
Unravelling the French National Fungal Database: Geography, Temporality, Taxonomy and Ecology of the Recorded Diversity


Abstract
:1. Introduction
2. Materials and Methods
2.1. National Fungal Database
2.2. Assembling of Analysis Matrices
2.3. Statistical Analyses
3. Results
3.1. The French National Dataset
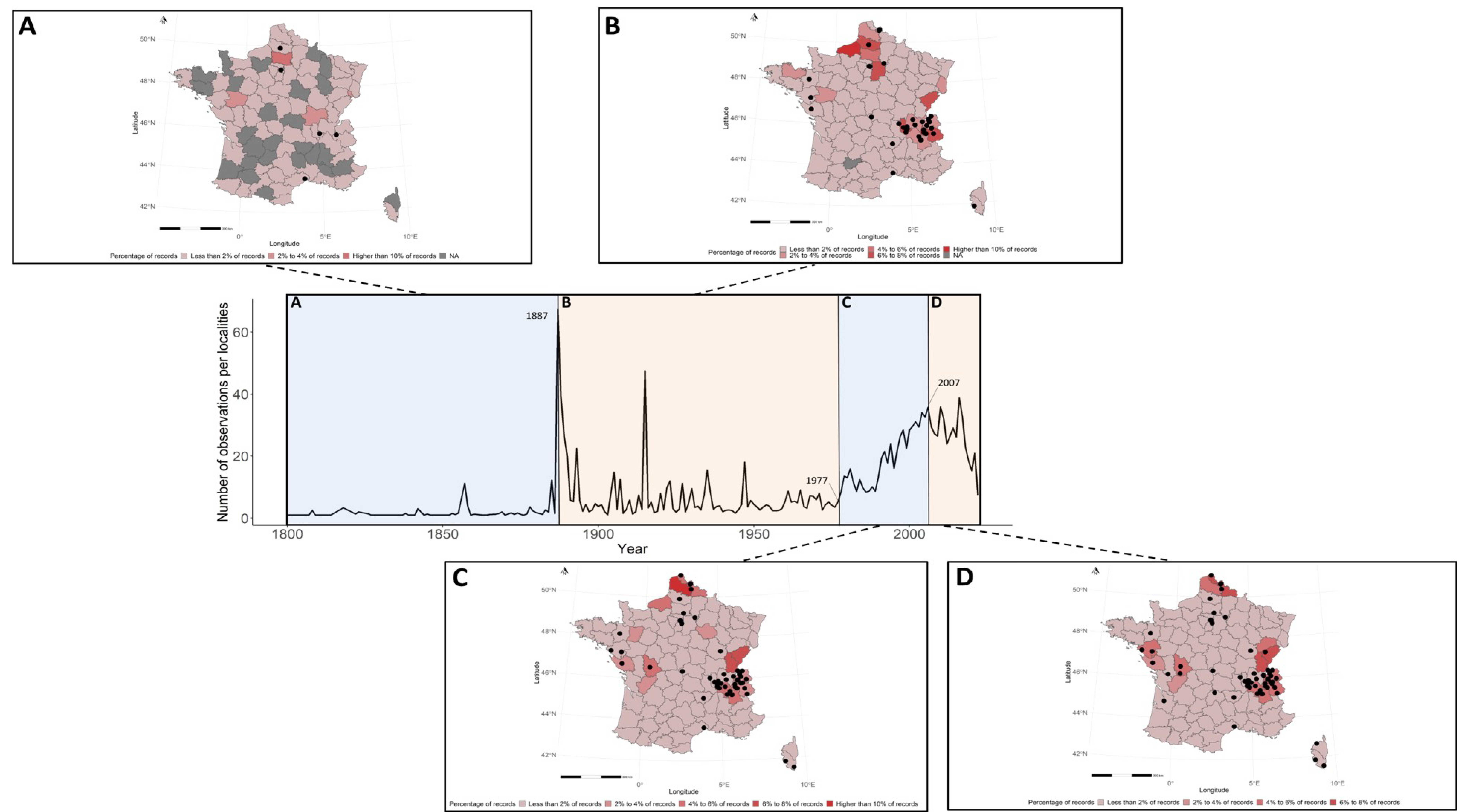
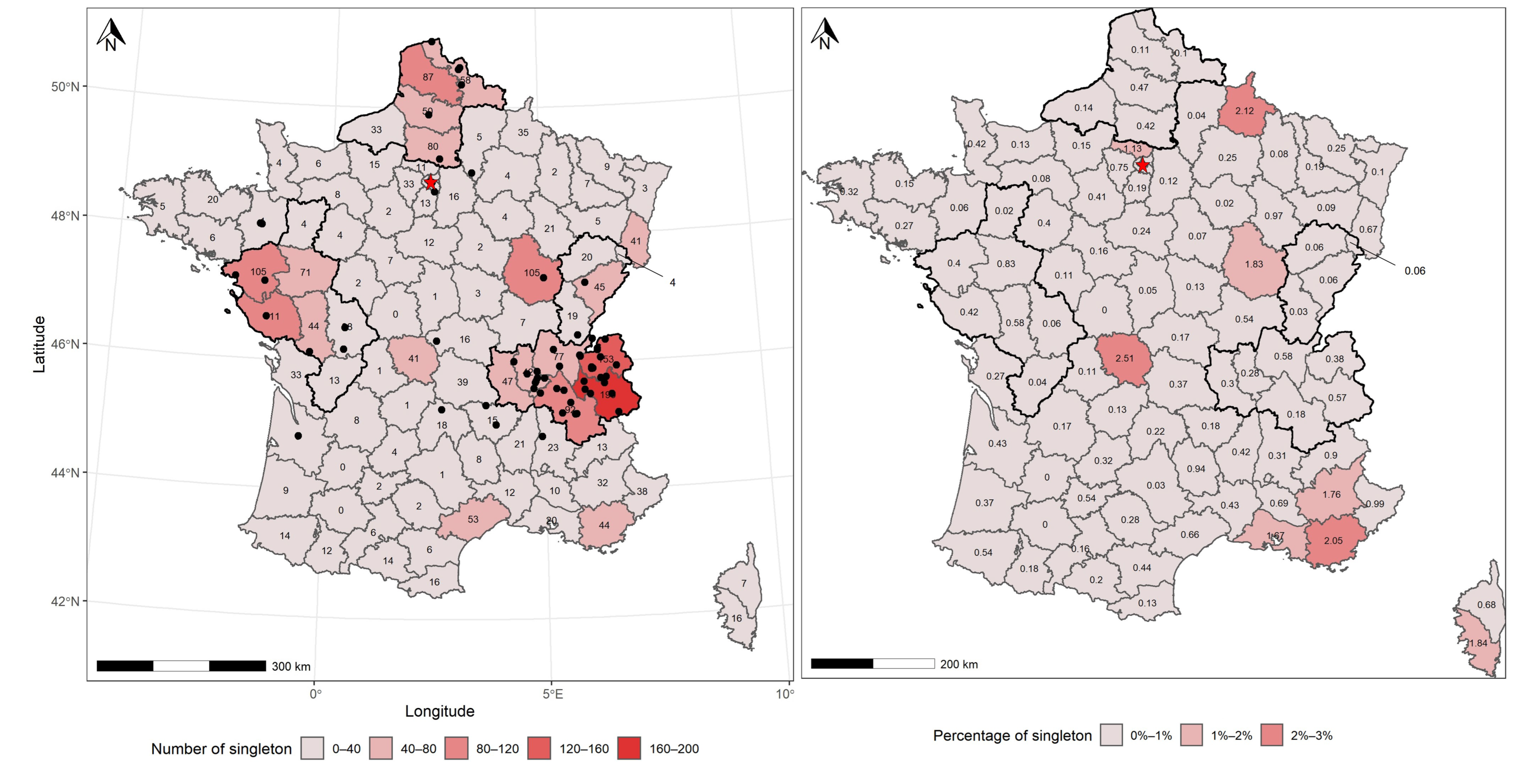
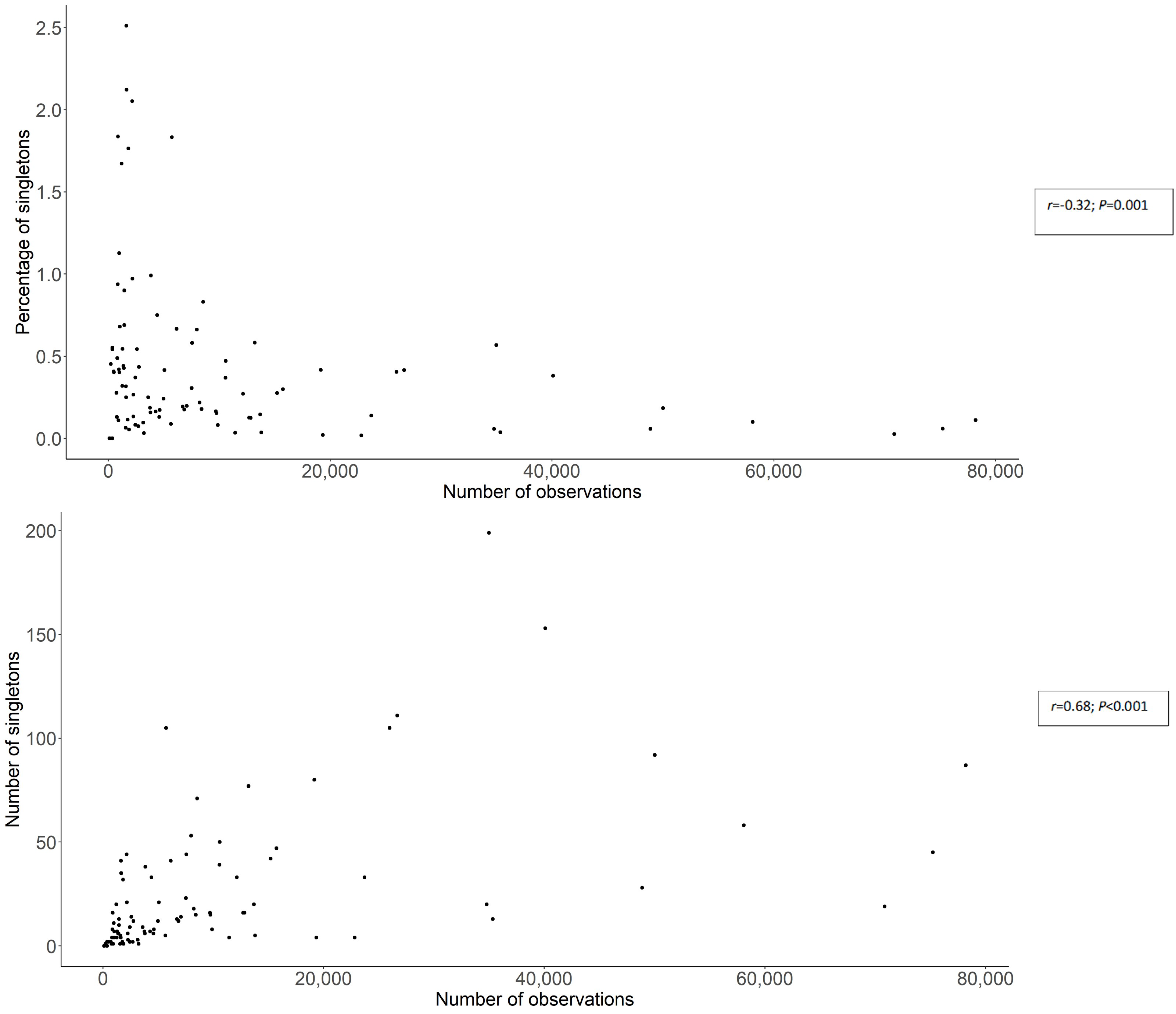
3.2. Spatial Patterns of Fungal Records
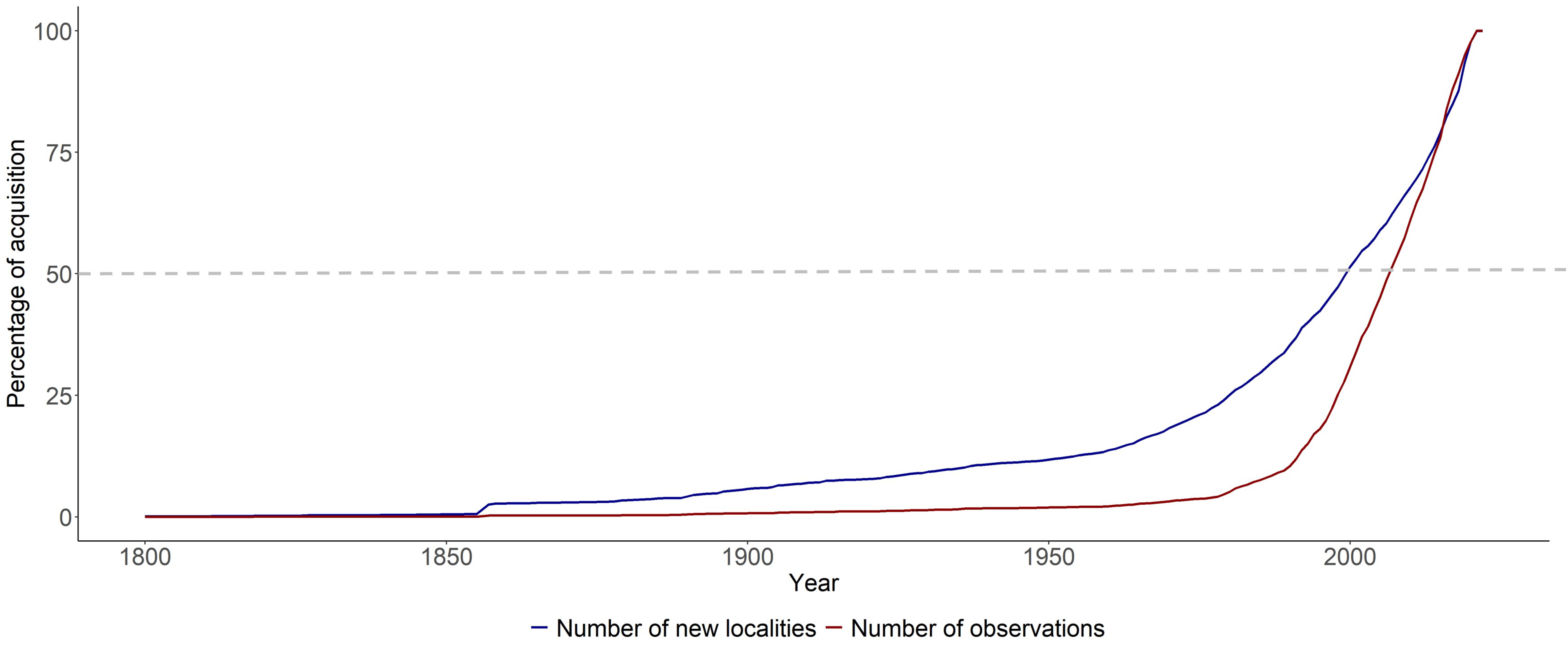
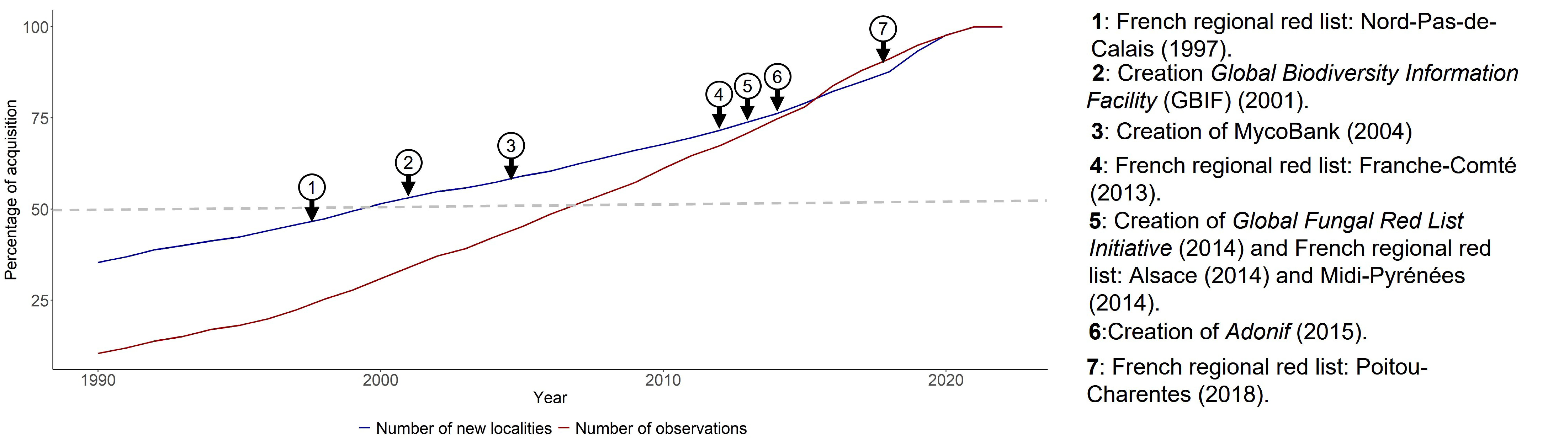
3.3. Temporal Patterns of Fungal Data Acquisition
3.4. Distribution of Fungal Alpha Diversity
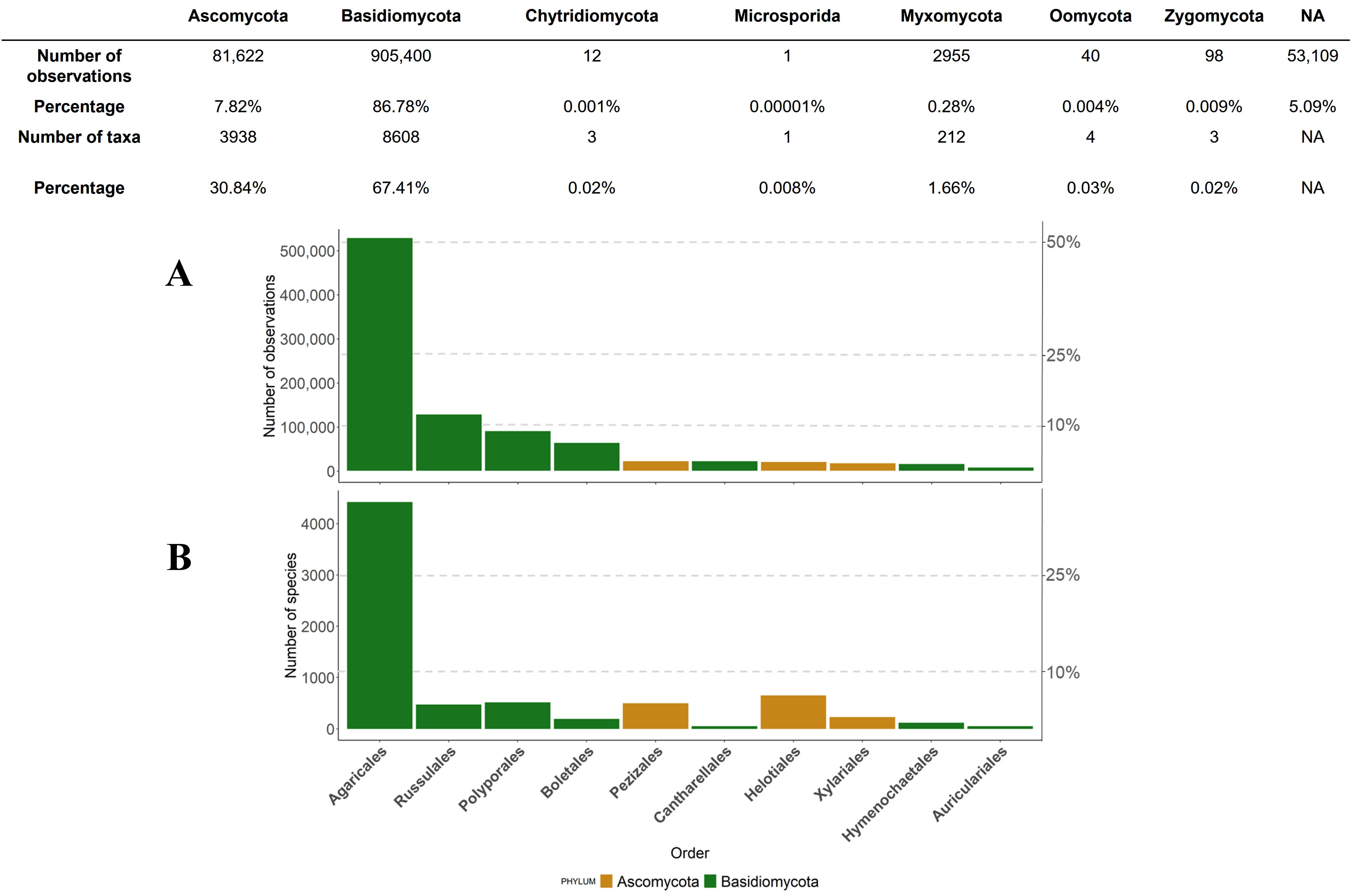
3.5. Taxonomy of Fungal Records
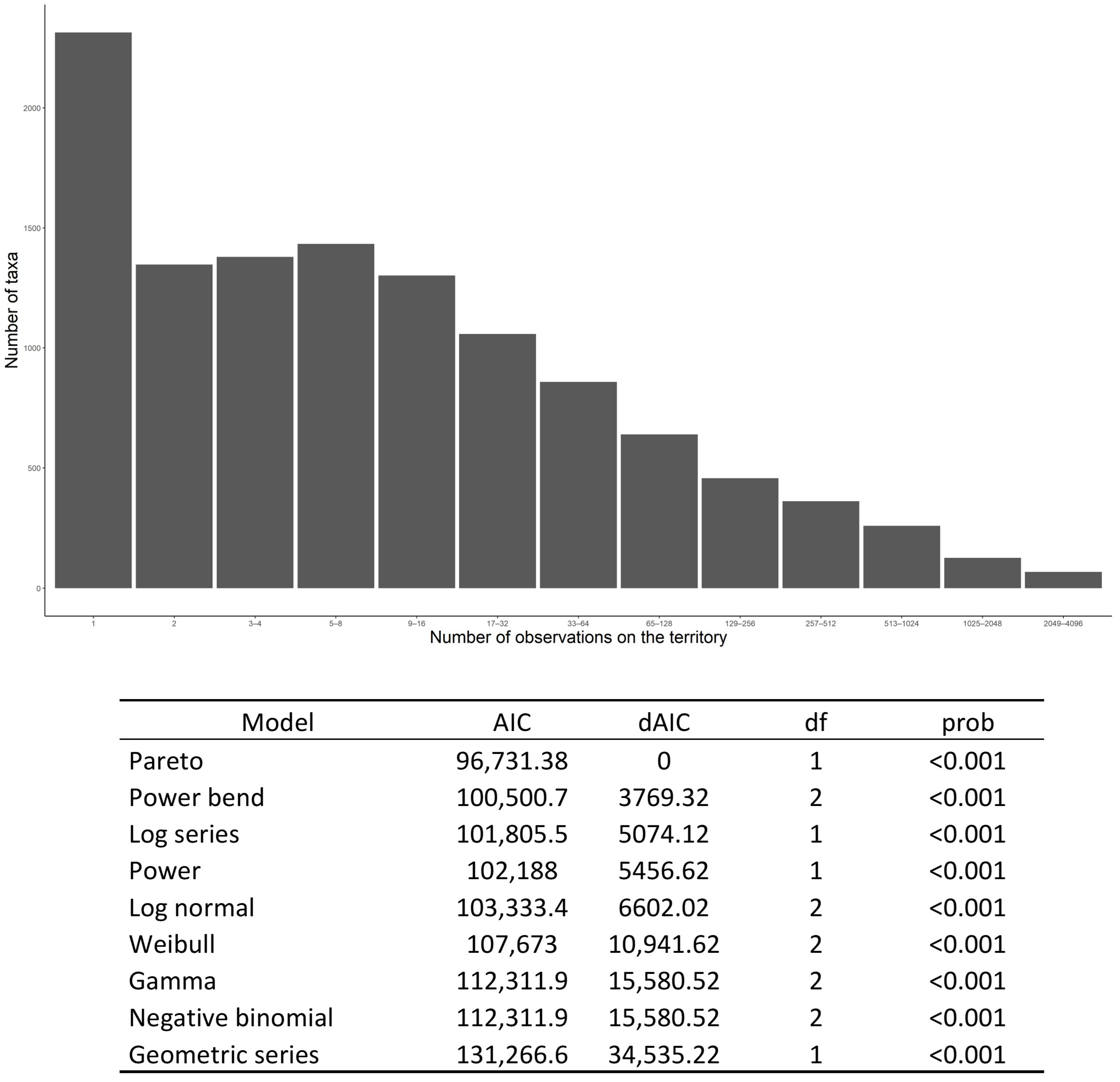
3.6. Distribution Patterns of Fungal Species Occurrence
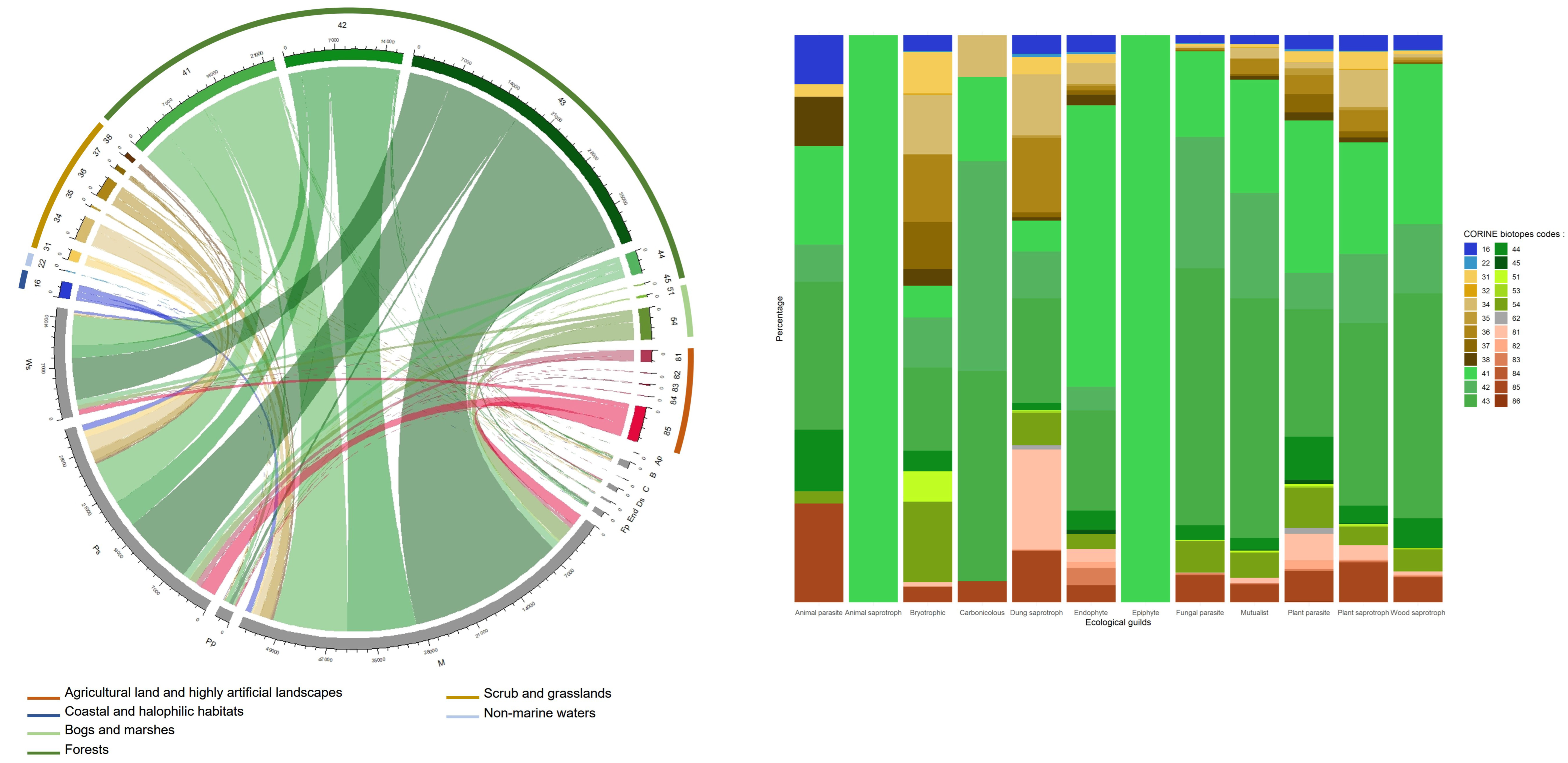
3.7. Ecology of Fungal Records
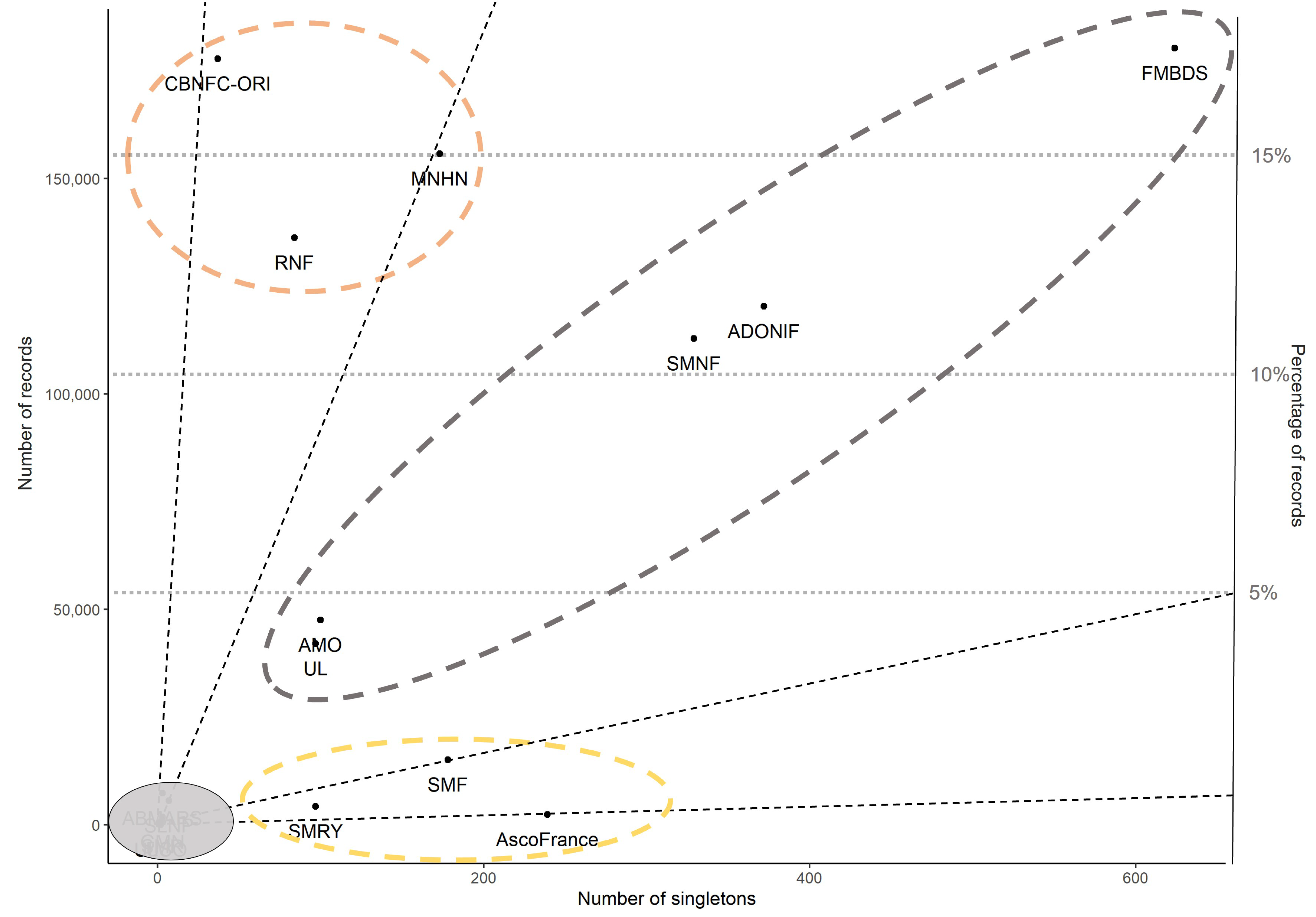
3.8. Distribution Patterns of Contributors
4. Discussion
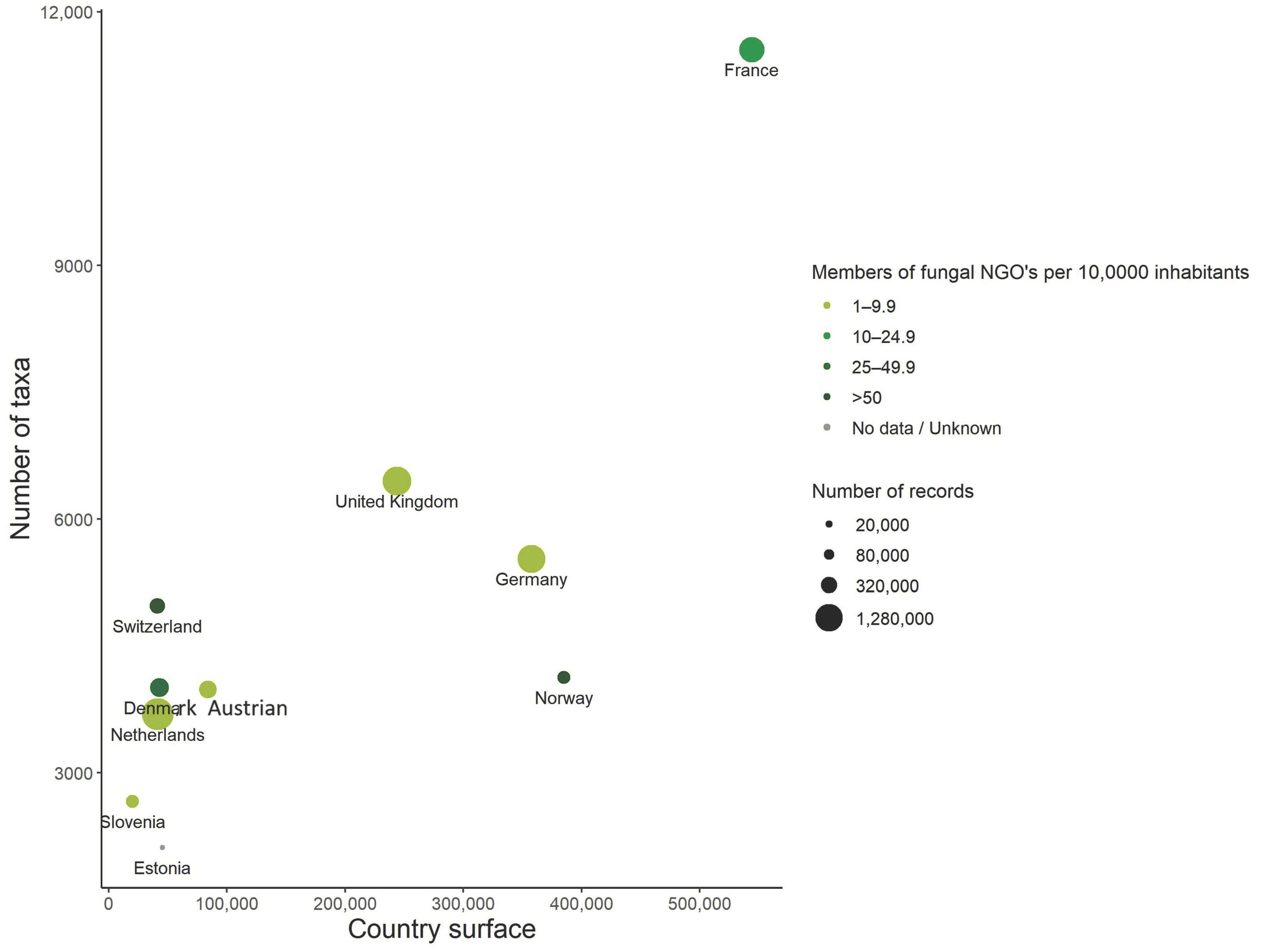
4.1. French Metropolitan Territory, a Species Rich Area for Fungi or a Mycologist-Rich Area?
4.2. Red List Initiatives Drive Fungal Recording Activity
4.3. A Two-Century Long Dynamic of Fungal Data Recording
4.4. A Deficit of Fungal Record in the French Mediterranean
4.5. Beyond the Data: An Unbalanced Taxonomy of Records
4.6. Beyond the Data: An Unbalanced Ecology of Records
4.7. Between Fungal Rarity and Sampling Activity (Methodological Development)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gange, A.; Gange, E.G.; Sparks, T.H.; Boddy, L. Rapid and recent changes in fungal fruiting patterns. Science 2007, 316, 71. [Google Scholar] [CrossRef] [PubMed]
