
Diversity of Colletotrichum Species Causing Apple Bitter Rot and Glomerella Leaf Spot in China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolates
2.2. DNA Extraction and PCR Amplification
2.3. Sequence Alignment and Phylogenetic Analysis
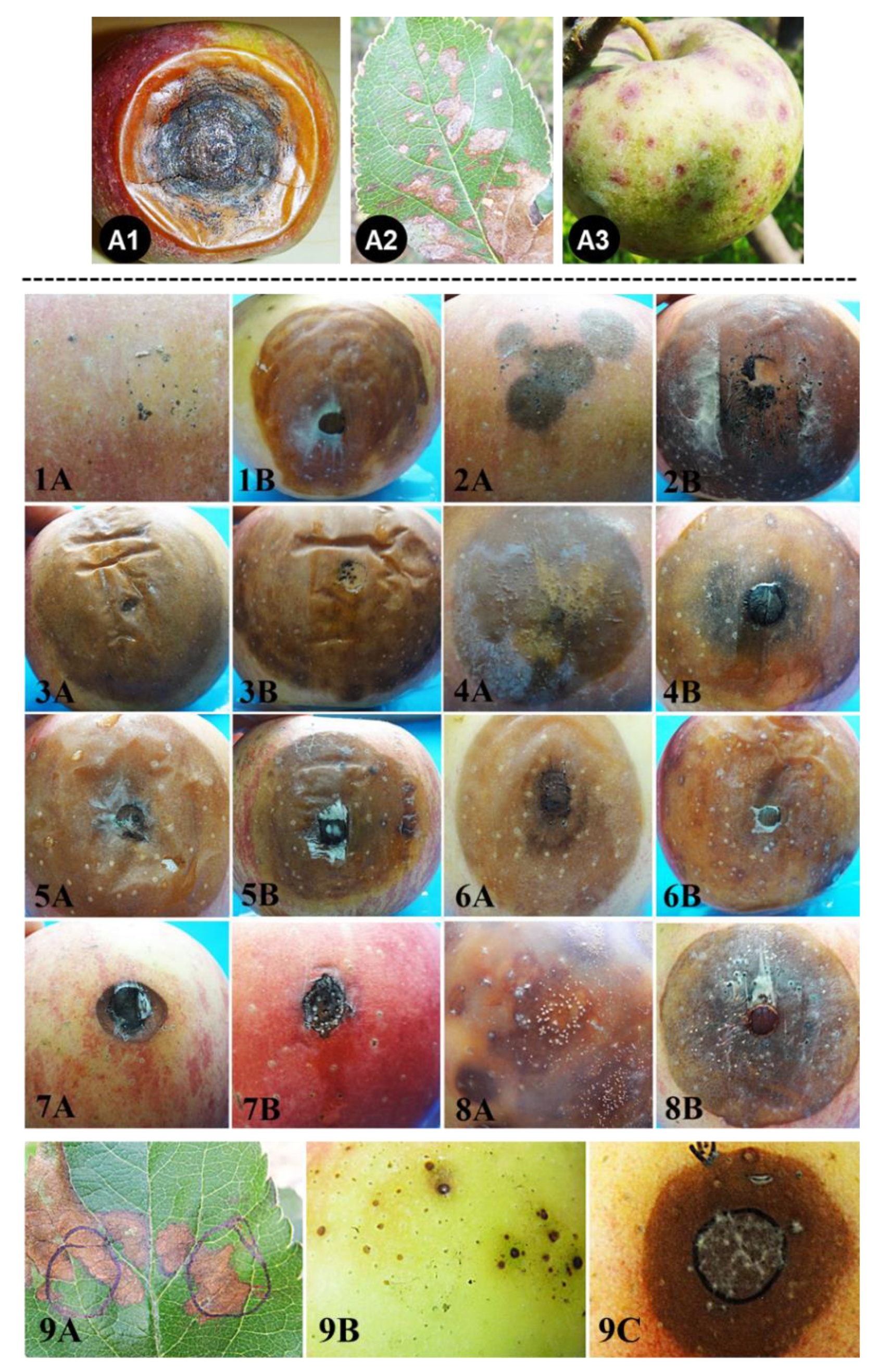
2.4. Pathogenicity Tests

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Type Strain | Host | County | GenBank No. | ||||
|---|---|---|---|---|---|---|---|---|
| ITS | ACT | GAPDH | CHS-1 | TUB2 | ||||
| C. acerbum | CBS 128530 1 | Malus domestica | New Zealand | JQ948459 | JQ949780 | JQ948790 | JQ949120 | JQ950110 |
| C. acutatum | CBS 112996 1 | Carica papaya | Australia | JQ005776 | JQ005839 | JQ948677 | JQ005797 | JQ005860 |
| CBS 126521 2 | Anemonehybride | Netherlands | JQ948366 | JQ949687 | JQ948697 | JQ949027 | JQ950017 | |
| C. aenigma | ICMP 18608 1 | Persea americana | Israel | JX010244 | JX009443 | JX010044 | JX009774 | JX010389 |
| ICMP 18686 2 | Pyrus pyrifolia | Japan | JX010243 | JX009519 | JX009913 | JX009789 | JX010390 | |
| F12PGXY03 | Malus domestica | China | KF772117 | KF772027 | KF772087 | KF772057 | KF772147 | |
| F12PGXY04 | Malus domestica | China | KF772118 | KF772028 | KF772088 | KF772058 | KF772148 | |
| W12PGYXY15 | Malus domestica | China | KF791590 | KF791569 | KF791583 | KF791576 | KF791597 | |
| C. aeschynomenes | ICMP 17673 1 | Aeschynomene sp. | USA | JX010176 | JX009483 | JX009930 | JX009799 | JX010392 |
| C. alienum | ICMP 12071 1 | Malus domestica | New Zealand | JX010251 | JX009572 | JX010028 | JX009882 | JX010411 |
| ICMP 18621 3 | Persea americana | New Zealand | JX010246 | JX009552 | JX009959 | JX009755 | JX010386 | |
| F11PGZH02 | Malus domestica | China | KF772119 | KF772029 | KF772089 | KF772059 | KF772149 | |
| C. asianum | ICMP 18580 1 | Coffea arabica | Thailand | FJ972612 | JX009584 | JX010053 | JX009867 | JX010406 |
| ICMP 18696 3 | Mangifera indica | Australia | JX010192 | JX009576 | JX009915 | JX009753 | JX010384 | |
| C. boninense | CBS 123755 1 | Crinum asiaticum | Japan | JQ005153 | JQ005501 | JQ005240 | JQ005327 | JQ005588 |
| C. cuscutae | IMI 304802 1 | Cuscuta sp. | Dominica | JQ948195 | JQ949516 | JQ948525 | JQ948856 | JQ949846 |
| C. fioriniae | CBS 128517 1 | Fiorinia externa | USA | JQ948292 | JQ949613 | JQ948622 | JQ948953 | JQ949943 |
| CBS 125396 2 | Malus domestica | USA | JQ948299 | JQ949620 | JQ948629 | JQ948960 | JQ949950 | |
| CBS 128517 1 | Fiorinia externa | USA | JQ948292 | JQ949613 | JQ948622 | JQ948953 | JQ949943 | |
| CBS 125396 2 | Malus domestica | USA | JQ948299 | JQ949620 | JQ948629 | JQ948960 | JQ949950 | |
| CBS 363003 2 | Camellia reticulata | China | JQ948339 | JQ949660 | JQ948669 | JQ949000 | JQ949990 | |
| ATCC 28992 2 | Malus domestica | USA | JQ948297 | JQ949618 | JQ948627 | JQ948958 | JQ949948 | |
