
Toward a Systematic Assessment of Sex Differences in Cystic Fibrosis
by
 , , , and
, , , and
Christiane Gärtner
1,2,3,*,
Jörg Fallmann
2,
Peter F. Stadler
2,
Thorsten Kaiser
3 and
Sarah J. Berkemer
4,5 1
Neuromorphic Information Processing, Institute of Computer Science, Leipzig University, Augustusplatz 10, D-04109 Leipzig, Germany
2
Bioinformatics Group, Institute of Computer Science, Interdisciplinary Center of Bioinformatics, Leipzig University, Härtelstraße 16-18, D-04107 Leipzig, Germany
3
Academic Department of Laboratory Medicine, Microbiology and Pathobiochemistry, Medical School and University Medical Center East Westphalia-Lippe, Hospital Lippe, Bielefeld University, Röntgenstraße 18, D-32756 Detmold, Germany
4
LIX CNRS UMR 7161, Ecole Polytechnique, Institut Polytechnique de Paris, 91120 Palaiseau, France
5
Earth-Life Science Institute, Tokyo Institute of Technology, 2-12-1-I7E-318 Ookayama, Tokyo 152-8550, Japan
*
Author to whom correspondence should be addressed.
J. Pers. Med. 2023, 13(6), 924; https://0-doi-org.brum.beds.ac.uk/10.3390/jpm13060924
Submission received: 30 April 2023
/
Revised: 26 May 2023
/
Accepted: 27 May 2023
/
Published: 31 May 2023
(This article belongs to the Special Issue Personalized Medicine Interdisciplinary Aspects and Sex Gender-Specific Differences)
Abstract
:(1) Background: Cystic fibrosis (CF) is a disease with well-documented clinical differences between female and male patients. However, this gender gap is very poorly studied at the molecular level. (2) Methods: Expression differences in whole blood transcriptomics between female and male CF patients are analyzed in order to determine the pathways related to sex-biased genes and assess their potential influence on sex-specific effects in CF patients. (3) Results: We identify sex-biased genes in female and male CF patients and provide explanations for some sex-specific differences at the molecular level. (4) Conclusion: Genes in key pathways associated with CF are differentially expressed between sexes, and thus may account for the gender gap in morbidity and mortality in CF.
1. Introduction
Cystic fibrosis (CF) is the most common inherited disease in the Caucasian population [1]. CF is caused by a mutation(s) in the cystic fibrosis transmembrane conductance regulator (CFTR) protein, which results in defects in the expression or activity of a chloride channel located in the cell membrane. CF affects all exocrine organs but mainly affects the lungs [2]. Severe symptoms include decreased chloride secretion, reduced airway surface liquid height, and increased mucus viscosity. This can lead to a range of CF-related health issues such as bacterial proliferation, sustained inflammation, epithelial lung tissue injury, fibrosis, and remodeling. Further problems include the development of CF-related diabetes (due to destroyed islets of Langerhans), high infertility rates, and malnutrition, as well as low socioeconomic status and decreased quality of life [2].
CF is a disease with well-documented clinical differences between female and male patients. This “gender gap”, however, is very poorly studied at the molecular level. We are aware of only a single study that addresses sex-biased gene expression in CF patients [3]. This contribution summarizes the current knowledge of sex-related differences in CF and analyzes RNA-seq data to identify patterns of differential gene expression that may be related to the known differences in clinical outcomes. Furthermore, it shows that the pathways that play key roles in CF and harbor genes with sex-biased expression can provide explanations for some sex-specific differences.
