
Improved In Vitro Anti-Mucorales Activity and Cytotoxicity of Amphotericin B with a Pegylated Surfactant

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Media
2.3. Solutions
2.4. Checkerboard Assays
2.5. Checkerboard Data Modeling
2.6. Hemolytic Activity
2.7. Cytotoxicity
2.8. Determination of AmB Aggregation States
2.9. Statistics
3. Results
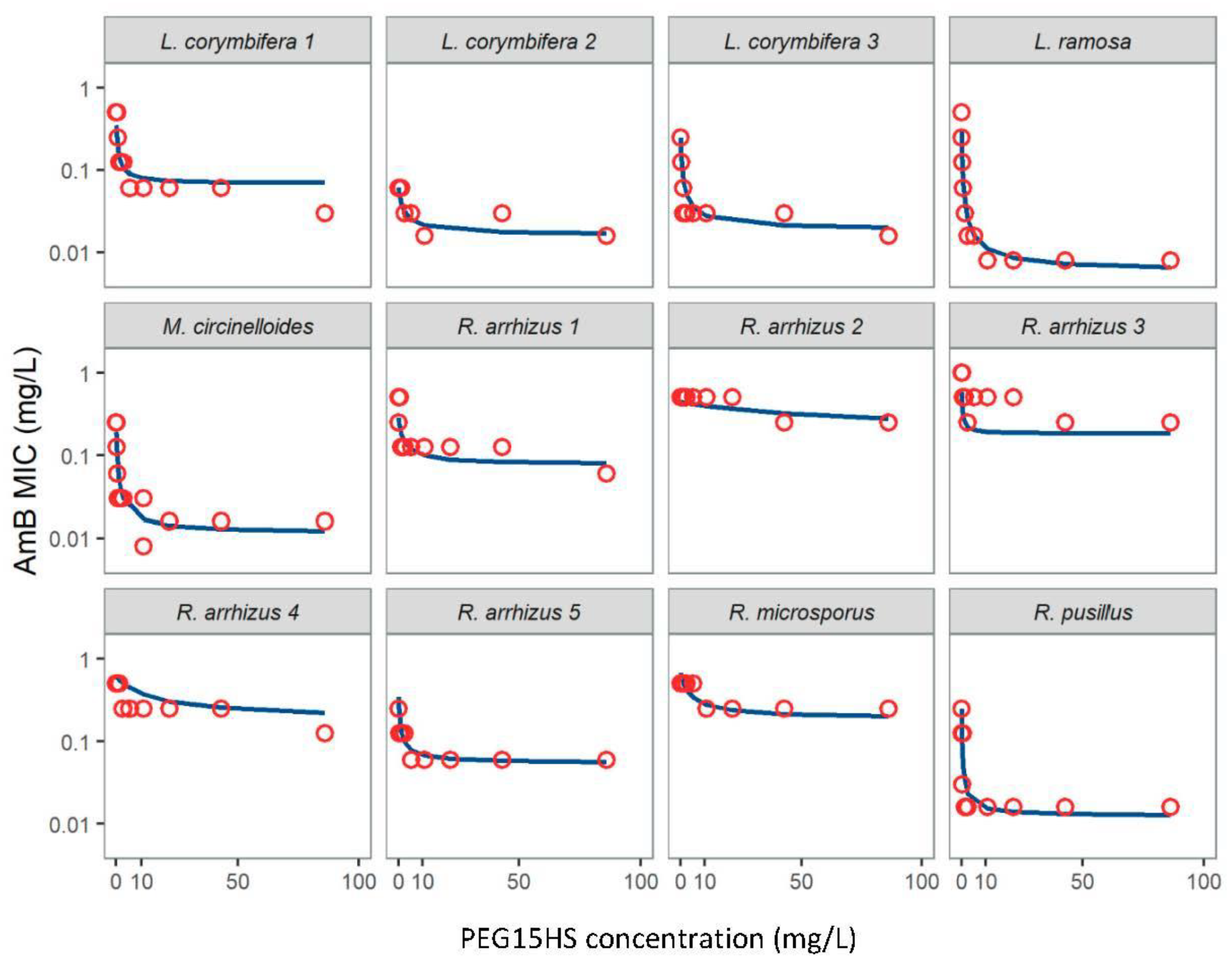
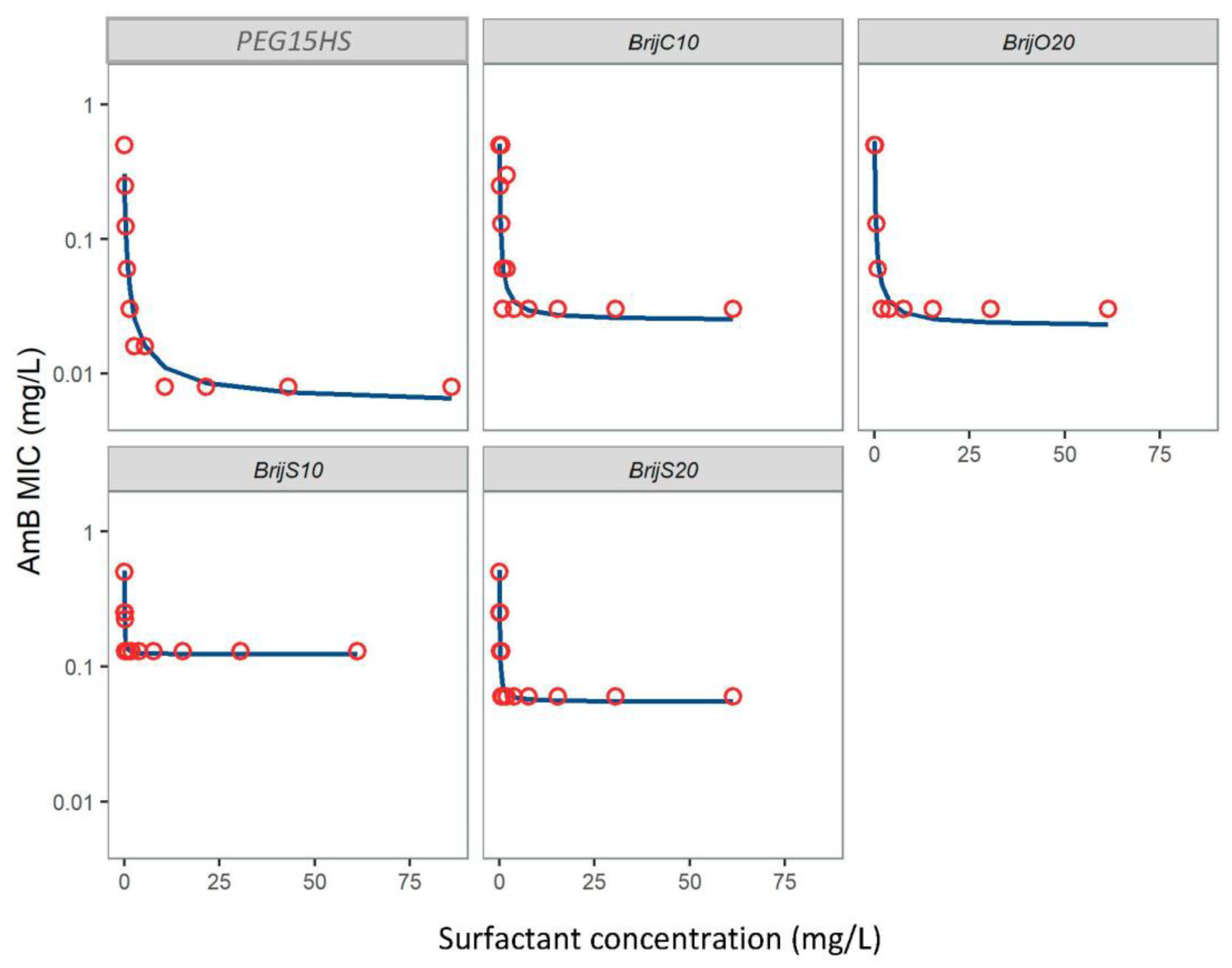
3.1. Nonionic Surfactants Enhanced AmB Activity against Various Mucorales Isolates
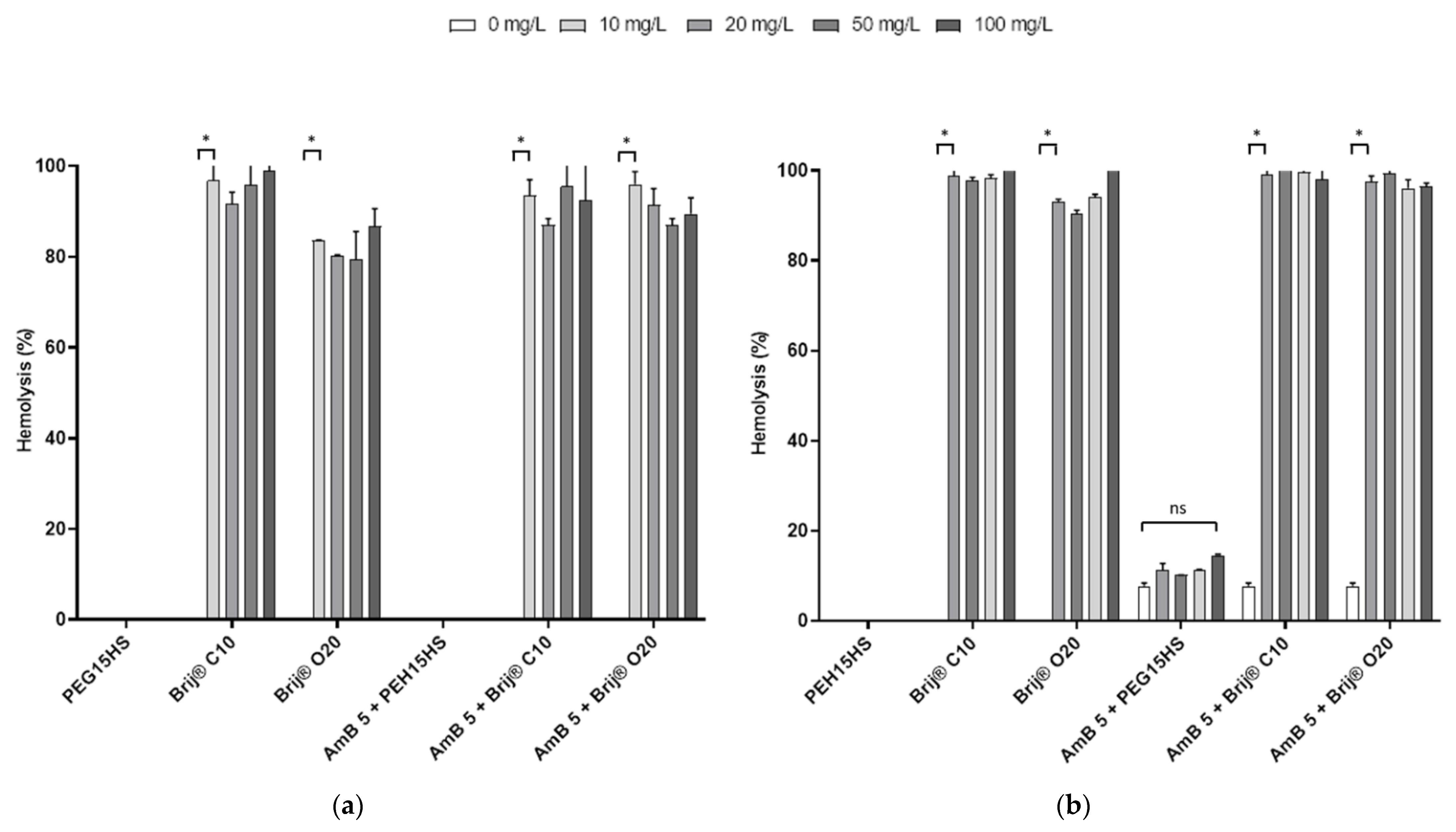
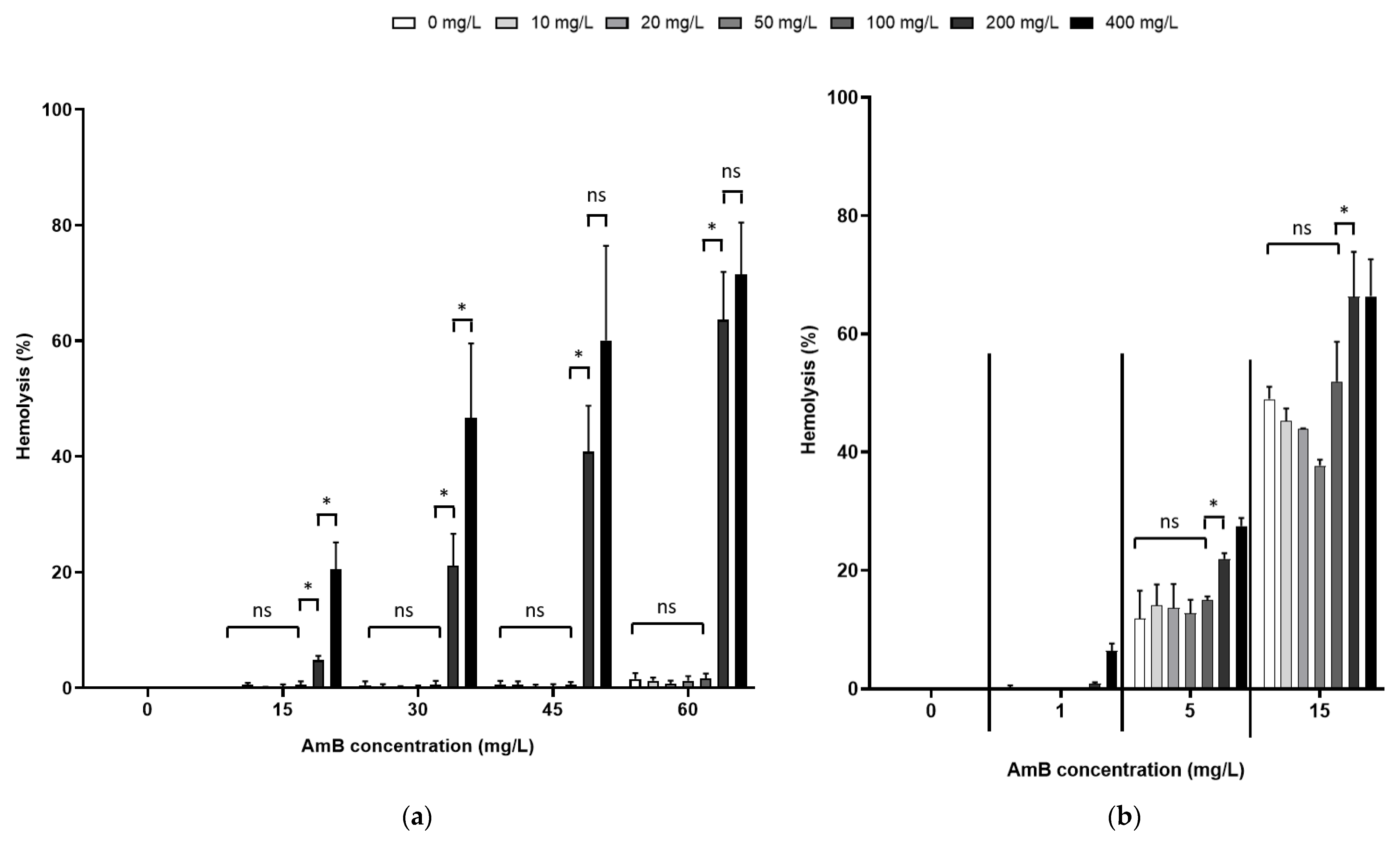
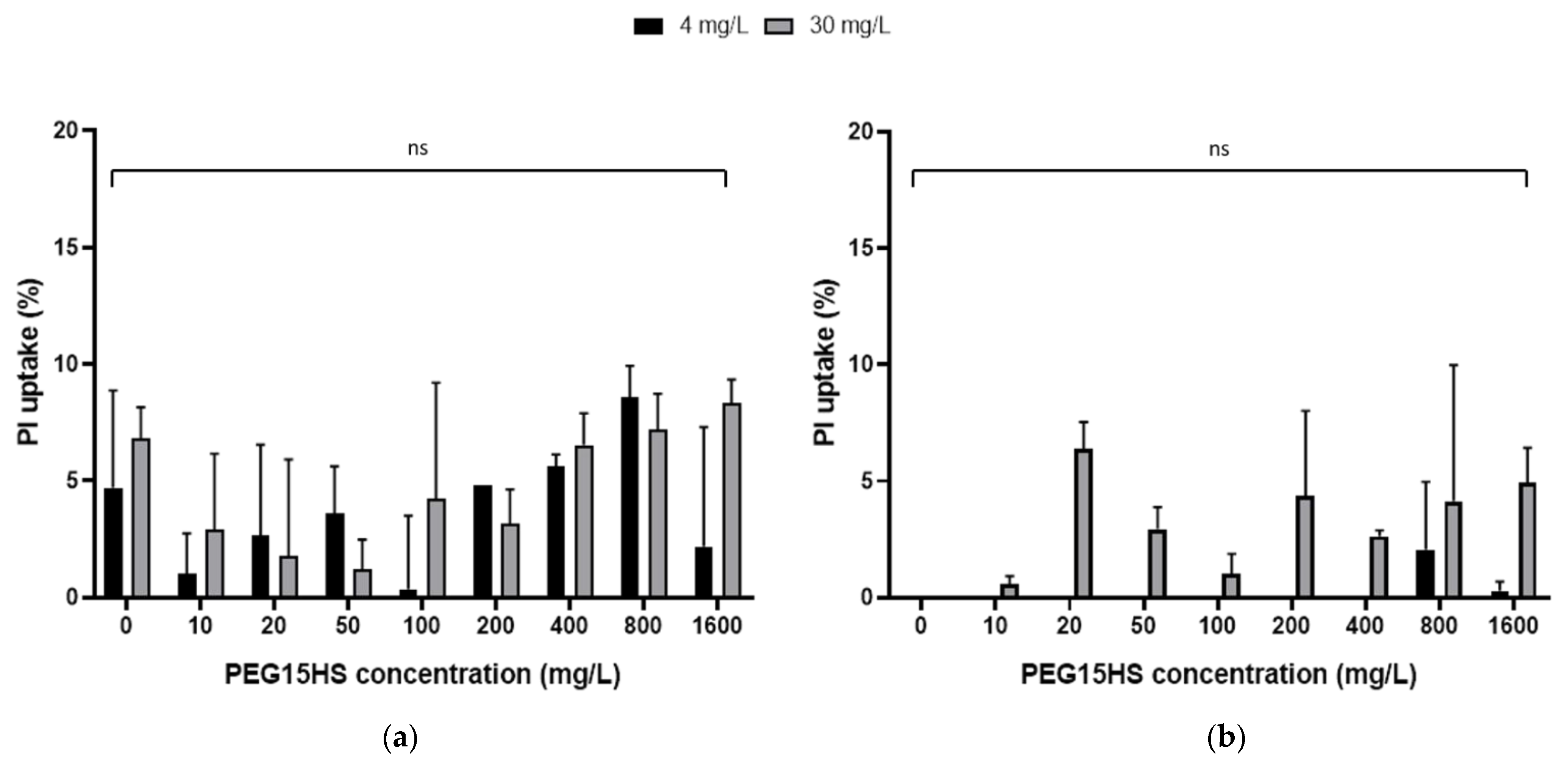
3.2. The Effect of AmB on the Integrity of Mammalian Cell Membranes Was Not Potentiated by PEG15HS at the Concentration Producing the Maximum Effect
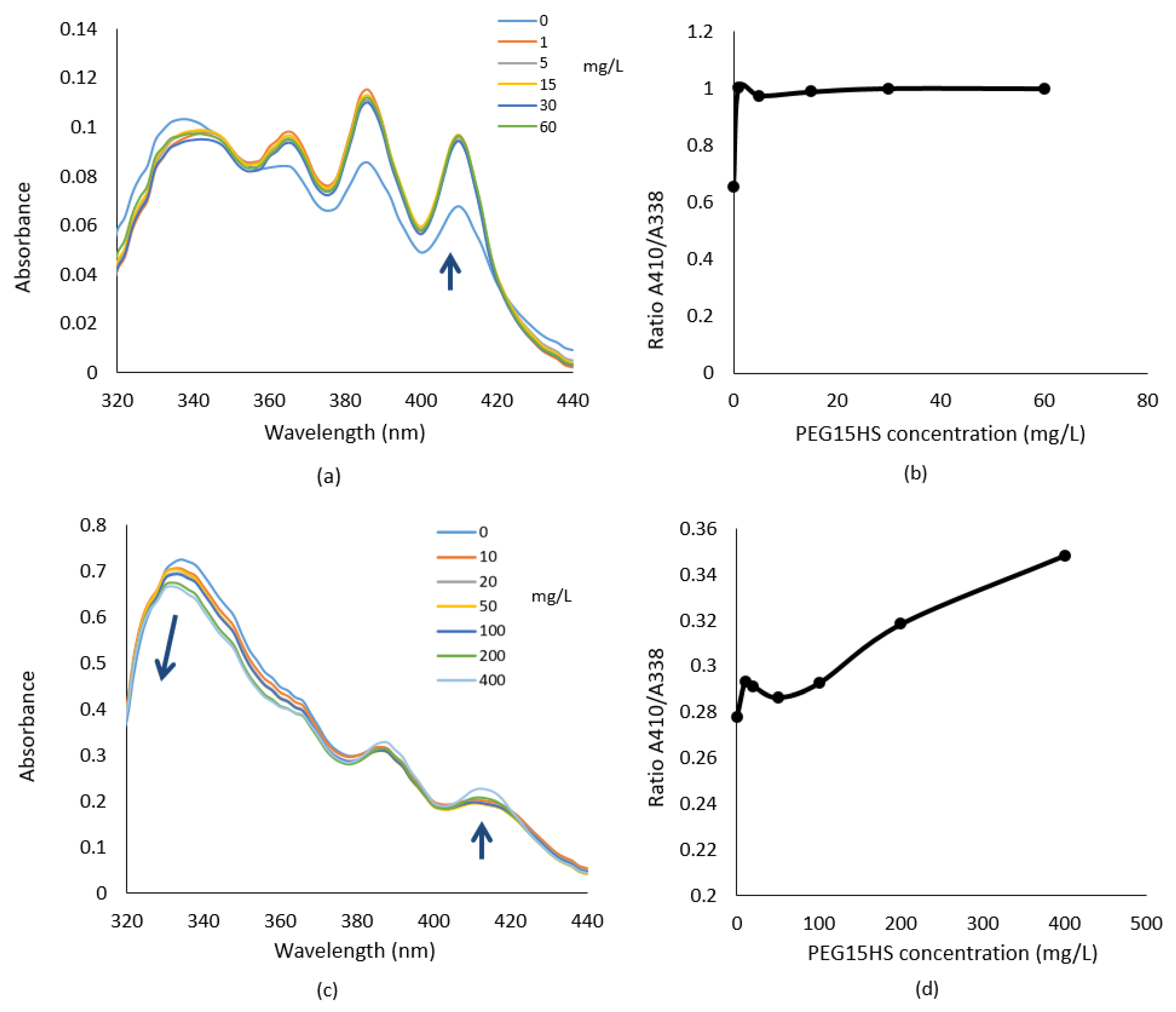
3.3. PEG15HS Alters Aggregation States of AmB
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farmakiotis, D.; Kontoyiannis, D.P. Mucormycoses. Infect. Dis. Clin. North Am. 2016, 30, 143–163. [Google Scholar] [CrossRef]
