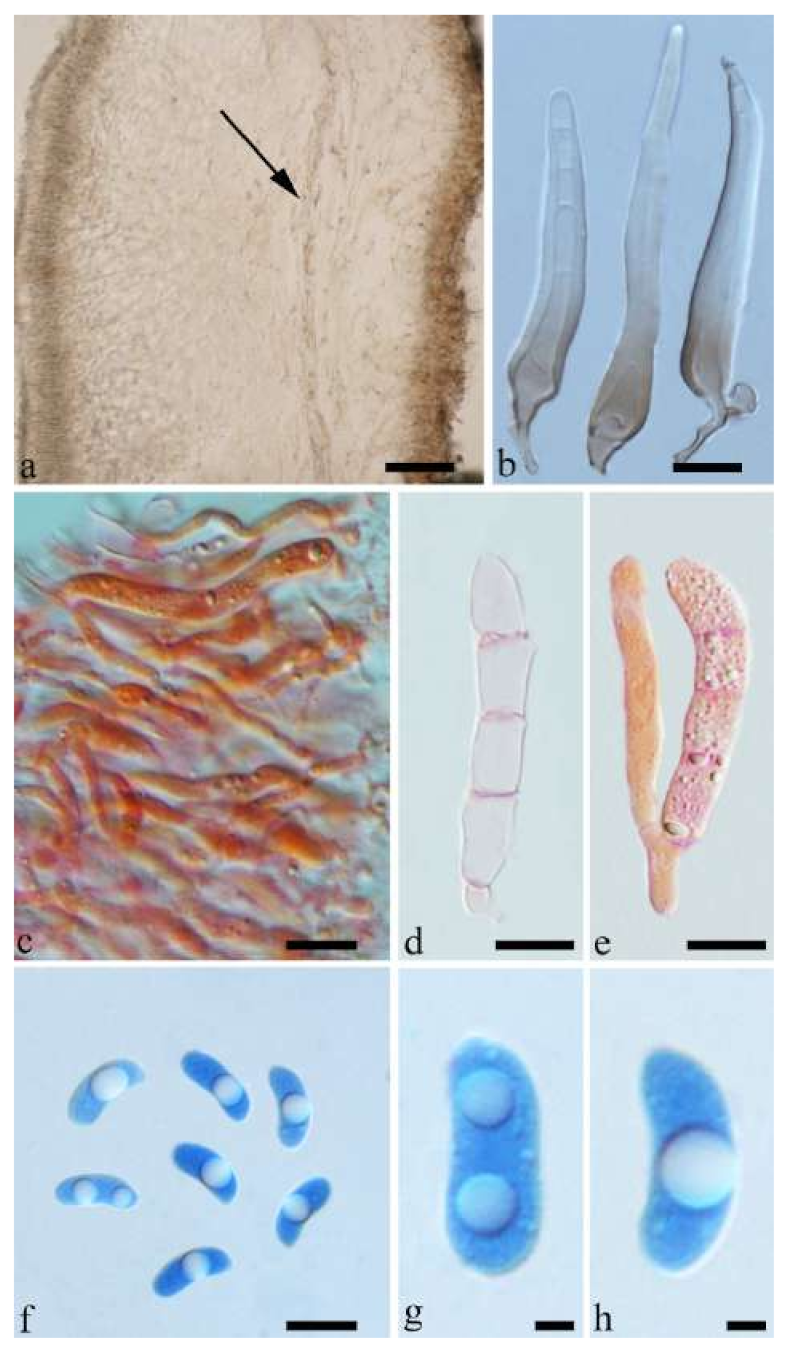{kind=link}
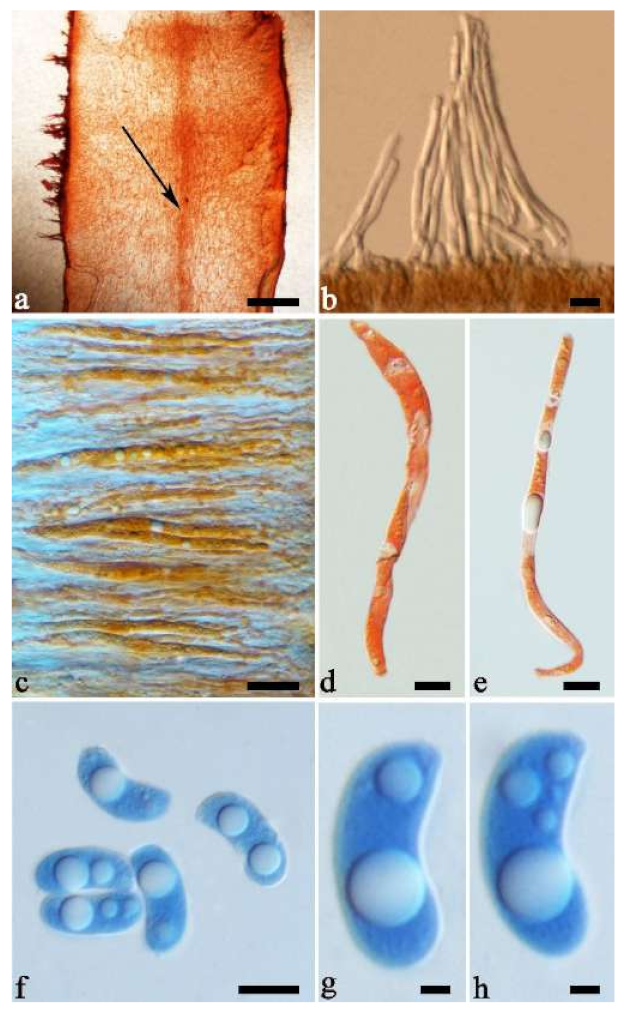{kind=link}
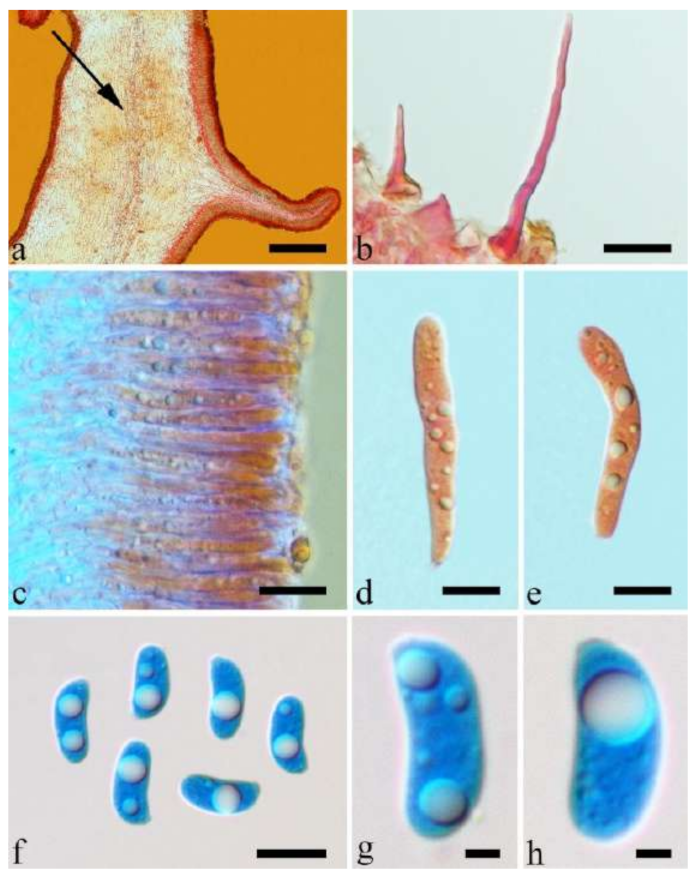