1.1. Sex Differences in CF
Women experience more severe effects of CF. The observed sex-related differences related to morbidity and mortality in CF include an extended length of hospitalization for women due to more complex pulmonary exacerbation treatment regimens. This may be a direct effect of the higher infection rate observed in women for, e.g., Pseudomonas aeruginosa or methicillin-resistant Staphylococcus aureus (MRSA) [4]. CF-related diabetes (CFRD) can lead to more severe complications, a more significant decline in lung function, and an increased mortality rate in women [5,6]. All the observed effects of CF contribute to a lower median survival age in women (49 years) compared to men (56 years) [7]. These sex-related differences have also been reported in other respiratory diseases [8]. Despite the existence of anatomical differences [9], sex hormones are assumed to be responsible for most of the symptoms, with significantly different effects observed between men and women [2].
1.2. Possible Explanations for the “Gender Gap”
Anatomical differences, in particular, the reduced airway diameter and lung volume in women, have been described as putative reasons for the differences in CF symptoms and complications [2,9]. Other differences have been described that can exacerbate asthma and autoimmune conditions in women such as more robust T-cell immunity, exuberant T-helper (Th)-2 response, eosinophilic inflammation, and IL-33 production [2]. Further risk factors include nutritional differences, which result in a body mass index (BMI) that is lower in female CF patients [4]. Social and behavioral studies show that women are more resistant to nutritional interventions and experience poorer medication and nutritional adherence. It is well-known that a reduced BMI and poor nutritional status contribute to decreased lung function and increased mortality [2].
1.3. Influence of Sexual Hormones on CF
Despite all the above-mentioned anatomical, social, and behavioral factors, recent developments show that puberty has an effect on CF and strongly indicate that being of the female sex is an independent risk factor in CF patients [10,11]. The following summarizes the recent results of studies regarding the influence of sex hormones on CF cells. For a more detailed summary of existing animal and human studies, see Table 1 in [2].
The effects of estrogen are mediated via estrogen receptors 1 and 2 (ESR1 and ESR2) and the 7-transmembrane G protein-coupled estrogen receptor (GPER). ESR1 and ESR2 are ligand-activated transcription factors that affect, e.g., proteins involved in cell cycle regulation and inflammation. On the other hand, GPER interacts with the epidermal growth factor receptor (EGFR) in transactivation, and it also influences a diverse array of signaling pathways [12]. In a microarray study by Clarke et al. [13] in human native nasal epithelial cells from CF patients compared to non-CF controls, ESR1 was upregulated 1.84-fold in CF patients. Additional targets for ESR1 were significantly enriched in samples from CF patients, whereas differential expression levels of both ESR2 and GPER were not observed.
In vitro tests have shown that estrogen decreases the innate immune response to bacterial infections [2]. In mice, the increased expression of IL-23 and IL-17 has been reported in response to estradiol stimulation in CFTR-deficient cells [14]. Hence, estrogen may directly alter bacterial behavior. In an in vitro study, it was shown that estrogen promotes the growth of P. aeruginosa, and this effect may vary depending on the phase of the menstrual cycle. Moreover, estrogen was found to reduce the intensity of ciliary beat frequency [15]. In another study, serum estradiol levels were found to be significantly higher during periods of CF exacerbation [16]. Further in vitro studies in human CF bronchial epithelial cells identified an estradiol-dependent upregulation of SLPI and an inhibition of IL-8, together with decreased ciliary beat frequency and reduced chloride transport in a CFTR-independent manner.
In a study focusing on the clinical outcomes for humans (no gene expression analyses), it was shown that treatment with Ivacaftor (CFTR potentiators) resulted in reduced rates of pulmonary exacerbation (PEx) in females. However, no significant sex-related differences were observed in terms of changes in forced expiratory volume in one second (FEV1) or body mass index (BMI) after the conclusion of the study [17].
2. Materials and Methods
2.1. Data Source
Data used in this study were obtained from the NCBI GEO database [18] under accession number GSE205161 [19]. The available data have been filtered to include only individuals without nontuberculous mycobacterial pulmonary disease infections. Additionally, individuals aged 18 and above were chosen to guarantee full adolescence. Next, the datasets were filtered to include individuals of Caucasian heritage, as this was a predominant factor in the available datasets and allowed for better comparison of medical parameters regarding lung status. Of the remainder, 5 female and 5 male whole blood transcriptomics bulk RNA-seq datasets were chosen randomly for this study. The available baseline characteristics of the patients subdivided according to sex are listed in Table 1.