- Serris, A.; Danion, F.; Lanternier, F. Disease Entities in Mucormycosis. J. Fungi 2019, 5, 23. [Google Scholar] [CrossRef] [Green Version]
- Cornely, O.A.; Alastruey-Izquierdo, A.; Arenz, D.; Chen, S.C.A.; Dannaoui, E.; Hochhegger, B.; Hoenigl, M.; Jensen, H.E.; Lagrou, K.; Lewis, R.E.; et al. Global Guideline for the Diagnosis and Management of Mucormycosis: An Initiative of the European Confederation of Medical Mycology in Cooperation with the Mycoses Study Group Education and Research Consortium. Lancet Infect. Dis. 2019, 19, e405–e421. [Google Scholar] [CrossRef]
- Tissot, F.; Agrawal, S.; Pagano, L.; Petrikkos, G.; Groll, A.H.; Skiada, A.; Lass-Flörl, C.; Calandra, T.; Viscoli, C.; Herbrecht, R. ECIL-6 Guidelines for the Treatment of Invasive Candidiasis, Aspergillosis and Mucormycosis in Leukemia and Hematopoietic Stem Cell Transplant Patients. Haematologica 2017, 102, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Sipsas, N.V.; Gamaletsou, M.N.; Anastasopoulou, A.; Kontoyiannis, D.P. Therapy of Mucormycosis. J. Fungi 2018, 4, 90. [Google Scholar] [CrossRef] [Green Version]
- Abidi, M.Z.; Sohail, M.R.; Cummins, N.; Wilhelm, M.; Wengenack, N.; Brumble, L.; Shah, H.; Jane Hata, D.; McCullough, A.; Wendel, A.; et al. Stability in the Cumulative Incidence, Severity and Mortality of 101 Cases of Invasive Mucormycosis in High-Risk Patients from 1995 to 2011: A Comparison of Eras Immediately before and after the Availability of Voriconazole and Echinocandin-Amphotericin Combination Therapies. Mycoses 2014, 57, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Kyvernitakis, A.; Torres, H.A.; Jiang, Y.; Chamilos, G.; Lewis, R.E.; Kontoyiannis, D.P. Initial Use of Combination Treatment Does Not Impact Survival of 106 Patients with Haematologic Malignancies and Mucormycosis: A Propensity Score Analysis. Clin. Microbiol. Infect. 2016, 22, 811.e1–811.e8. [Google Scholar] [CrossRef] [Green Version]