- Kauserud, H.; Heegaard, E.; Halvorsen, R.; Boddy, L.; Høiland, K.; Stenseth, N. Mushroom’s Spore Size and Time of Fruiting Are Strongly Related: Is Moisture Important? Biol. Lett. 2010, 7, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Gange, A.; Gange, E.G.; Mohammad, A.B.; Boddy, L. Host Shifts in Fungi Caused by Climate Change? Fungal Ecol. 2011, 4, 184–190. [Google Scholar] [CrossRef]
- Büntgen, U.; Kauserud, H.; Egli, S. Linking Climate Variability to Mushroom Productivity and Phenology. Front. Ecol. Environ. 2012, 10, 14–19. [Google Scholar] [CrossRef]
- Diez, J.M.; James, T.Y.; McMunn, M.; Ibáñez, I. Predicting Species-Specific Responses of Fungi to Climatic Variation Using Historical Records. Glob. Chang. Biol. 2013, 19, 3145–3154. [Google Scholar] [CrossRef]
- Geldmann, J.; Heilmann-Clausen, J.; Holm, T.E.; Levinsky, I.; Markussen, B.; Olsen, K.; Rahbek, C.; Tøttrup, A.P. What Determines Spatial Bias in Citizen Science? Exploring Four Recording Schemes with Different Proficiency Requirements. Divers. Distrib. 2016, 22, 1139–1149. [Google Scholar] [CrossRef]
- Andrew, C.; Heegaard, E.; Kirk, P.M.; Bässler, C.; Heilmann-Clausen, J.; Krisai-Greilhuber, I.; Kuyper, T.W.; Senn-Irlet, B.; Büntgen, U.; Diez, J.; et al. Big Data Integration: Pan-European Fungal Species Observations’ Assembly for Addressing Contemporary Questions in Ecology and Global Change Biology. Fungal Biol. Rev. 2017, 31, 88–98. [Google Scholar] [CrossRef]
- Bates, S.T.; Miller, A.N. The M acrofungi Collections and Microfungi Collections Consortia. The Protochecklist of North American Nonlichenized Fungi. Mycologia 2018, 110, 1222–1348. [Google Scholar] [CrossRef]
- Öpik, M.; Vanatoa, A.; Vanatoa, E.; Moora, M.; Davison, J.; Kalwij, J.M.; Reier, Ü.; Zobel, M. The Online Database MaarjAM Reveals Global and Ecosystemic Distribution Patterns in Arbuscular Mycorrhizal Fungi (Glomeromycota). New Phytol. 2010, 188, 223–241. [Google Scholar] [CrossRef]
