
Sensing and Responding to Hypersaline Conditions and the HOG Signal Transduction Pathway in Fungi Isolated from Hypersaline Environments: Hortaea werneckii and Wallemia ichthyophaga

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Molecular Adaptations of Halotolerant H. werneckii
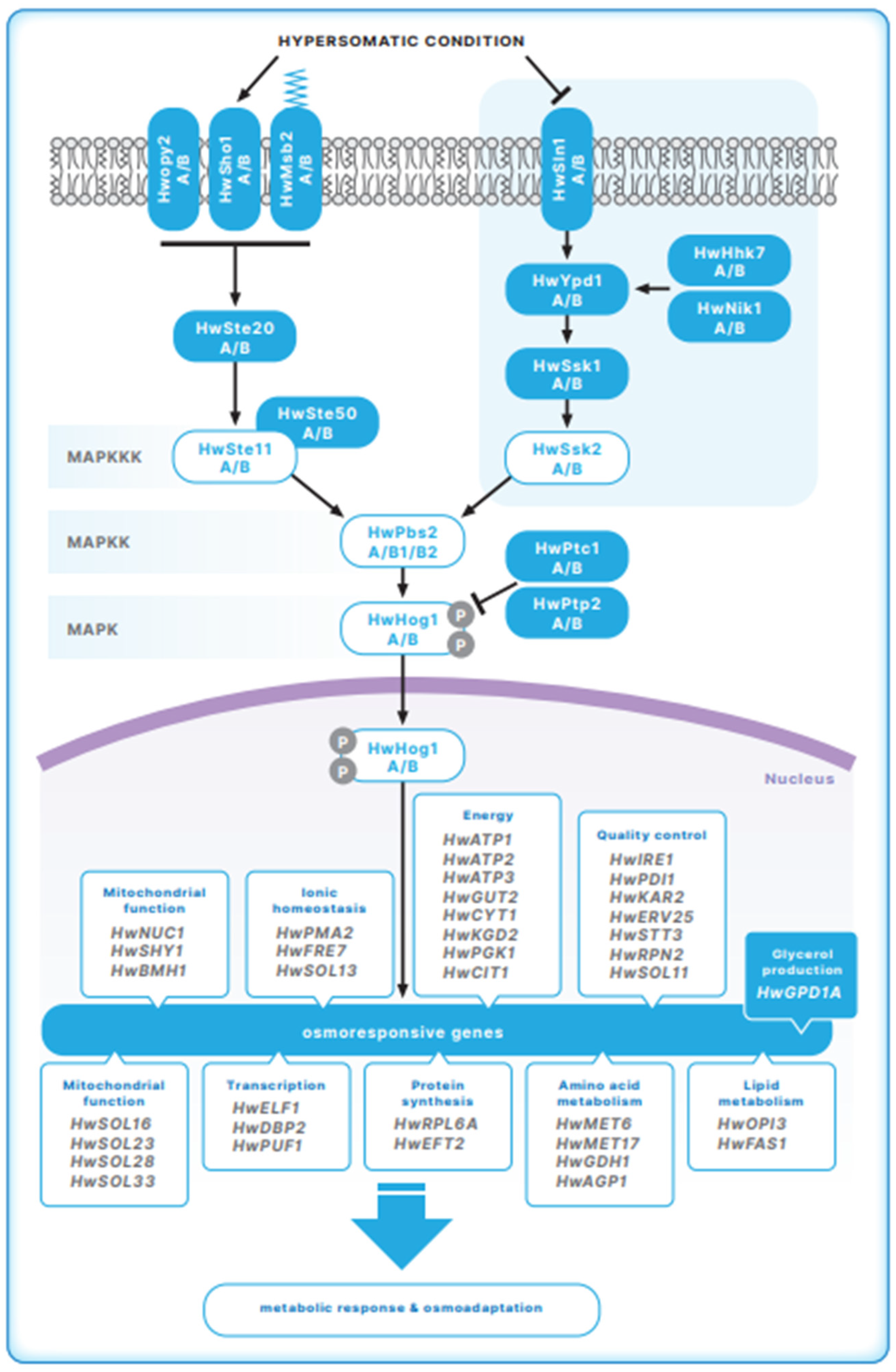
3. The HOG Signal Transduction Pathway in Halotolerant H. werneckii
3.1. The Sensory Apparatus of the HOG Pathway in H. werneckii
3.2. The MAP Kinase Module of the HOG Pathway in H. werneckii
4. Molecular Adaptations of the Halophilic Fungus W. ichthyophaga
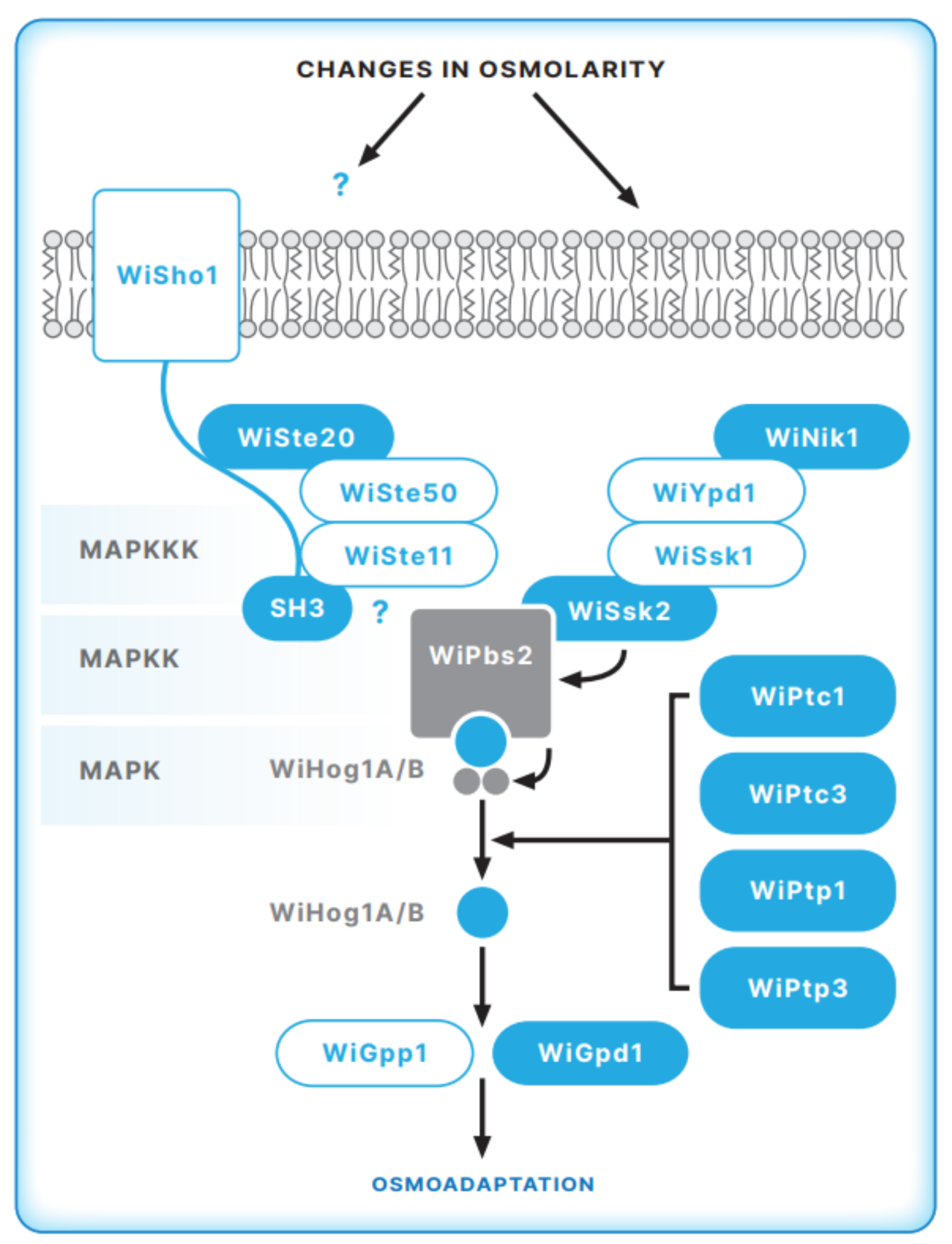
5. The HOG Signal Transduction Pathway in the Halophilic Fungus W. ichthyophaga
5.1. The Sensory Apparatus of the HOG Pathway in W. ichthyophaga
5.2. The MAP Kinase Module of the HOG Pathway in W. ichthyophaga
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Gunde-Cimerman, N.; Ramos, J.; Plemenitaš, A. Halotolerant and halophilic fungi. Mycol. Res. 2009, 113, 1231–1241. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Zalar, P. Extremely halotolerant and halophilic fungi inhabit brine in solar salterns around the globe. Food Technol. Biotechnol. 2014, 52, 170–179. [Google Scholar]
- Gunde-Cimerman, N.; Plemenitaš, A.; Oren, A. Strategies of adaptation of microorganisms of the three domains of life to high salt concentrations. FEMS Microbiol. Rev. 2018, 42, 353–375. [Google Scholar] [CrossRef] [PubMed]
- Petrovicˇ, U.; Gunde-Cimerman, N.; Plemenitaš, A. Cellular responses to environmental salinity in the halophilic black yeast Hortaea werneckii. Mol. Microbiol. 2002, 45, 665–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plemenitaš, A.; Lenassi, M.; Konte, T.; Kejžar, A.; Zajc, J.; Gostinčar, C.; Gunde-Cimerman, N. Adaptation to high salt concentrations in halotolerant/halophilic fungi: A molecular perspective. Front. Microbiol. 2014, 5, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zajc, J.; Kogej, T.; Galinski, E.A.; Ramos, J.; Gunde-Cimerman, N. Osmoadaptation Strategy of the Most Halophilic Fungus, Wallemia ichthyophaga, Growing Optimally at Salinities above 15% NaCl. Appl. Environ. Microbiol. 2014, 80, 247–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butinar, L.; Sonjak, S.; Zalar, P.; Plemenitaš, A.; Gunde-Cimerman, N. Melanized halophilic fungi are eukaryotic members of microbial communities in hypersaline waters of solar salterns. Bot. Mar. 2005, 48, 73–79. [Google Scholar] [CrossRef]
- Lenassi, M.; Gostinčar, C.; Jackman, S.; Turk, M.; Sadowski, I.; Nislow, C.; Jones, S.; Birol, I.; Cimerman, N.G.; Plemenitaš, A. Whole Genome Duplication and Enrichment of Metal Cation Transporters Revealed by De Novo Genome Sequencing of Extremely Halotolerant Black Yeast Hortaea werneckii. PLoS ONE 2013, 8, e71328. [Google Scholar] [CrossRef] [Green Version]