- van Burik, J.-A.H.; Hare, R.S.; Solomon, H.F.; Corrado, M.L.; Kontoyiannis, D.P. Posaconazole Is Effective as Salvage Therapy in Zygomycosis: A Retrospective Summary of 91 Cases. Clin. Infect. Dis. 2006, 42, e61–e65. [Google Scholar] [CrossRef] [Green Version]
- Lanternier, F.; Dannaoui, E.; Morizot, G.; Elie, C.; Garcia-Hermoso, D.; Huerre, M.; Bitar, D.; Dromer, F.; Lortholary, O.; French Mycosis Study Group. A Global Analysis of Mucormycosis in France: The RetroZygo Study (2005–2007). Clin. Infect. Dis. 2012, 54 (Suppl. 1), S35–S43. [Google Scholar] [CrossRef] [Green Version]
- Reed, C.; Bryant, R.; Ibrahim, A.S.; Edwards, J., Jr.; Filler, S.G.; Goldberg, R.; Spellberg, B. Combination Polyene-Caspofungin Treatment of Rhino-Orbital-Cerebral Mucormycosis. Clin. Infect. Dis. 2008, 47, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Brunet, K.; Rammaert, B. Mucormycosis Treatment: Recommendations, Latest Advances, and Perspectives. J. Mycol. Med. 2020, 30, 101007. [Google Scholar] [CrossRef]