- Větrovský, T.; Morais, D.; Kohout, P.; Lepinay, C.; Algora, C.; Awokunle Hollá, S.; Bahnmann, B.D.; Bílohnědá, K.; Brabcová, V.; D’Alò, F.; et al. GlobalFungi, a Global Database of Fungal Occurrences from High-Throughput-Sequencing Metabarcoding Studies. Sci. Data 2020, 7, 228. [Google Scholar] [CrossRef]
- Primicia, I.; Camarero, J.J.; Martínez de Aragón, J.; de-Miguel, S.; Bonet, J.A. Linkages between Climate, Seasonal Wood Formation and Mycorrhizal Mushroom Yields. Agric. For. Meteorol. 2016, 228–229, 339–348. [Google Scholar] [CrossRef]
- Alday, J.G.; Bonet, J.A.; Oria-de-Rueda, J.A.; Martínez-de-Aragón, J.; Aldea, J.; Martín-Pinto, P.; de-Miguel, S.; Hernández-Rodríguez, M.; Martínez-Peña, F. Record Breaking Mushroom Yields in Spain. Fungal Ecol. 2017, 26, 144–146. [Google Scholar] [CrossRef]
- Taşkın, H.; Büyükalaca, S.; Hansen, K.; O’Donnell, K. Multilocus phylogenetic analysis of true morels (Morchella) reveals high levels of endemics in Turkey relative to other regions of Europe. Mycologia 2012, 104, 446–461. [Google Scholar] [CrossRef] [PubMed]
- Loizides, M.; Bellanger, J.M.; Clowez, P.; Richard, F.; Moreau, P.A. Combined phylogenetic and morphological studies of true morels (Pezizales, Ascomycota) in Cyprus reveal significant diversity, including Morchella arbutiphila and M. disparilis spp. nov. Mycol. Prog. 2016, 15, 1–28. [Google Scholar] [CrossRef]
- Loizides, M.; Bellanger, J.M.; Assyov, B.; Moreau, P.A.; Richard, F. Present status and future of boletoid fungi (Boletaceae) on the island of Cyprus: Cryptic and threatened diversity unravelled by ten-year study. Fungal Ecol. 2019, 41, 65–81. [Google Scholar] [CrossRef]
- Vogt-Schilb, H.; Richard, F.; Malaval, J.-C.; Rapior, S.; Fons, F.; Bourgade, V.; Schatz, B.; Buentgen, U.; Moreau, P.-A. Climate-Induced Long-Term Changes in the Phenology of Mediterranean Fungi. Fungal Ecol. 2022, 60, 101166. [Google Scholar] [CrossRef]