The numbers of uniquely mapped reads were similar between sexes, with the exception of an upwards outlier in female sample no. SRR19451822, which contained 2–4× we confirm the change. more unique mappers compared to the other samples. However, in terms of the relative numbers, all samples contained between 60% and 75% unique mappers.
2.2. Data Processing and Analysis
The downloading, processing, and analysis of the data used in this study were conducted using MONSDA [20] release v1.1.0. The corresponding configuration file is available at https://raw.githubusercontent.com/jfallmann/MONSDA_companion/main/CysticFibrosisGenderBias/config_cystic_fibrosis_monsda.json, accessed on 26 May 2023.
2.2.1. Pre-Processing and Mapping
GEO Series GSE205161 was downloaded using sra-tools [21] version 2.11.0. The raw reads were trimmed using cutadapt [22] version 4.1 and mapped using star [23] version 2.7.10b. Quality control was conducted for the raw, trimmed, and mapped reads using FastQC [24] version 0.11.9 and MultiQC [25] version 1.14.
2.2.2. Detection of Differentially Expressed Genes between Females and Males
To identify differentially expressed genes, the uniquely mapped reads were quantified using FeatureCounts [26] version 1.6.4 and analyzed for DE using DESeq2 [27] version 1.32.0. The expression of genes from GENCODE v37 was compared between CF-diseased female and male samples, with the former regarded as the baseline condition. Genes that were significantly (absolute fold-change > 2, FDR < 0.05) differentially expressed were classified into two categories, male-biased genes (MG) and female-biased genes (FG), based on their overexpression in comparison to the other condition [28]. Therefore, MG is the set of genes overexpressed in CF-diseased males compared to CF-diseased females, and vice versa for FG.
2.2.3. Detection of Sex-Biased Alternatively Spliced Genes
Alternative splicing analysis was conducted using EdgeR [29] version 3.28.0, following the quantification of the exon expression using FeatureCounts [26] version 1.6.4. The definitions of FG and MG are consistent with the information presented in Section 2.2.2.
2.2.4. Network analysis and subcellular locations
3. Results
3.1. Differentially Expressed Genes in Female versus Male Patients with Cystic Fibrosis
We found 1140 genes expressed in a sex-biased manner: 999 male-biased genes (MG) and 141 female-biased genes (FG). The distribution of FG and MG across the different chromosomes is shown in Figure 1. Overall, most sex-biased genes were MG and were found in autosomes; however, some MG were found on the X chromosome. The relative distribution of sex-biased genes to the total number of genes of a chromosome showed that most sex-biased genes were located on chromosome 4 and chromosome 13.
3.2. Enrichment Analysis of Sex-Biased Expressed Genes in CF
During the investigation of enriched interactions, pathways, cellular processes, and cellular compartments within the set of sex-biased expressed genes, significant enrichments of pathways and cellular functions were observed, which provide some insights into CF-specific sex biases. The results are summarized in Table 2.
3.2.1. Pathways
Ideozu et al. [33] summarized several expression studies in CF and showed the following key pathways in CF: cytokine signaling, inflammatory response, cell-to-cell signaling, TLR signaling, chemokine signaling, AMPK-Akt signaling, glycosylation of biopolymers, E1F2 signaling, and IL-8 signaling. Regarding sex-biased genes in CF, our results align with theirs in terms of glycosylation, cell-to-cell signaling, multicellular organismal signaling, cAMP signaling, and the estrogen pathway, as shown in Table 2.