- Schwarz, P.; Cornely, O.A.; Dannaoui, E. Antifungal Combinations in Mucorales: A Microbiological Perspective. Mycoses 2019, 62, 746–760. [Google Scholar] [CrossRef]
- Kamiński, D.M. Recent Progress in the Study of the Interactions of Amphotericin B with Cholesterol and Ergosterol in Lipid Environments. Eur. Biophys. J. 2014, 43, 453–467. [Google Scholar] [CrossRef] [Green Version]
- Gray, K.C.; Palacios, D.S.; Dailey, I.; Endo, M.M.; Uno, B.E.; Wilcock, B.C.; Burke, M.D. Amphotericin Primarily Kills Yeast by Simply Binding Ergosterol. Proc. Natl. Acad. Sci. USA 2012, 109, 2234–2239. [Google Scholar] [CrossRef] [Green Version]
- Adler-Moore, J.P.; Gangneux, J.-P.; Pappas, P.G. Comparison between Liposomal Formulations of Amphotericin B. Med. Mycol. 2016, 54, 223–231. [Google Scholar] [CrossRef]
- Espada, R.; Valdespina, S.; Alfonso, C.; Rivas, G.; Ballesteros, M.P.; Torrado, J.J. Effect of Aggregation State on the Toxicity of Different Amphotericin B Preparations. Int. J. Pharm. 