- Kauserud, H.; Stige, L.; Vik, J.O.; Halvorsen, R.; Høiland, K.; Stenseth, N. Mushroom Fruiting and Climate Change. Proc. Natl. Acad. Sci. USA 2008, 105, 3811–3814. [Google Scholar] [CrossRef]
- Boddy, L.; Buentgen, U.; Egli, S.; Gange, A.; Heegaard, E.; Kirk, P.M.; Mohammad, A.; Kauserud, H. Climate Variation Effects on Fungal Fruiting. Fungal Ecol. 2014, 10, 20–33. [Google Scholar] [CrossRef]
- Heilmann-Clausen, J.; Læssøe, T. On Species Richness Estimates, Climate Change and Host Shifts in Wood-Inhabiting Fungi. Fungal Ecol. 2012, 5, 641–646. [Google Scholar] [CrossRef]
- Horton, T.; Bruns, T.D. The Molecular Revolution in Ectomycorrhizal Ecology: Peeking into the Black-Box. Mol. Ecol. 2001, 10, 1855–1871. [Google Scholar] [CrossRef] [Green Version]
- Richard, F.; Millot, S.; Gardes, M.; Selosse, M.-A. Diversity and Specificity of Ectomycorrhizal Fungi Retrieved from an Old-Growth Mediterranean Forest Dominated by Quercus Ilex. New Phytol. 2005, 166, 1011–1023. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.H.; Pérez-Pérez, M.A.; Smith, M.E.; Bledsoe, C.S. Multiple Species of Ectomycorrhizal Fungi Are Frequently Detected on Individual Oak Root Tips in a Tropical Cloud Forest. Mycorrhiza 2008, 18, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Pierre-Emmanuel, C.; François, M.; Marc-André, S.; Myriam, D.; Stéven, C.; Fabio, Z.; Marc, B.; Claude, P.; Adrien, T.; Jean, G.; et al. Into the Functional Ecology of Ectomycorrhizal Communities: Environmental Filtering of Enzymatic Activities. J. Ecol. 2016, 104, 1585–1598. [Google Scholar] [CrossRef]
- Hubbell, S.P. A Unified Theory of Biogeography and Relative Species Abundance and Its Application to Tropical Rain Forests and Coral Reefs. Coral Reefs 1997, 16, S9–S21. [Google Scholar] [CrossRef]
- Buée, M.; Reich, M.; Murat, C.; Morin, E.; Nilsson, R.H.; Uroz, S.; Martin, F. 454 Pyrosequencing Analyses of Forest Soils Reveal an Unexpectedly High Fungal Diversity. New Phytol. 2009, 184, 449–456. [Google Scholar] [CrossRef]