| CBS 129938 2 | Malus domestica | USA | JQ948296 | JQ949617 | JQ948626 | JQ948957 | JQ949947 | |
| CBS 129948 2 | Tulipa sp. | UK | JQ948344 | JQ949665 | JQ948674 | JQ949005 | JQ949995 | |
| IMI 324996 2 | Malus pumila | USA | JQ948301 | JQ949622 | JQ948631 | JQ948962 | JQ949952 | |
| ATCC 12097 2 | Rhododendron sp. | USA | JQ948307 | JQ949628 | JQ948637 | JQ948968 | JQ949958 | |
| CBS 200.35 2 | Rubus sp. | USA | JQ948293 | JQ949614 | JQ948623 | JQ948954 | JQ949944 | |
| CBS 490.92 2 | Solanum lycopersicum | New Zealand | JQ948326 | JQ949647 | JQ948656 | JQ948987 | JQ949977 | |
| CBS 119293 2 | Vaccinium corymbosum (blueberry) | New Zealand | JQ948314 | JQ949635 | JQ948644 | JQ948975 | JQ949965 | |
| C. fructicola | CBS 130416 1 | Coffea arabica | Thailand | JX010165 | FJ907426 | JX010033 | JX009866 | JX010405 |
| F12PGSQ01 | Malus domestica | China | KF772124 | KF772034 | KF772094 | KF772064 | KF772154 | |
| F12PGSQ05 | Malus domestica | China | KF772125 | KF772035 | KF772095 | KF772065 | KF772155 | |
| F12PGXY01 | Malus ×domestica | China | KF772126 | KF772036 | KF772096 | KF772066 | KF772156 | |
| W12PGYSQ06 | M. ×domestica | China | KF791591 | KF791570 | KF791584 | KF791577 | KF791598 | |
| F10PGCJJ1 | M. ×domestica | China | KF772128 | KF772038 | KF772098 | KF772068 | KF772158 | |
| F10PGCJJ3 | M. ×domestica | China | KF772129 | KF772039 | KF772099 | KF772069 | KF772159 | |
| F10PGHLD1 | M. ×domestica | China | KF772130 | KF772040 | KF772100 | KF772070 | KF772160 | |
| F11PGYT02 | M. ×domestica | China | KF772131 | KF772041 | KF772101 | KF772071 | KF772161 | |
| F11PGYT04 | M. ×domestica | China | KF772132 | KF772042 | KF772102 | KF772072 | KF772162 | |
| C. gloeosporioides | CBS 112999 1 | Citrus sinensis | Italy | JX010152 | JX009531 | JX010056 | JX009818 | JX010445 |
| CBS 119204 3 | Pueraria lobata | USA | JX010150 | JX009502 | JX010013 | JX009790 | GQ849434 | |
| F11PGQX17 | M. ×domestica | China | KF772111 | KF772021 | KF772081 | KF772051 | KF772141 | |
| F12PGDL01 | M. ×domestica | China | KF772112 | KF772022 | KF772082 | KF772052 | KF772142 | |
| F12PGLQ30 | M. ××domestica | China | KF772113 | KF772023 | KF772083 | KF772053 | KF772143 | |
| F12PGLQ33 | M. ×domestica | China | KF772114 | KF772024 | KF772084 | KF772054 | KF772144 | |
| F12PGLQ34 | M. ×domestica | China | KF772115 | KF772025 | KF772085 | KF772055 | KF772145 | |
| F11PGZH23 | M. ×domestica | China | KF772116 | KF772026 | KF772086 | KF772056 | KF772146 | |
| C. godetiae | CBS 133.44 1 | Clarkia hybrida | Denmark | JQ948402 | JQ949723 | JQ948733 | JQ949063 | JQ950053 |
| CBS 198.53 2 | M. sylvestris | Netherlands | JQ948432 | JQ949753 | JQ948763 | JQ949093 | JQ950083 | |
| C. horii | ICPM 10492 1 | Diospyros kaki | Japan | GQ329690 | JX009438 | GQ329681 | JX009752 | JX010450 |