2008, 361, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Barwicz, J.; Christian, S.; Gruda, I. Effects of the Aggregation State of Amphotericin B on Its Toxicity to Mice. Antimicrob. Agents Chemother. 1992, 36, 2310–2315. [Google Scholar] [CrossRef] [Green Version]
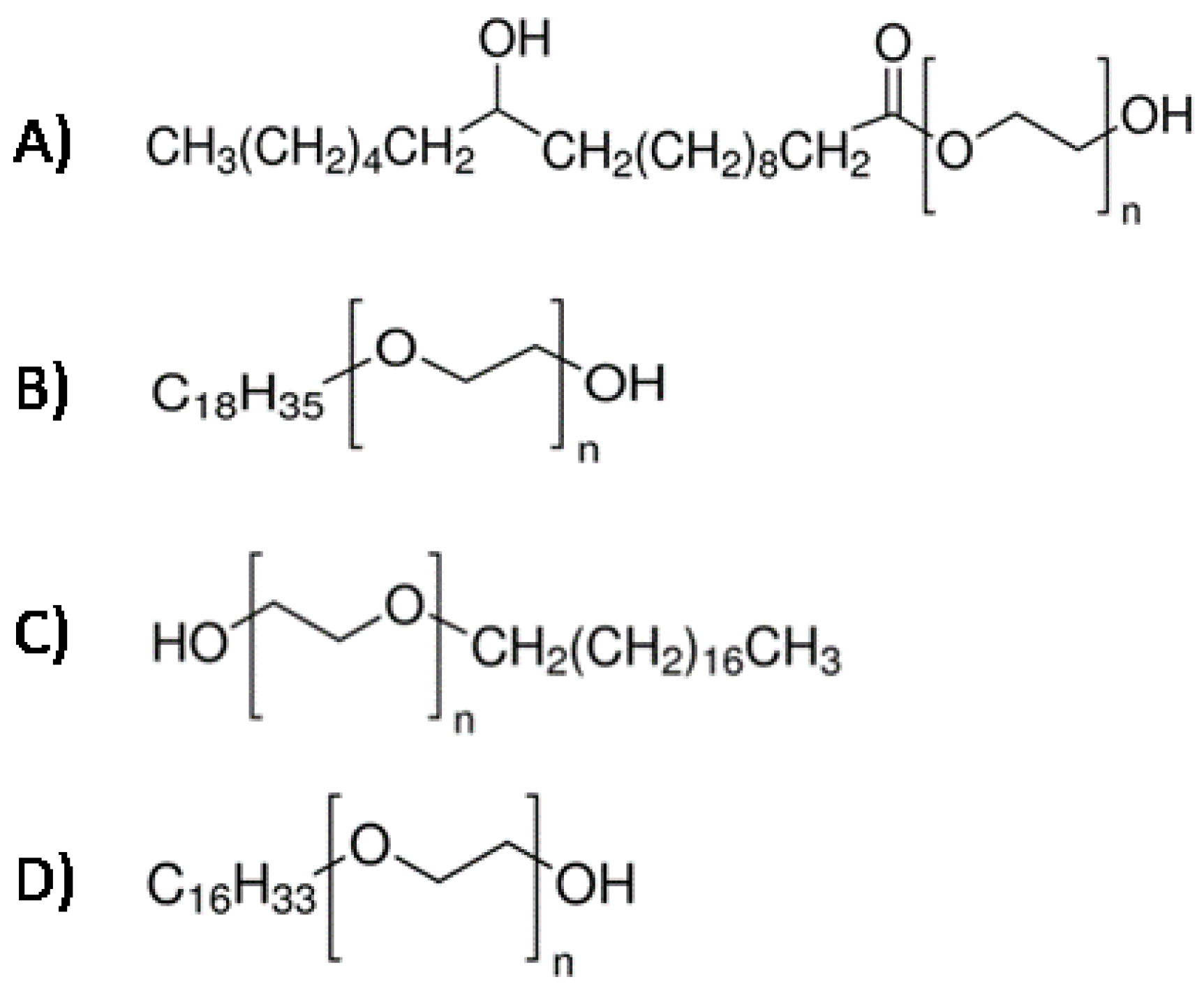
- Tewes, F.; Corrigan, O.I.; Healy, A.M. Surfactants in Pharmaceutical Products and Systems. In Encyclopedia of Pharmaceutical Science and Technology, 4th ed.; CRC Press: Boca Raton, FL, USA, 2015; ISBN 978-1-351-12487-4. [Google Scholar]
- Jiao, J. Polyoxyethylated Nonionic Surfactants and Their Applications in Topical Ocular Drug Delivery. Adv. Drug Deliv. Rev. 2008, 60, 1663–1673. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing. EUCAST DEFINITIVE DOCUMENT E.DEF 9.3.2. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/AFST/Files/EUCAST_E_Def_9.3.2_Mould_testing_definitive_revised_2020.pdf (accessed on 16 January 2022).
