Germline Genetic Association between Stromal Interaction Molecule 1 (STIM1) and Clinical Outcomes in Breast Cancer Patients

 , ,
, ,
Abstract
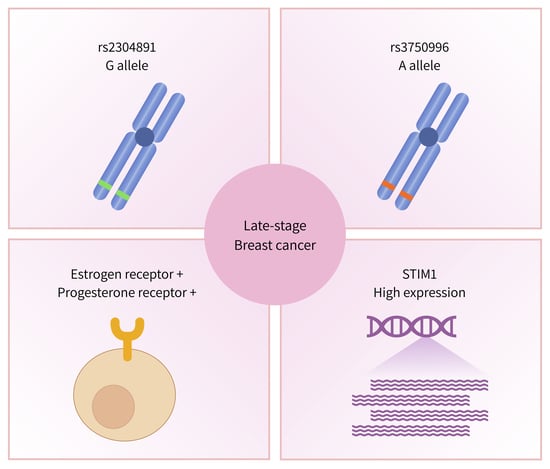
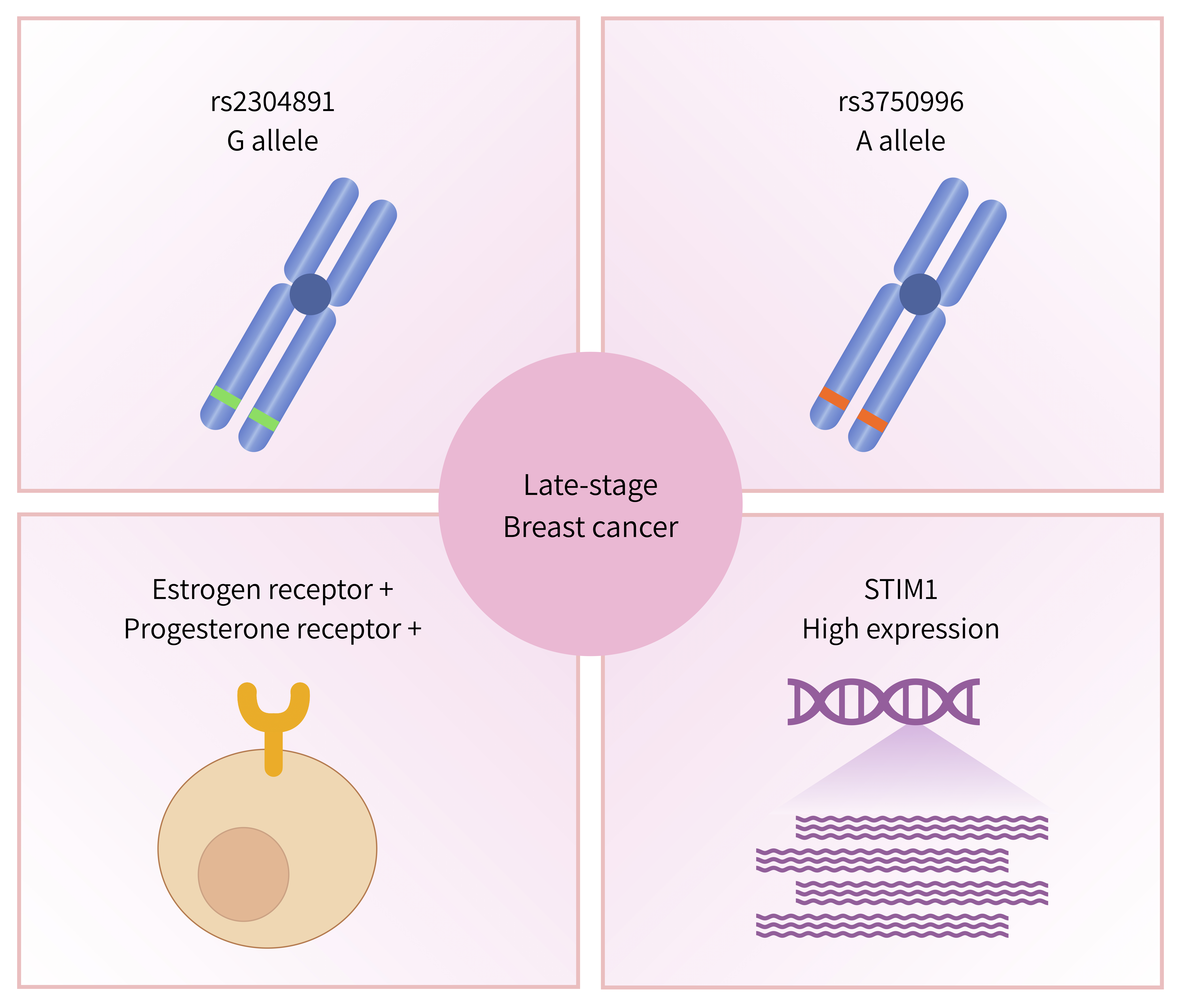
:
1. Introduction
2. Materials and Methods
2.1. Study Cohorts
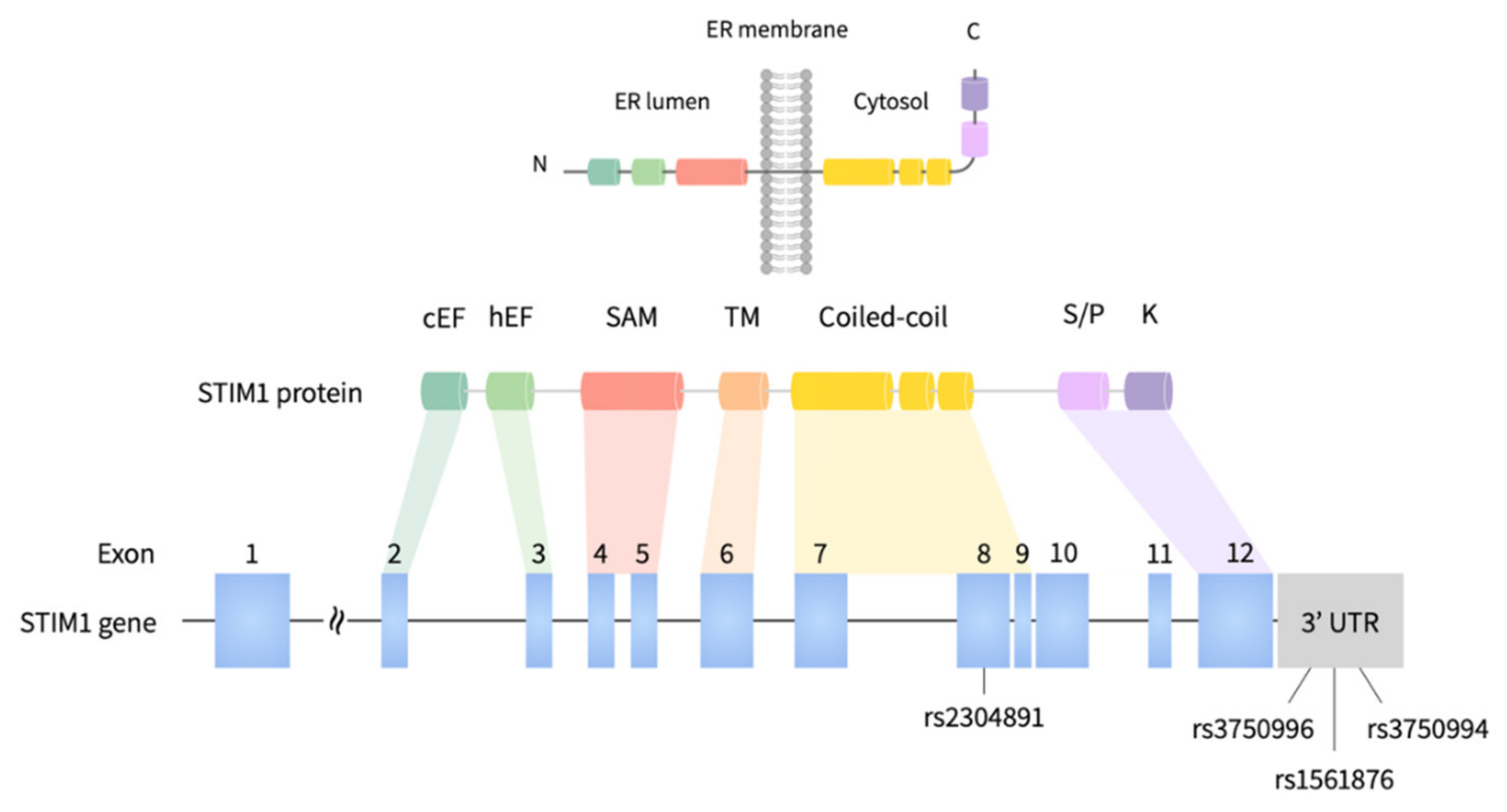
2.2. Selection of Germline Tagging Single Nucleotide Variants (SNVs)
2.3. TaqMan Genotyping Assay
2.4. Statistical Analysis
3. Results
3.1. Basal Characteristic of the Breast Cancer Patients
3.2. Association of STIM1 Genetic Variants with BC Disease Staging
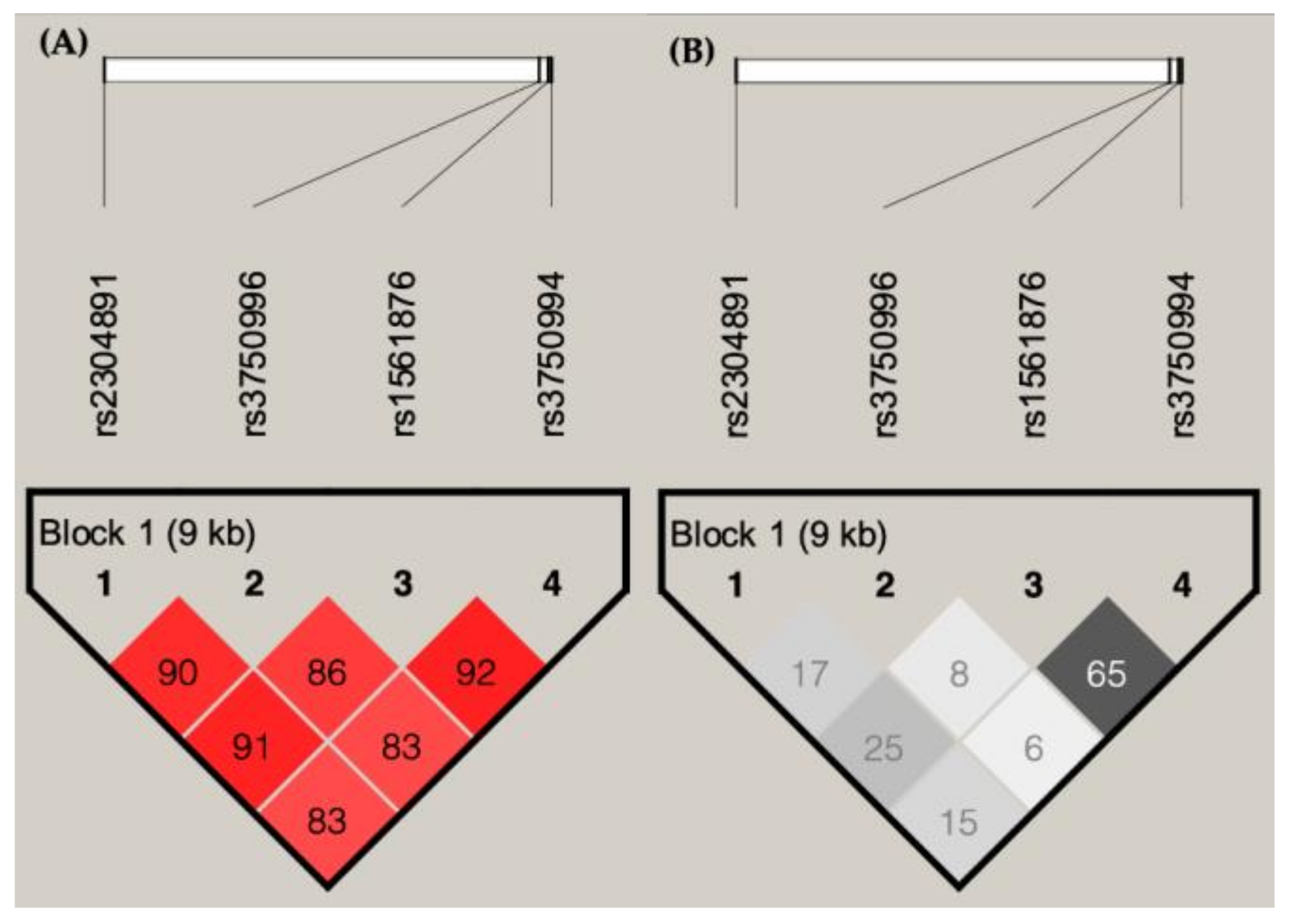
3.3. Association of STIM1 Haplotypes with Disease Staging in BC Patient
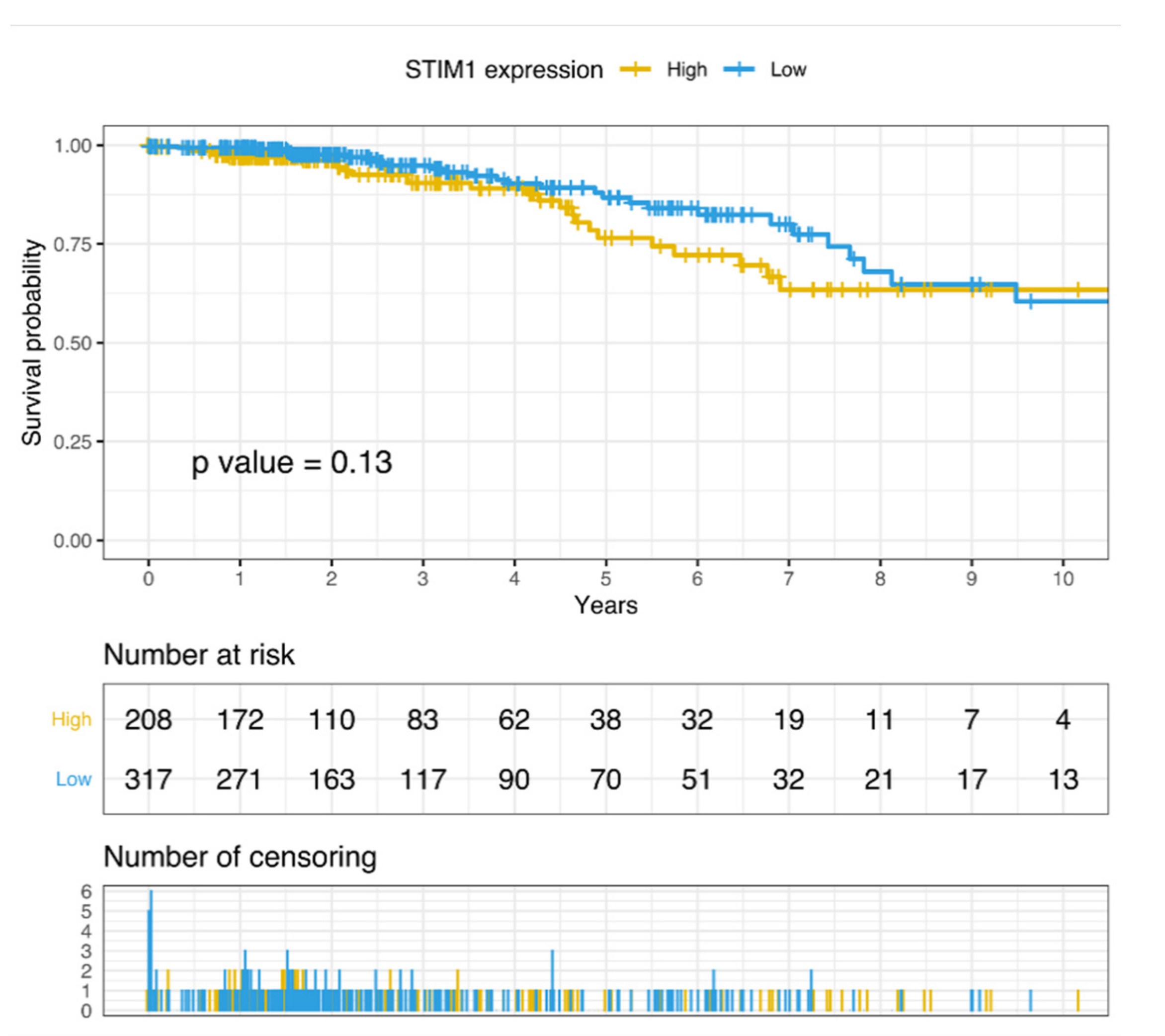
3.4. Correlation between STIM1 Expression and the Survival of Breast Cancer Patients
3.5. eQTL Analysis of rs2304891 Confirmed the Correlations between Genotypes and Tissue-Specific Gene Expression Levels
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ghoncheh, M.; Pournamdar, Z.; Salehiniya, H. Incidence and Mortality and Epidemiology of Breast Cancer in the World. Asian Pac. J. Cancer Prev. 2016, 17, 43–46. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.N.; Liao, Y.M.; Kuo, L.N.; Tsai, H.J.; Chang, W.C.; Yen, Y. Cancers in Taiwan: Practical insight from epidemiology, treatments, biomarkers, and cost. J. Formos Med. Assoc. 2020, 119, 1731–1741. [Google Scholar] [CrossRef]
- Goetz, M.P.; Gradishar, W.J.; Anderson, B.O.; Abrahacm, J.; Aft, R.; Allison, K.H.; Blair, S.L.; Burstein, H.J.; Dang, C.; Elias, A.D.; et al. NCCN Guidelines Insights: Breast Cancer, Version 3.2018. J. Natl. Compr. Canc. Netw. 2019, 17, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Anderson, K.N.; Schwab, R.B.; Martinez, M.E. Reproductive risk factors and breast cancer subtypes: A review of the literature. Breast Cancer Res. Treat. 2014, 144, 1–10. [Google Scholar] [CrossRef]