- Sinha, S.; Flibotte, S.; Neira, M.; Formby, S.; Plemenitaš, A.; Cimerman, N.G.; Lenassi, M.; Gostinčar, C.; Stajich, J.E.; Nislow, C. Insight into the Recent Genome Duplication of the Halophilic Yeast Hortaea werneckii: Combining an Improved Genome with Gene Expression and Chromatin Structure. G3: Genes|Genomes|Genet. 2017, 7, 2015–2022. [Google Scholar] [CrossRef] [Green Version]
- Gostinčar, C.; Stajich, J.E.; Kejžar, A.; Sinha, S.; Nislow, C.; Lenassi, M.; Gunde-Cimerman, N. Seven Years at High Salinity—Experimental Evolution of the Extremely Halotolerant Black Yeast Hortaea werneckii. J. Fungi 2021, 7, 723. [Google Scholar] [CrossRef]
- Lenassi, M.; Zajc, J.; Gostinčar, C.; Gorjan, A.; Gunde-Cimerman, N.; Plemenitaš, A. Adaptation of the glycerol-3-phosphate dehydrogenase Gpd1 to high salinities in the extremely halotolerant Hortaea werneckii and halophilic Wallemia ichthyophaga. Fungal Biol. 2011, 115, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Gostinčar, C.; Turk, M.; Plemenitaš, A.; Gunde-Cimerman, N. The expressions of [delta]9-,[delta]12-desaturases and an elongase by the extremely halotolerant Hortaea werneckii are salt dependent. FEMS Yeast Res. 2009, 9, 247–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turk, M.; Méjanelle, L.; Šentjurc, M.; Grimalt, J.O.; Gunde-Cimerman, N.; Plemenitaš, A. Salt-induced changes in lipid composition and membrane fluidity of halophilic yeast-like melanized fungi. Extremophiles 2003, 8, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Turk, M.; Plemenitaš, A.; Gunde-Cimerman, N. Extremophilic yeasts: Plasma-membrane fluidity as determinant of stress tolerance. Fungal Biol. 2011, 115, 950–958. [Google Scholar] [CrossRef] [PubMed]
- Gorjan, A.; Plemenitaš, A. Identification and characterization of ENA ATPases HwENA1 and HwENA2 from the halophilic black yeast Hortaea werneckii. FEMS Microbiol. Lett. 2006, 265, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kogej, T.; Ramos, J.; Plemenitaš, A.; Gunde-Cimerman, N. The Halophilic Fungus Hortaea werneckii and the Halotolerant Fungus Aureobasidium pullulans Maintain Low Intracellular Cation Concentrations in Hypersaline Environments. Appl. Environ. Microbiol. 2005, 71, 6600–6605. [Google Scholar] [CrossRef] [Green Version]
- Plemenitaš, A.; Konte, T.; Gostinčar, C.; Cimerman, N.G. Transport systems in halophilic fungi. In Yeast Membrane Transport—Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2016; Volume 892, pp. 307–325. [Google Scholar]