| C. hymenocallidis | CBS 125378 1 | Hymenocallis | China | JX010278 | GQ856775 | JX010019 | GQ856730 | JX010410 |
| C. kahawae subsp. kahawae | IMI 319418 1 | Coffea arabica | Kenya | JX010231 | JX009452 | JX010012 | JX009813 | JX010444 |
| C. karstii | CBS 132134 4 | Malus sp. | USA | JQ005181 | JQ005529 | JQ005268 | JQ005355 | JQ005615 |
| C. lupini | CBS 109225 1 | Lupinus albus | Ukraine | JQ948155 | JQ949476 | JQ948485 | JQ948816 | JQ949806 |
| C. musae | CBS 116870 1 | Musa sp. | USA | JX010146 | JX009433 | JX010050 | JX009896 | HQ596280 |
| ICMP 17817 3 | Musa sapientum | Kenya | JX010142 | JX009432 | JX010015 | JX009815 | JX010395 | |
| C. nymphaeae | CBS 515.78 1 | Nymphaea alba | Netherlands | JQ948197 | JQ949518 | JQ948527 | JQ948858 | JQ949848 |
| IMI 370491 2 | M. pumila | Brazil | JQ948204 | JQ949525 | JQ948534 | JQ948865 | JQ949855 | |
| F10PGBYS12 | M. ×domestica | China | KF772133 | KF772043 | KF772103 | KF772073 | KF772163 | |
| C. orchidophilum | CBS 632.80 1 | Dendrobium sp. | USA | JQ948151 | JQ949472 | JQ948481 | JQ948812 | JQ949802 |
| C. orientalis | F10PGBYS1 | M. ×domestica | China | KF772134 | KF772044 | KF772104 | KF772074 | KF772164 |
| F10PGBYS2 | M. ×domestica | China | KF772135 | KF772045 | KF772105 | KF772075 | KF772165 | |
| F10PGBYS3 | M. ×domestica | China | KF772136 | KF772046 | KF772106 | KF772076 | KF772166 | |
| F10PGBYS4 | M. ×domestica | China | KF772137 | KF772047 | KF772107 | KF772077 | KF772167 | |
| F10PGBYS7 | M. ×domestica | China | KF772138 | KF772048 | KF772108 | KF772078 | KF772168 | |
| F10PGBYS8 | M. ×domestica | China | KF772139 | KF772049 | KF772109 | KF772079 | KF772169 | |
| F10PGBYS10 | M. ×domestica | China | KF772140 | KF772050 | KF772110 | KF772080 | KF772170 | |
| CBS 128555 2 | Malus domestica | New Zealand | JQ948305 | JQ949626 | JQ948635 | JQ948966 | JQ949956 | |
| C. queenslandicum | ICMP 1778 1 | Carica papaya | Australia | JX010276 | JX009447 | JX009934 | JX009899 | JX010414 |
| ICMP 18705 3 | Coffea sp. | Fiji | JX010185 | JX009490 | JX010036 | JX009890 | JX010412 | |
| C. salicis | CBS 113.14 2 | M. ×domestica | Germany | JQ948478 | JQ949799 | JQ948809 | JQ949139 | JQ950129 |
| IMI 385055 2 | M. ×domestica | New Zealand | JQ948472 | JQ949793 | JQ948803 | JQ949133 | JQ950123 | |
| C. salsolae | ICMP 19051 1 | Salsola tragus | Hungary | JX010242 | JX009562 | JX009916 | JX009863 | JX010403 |
| C. siamense | CBS 130417 1 | Coffea arabica | Thailand | JX010171 | FJ907423 | JX009924 | JX009865 | JX010404 |
| ICMP 17795 3 | M. ×domestica | USA | JX010162 | JX009506 | JX010051 | JX009805 | JX010393 | |
| F12PGSQ02 | M. ×domestica | China | KF772127 | KF772037 | KF772097 | KF772067 | KF772157 | |
| F10PGWFT2 | M. ×domestica | China | KF772120 | KF772030 | KF772090 | KF772060 | KF772150 | |