- Dai, X.; Li, T.; Bai, Z.; Yang, Y.; Liu, X.; Zhan, J.; Shi, B. Breast cancer intrinsic subtype classification, clinical use and future trends. Am. J. Cancer Res. 2015, 5, 2929–2943. [Google Scholar]
- Miller, K.D.; Siegel, R.L.; Lin, C.C.; Mariotto, A.B.; Kramer, J.L.; Rowland, J.H.; Stein, K.D.; Alteri, R.; Jemal, A. Cancer treatment and survivorship statistics, 2016. CA Cancer J. Clin. 2016, 66, 271–289. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.C.; Tu, S.H.; Lien, H.H.; Jeng, J.Y.; Huang, C.S.; Huang, C.J.; Lai, L.C.; Chuang, E.Y. Concurrent gene signatures for han chinese breast cancers. PLoS ONE 2013, 8, e76421. [Google Scholar] [CrossRef] [Green Version]
- Slade, D. PARP and PARG inhibitors in cancer treatment. Genes Dev. 2020, 34, 360–394. [Google Scholar] [CrossRef] [Green Version]
- Prevarskaya, N.; Skryma, R.; Shuba, Y. Calcium in tumour metastasis: New roles for known actors. Nat. Rev. Cancer 2011, 11, 609–618. [Google Scholar] [CrossRef]
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11–21. [Google Scholar] [CrossRef]
- Ray, K. Calcium-Sensing Receptor: Trafficking, Endocytosis, Recycling, and Importance of Interacting Proteins. Prog. Mol. Biol. Transl. Sci. 2015, 132, 127–150. [Google Scholar] [CrossRef]
- Schulz, R.A.; Yutzey, K.E. Calcineurin signaling and NFAT activation in cardiovascular and skeletal muscle development. Dev. Biol. 2004, 266, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Zhang, J.J.; Huang, X.Y. Orai1 and STIM1 are critical for breast tumor cell migration and metastasis. Cancer Cell 2009, 15, 124–134. [Google Scholar] [CrossRef] [Green Version]
- Roos, J.; DiGregorio, P.J.; Yeromin, A.V.; Ohlsen, K.; Lioudyno, M.; Zhang, S.; Safrina, O.; Kozak, J.A.; Wagner, S.L.; Cahalan, M.D.; et al. STIM1, an essential and conserved component of store-operated Ca2+ channel function. J. Cell Biol. 2005, 169, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Liou, J.; Kim, M.L.; Heo, W.D.; Jones, J.T.; Myers, J.W.; Ferrell, J.E., Jr.; Meyer, T. STIM is a Ca2+ sensor essential for Ca2+-store-depletion-triggered Ca2+ influx. Curr. Biol. 2005, 15, 1235–1241. [Google Scholar] [CrossRef] [Green Version]