- Vaupotic, T.; Plemenitaš, A. Osmoadaptation-dependent activity of microsomal HMG-CoA reductase in the extremely halotolerant black yeastHortaea werneckiiis regulated by ubiquitination. FEBS Lett. 2007, 581, 3391–3395. [Google Scholar] [CrossRef] [Green Version]
- Vaupotič, T.; Veranic, P.; Petrovič, U.; Gunde-Cimerman, N.; Plemenitaš, A. HMG-CoA reductase is regulated by environmental salinity and its activity is essential for halotolerance in halophilic fungi. Stud. Mycol. 2008, 61, 61–66. [Google Scholar] [CrossRef]
- Vaupotič, T.; Gunde-Cimerman, N.; Plemenitaš, A. Novel 3′-phosphoadenosine-5′-phosphatases from extremely halotolerant Hortaea werneckii reveal insight into molecular determinants of salt tolerance of black yeasts. Fungal Genet. Biol. 2007, 44, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Gašparič, M.B.; Lenassi, M.; Gostinčar, C.; Rotter, A.; Plemenitaš, A.; Gunde-Cimerman, N.; Gruden, K.; Žel, J. Insertion of a Specific Fungal 3′-phosphoadenosine-5′-phosphatase Motif into a Plant Homologue Improves Halotolerance and Drought Tolerance of Plants. PLOS ONE 2013, 8, e81872. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, S. Osmotic Stress Signaling and Osmoadaptation in Yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 300–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohmann, S. Control of high osmolarity signalling in the yeastSaccharomyces cerevisiae. FEBS Lett. 2009, 583, 4025–4029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, H.; Posas, F. Response to Hyperosmotic Stress. Genetics 2012, 192, 289–318. [Google Scholar] [CrossRef] [Green Version]
- Tatebayashi, K.; Yamamoto, K.; Nagoya, M.; Takayama, T.; Nishimura, A.; Sakurai, M.; Momma, T.; Saito, H. Osmosensing and scaffolding functions of the oligomeric four-transmembrane domain osmosensor Sho1. Nat. Commun. 2015, 6, 6975. [Google Scholar] [CrossRef] [Green Version]
- Warmka, J.; Hanneman, J.; Lee, J.; Amin, D.; Ota, I. Ptc1, a Type 2C Ser/Thr Phosphatase, Inactivates the HOG Pathway by Dephosphorylating the Mitogen-Activated Protein Kinase Hog1. Mol. Cell. Biol. 2001, 21, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacoby, T.; Flanagan, H.; Faykin, A.; Seto, A.G.; Mattison, C.; Ota, I. Two Protein-tyrosine Phosphatases Inactivate the Osmotic Stress Response Pathway in Yeast by Targeting the Mitogen-activated Protein Kinase, Hog1. J. Biol. Chem. 1997, 272, 17749–17755. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, K.; Hoshi, Y.; Maeda, T.; Nakajima, T.; Abe, K. Aspergillus nidulans HOG pathway is activated only by two-component signalling pathway in response to osmotic stress. Mol. Microbiol. 2005, 56, 1246–1261. [Google Scholar] [CrossRef]