- Baptista, P.; Reis, F.; Pereira, E.; Tavares, R.M.; Santos, P.M.; Richard, F.; Selosse, M.-A.; Lino-Neto, T. Soil DNA Pyrosequencing and Fruitbody Surveys Reveal Contrasting Diversity for Various Fungal Ecological Guilds in Chestnut Orchards. Environ. Microbiol. Rep. 2015, 7, 946–954. [Google Scholar] [CrossRef]
- Taudière, A.; Bellanger, J.-M.; Carcaillet, C.; Hugot, L.; Kjellberg, F.; Lecanda, A.; Lesne, A.; Moreau, P.-A.; Scharmann, K.; Leidel, S.; et al. Diversity of Foliar Endophytic Ascomycetes in the Endemic Corsican Pine Forests. Fungal Ecol. 2018, 36, 128–140. [Google Scholar] [CrossRef]
- Frøslev, T.G.; Kjøller, R.; Bruun, H.H.; Ejrnæs, R.; Hansen, A.J.; Læssøe, T.; Heilmann-Clausen, J. Man against Machine: Do Fungal Fruitbodies and EDNA Give Similar Biodiversity Assessments across Broad Environmental Gradients? Biol. Conserv. 2019, 233, 201–212. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.L.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global Diversity and Geography of Soil Fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Toots, M.; Diédhiou, A.G.; Henkel, T.W.; Kjøller, R.; Morris, M.H.; Nara, K.; Nouhra, E.; Peay, K.G.; et al. Towards Global Patterns in the Diversity and Community Structure of Ectomycorrhizal Fungi. Mol. Ecol. 2012, 21, 4160–4170. [Google Scholar] [CrossRef]
- Gange, A.; Allen, L.P.; Nussbaumer, A.; Gange, E.G.; Andrew, C.; Egli, S.; Senn-Irlet, B.; Boddy, L. Multiscale Patterns of Rarity in Fungi, Inferred from Fruiting Records. Glob. Ecol. Biogeogr. 2019, 28, 1106–1117. [Google Scholar] [CrossRef]
- Ridley, G. The New Zealand Connection—Amanita Inopinata—the Mystery Deepens. Field Mycol. 2000, 1, 117–118. [Google Scholar] [CrossRef]
- Pringle, A.; Bever, J.D.; Gardes, M.; Parrent, J.L.; Rillig, M.; Klironomos, J.N. Mycorrhizal Symbioses and Plant Invasions. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 699–715. [Google Scholar] [CrossRef]