- Giachini, F.R.; Webb, R.C.; Tostes, R.C. STIM and Orai proteins: Players in sexual differences in hypertension-associated vascular dysfunction? Clin. Sci. (Lond.) 2009, 118, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.L.; Yeromin, A.V.; Zhang, X.H.; Yu, Y.; Safrina, O.; Penna, A.; Roos, J.; Stauderman, K.A.; Cahalan, M.D. Genome-wide RNAi screen of Ca(2+) influx identifies genes that regulate Ca(2+) release-activated Ca(2+) channel activity. Proc. Natl. Acad. Sci. USA 2006, 103, 9357–9362. [Google Scholar] [CrossRef] [Green Version]
- Vig, M.; Peinelt, C.; Beck, A.; Koomoa, D.L.; Rabah, D.; Koblan-Huberson, M.; Kraft, S.; Turner, H.; Fleig, A.; Penner, R.; et al. CRACM1 is a plasma membrane protein essential for store-operated Ca2+ entry. Science 2006, 312, 1220–1223. [Google Scholar] [CrossRef] [Green Version]
- Lewis, R.S. The molecular choreography of a store-operated calcium channel. Nature 2007, 446, 284–287. [Google Scholar] [CrossRef]
- Sun, J.; Lu, F.; He, H.; Shen, J.; Messina, J.; Mathew, R.; Wang, D.; Sarnaik, A.A.; Chang, W.C.; Kim, M.; et al. STIM1- and Orai1-mediated Ca(2+) oscillation orchestrates invadopodium formation and melanoma invasion. J. Cell Biol. 2014, 207, 535–548. [Google Scholar] [CrossRef] [Green Version]
- Ho, K.H.; Chang, C.K.; Chen, P.H.; Wang, Y.J.; Chang, W.C.; Chen, K.C. miR-4725-3p targeting stromal interacting molecule 1 signaling is involved in xanthohumol inhibition of glioma cell invasion. J. Neurochem. 