| F11PGQX26 | M. ×domestica | China | KF772121 | KF772031 | KF772091 | KF772061 | KF772151 | |
| F11PGLQ22 | M. ×domestica | China | KF772122 | KF772032 | KF772092 | KF772062 | KF772152 | |
| F12PGMJ01 | M. ×domestica | China | KF772123 | KF772033 | KF772093 | KF772063 | KF772153 | |
| C. simmondsii | CBS 126524 2 | Cyclamen sp. | Netherlands | JQ948281 | JQ949602 | JQ948611 | JQ948942 | JQ949932 |
| CBS 122122 1 | Carica papaya | Australia | JQ948276 | JQ949597 | JQ948606 | JQ948937 | ||
| C. tropicale | CBS 124949 1 | Theobroma cacao | Panama | JX010264 | JX009489 | JX010007 | JX009870 | |
| ICMP 18672 3 | Litchi chinensis | Japan | JX010275 | JX009480 | JX010020 | JX009826 | ||
3. Results
3.1. Isolate Isolation
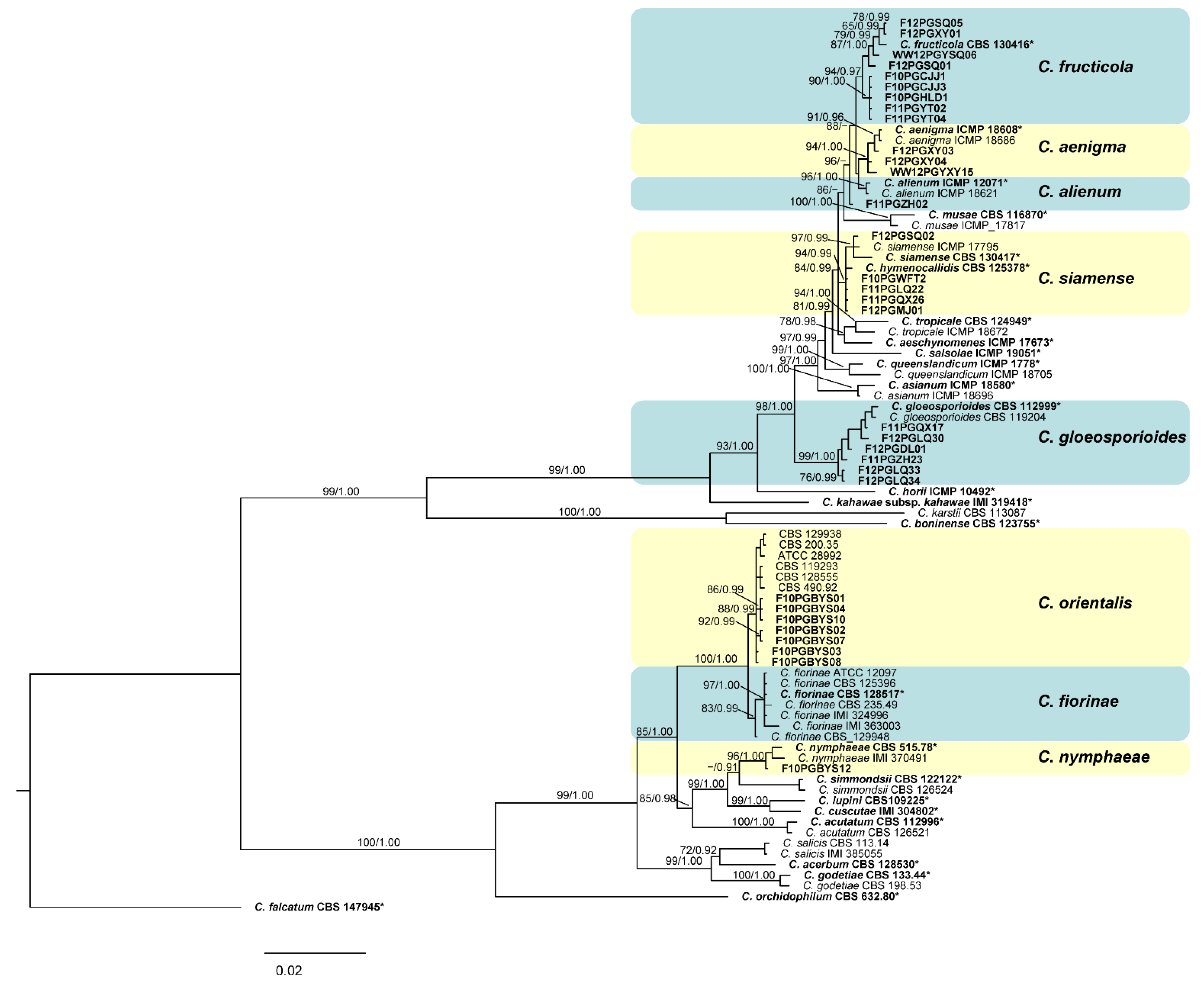
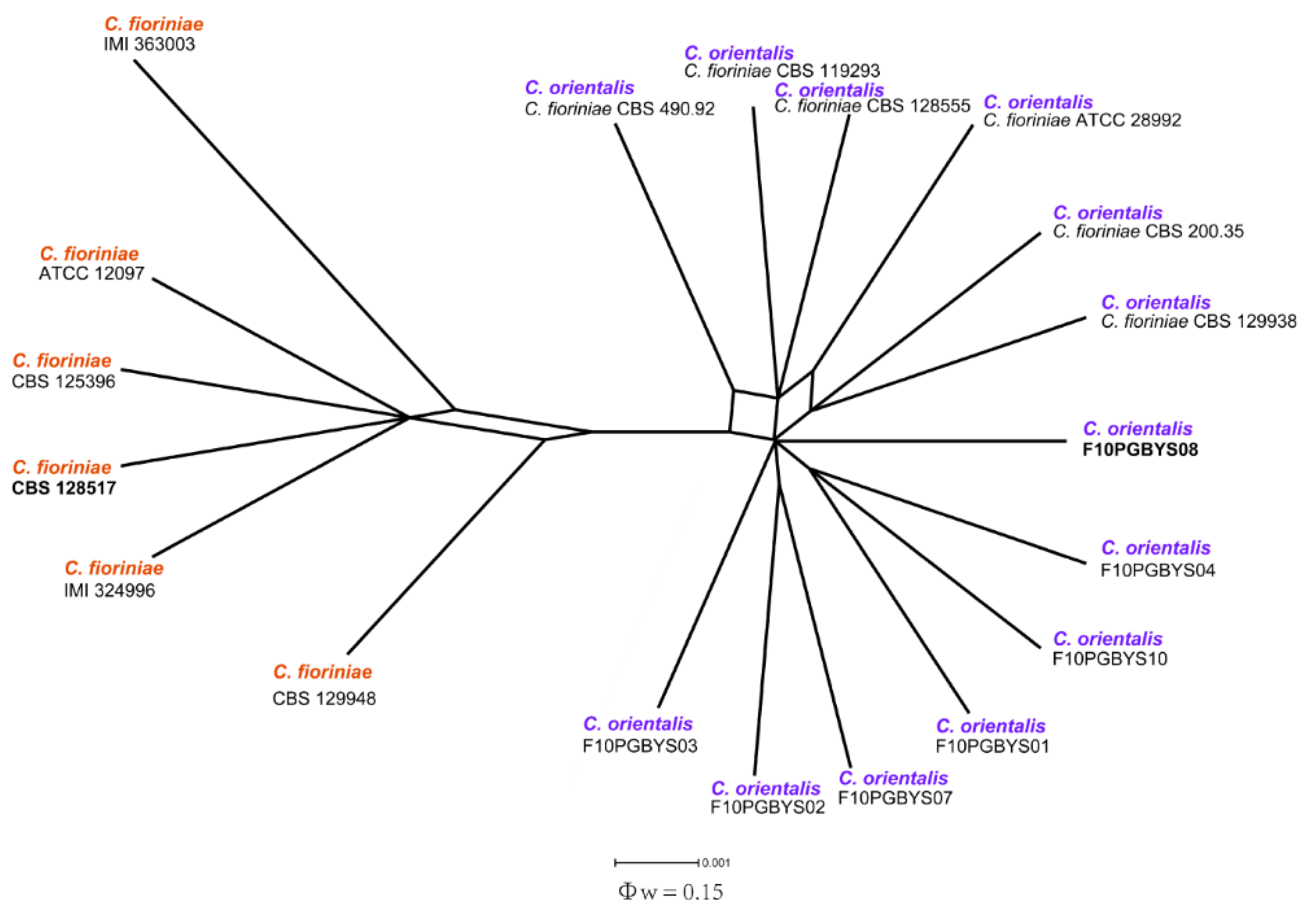
3.2. Phylogenetic Analysis
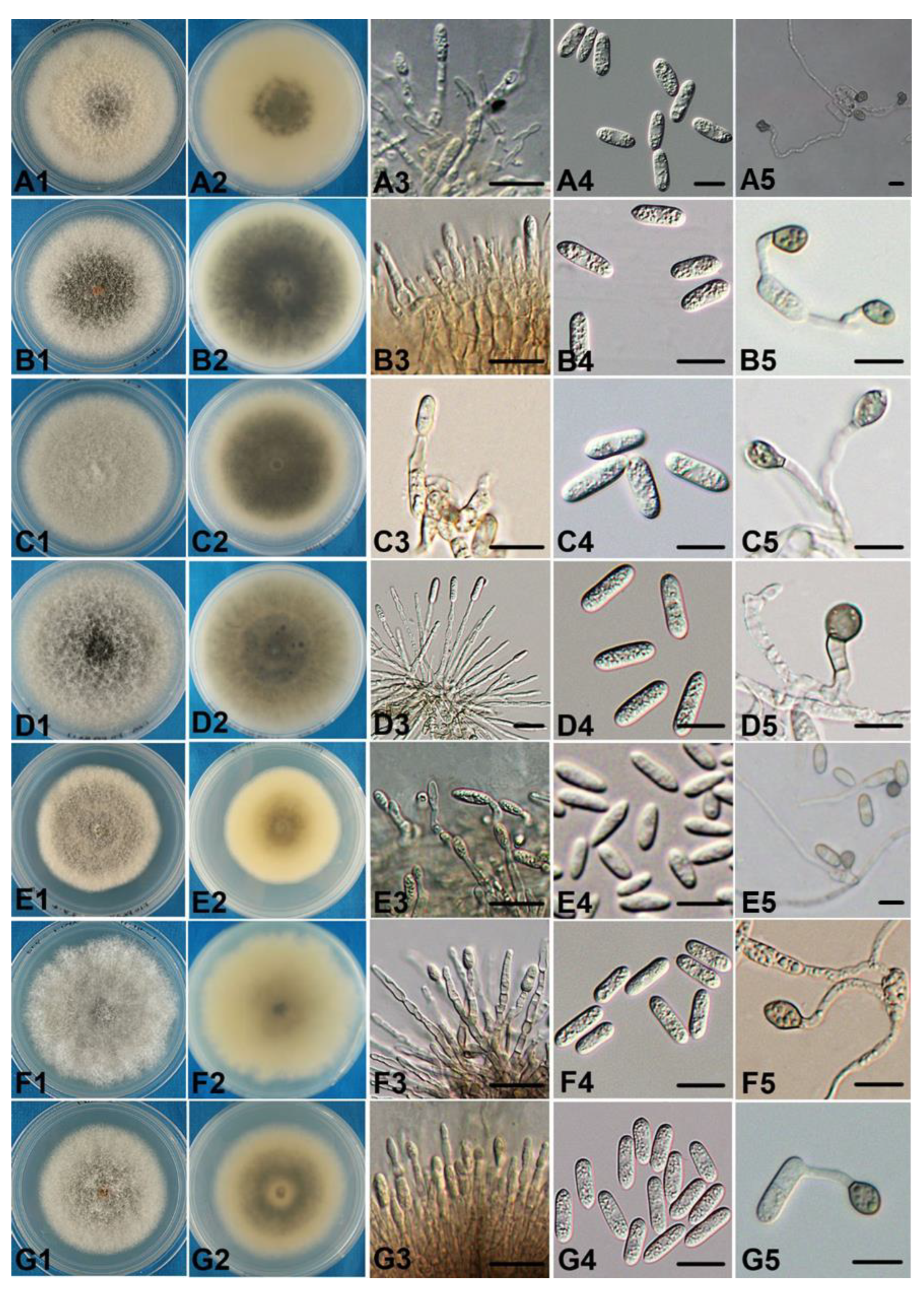
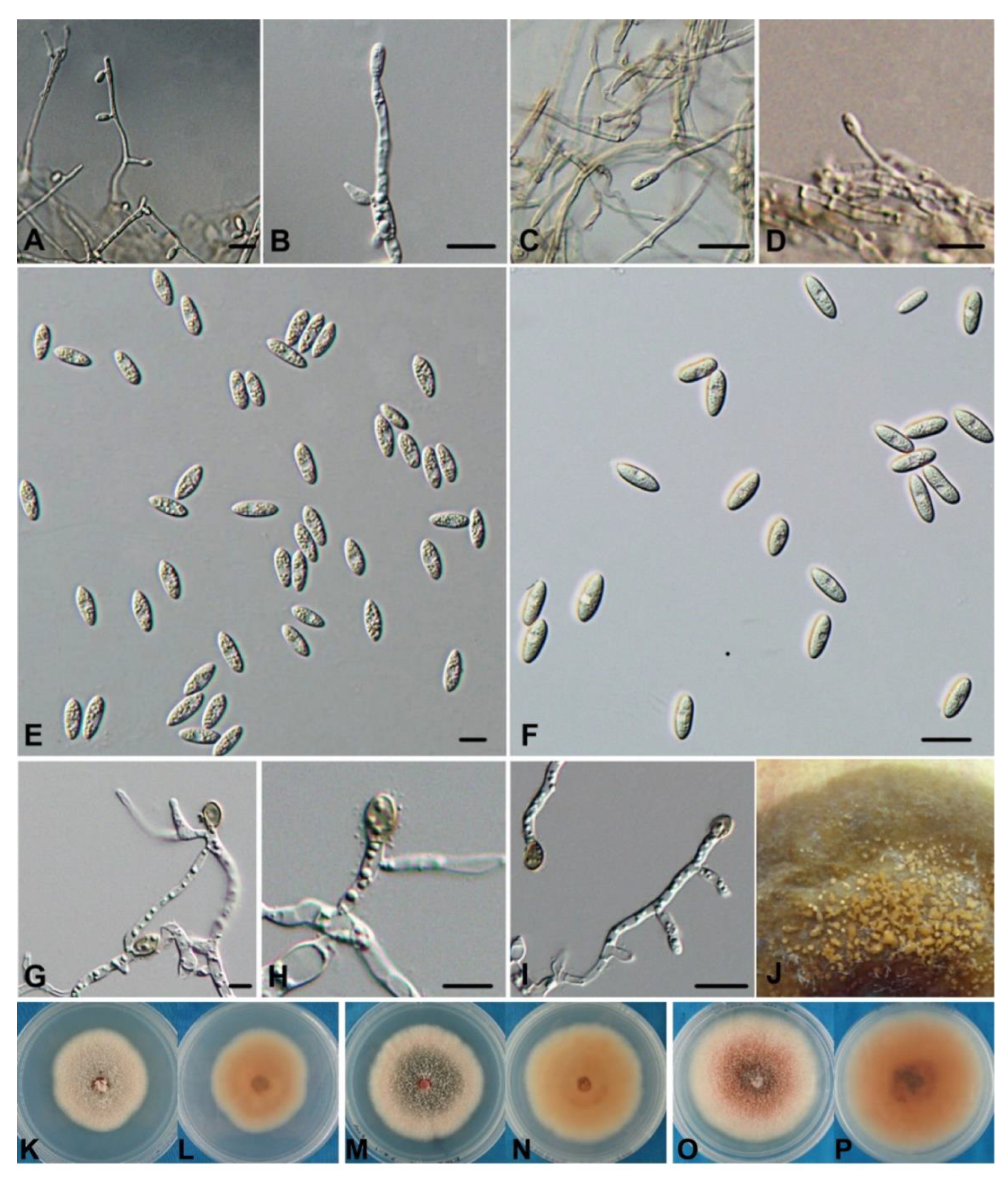
3.3. Taxonomy
- Colletotrichum orientalisDandan Fu & G.Y. Sun, sp. nov. Figure 4.
3.4. Pathogenicity Tests
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- González, E.; Sutton, T.B. Population diversity within isolates of Colletotrichum spp. causing Glomerella leaf spot and bitter rot of apple in three orchards in North Carolina. Plant Dis. 2004, 88, 1335–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrenk, H.V.; Spaulding, P. The bitter rot of apples. USDA Bur. Plant Ind. Bull. 1903, 44, 1–54. [Google Scholar]
- Simmond, J.H. A study of the species of Colletotrichum causing ripe fruit rots in Queensland. Qld. J. Agric. Anim. Sci. 1966, 22, 437–459. [Google Scholar]
- Sutton, T.B. Bitter rot. In Compendium of Apple and Pear Disease; Jones, A.L., Aldwinckle, H.S., Eds.; The American Phytopathological Society: St. Paul, MN, USA, 1990; pp. 15–26. [Google Scholar]
- Jones, A.L.; Ehret, G.R.; Meyer, M.P.; Shane, W.W. Occurrence of bitter rot on apple in Michigan. Plant Dis. 1996, 80, 1294–1297. [Google Scholar] [CrossRef]
- Shi, Y.; Correll, J.C.; Guerber, J.C.; Rom, C.R. Frequency of Colletotrichum species causing bitter rot of apple in the southeastern United States. Plant Dis. 1996, 80, 692–696. [Google Scholar] [CrossRef]
- González, E.; Sutton, T.B.; Correll, J.C. Clarification of the etiology of Glomerella leaf spot and bitter rot of apple caused by Colletotrichum spp. based on morphology and genetic, molecular, and pathogenicity tests. Phytopathology 2006, 96, 982–992. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.H.; Kim, D.H.; Jeon, Y.A.; Uhm, J.Y.; Hong, S.B. Molecular and cultural characterization of Colletotrichum spp. causing bitter rot of apples in Korea. Plant Pathol. 2007, 23, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Giaretta, D.R.; Bogo, A.; Coelho, C.; Guidolin, A.F.; de Mesquita-Dantas, A.C.; Gomes, E.A. ITS-rDNA phylogeny of Colletotrichum spp. causal agent of apple Glomerella leaf spot. Cienc. Rural 2010, 40, 806–812. [Google Scholar] [CrossRef]