- Ma, D.; Li, R. Current Understanding of HOG-MAPK Pathway in Aspergillus fumigatus. Mycopathologia 2012, 175, 13–23. [Google Scholar] [CrossRef]
- Bahn, Y.-S. Master and Commander in Fungal Pathogens: The Two-Component System and the HOG Signaling Pathway. Eukaryot. Cell 2008, 7, 2017–2036. [Google Scholar] [CrossRef] [Green Version]
- Jacob, S.; Foster, A.J.; Yemelin, A.; Thines, E. High osmolarity glycerol (HOG) signalling in Magnaporthe oryzae: Identification of MoYPD1 and its role in osmoregulation, fungicide action, and pathogenicity. Fungal Biol. 2015, 119, 580–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Román, E.; Correia, I.; Prieto, A.D.; Alonso, R.; Pla, J. The HOG MAPK pathway in Candida albicans: More than an osmosensing pathway. Int. Microbiol. 2019, 23, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Yamada-Okabe, T.; Mio, T.; Ono, N.; Kashima, Y.; Matsui, M.; Arisawa, M.; Yamada-Okabe, H. Roles of Three Histidine Kinase Genes in Hyphal Development and Virulence of the Pathogenic Fungus Candida albicans. J. Bacteriol. 1999, 181, 7243–7247. [Google Scholar] [CrossRef] [Green Version]
- Fettich, M.; Lenassi, M.; Veranič, P.; Gunde-Cimerman, N.; Plemenitaš, A. Identification and characterization of putative osmosensors, HwSho1A and HwSho1B, from the extremely halotolerant black yeast Hortaea werneckii. Fungal Genet. Biol. 2011, 48, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Kejžar, A.; Grötli, M.; Tamás, M.J.; Plemenitaš, A.; Lenassi, M. HwHog1 kinase activity is crucial for survival of Hortaea werneckii in extremely hyperosmolar environments. Fungal Genet. Biol. 2015, 74, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Turk, M.; Plemenitaš, A. The HOG pathway in the halophilic black yeastHortaea werneckii: Isolation of theHOG1homolog gene and activation of HwHog1p. FEMS Microbiol. Lett. 2002, 216, 193–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaupotič, T.; Plemenitaš, A. Differential gene expression and Hog1 interaction with osmoresponsive genes in the extremely halotolerant black yeast Hortaea werneckii. BMC Genom. 2007, 8, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenassi, M.; Plemenitaš, A. Novel group VII histidine kinase HwHhk7B from the halophilic fungi Hortaea werneckii has a putative role in osmosensing. Curr. Genet. 2007, 51, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Randhawa, A.; Chawla, S.; Mondal, A.K. Functional dissection of HAMP domains in NIK1 ortholog from pathogenic yeast Candida lusitaniae. Gene 2016, 577, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Catlett, N.L.; Yoder, O.C.; Turgeon, B.G. Whole-Genome Analysis of Two-Component Signal Transduction Genes in Fungal Pathogens. Eukaryot. Cell 2003, 2, 1151–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, K.-H.; Prade, R.A. Osmotic stress-coupled maintenance of polar growth in Aspergillus nidulans. Mol. Microbiol. 2002, 43, 1065–1078. [Google Scholar] [CrossRef] [PubMed]