2018, 146, 269–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.Y.; Sun, J.; Huang, M.Y.; Wang, Y.S.; Hou, M.F.; Sun, Y.; He, H.; Krishna, N.; Chiu, S.J.; Lin, S.; et al. STIM1 overexpression promotes colorectal cancer progression, cell motility and COX-2 expression. Oncogene 2015, 34, 4358–4367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, I.H.; Tsai, Y.T.; Chiu, S.J.; Liu, L.T.; Lee, H.H.; Hou, M.F.; Hsu, W.L.; Chen, B.K.; Chang, W.C. Involvement of STIM1 and Orai1 in EGF-mediated cell growth in retinal pigment epithelial cells. J. Biomed. Sci. 2013, 20, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.C.; Chai, C.Y.; Chen, W.C.; Hou, M.F.; Wang, Y.S.; Chiu, Y.C.; Lu, S.R.; Chang, W.C.; Juo, S.H.; Wang, J.Y.; et al. Histamine regulates cyclooxygenase 2 gene activation through Orai1-mediated NFκB activation in lung cancer cells. Cell Calcium 2011, 50, 27–35. [Google Scholar] [CrossRef]
- Wang, J.Y.; Chen, B.K.; Wang, Y.S.; Tsai, Y.T.; Chen, W.C.; Chang, W.C.; Hou, M.F.; Wu, Y.C.; Chang, W.C. Involvement of store-operated calcium signaling in EGF-mediated COX-2 gene activation in cancer cells. Cell Signal 2012, 24, 162–169. [Google Scholar] [CrossRef]
- Chang, W.C.; Parekh, A.B. Close functional coupling between Ca2+ release-activated Ca2+ channels, arachidonic acid release, and leukotriene C4 secretion. J. Biol. Chem. 2004, 279, 29994–29999. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.C.; Nelson, C.; Parekh, A.B. Ca2+ influx through CRAC channels activates cytosolic phospholipase A2, leukotriene C4 secretion, and expression of c-fos through ERK-dependent and -independent pathways in mast cells. FASEB J. 2006, 20, 2381–2383. [Google Scholar] [CrossRef]
- Parekh, A.B.; Putney, J.W., Jr. Store-operated calcium channels. Physiol. Rev. 2005, 85, 757–810. [Google Scholar] [CrossRef] [Green Version]
- Edge, S.B.; Compton, C.C. The American Joint Committee on Cancer: The 7th edition of the AJCC cancer staging manual and the future of TNM. Ann. Surg. Oncol. 2010, 17, 1471–1474. [Google Scholar] [CrossRef]