The expression differences of genes encoding for estrogen receptors 1 and 2 (ESR1 and ESR2) were not significant between the sexes according to the strict logFC cutoffs. However, the general expression of ESR2 was very high (base mean 1576) and the logFC at was significant without the cutoffs (adjusted p-value < 0.001). Therefore, assuming relevant effects is not unfounded. ESR1 was also expressed at a high level, although the level was lower compared to ESR2 (base mean 540) with a logFC of . In this case, no statistical significance (adjusted p-value ) was found. G protein-coupled estrogen receptor (GPER1) was found to be expressed only at low levels (base mean ), without any fold changes observed between the sexes.
In addition, genes involved in inflammatory processes (IL-17 signaling) were found to exhibit sex-biased expression. Among the FG, we detected CXCL5, a known chemokine with IL-17 upregulating function. Among the MG, genes were identified that affect the O-glycosylation of TSR domain-containing proteins. These genes play a role in regulating inflammation and are involved in the functioning of NOD-like receptors, which are essential for recognizing pathogen-associated molecular patterns that are crucial for infection response.
3.2.2. Subcellular Location of Sex-Biased Genes
Enrichment analysis showed that the sex-biased genes were located close to or inside the cell membrane, with some genes even being integral components of the membrane. In addition, most of them were MG (18 FG vs. 111 MG), as depicted in Figure 2. Enrichment of this cellular compartment appears conclusive since CF is based on a mutation of an ion channel localized in the cell membrane.
3.3. Alternatively Spliced Genes between Female and Male Patients with Cystic Fibrosis
Table A5 and Table A6 list the genes that were detected as differentially expressed in male and female CF patients. All of these genes showed enrichment in pathways related to alternative splicing based on the results from StringDB. In contrast, our own analysis only showed 33 alternatively spliced genes regarding sex bias (16 MG, 17 FG; see also Table A7 and Table A8). No overlaps between the StringDB results and our set of genes were detected. This is not surprising because the comparison was performed on two levels. The list of MG and FG differentially expressed genes contained some genes that regulate the alternative splicing process, whereas this analysis detected genes that are potential targets of alternative splicing.
4. Discussion
Cystic fibrosis is a disease with well-documented clinical differences between female and male patients. In this contribution, we analyzed the RNA-seq data of five female and five male CF patients in order to detect differentially expressed genes regarding patient sex. The results demonstrate the presence of significantly sex-biased genes. Therefore, sex-specific clinical treatment and medication may decrease differences in clinical outcomes and symptoms and improve the living conditions of CF patients regardless of gender.
There was minimal overlap observed when comparing the overall differential gene expression between healthy female and male subjects, similar to what was found in a study by Talebizadeh et al. [34] on X chromosome gene expression in female and male tissues. Comparing sex-biased genes in healthy individuals revealed a minimal overlap with those identified in CF-affected individuals. Most sex-biased expressed genes in CF patients were not found to be differentially expressed between healthy individuals of both sexes, which highlights the need for specific studies, as the transferability of results is not guaranteed. Only 30 out of 883 FG and 21 out of 894 MG were found in peripheral mononuclear blood cells, overlapping with those of healthy subjects described in Guo et al. [28]. Therefore, a CF-specific sex bias can be derived.
Ogilvie et al. reported a total of 863 differentially expressed genes between bronchial epithelium samples of CF vs. healthy individuals [35], only a fraction of which were found in this study. This was also the case when comparing our results to genes differentially expressed between CF and non-CF patients in the microarray study by Clarke et al. [13], where only a few additional genes were sex-biased expressed. Acknowledging the differences between the technologies used in these studies and the analysis presented here (microarray vs. RNA-Seq), as well as the difference in the tissue under consideration (epithelium vs. plasma), we conclude that sex plays an important role as a confounding factor in differential expression analysis and should not be ignored. Furthermore, the differences between sexes seem to be even more pronounced than the overall differences observed between pooled healthy and diseased subjects.