- O’Rourke, S.M.; Herskowitz, I. Unique and Redundant Roles for HOG MAPK Pathway Components as Revealed by Whole-Genome Expression Analysis. Mol. Biol. Cell 2004, 15, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Bahn, Y.-S.; Geunes-Boyer, S.; Heitman, J. Ssk2 Mitogen-Activated Protein Kinase Kinase Kinase Governs Divergent Patterns of the Stress-Activated Hog1 Signaling Pathway in Cryptococcus neoformans. Eukaryot. Cell 2007, 6, 2278–2289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kejžar, A.; Cibic, M.; Grøtli, M.; Plemenitaš, A.; Lenassi, M. The unique characteristics of HOG pathway MAPKs in the extremely halotolerant Hortaea werneckii. FEMS Microbiol. Lett. 2015, 362, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Gostinčar, C.; Lenassi, M.; Gunde-Cimerman, N.; Plemenitaš, A. Fungal Adaptation to Extremely High Salt Concentrations. Int. Rev. Cytol. 2011, 77, 71–96. [Google Scholar] [CrossRef]
- Vaupotic, T.; Veranic, P.; Jenoe, P.; Plemenitaš, A. Mitochondrial mediation of environmental osmolytes discrimination during osmoadaptation in the extremely halotolerant black yeast Hortaea werneckii. Fungal Genet. Biol. 2008, 45, 994–1007. [Google Scholar] [CrossRef]
- Jančič, S.; Nguyen, H.D.T.; Frisvad, J.; Zalar, P.; Schroers, H.-J.; Seifert, K.A.; Gunde-Cimerman, N. A Taxonomic Revision of the Wallemia sebi Species Complex. PLOS ONE 2015, 10, e0125933. [Google Scholar] [CrossRef] [Green Version]
- Jančič, S.; Zalar, P.; Kocev, D.; Schroers, H.-J.; Džeroski, S.; Gunde-Cimerman, N. Halophily reloaded: New insights into the extremophilic life-style of Wallemia with the description of Wallemia hederae sp. nov. Fungal Divers. 2016, 76, 97–118. [Google Scholar] [CrossRef] [Green Version]
- Zajc, J.; Liu, Y.; Dai, W.; Yang, Z.; Hu, J.; Gostinčar, C.; Gunde-Cimerman, N. Genome and transcriptome sequencing of the halophilic fungus Wallemia ichthyophaga: Haloadaptations present and absent. BMC Genom. 2013, 14, 617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madern, D.; Ebel, C.; Zaccai, G. Halophilic adaptation of enzymes. Extremophiles 2000, 4, 91–98. [Google Scholar] [CrossRef]
- Konte, T.; Plemenitaš, A. The HOG signal transduction pathway in the halophilic fungus Wallemia ichthyophaga: Identification and characterisation of MAP kinases WiHog1A and WiHog1B. Extremophiles 2013, 17, 623–636. [Google Scholar] [CrossRef] [PubMed]