- Network, C.G.A. Comprehensive molecular portraits of human breast tumours. Nature 2012, 490, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Colaprico, A.; Silva, T.C.; Olsen, C.; Garofano, L.; Cava, C.; Garolini, D.; Sabedot, T.S.; Malta, T.M.; Pagnotta, S.M.; Castiglioni, I.; et al. TCGAbiolinks: An R/Bioconductor package for integrative analysis of TCGA data. Nucleic Acids Res. 2016, 44, e71. [Google Scholar] [CrossRef] [PubMed]
- Yates, A.D.; Achuthan, P.; Akanni, W.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; et al. Ensembl 2020. Nucleic Acids Res. 2020, 48, D682–D688. [Google Scholar] [CrossRef]
- Consortium, G. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.C.; Hung, K.S.; Hsu, Y.W.; Wong, R.H.; Huang, C.H.; Jan, M.S.; Wu, S.J.; Juan, Y.S.; Chang, W.C. Genetic polymorphisms of stromal interaction molecule 1 associated with the erythrocyte sedimentation rate and C-reactive protein in HLA-B27 positive ankylosing spondylitis patients. PLoS ONE 2012, 7, e49698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.C.; Lee, C.H.; Hirota, T.; Wang, L.F.; Doi, S.; Miyatake, A.; Enomoto, T.; Tomita, K.; Sakashita, M.; Yamada, T.; et al. ORAI1 genetic polymorphisms associated with the susceptibility of atopic dermatitis in Japanese and Taiwanese populations. PLoS ONE 2012, 7, e29387. [Google Scholar] [CrossRef] [Green Version]
- Onouchi, Y.; Fukazawa, R.; Yamamura, K.; Suzuki, H.; Kakimoto, N.; Suenaga, T.; Takeuchi, T.; Hamada, H.; Honda, T.; Yasukawa, K.; et al. Variations in ORAI1 Gene Associated with Kawasaki Disease. PLoS ONE 2016, 11, e0145486. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Guo, H.; Chen, D.; Chen, X.; Jin, Q. MUTYH and ORAI1 polymorphisms are associated with susceptibility to osteoarthritis in the Chinese Han population. J. Int. Med. Res. 2018, 46, 2292–2300. [Google Scholar] [CrossRef] [Green Version]