In severe CF, differentially expressed genes regarding CF vs. non-CF patients were reported to be involved in protein ubiquitination, mitochondrial oxidoreductase activity, and lipid metabolism [36]. Our study identified an overlap in sex-biased expressed genes, specifically CALR3 (Calreticulin, Ca2 binding, and storage) as a male-biased expressed gene and IGFBP3 as a female-biased expressed gene.
When comparing our results with other studies on the gender gap in CF, we found no overlap between the miRNAs involved in inflammatory processes [3] and the sex-biased miRNAs detected here or the list of hub genes identified in the meta-analysis by Trivedi et al. [37]. In that study, a hub gene analysis was conducted on microarray-derived gene expression datasets and hub genes were identified based on a protein–protein interaction network. As stated by the authors, the identified hub genes (MYC, EZR, S100A9, S100A8, TF, TIA1, KYNU, KLF6, CSTA, and LRRFIP1) were present in the IL-17 signaling pathway and the mineral absorption and gastric-acid secretion pathways [37]. However, the study focused on healthy vs. affected individuals and we found no overlaps with the sex-biased genes identified in our study. This implies that hub gene expression is not sex-biased within CF patients. Given their central role in sex-independent molecular mechanisms, this is not surprising.
Analysis of the subcellular location of sex-biased genes, which were mostly MG, showed a strong trend toward localization in or around the cell membrane. We hypothesize that an increased expression of different ion channels in males, especially CFTR, leads to a better compensatory response for the mutated CFTR protein (see Table A3 and Table A4).
4.1. IL-17 Signaling Pathway
In their study using Luminex multiplex assays, Deny et al. [3] reported that a few inflammatory mediators were more highly expressed in the plasma of females (TNF-, IL-1b, IL-8, IL-10, IL-12p70, IL-17A, and CXCL10). They mainly appeared in the IL-17 pathway (TNF-, IL-1b, IL-17A, and CXCL10) involved in the response to inflammation and as a host defense. As described above, our results also included genes involved in the IL-17 pathway (MAPK and MMP9 (both MG) and CXCL5 (FG)). Here, MMP9 is responsible for tissue remodeling, hence upregulation in male patients may cause severe effects. Vermeer et al. [38] reported that higher expression of MMP9 in asthma patients was shown to decrease the functionality of cell barriers (specifically the tight junction pathway), which can limit the protection against bacteria entering the cells. However, MMP9 was shown to correlate adversely with lung function in CF patients [39] and has been discussed as a potential future therapeutic target in CF [40]. Female-biased expression of the chemokine CXCL5, also part of the IL-17 signaling pathway, could partly account for more intense inflammation reactions in females.
The dataset used in this study consists of whole blood transcriptomes, whereas cystic fibrosis is known to affect mostly epithelial cells. However, the IL-17 signaling pathway, as well as the pathways described in the following subsections, play significant roles in signaling cascades in both epithelial and whole blood cells. IL-17 can be produced by a wide range of immune cells present in the blood, e.g., Th17 cells, natural killer T cells, group 3 innate lymphoid cells, CD8+ cells, and neutrophils; [41], whereas IL-17 receptor A is widely expressed in epithelial cells, fibroblasts, and blood cells, e.g., macrophages, dendritic cells, and peripheral blood T lymphocytes [42]. The IL-17 downstream pathways induce the production of inflammatory molecules and chemokines, which leads to the recruitment of inflammatory cells. This process mediates the inflammatory response and contributes to the genesis of autoimmune disorders [41].
4.2. cAMP Signaling Pathway
The cAMP signaling pathway regulates homeostasis through direct or indirect modulation of transmembrane ion channels, including those for Ca2+, Na+, and K+, as well as CFTR via PKA activation. In our dataset, CFTR is a male-biased gene, suggesting a potential mechanism in males to compensate for the loss of function due to mutations. The beta-adrenoreceptor agonists are known as therapeutic targets in CF and act on the cAMP pathway via ADRB2 [43]. Figure A1 depicts the cAMP pathway (KEGG mapper). The genes mentioned in our study are marked in different colors. The targeted ADRB2 gene is marked in blue, yellow indicates the sex-biased genes detected in our study (all MG; see also Table 2), and pink indicates differentially expressed genes between CF and non-CF patients reported by other studies. The CFTR gene is marked in yellow/red and is located in the lower part of the figure (inside the cell membrane).