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum: Current status and future directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef] [Green Version]
- Velho, A.C.; Stadnik, M.J.; Wallhead, M. Unraveling Colletotrichum species associated with Glomerella leaf spot of apple. Trop. Plant Pathol. 2019, 44, 197–204. [Google Scholar] [CrossRef]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.; Crous, P.W. The Colletotrichum acutatum species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mari, M.; Guidarelli, M.; Martini, C.; Spadoni, A. First report of Colletotrichum acutatum causing bitter rot on apple in Italy. Plant Dis. 2011, 96, 144. [Google Scholar] [CrossRef] [PubMed]
- Cheon, W.; Kim, Y.S.; Jeon, Y.H. First report of anthracnose caused by Colletotrichum gloeosporioides on Malus prunifolia in Korea. Plant Dis. 2012, 96, 766. [Google Scholar] [CrossRef]
- Kou, L.P.; Gaskins, V.; Luo, Y.G.; Jurick, W.M. First report of Colletotrichum fioriniae causing postharvest decay on ‘Nittany’ apple fruit in the United States. Plant Dis. 2013, 98, 993. [Google Scholar] [CrossRef] [PubMed]
- Baroncelli, R.; Sreenivasaprasad, S.; Thon, M.R.; Sukno, S.A. First report of apple bitter rot caused by Colletotrichum godetiae in the United Kingdom. Plant Dis. 2014, 98, 1000–1001. [Google Scholar] [CrossRef]
- Velho, A.C.; Stadnik, M.J.; Casanova, L.; Mondino, P.; Alaniz, S. First report of Colletotrichum karstii causing Glomerella leaf spot on apple in Santa Catarina State, Brazil. Plant Dis. 2014, 98, 157–158. [Google Scholar] [CrossRef]
- Wallhead, M.; Broders, G.; Beaudoin, E.; Peralta, C.; Broders, K. Phylogenetic assessment of Colletotrichum species associated with bitter rot and Glomerella leaf spot in the northeastern US. Phytopathology 2014, 104, 123–124. [Google Scholar]
- Munir, M.; Amsden, B.; Dixon, E.; Vaillancourt, L.; Gauthier, N.A. Characterization of Colletotrichum species causing bitter rot of apples in Kentucky orchards. Plant Dis. 2016, 100, 2194–2203. [Google Scholar] [CrossRef] [Green Version]
- Velho, A.C.; Alaniz, S.; Casanova, L.; Mondino, P.; Stadnik, M.J. New insights into the characterization of Colletotrichum species associated with apple diseases in southern Brazil and Uruguay. Fungal Biol. 2015, 119, 229–244. [Google Scholar] [CrossRef]
- Wenneker, M.; Pham, K.; Kerkhof, E.; Harteveld, D.O. First report of preharvest fruit rot of ’Pink Lady’ apples caused by Colletotrichum fructicola in Italy. Plant Dis. 2021, 105, 1561. [Google Scholar] [CrossRef]