- Wolfe, B.E.; Richard, F.; Cross, H.B.; Pringle, A. Distribution and Abundance of the Introduced Ectomycorrhizal Fungus Amanita Phalloides in North America. New Phytol. 2010, 185, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Alphandéry, P.; Pinton, F. Des Znieff à Natura 2000, connaissances naturalistes et conservation de la biodiversité. L’exemple icaunois. Bourgogne Nat. 2007, 6, 59–75. [Google Scholar]
- Evans, D. Building the European Union’s Natura 2000 Network. Nat. Conserv. 2012, 11–26. [Google Scholar] [CrossRef]
- Moreau, P.-A.; Hériveau, P.; Bourgade, V.; Bellanger, J.-M.; Courtecuisse, R.; Fons, F.; Rapior, S. Redécouverte et Typification Des Champignons de La Région de Montpellier Illustrés Par Michel-Félix Dunal et Alire Raffeneau-Delile. Cryptogam. Mycol. 2011, 32, 255–276. [Google Scholar] [CrossRef]
- Courtecuisse, R. Le programme d’inventaire mycologique national et de cartographie des Mycota français. 1 ère note: Présentation générale. Bullet. Soc. Mycol. Fr. 1992, 107, 161–203. [Google Scholar]
- Boury, B.; Bondu, A.; Chalange, R.; Courtecuisse, R.; Delannoy, A.; Lechat, C.; Lecuru, C.; Sellier, Y.; Vidonne, J.-P.; Moreau, P.-A. La Base Mycologique Nationale: Un projet interactif pour la gestion des données mycologiques en France. Doc. Mycol. 2018, 37, 3–14. [Google Scholar]
- FongiFrance. Available online: http://fongibase.fongifrance.fr (accessed on 22 July 2022).
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An Open Annotation Tool for Parsing Fungal Community Datasets by Ecological Guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Rdrr.io. Available online: https://rdrr.io/github/brendanf/FUNGuildR/f/README.Rmd (accessed on 22 July 2022).