- Abbate, A.; Trankle, C.R.; Buckley, L.F.; Lipinski, M.J.; Appleton, D.; Kadariya, D.; Canada, J.M.; Carbone, S.; Roberts, C.S.; Abouzaki, N.; et al. Interleukin-1 Blockade Inhibits the Acute Inflammatory Response in Patients With ST-Segment-Elevation Myocardial Infarction. J. Am. Heart Assoc. 2020, 9, e014941. [Google Scholar] [CrossRef]
- Thiha, K.; Mashimo, Y.; Suzuki, H.; Hamada, H.; Hata, A.; Hara, T.; Tanaka, T.; Ito, K.; Onouchi, Y. Investigation of novel variations of ORAI1 gene and their association with Kawasaki disease. J. Hum. Genet. 2019, 64, 511–519. [Google Scholar] [CrossRef]
- Clarke, R.; Leonessa, F.; Welch, J.N.; Skaar, T.C. Cellular and molecular pharmacology of antiestrogen action and resistance. Pharmacol. Rev. 2001, 53, 25–71. [Google Scholar] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgarten, S.C.; Frasor, J. Minireview: Inflammation: An instigator of more aggressive estrogen receptor (ER) positive breast cancers. Mol. Endocrinol. 2012, 26, 360–371. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Jiang, Z.; Ma, N.; Wang, B.; Liu, J.; Zhang, L.; Gu, L. MicroRNA-223 Targeting STIM1 Inhibits the Biological Behavior of Breast Cancer. Cell. Physiol. Biochem. 2018, 45, 856–866. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Total No. = 348 |
|---|---|
| Gender (female; no.) | 348 (100) |
| Age (years old, mean ± sd (range)) | 53.13 ± 11.17 (23–89) |
| ER a (no.) | |
| Positive | 231 (66.38) |
| Negative | 64 (18.39) |
| n.a. | 53 (15.23) |
| PR b (no.) | |
| Positive | 189 (54.31) |
| Negative | 105 (30.17) |
| n.a. | 54 (15.52) |
| HER2 c (no.) | |
| Positive | 195 (56.03) |
| Negative | 98 (28.16) |
| n.a. | 55 (15.80) |
| AJCC d/TNM e stage (no.) | |
| 1 | 138 (39.66) |
| 2 | 97 (27.87) |
| 3 | 48 (13.79) |
| n.a. | 65 (18.68) |
| Population Genotypic Frequency a | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| dbSNV ID | Genotype | Chr:Pos b | AFR | AMR | EUR | SAS | EAS | TWB | Our Cohort |
| rs2304891 | G/G | 11:4103524 | 0.020 | 0.236 | 0.336 | 0.225 | 0.210 | 0.199 | 0.183 |