Adenylate cyclase (AC) produces cAMP, which affects ciliary beat frequency (lower in CF). AC function is impaired in CF but is more highly expressed in males, which may partly compensate for the lower functionality in CF [44]. The importance of the cAMP signaling pathway in CF is evident, as the inhibition of cAMP degradation via different phosphodiesterase inhibitors has been discussed and investigated as a therapeutic target (summarized in [45]). Sun et al. [46] reported that SNPs in ATP2B2 showed an association with the development of meconium ileus in CF patients.
Notably, all sex-biased expressed genes involved in the cAMP signaling pathway were found to be MG and mostly encode proteins located within or at the cell membrane, directly influencing cell homeostasis via ion channel regulation. Some of them are part of tight junctions or play a role in their regulation, with the latter being important in maintaining the function of epithelial tissue of the lungs, among others. However, we also observed a direct impact of cAMP signaling on whole blood cells. Chronic inflammation is characterized by the excessive migration of leukocytes from the peripheral blood into the tissues. The leukocyte extravasation process is regulated by signaling pathways in both leukocytes and vascular endothelium, which involve cAMP and calcium as intracellular messengers [47].
4.3. Estrogen Pathway
When comparing the sex-biased genes identified in this study to genes involved in the estrogen pathway, an overlap was observed only for MG (see also Figure A2). However, genes can be considered protective if expressed at higher levels. As reported by Kim et al. [48], non-mutated CFTR suppresses airway epithelial IL-8 production that occurs via a stimulatory EGFR cascade. A loss of normal CFTR activity exaggerates IL-8 production via the activation of a pro-inflammatory EGFR cascade. Both CFTR and EGFR are MG, possibly resulting in lower IL-8 production and less aggressive inflammation in males. This indicates a plausible protective characteristic and further hints at the stimulation of ciliary beat frequency, which is otherwise impaired by low adenylate cyclase levels.
Changes in the expression of genes involved in the estrogen pathway can also directly influence blood cells. Neutrophils, monocytes, macrophages, and lymphocytes, among others, express estrogen receptors [49], suggesting that estrogen directly affects the function of these cells. Estrogen has been found to suppress the development of B cells, but on the other hand, it has also been found to augment B cell functions, eventually leading to higher levels of antibodies observed in females. The number of circulating neutrophils is also influenced by the different levels of estrogen present during the different phases of the menstruation cycle in females [50]. Interestingly, estrogen treatment was shown to increase Th17 cells in the early phase of collagen-induced arthritis [51]. As IL-17 is essential for Th17 cells, this indicates a tight connection between estrogen and the IL-17 pathways.
4.4. Calcium Signaling Pathway
As shown in Table 2, a large number of MG genes were found to be involved in the calcium signaling pathway, many of which were located within the membrane of the cell. Here, EGFR, which is also involved in the estrogen pathway, and ADRB2, which is also involved in the cAMP pathway, were found to play a role in the calcium signaling pathway, indicating their involvement in multiple pathways. Moreover, calcium signals are also crucial for blood cells, such as those investigated in this study. They mediate the production of cytokines and the reprogramming of T cells, leading to their differentiation into various T cell subsets [52].
4.5. Further Targets
A known and reported target for future CF therapeutics [53,54] is the gene ANO1, which encodes for TMEM16A, an anion-selective channel activated by the binding of Ca2+ from the cytoplasm. ANO1 (logFC 1.9) was identified as MG in our data; therefore, targeting ANO1 may lead to different outcomes for male and female patients.