- Moss, D.; Wyatt, B.K. The CORINE Biotopes Project: A Database for Conservation of Nature and Wildlife in the European Community. Appl. Geogr. 1994, 14, 327–349. [Google Scholar] [CrossRef]
- Noirfalise, A. Map of the Natural Vegetation of the Member Countries of the European Community and Council of Europe; Council of Europe: Strasbourg, France, 1987. [Google Scholar]
- R Core Team. A Language and Environment for Statistical Computing. 2013. Available online: https://www.r-project.org/ (accessed on 22 July 2022).
- R.studio Team. Studio: Integrated Development for R.studio Team; PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com (accessed on 22 July 2022).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Bivand, R.S. Progress in the R Ecosystem for Representing and Handling Spatial Data. J. Geogr. Syst. 2021, 23, 515–546. [Google Scholar] [CrossRef]
- Dunnington, D.; Thorne, B. Ggspatial: Spatial Data Framework for Ggplot2. Package Version 1. 2020. Available online: https://rdrr.io/cran/ggspatial/ (accessed on 22 July 2022).
- INSEE. Available online: https://www.insee.fr/fr/accueil (accessed on 22 July 2022).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.B.; Simpson, G.L.; Sólymos, P.; Stevens, M.H.H.; Wagner, H. vegan: Community Ecology Package. 2012. Available online: https://www.researchgate.net/publication/313502495_Vegan_Community_Ecology_Package (accessed on 22 July 2022).
- Venables, W.; Ripley, B. Modern Applied Statistics with S; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P.; Heiberger, R.M.; Schuetzenmeister, A.; Scheibe, S. Multcomp: Simultaneous Inference in General Parametric Models. Package Version 1.4–5. 2016. Available online: http://cran.stat.sfu.ca/web/packages/multcomp/ (accessed on 22 July 2022).
- Prado, P.; Miranda, M. Fitting Species Abundance Models with Maximum Likelihood—Quick Reference for Sads Package 2014. Available online: http://piklprado.github.io/sads/ (accessed on 22 July 2022).