| A/G | 0.156 | 0.496 | 0.463 | 0.503 | 0.464 | 0.481 | 0.477 | ||
| A/A | 0.825 | 0.268 | 0.201 | 0.272 | 0.325 | 0.320 | 0.340 | ||
| rs3750996 | G/G | 11:4113200 | 0 | 0 | 0 | 0.010 | 0.060 | 0.050 | 0.081 |
| A/G | 0 | 0.003 | 0 | 0.147 | 0.315 | 0.339 | 0.325 | ||
| A/A | 1 | 0.997 | 1 | 0.843 | 0.625 | 0.611 | 0.594 | ||
| rs1561876 | G/G | 11:4113395 | 0.498 | 0.061 | 0.012 | 0.004 | 0.067 | 0.079 | 0.059 |
| A/G | 0.415 | 0.326 | 0.209 | 0.182 | 0.331 | 0.381 | 0.422 | ||
| A/A | 0.088 | 0.614 | 0.779 | 0.814 | 0.601 | 0.540 | 0.519 | ||
| rs3750994 | G/G | 11:4113470 | 0 | 0.012 | 0.002 | 0 | 0.052 | 0.054 | 0.031 |
| T/G | 0.015 | 0.205 | 0.056 | 0.055 | 0.282 | 0.323 | 0.350 | ||
| T/T | 0.985 | 0.784 | 0.942 | 0.945 | 0.667 | 0.623 | 0.619 | ||
| dbSNV ID | Genotype | Late Stage a N (%) | Early Stage b N (%) | Genotype Model | |
|---|---|---|---|---|---|
| OR (95% C.I.) c | p Value | ||||
| rs2304891 | G/G | 15 (31.9) | 37 (16.4) | 4.48 (1.73~11.62) | 5.17 × 10−3 * |
| A/G | 24 (51.1) | 104 (46.2) | 2.49 (1.06~5.86) | ||
| A/A | 8 (17.0) | 84 (37.3) | Reference | ||
| rs3750996 | G/G | 0 (0) | 19 (8.3) | 0 | 5.50 × 10−4 ** |
| A/G | 8 (17.0) | 75 (32.9) | 0.35 (0.15~0.79) | ||
| A/A | 39 (83.0) | 134 (58.8) | Reference | ||
| rs1561876 | G/G | 1 (2.3) | 16 (7.3) | 0.42 (0.05~3.40) | 0.2631 |
| A/G | 24 (54.5) | 94 (42.9) | 1.47 (0.76~2.87) | ||
| A/A | 19 (43.2) | 109 (49.8) | Reference | ||
| rs3750994 | G/G | 2 (4.4) | 10 (4.4) | 1.24 (0.25~6.15) | 0.9488 |
| T/G | 17 (37.8) | 81 (35.8) | 1.08 (0.55~2.12) | ||
| T/T | 26 (57.8) | 135 (59.7) | Reference | ||
| dbSNV ID | Genotype | Late Stage a N (%) | Early Stage b N (%) | Genotype Model | |
|---|---|---|---|---|---|
| OR (95% C.I.) c | p Value | ||||
| rs2304891 | G/G | 11 (33.3) | 20 (14.4) | 9.86 (2.47~39.39) | 9.18 × 10−4 ** |
| A/G | 19 (57.6) | 67 (48.2) | 4.97 (1.39~17.75) | ||
| A/A | 3 (9.1) | 52 (37.4) | Reference | ||