{kind=link}
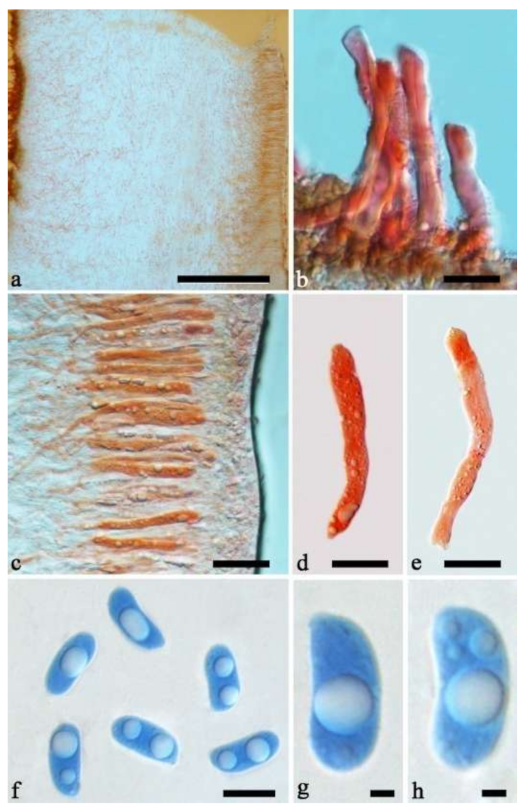{kind=link}
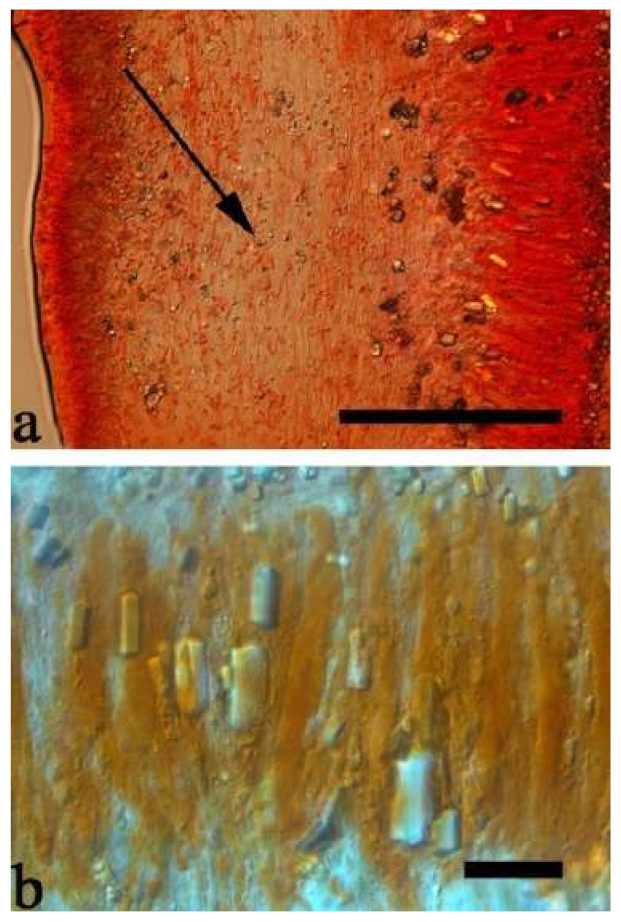{kind=link}