- Bragança, C.A.; Damm, U.; Baroncelli, R.; Massola Júnior, N.S.; Crous, P.W. Species of the Colletotrichum acutatum complex associated with anthracnose diseases of fruit in Brazil. Fungal Biol. 2016, 120, 547–561. [Google Scholar] [CrossRef] [PubMed]
- Nodet, P.; Baroncelli, R.; Faugère, D.; Le Floch, G. First report of apple bitter rot caused by Colletotrichum fioriniae in Brittany, France. Plant Dis. 2016, 100, 1497. [Google Scholar] [CrossRef]
- Khodadadi, F.; González, J.B.; Martin, P.L.; Giroux, E.; Bilodeau, G.J.; Peter, K.A.; Doyle, V.P.; Aćimović, S.G. Identification and characterization of Colletotrichum species causing apple bitter rot in New York and description of C. noveboracense sp. nov. Sci. Rep. 2020, 10, 11043. [Google Scholar] [CrossRef] [PubMed]
- Cabrefiga, J.; Pizà, D.; Vilardell, P.; Luque, J. First report of Colletotrichum chrysophilum causing apple bitter rot in Spain. Plant Dis. 2022, PDIS07211578PDN. [Google Scholar] [CrossRef] [PubMed]
- Wenneker, M.; Pham, K.; Lemmers, M.; de Boer, F.A.; van der Lans, A.M.; van Leeuwen, P.J.; Hollinger, T.C. First report of Colletotrichum godetiae causing bitter rot on ‘Golden Delicious’ apples in the Netherlands. Plant Dis. 2015, 100, 218. [Google Scholar] [CrossRef]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Fisher, M.C. Phylogenetic species recognition and species concepts in fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.X.; Zhang, Z.F.; Li, B.H.; Wang, H.Y.; Dong, X.L. First report of Glomerella leaf spot of apple caused by Glomerella cingulata in China. Plant Dis. 2012, 96, 912–913. [Google Scholar] [CrossRef]
- Velho, A.C.; Stadnik, M.J.; Casanova, L.; Mondino, P.; Alaniz, S. First report of Colletotrichum nymphaeae causing apple bitter rot in Southern Brazil. Plant Dis. 2014, 98, 567–568. [Google Scholar] [CrossRef]
- Moreira, R.R.; Peres, N.A.; May De Mio, L.L. Colletotrichum acutatum and C. gloeosporioides species complexes associated with apple in Brazil. Plant Dis. 2019, 103, 268–275. [Google Scholar] [CrossRef] [Green Version]
- Alaniz, S.; Cuozzo, V.; Martínez, V.; Stadnik, M.J.; Mondino, P. Ascospore infection and Colletotrichum species causing Glomerella leaf spot of apple in Uruguay. Plant Pathol. J. 2019, 35, 100–111. [Google Scholar] [CrossRef]
- Zhang, Z.; Yan, M.; Li, W.; Guo, Y.; Liang, X. First report of Colletotrichum aenigma causing apple Glomerella leaf spot on the Granny Smith cultivar in China. Plant Dis. 2021, 105, 1563. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Li, J.Y. Studies on species of Colletotrichum in Province Shaanxi, China. Acta Mycol. Sin. 1987, 6, 211–218. [Google Scholar]
- Zhang, R.; Wang, S.F.; Cui, J.Q.; Sun, G.Y. First report of bitter rot caused by Colletotrichum acutatum on apple in China. Plant Dis. 2008, 92, 1471. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Wang, S.F.; Cui, J.Q.; Sun, G.Y. Identification of pathogens causing apple fruit bitter rot in Shaanxi and Henan Provinces. Sci. Agric. Sin. 2009, 42, 3224–3229. [Google Scholar]