- Chauzy, A.; Buyck, J.; de Jonge, B.L.M.; Marchand, S.; Grégoire, N.; Couet, W. Pharmacodynamic Modelling of β-Lactam/β-Lactamase Inhibitor Checkerboard Data: Illustration with Aztreonam-Avibactam. Clin. Microbiol. Infect. 2019, 25, 515.e1–515.e4. [Google Scholar] [CrossRef] [Green Version]
- Serrano, D.R.; Hernández, L.; Fleire, L.; González-Alvarez, I.; Montoya, A.; Ballesteros, M.P.; Dea-Ayuela, M.A.; Miró, G.; Bolás-Fernández, F.; Torrado, J.J. Hemolytic and Pharmacokinetic Studies of Liposomal and Particulate Amphotericin B Formulations. Int. J. Pharm. 2013, 447, 38–46. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, L.; Wang, H.; Lv, X.; Ren, Q.; Zheng, G. The Effects of Legumain in THP1 Leukemia Cells. Biocell 2021, 45, 149–156. [Google Scholar] [CrossRef]
- Lankoff, A.; Sandberg, W.J.; Wegierek-Ciuk, A.; Lisowska, H.; Refsnes, M.; Sartowska, B.; Schwarze, P.E.; Męczyńska-Wielgosz, S.; Wojewódzka, M.; Kruszewski, M. The Effect of Agglomeration State of Silver and Titanium Dioxide Nanoparticles on Cellular Response of HepG2, A549 and THP-1 Cells. Toxicol. Lett. 2012, 208, 197–213. [Google Scholar] [CrossRef]
- Gagoś, M.; Arczewska, M. Influence of K+ and Na+ Ions on the Aggregation Processes of Antibiotic Amphotericin B: Electronic Absorption and FTIR Spectroscopic Studies. J. Phys. Chem. B 2011, 115, 3185–3192. [Google Scholar] [CrossRef]
- Gruda, I.; Dussault, N. Effect of the Aggregation State of Amphotericin B on Its Interaction with Ergosterol. Biochem. Cell Biol. 1988, 66, 177–183. [Google Scholar] [CrossRef]
- Gagoś, M.; Hereć, M.; Arczewska, M.; Czernel, G.; Dalla Serra, M.; Gruszecki, W.I. Anomalously High Aggregation Level of the Polyene Antibiotic Amphotericin B in Acidic Medium: Implications for the Biological Action. Biophys. Chem. 2008, 136, 44–49. [Google Scholar] [CrossRef] [Green Version]
- White, R.L.; Burgess, D.S.; Manduru, M.; Bosso, J.A. Comparison of Three Different in Vitro Methods of Detecting Synergy: Time-Kill, Checkerboard, and E Test. Antimicrob. Agents Chemother. 1996, 40, 1914–1918. [Google Scholar] [CrossRef] [Green Version]
- Bidaud, A.L.; Botterel, F.; Chowdhary, A.; Dannaoui, E. In Vitro Antifungal Combination of Flucytosine with Amphotericin B, Voriconazole, or Micafungin against Candida Auris Shows No Antagonism. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef]
- Schneider, E.K.; Reyes-Ortega, F.; Velkov, T.; Li, J. Antibiotic-Non-Antibiotic Combinations for Combating Extremely Drug-Resistant Gram-Negative “Superbugs”. Essays Biochem. 2017, 61, 115–125. [Google Scholar] [CrossRef]
- Shaukat Ali Kolliphor® HS 15—An Enabler for Parenteral and Oral Formulations. Available online: http://www.americanpharmaceuticalreview.com/Featured-Articles/358749-Kolliphor-HS-15-An-Enabler-for-Parenteral-and-Oral-Formulations/ (accessed on 10 December 2021).
- U.S. Food and Drug Administration. Inactive Ingredient Search for Approved Drug Products. Available online: https://www.accessdata.fda.gov/scripts/cder/iig/index.cfm (accessed on 21 August 2020).
- Younes, N.F.; Abdel-Halim, S.A.; Elassasy, A.I. Solutol HS15 Based Binary Mixed Micelles with Penetration Enhancers for Augmented Corneal Delivery of Sertaconazole Nitrate: Optimization, in Vitro, Ex Vivo and in Vivo Characterization. Drug Deliv. 2018, 25, 1706–1717. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Zhang, F.; Lan, J.; Sun, F.; Li, J.; Li, M.; Song, K.; Wu, X. Ultra-Small Micelles Based on Polyoxyl 15 Hydroxystearate for Ocular Delivery of Myricetin: Optimization, in Vitro, and in Vivo Evaluation. Drug Deliv. 2019, 26, 158–167. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Zhang, Z.; Jia, X.; Song, J. D-α-Tocopheryl Polyethylene Glycol Succinate/Solutol HS 15 Mixed Micelles for the Delivery of Baohuoside I against Non-Small-Cell Lung Cancer: Optimization and in Vitro, in Vivo Evaluation. Int. J. Nanomed. 2016, 11, 4563–4571. [Google Scholar] [CrossRef]
- Coon, J.S.; Knudson, W.; Clodfelter, K.; Lu, B.; Weinstein, R.S. Solutol HS 15, Nontoxic Polyoxyethylene Esters of 12-Hydroxystearic Acid, Reverses Multidrug Resistance. Cancer Res. 1991, 51, 897–902. [Google Scholar]
- Weete, J.D.; Gandhi, S.R. Sterols of the Phylum Zygomycota: Phylogenetic Implications. Lipids 1997, 32, 1309–1316. [Google Scholar] [CrossRef]