| rs3750996 | G/G | 0 (0) | 10 (7.0) | 0 | 3.16 × 10−3 ** |
| A/G | 5 (15.2) | 51 (35.7) | 0.28 (0.10~0.78) | ||
| A/A | 28 (84.8) | 82 (57.3) | Reference | ||
| rs1561876 | G/G | 0 (0) | 11 (8.1) | 0 | 0.0909 |
| A/G | 16 (53.3) | 59 (43.7) | 1.26 (0.56~2.80) | ||
| A/A | 14 (46.7) | 65 (48.1) | Reference | ||
| rs3750994 | G/G | 1 (3.2) | 5 (3.5) | 0.87 (0.09~8.04) | 0.9897 |
| T/G | 11 (35.5) | 52 (36.4) | 0.97 (0.42~2.20) | ||
| T/T | 19 (61.3) | 86 (60.1) | Reference | ||
| Haplotype a | Frequency | OR (95% C.I.) d | p Value e | |
|---|---|---|---|---|
| rs2304891/rs3750996/rs1561876/rs3750994 | Late Stage b | Early Stage c | ||
| G/A/A/T | 0.548 | 0.367 | 3.84 (1.63~9.03) | 0.0028 * |
| A/A/G/G | 0.222 | 0.201 | 2.84 (1.11~7.28) | 0.6631 |
| A/A/G/T | 0.065 | 0.066 | 2.74 (0.74~10.09) | 0.9261 |
| A/A/A/T | 0.050 | 0.097 | 1.49 (0.41~5.37) | 0.2104 |
| A/G/A/T | 0.088 | 0.230 | Reference | Reference |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.-C.; Lin, M.-R.; Yang, Y.-C.; Hsu, Y.-W.; Wong, H.S.-C.; Chang, W.-C. Germline Genetic Association between Stromal Interaction Molecule 1 (STIM1) and Clinical Outcomes in Breast Cancer Patients. J. Pers. Med. 2020, 10, 287. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm10040287
Huang C-C, Lin M-R, Yang Y-C, Hsu Y-W, Wong HS-C, Chang W-C. Germline Genetic Association between Stromal Interaction Molecule 1 (STIM1) and Clinical Outcomes in Breast Cancer Patients. Journal of Personalized Medicine. 2020; 10(4):287. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm10040287
Chicago/Turabian StyleHuang, Chi-Cheng, Min-Rou Lin, Yu-Chen Yang, Yu-Wen Hsu, Henry Sung-Ching Wong, and Wei-Chiao Chang. 2020. "Germline Genetic Association between Stromal Interaction Molecule 1 (STIM1) and Clinical Outcomes in Breast Cancer Patients" Journal of Personalized Medicine 10, no. 4: 287. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm10040287