- Than, P.P.; Shivas, R.G.; Jeewon, R.; Pongsupasamit, S.; Marney, T.S.; Taylor, P.W.; Hyde, K.D. Epitypification and phylogeny of Colletotrichum acutatum J.H. Simmonds. Fungal Divers. 2008, 28, 97–108. [Google Scholar]
- Barnes, I.; Roux, J.; Wingfield, M.J.; Coetzee, M.P.; Brenda, D.; Wingfield, B.D. Characterization of Seiridium spp. associated with cypress canker based on β-tubulin and histone sequences. Plant Dis. 2001, 85, 317–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes -- application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Guerber, J.C.; Liu, B.; Correll, J.C.; Johnston, P.R. Characterization of diversity in Colletotrichum acutatum sensu lato by sequence analysis of two introns, mtDNA and intron RFLPs, and mating compatibility. Mycologia 2003, 95, 872–895. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Damm, U.; Woudenberg, J.H.; Cannon, P.F.; Crous, P.W. Colletotrichum species with curved conidia from herbaceous hosts. Fungal Divers. 2009, 39, 45–87. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H. SplitsTree: Analyzing and visualizing evolutionary data. Bioinformatics 1998, 14, 68–73. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [Green Version]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.; Johnston, P.R.; Weir, B.S.; Tan, Y.P.; Shivas, R.G.; Crous, P.W. The Colletotrichum boninense species complex. Stud. Mycol. 2012, 73, 1–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prihastuti, H.; Cai, L.; Chen, H.; McKenzie, E.H.; Hyde, K.D. Characterization of Colletotrichum species associated with coffee berries in northern Thailand. Fungal Divers. 2009, 39, 89–109. [Google Scholar]
- Penzig, A.G.O. Fungi agrumicoli. Contribuzione allo studio dei funghi parassiti degli agrumi. In Michelia; Patavii: Berlin, Germany, 1882; pp. 385–508, Sumptibus auctoris, Typis seminarii. [Google Scholar]
- van der Aa, H.A. A leaf spot of Nymphaea alba in the Netherlands. Netherlands J Plant Pathol. 1978, 84, 109–115. [Google Scholar]
- Yang, Y.L.; Liu, Z.Y.; Cai, L.; Hyde, K.D.; Yu, Z.N.; McKenzie, E.H. Colletotrichum anthracnose of Amaryllidaceae. Fungal Divers. 2009, 39, 123–146. [Google Scholar]
- Sharma, G.; Kumar, N.; Weir, B.S.; Hyde, K.D.; Shenoy, B.D. The ApMat marker can resolve Colletotrichum species: A case study with Mangifera indica. Fungal Divers. 2013, 61, 117–138. [Google Scholar] [CrossRef]
- Liu, F.; Damm, U.; Cai, L.; Crous, P.W. Species of the Colletotrichum gloeosporioides complex associated with anthracnose diseases of Proteaceae. Fungal Divers. 2013, 61, 89–105. [Google Scholar] [CrossRef]