- Weete, J.D.; Abril, M.; Blackwell, M. Phylogenetic Distribution of Fungal Sterols. PLoS ONE 2010, 5, e10899. [Google Scholar] [CrossRef] [Green Version]
- Müller, C.; Neugebauer, T.; Zill, P.; Lass-Flörl, C.; Bracher, F.; Binder, U. Sterol Composition of Clinically Relevant Mucorales and Changes Resulting from Posaconazole Treatment. Molecules 2018, 23, 1218. [Google Scholar] [CrossRef] [Green Version]
- Weete, J.D.; Lawler, G.C.; Laseter, J.L. Total Lipid and Sterol Components of Rhizopus Arrhizus: Identification and Metabolism. Arch. Biochem. Biophys. 1973, 155, 411–419. [Google Scholar] [CrossRef]
- Dannaoui, E.; Afeltra, J.; Meis, J.F.G.M.; Verweij, P.E.; Eurofung Network. In Vitro Susceptibilities of Zygomycetes to Combinations of Antimicrobial Agents. Antimicrob. Agents Chemother. 2002, 46, 2708–2711. [Google Scholar] [CrossRef] [Green Version]
- van der Linden, J.W.M.; Snelders, E.; Kampinga, G.A.; Rijnders, B.J.A.; Mattsson, E.; Debets-Ossenkopp, Y.J.; Kuijper, E.J.; Van Tiel, F.H.; Melchers, W.J.G.; Verweij, P.E. Clinical Implications of Azole Resistance in Aspergillus Fumigatus, The Netherlands, 2007–2009. Emerg. Infect. Dis. 2011, 17, 1846–1854. [Google Scholar] [CrossRef]
- Hamill, R.J. Amphotericin B Formulations: A Comparative Review of Efficacy and Toxicity. Drugs 2013, 73, 919–934. [Google Scholar] [CrossRef]
- Meyer, H.W.; Richter, W.; Winkelmann, H. Nystatin- and Amphotericin B-Induced Structural Alterations of the Erythrocyte Membrane: Importance of Reduced Ionic Strength. Exp. Pathol. 1983, 24, 163–166. [Google Scholar] [CrossRef]
- Tancrède, P.; Barwicz, J.; Jutras, S.; Gruda, I. The Effect of Surfactants on the Aggregation State of Amphotericin B. Biochim. Biophys. Acta 1990, 1030, 289–295. [Google Scholar] [CrossRef]
- Huang, W.; Zhang, Z.; Han, X.; Tang, J.; Wang, J.; Dong, S.; Wang, E. Ion Channel Behavior of Amphotericin B in Sterol-Free and Cholesterol- or Ergosterol-Containing Supported Phosphatidylcholine Bilayer Model Membranes Investigated by Electrochemistry and Spectroscopy. Biophys. J. 2002, 83, 3245–3255. [Google Scholar] [CrossRef] [Green Version]
- Golenser, J.; Domb, A. New Formulations and Derivatives of Amphotericin B for Treatment of Leishmaniasis. Mini Rev. Med. Chem. 2006, 6, 153–162. [Google Scholar] [CrossRef]
- Chéron, M.; Petit, C.; Bolard, J.; Gaboriau, F. Heat-Induced Reformulation of Amphotericin B-Deoxycholate Favours Drug Uptake by the Macrophage-like Cell Line J774. J. Antimicrob. Chemother. 2003, 52, 904–910. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Surfactant | Isolates | MIC0 (mg/L) | EC50 (mg/L) | MIC∞ (mg/L) | Rmax (MIC0/MIC∞) |
|---|---|---|---|---|---|
| PEG15HS | L. corymbifera 1 | 0.36 | 0.44 | 0.068 | 5.2 |
| L. corymbifera 2 | 0.06 | 1.33 | 0.016 | 3.8 | |
| L. corymbifera 3 | 0.25 | 0.42 | 0.019 | 13.0 | |
| L. ramosa | 0.51 | 0.19 | 0.008 | 63.8 | |
| M. circinelloides | 0.19 | 0.32 | 0.012 | 16.5 | |
| R. arrhizus 1 | 0.28 | 1.34 | 0.077 | 3.7 | |
| R. arrhizus 2 | 0.44 | 53.45 | 0.177 | 2.5 | |
| R. arrhizus 3 | 0.59 | 0.28 | 0.183 | 3.2 | |
| R. arrhizus 4 | 0.61 | 8.57 | 0.186 | 3.3 | |
| R. arrhizus 5 | 0.35 | 0.47 | 0.055 | 6.4 | |
| R. microsporus | 0.68 | 2.32 | 0.190 | 3.6 | |
| R. pusillus | 0.25 | 0.13 | 0.012 | 19.9 | |
| Brij® S10 | L. ramosa | 0.51 | 0.02 | 0.123 | 4.1 |
| Brij® S20 | L. ramosa | 0.51 | 0.02 | 0.056 | 9.2 |
| Brij® O20 | L. ramosa | 0.51 | 0.06 | 0.022 | 23.2 |
| Brij® C10 | L. ramosa | 0.51 | 0.05 | 0.025 | 20.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brunet, K.; Diop, C.A.B.; Chauzy, A.; Prébonnaud, N.; Marchand, S.; Rammaert, B.; Tewes, F. Improved In Vitro Anti-Mucorales Activity and Cytotoxicity of Amphotericin B with a Pegylated Surfactant. J. Fungi 2022, 8, 121. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8020121
Brunet K, Diop CAB, Chauzy A, Prébonnaud N, Marchand S, Rammaert B, Tewes F. Improved In Vitro Anti-Mucorales Activity and Cytotoxicity of Amphotericin B with a Pegylated Surfactant. Journal of Fungi. 2022; 8(2):121. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8020121
Chicago/Turabian StyleBrunet, Kévin, Cheikh A. B. Diop, Alexia Chauzy, Noémie Prébonnaud, Sandrine Marchand, Blandine Rammaert, and Frédéric Tewes. 2022. "Improved In Vitro Anti-Mucorales Activity and Cytotoxicity of Amphotericin B with a Pegylated Surfactant" Journal of Fungi 8, no. 2: 121. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8020121