- Konte, T.; Terpitz, U.; Plemenitaš, A. Reconstruction of the High-Osmolarity Glycerol (HOG) Signaling Pathway from the Halophilic Fungus Wallemia ichthyophaga in Saccharomyces cerevisiae. Front. Microbiol. 2016, 7, 901. [Google Scholar] [CrossRef]
- Meena, N.; Kaur, H.; Mondal, A.K. Interactions among HAMP Domain Repeats Act as an Osmosensing Molecular Switch in Group III Hybrid Histidine Kinases from Fungi. J. Biol. Chem. 2010, 285, 12121–12132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srikantha, T.; Tsai, L.; Daniels, K.; Enger, L.; Highley, K.; Soll, D.R. The two-component hybrid kinase regulator CaNIKl of Candida albicans. Microbiology 1998, 144, 2715–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, K.; Randhawa, A.; Kaur, H.; Mondal, A.K.; Hohmann, S. Fungal fludioxonil sensitivity is diminished by a constitutively active form of the group III histidine kinase. FEBS Lett. 2012, 586, 2417–2422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krantz, M.; Becit, E.; Hohmann, S. Comparative analysis of HOG pathway proteins to generate hypotheses for functional analysis. Curr. Genet. 2006, 49, 152–165. [Google Scholar] [CrossRef] [PubMed]
- A Marles, J.; Dahesh, S.; Haynes, J.; Andrews, B.J.; Davidson, A.R. Protein-Protein Interaction Affinity Plays a Crucial Role in Controlling the Sho1p-Mediated Signal Transduction Pathway in Yeast. Mol. Cell 2004, 14, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Zarrinpar, A.; Bhattacharyya, R.P.; Nittler, M.; A Lim, W. Sho1 and Pbs2 Act as Coscaffolds Linking Components in the Yeast High Osmolarity MAP Kinase Pathway. Mol. Cell 2004, 14, 825–832. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plemenitaš, A. Sensing and Responding to Hypersaline Conditions and the HOG Signal Transduction Pathway in Fungi Isolated from Hypersaline Environments: Hortaea werneckii and Wallemia ichthyophaga. J. Fungi 2021, 7, 988. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7110988
Plemenitaš A. Sensing and Responding to Hypersaline Conditions and the HOG Signal Transduction Pathway in Fungi Isolated from Hypersaline Environments: Hortaea werneckii and Wallemia ichthyophaga. Journal of Fungi. 2021; 7(11):988. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7110988
Chicago/Turabian StylePlemenitaš, Ana. 2021. "Sensing and Responding to Hypersaline Conditions and the HOG Signal Transduction Pathway in Fungi Isolated from Hypersaline Environments: Hortaea werneckii and Wallemia ichthyophaga" Journal of Fungi 7, no. 11: 988. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7110988