- Senn-Irlet, B.; Heilmann-Clausen, J.; Genney, D.; Dahlberg, A. Guidance for Conservation of Macrofungi in Europe. In Document prepared for the European Council for Conservation of Fungi (ECCF) within the European Mycological Association (EMA) and the Directorate of Culture and Cultural and Natural Heritage; Council of Europe: Strasbourg, France, 2007. [Google Scholar]
- Barnes, M.L.; Bodin, Ö.; McClanahan, T.; Kittinger, J.N.; Hoey, A.S.; Gaoue, O.G.; Graham, N.A. Social-ecological alignment and ecological conditions in coral reefs. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavoie, C. Biological Collections in an Ever Changing World: Herbaria as Tools for Biogeographical and Environmental Studies. Perspect. Plant Ecol. Evol. Syst. 2013, 15, 68–76. [Google Scholar] [CrossRef]
- Bidaud, A.; Loizides, M.; Armada, F.; García, J.; Carteret, X.; Corriol, G.; Consiglio, G.; Reumaux, P.; Bellanger, J.-M. Cortinarius Subgenus Leprocybe in Europe: Expanded Sanger and Next Generation Sequencing Unveil Unexpected Diversity in the Mediterranean. Pers.—Mol. Phylogeny Evol. Fungi 2021, 46, 188. [Google Scholar]
- Lõhmus, A. Factors of Species-Specific Detectability in Conservation Assessments of Poorly Studied Taxa: The Case of Polypore Fungi. Biol. Conserv. 2009, 142, 2792–2796. [Google Scholar] [CrossRef]
- Malaval, J.-C. Liste rouge Ascomycotina, Basidiomycotina, Myxostelidae menacés de Haute-Normandie. Bull. Société Amis Sci. Nat. Muséum Rouen 2000, 2000–2001, 73–90. Available online: https://sasnmr.fr/ressources/le-coin-des-lecteurs (accessed on 22 July 2022).
- Dahlberg, A.; Mueller, G.M. Applying IUCN Red-Listing Criteria for Assessing and Reporting on the Conservation Status of Fungal Species. Fungal Ecol. 2011, 4, 147–162. [Google Scholar] [CrossRef]
- O’Dell, T.; Ammirati, J.; Schreiner, E. Species Richness and Abundance of Ectomycorrhizal Basidiomycete Sporocarps on a Moisture Gradient in the Tsuga Heterophylla Zone. Can. J. Bot. 1999, 77, 1699–1711. [Google Scholar] [CrossRef]
- Martínez de Aragón, J.; Bonet, J.A.; Fischer, C.; Colinas, C. Productivity of Ectomycorrhizal and Selected Edible Saprotrophic Fungi in Pine Forests of the Pre-Pyrenees Mountains, Spain: Predictive Equations for Forest Management of Mycological Resources. For. Ecol. Manag. 2007, 252, 239–256. [Google Scholar] [CrossRef]
- Straatsma, G.; Ayer, F.; Egli, S. Species Richness, Abundance, and Phenology of Fungal Fruit Bodies over 21 Years in a Swiss Forest Plot. Mycol. Res. 2001, 105, 515–523. [Google Scholar] [CrossRef]
- Egli, S.; Peter, M.; Buser, C.; Stahel, W.; Ayer, F. Mushroom Picking Does Not Impair Future Harvests—Results of a Long-Term Study in Switzerland. Biol. Conserv. 2006, 129, 271–276. [Google Scholar] [CrossRef]
- Kranabetter, J.M.; Durall, D.M.; MacKenzie, W.H. Diversity and Species Distribution of Ectomycorrhizal Fungi along Productivity Gradients of a Southern Boreal Forest. Mycorrhiza 2009, 19, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2010. [Google Scholar]
- Kertész, Á.; Madarász, B. Conservation Agriculture in Europe. Int. Soil Water Conserv. Res. 2014, 2, 91–96. [Google Scholar] [CrossRef]
- Richard, F.; Moreau, P.-A.; Selosse, M.-A.; Gardes, M. Diversity and Fruiting Patterns of Ectomycorrhizal and Saprobic Fungi in an Old-Growth Mediterranean Forest Dominated by Quercus ilex L. Can. J. Bot. 2004, 82, 1711–1729. [Google Scholar] [CrossRef] [Green Version]
- Baptista, P.; Martins, A.; Tavares, R.M.; Lino-Neto, T. Diversity and Fruiting Pattern of Macrofungi Associated with Chestnut (Castanea sativa) in the Trás-Os-Montes Region (Northeast Portugal). Fungal Ecol. 2010, 3, 9–19. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Ecoregion | ||||
|---|---|---|---|---|---|
| Atlantic | Continental | Mediterranean | Montane | Total France | |
| Number of observations (n) | |||||
| Total | 492,014 | 342,124 | 35,202 | 157,558 | 1,043,262 |
| Mean per department | 10,696 a | 11,404 a | 3200 b | 17,506 a | |
| Standard deviation | 16,343.08 | 18,262.98 | 2594.12 | 18,861.16 | |
| Observation density (n·km−2) | |||||
| Mean per department | 2.06 a | 2.72 a | 0.58 b | 3.18 a | |
| Standard deviation | 2.68 | 4.71 | 0.41 | 3.28 | |
| Specific richness (S) | |||||
| Mean per department | 1274.2 a | 1400.03 a | 1045.46 a | 2099.56 a | 12,055 |
| Standard deviation | 941.32 | 808.49 | 503 | 1605.13 | |
| Rarefied species richness | 3632.17 | 3686.14 | 4400 | 478.96 | |
| Sampling effort | |||||
| Number of communes | 16,815 | 12,258 | 2771 | 2992 | 34,836 |
| Number of sampled communes | 5147 | 3912 | 1081 | 1396 | 11,536 |
| % of sampled communes | 30.6 | 31.9 | 39 | 46.7 | 33.1 |
| Coefficient | Estimate | Standard Error | z Value | p-Value |
|---|---|---|---|---|
| (a) | ||||
| Intercept | −4.04 | 2.97 | −1.36 | 0.17 |
| Nb of records (log) | 1.20 | 0.35 | 3.39 | 0.0007 *** |
| Nb of inhabitants (log) | 0.53 | 0.23 | 2.29 | 0.02 * |
| Nb of contributors | 0.0344 | 0.02 | 1.56 | 0.12 |
| Nb of records (log) x nb of inhabitants (log) | −0.05 | 0.03 | −1.98 | 0.04 * |
| (b) | ||||
| Intercept | −2.60 | 1.13 | −2.30 | 0.02 * |
| Record density (log) | 0.50 | 0.16 | 3.12 | 0.0018 ** |
| Density of inhabitants (log) | 0.25 | 0.24 | 1.05 | 0.29 |
| Nb of contributors | −0.01 | 0.08 | −0.09 | 0.93 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gautier, M.; Moreau, P.-A.; Boury, B.; Richard, F. Unravelling the French National Fungal Database: Geography, Temporality, Taxonomy and Ecology of the Recorded Diversity. J. Fungi 2022, 8, 926. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8090926
Gautier M, Moreau P-A, Boury B, Richard F. Unravelling the French National Fungal Database: Geography, Temporality, Taxonomy and Ecology of the Recorded Diversity. Journal of Fungi. 2022; 8(9):926. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8090926
Chicago/Turabian StyleGautier, Montan, Pierre-Arthur Moreau, Béatrice Boury, and Franck Richard. 2022. "Unravelling the French National Fungal Database: Geography, Temporality, Taxonomy and Ecology of the Recorded Diversity" Journal of Fungi 8, no. 9: 926. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8090926