- Freeman, S.; Shabi, E. Cross-infection of subtropical and temperate fruits by Colletotrichum species from various hosts. Physiol. Mol. Plant Pathol. 1996, 49, 395–404. [Google Scholar] [CrossRef]
- Liu, Y.; Li, B.; Wang, C.; Liu, C.; Kong, X.; Zhu, J.; Dai, H. Genetics and molecular marker identification of a resistance to Glomerella leaf spot in apple. Hortic. Plant J. 2016, 2, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Rockenbach, M.F.; Velho, A.C.; Gonçalves, A.E.; Mondino, P.E.; Alaniz, S.M.; Stadnik, M.J. Genetic structure of Colletotrichum fructicola associated to apple bitter rot and Glomerella leaf spot in southern Brazil and Uruguay. Phytopathology 2016, 106, 774–781. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Wang, M.; Damm, U.; Crous, P.W.; Cai, L. Species boundaries in plant pathogenic fungi: A Colletotrichum case study. BMC Evol. Biol. 2016, 16, 1–14. [Google Scholar] [CrossRef] [Green Version]





| Species | Isolate | Non-Wounded | Wounded |
|---|---|---|---|
| C. alienum | F11PGZH02 | +++ | +++ |
| C. fructicola | F12PGSQ01 | ++ | +++ |
| C. gloeosporioides | F11PGQX17 | + | +++ |
| C. nymphaeae | F10PGBYS12 | ++ | +++ |
| C. siamense | F12PGSQ02 | ++ | +++ |
| C. orientalis | F10PGBYS08 | + | +++ |
| Species | Isolate | Origin | Inoculation Cultivar | Inoculation Outcome |
|---|---|---|---|---|
| C. aenigma | F12PGXY03 | GLS lesion | Fuji | − |
| Gala | + | |||
| W12PGYXY15 | GLS lesion | Fuji | − | |
| Gala | + | |||
| C. fructicola | F12PGSQ05 | GLS lesion | Fuji | − |
| Gala | + | |||
| W12PGYSQ06 | GLS lesion | Fuji | − | |
| Gala | + | |||
| C. alienum | F11PGZH02 | ABR lesion | Fuji | − |
| Gala | − | |||
| C. fructicola | F12PGSQ01 | ABR lesion | Fuji | − |
| Gala | − | |||
| C. gloeosporioides | F11PGQX17 | ABR lesion | Fuji | − |
| Gala | − | |||
| C. nymphaeae | F10PGBYS12 | ABR lesion | Fuji | − |
| Gala | − | |||
| C. siamense | F12PGSQ02 | ABR lesion | Fuji | − |
| Gala | − | |||
| C. orientalis | F10PGBYS08 | ABR lesion | Fuji | − |
| Gala | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Fu, D.; Wang, W.; Gleason, M.L.; Zhang, R.; Liang, X.; Sun, G. Diversity of Colletotrichum Species Causing Apple Bitter Rot and Glomerella Leaf Spot in China. J. Fungi 2022, 8, 740. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8070740
Chen Y, Fu D, Wang W, Gleason ML, Zhang R, Liang X, Sun G. Diversity of Colletotrichum Species Causing Apple Bitter Rot and Glomerella Leaf Spot in China. Journal of Fungi. 2022; 8(7):740. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8070740
Chicago/Turabian StyleChen, Yang, Dandan Fu, Wei Wang, Mark L. Gleason, Rong Zhang, Xiaofei Liang, and Guangyu Sun. 2022. "Diversity of Colletotrichum Species Causing Apple Bitter Rot and Glomerella Leaf Spot in China" Journal of Fungi 8, no. 7: 740. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8070740