4.6. Outlook
Although Basu et al. [55] reported that approximately 75% of differentially expressed genes from whole blood analysis can be transferred to lung tissue, the field would benefit from a dedicated follow-up study that analyses sex-biased genes in bronchial epithelium. Halloran et al. showed a strong correlation (Spearman’s rank correlation coefficient of ) between mean expression levels in lung tissue and whole blood samples [56]. According to a study by Rotunno et al., the gene expression signature derived from blood samples reflects cancer-related gene expression changes in lung tissue, particularly in stage I lung adenocarcinoma [57]. A similar study was conducted to differentiate groups of patients with idiopathic pulmonary fibrosis, where a 13-gene cluster expressed in blood was used as a classifier to separate the groups. Yang et al. [58] presented a similar method to distinguish individuals with idiopathic pulmonary fibrosis from others.
Our analysis revealed that alternative splicing was a mechanism regulated by many of the sex-biased differentially expressed genes. Although our investigation only detected a small number of these sex-biased alternatively spliced genes, we conclude that further investigation into this topic is worthwhile. Datasets with higher sequencing depth would provide more statistical power and should be combined with dedicated analysis workflows to analyze this mechanism and its role in the regulation of sex-biased gene expression in more detail.
5. Conclusions
Our results show that some important pathways in CF pathology are MG expressed (IL-17 signaling pathway, cAMP signaling pathway, estrogen pathway, and calcium signaling pathway) and thus can account for the gender gap in mortality and morbidity in CF. The low number of sex-specific patient samples that limits our results emphasizes the importance of sex-specific studies in CF and other diseases to develop therapeutics that take into account patients’ sex and the corresponding biases.
Author Contributions
Conceptualization, C.G. and S.J.B.; methodology, C.G., S.J.B. and J.F.; validation, C.G., S.J.B. and J.F.; formal analysis, C.G. and S.J.B.; resources, T.K. and P.F.S.; data curation, C.G.; writing—original draft preparation, C.G., S.J.B. and J.F.; writing—review and editing, T.K., P.F.S., C.G., S.J.B. and J.F.; visualization, C.G. and S.J.B.; supervision, T.K., P.F.S. and J.F.; funding acquisition, S.J.B. and P.F.S. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by the European Union’s Horizon 2020 research and innovation program under the Marie Skłodowska-Curie grant (agreement no. 101029676) and the Open Access Publishing Fund of Leipzig University, which is supported by the German Research Foundation within the program Open Access Publication Funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Stephanie Kehr, Stephan Bernhardt, Rima Chakaroun, and the Ph.D. students of the Bioinformatics Group, Leipzig, for their fruitful discussions on gender-sensitive medicine.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| CF | cystic fibrosis |
| FG | female-biased genes |
| MG | male-biased genes |
| hsa | homo sapiens |
| CFTR | cystic fibrosis transmembrane conductance regulator |
| SABV | sex as biological variable |
| ESR | estrogen receptor |
| GPER | G protein-coupled estrogen receptor |
Appendix A. Additional Figure A1
Figure A1.
Sex-biased expressed genes in cystic fibrosis and their involvement in the cAMP signaling pathway (from KEGG Mapper). Pink indicates differentially expressed genes in CF vs. non-CF [3], yellow indicates sex-related differentially expressed genes in CF patients detected here (all MG, see also Table 2), and blue indicates potential targets for therapeutics (-2 adrenergic receptor agonists (bronchodilatation): ADRB2), as described in the main text. The CFTR gene (yellow/red) is the gene whose mutation causes CF. It appears as a male-biased gene but is also used as a target in therapeutics to restore activity. The double line represents the cell membrane. Genes marked in yellow: HCN4, GRIN2A, GRIN2B, GRIA1, GRIA2, ATP2B2, PDE10A. Genes marked in blue: ADRB2. Genes marked in pink: GHRL, PIK3R1, NFKBIA, NFKB1, SOX9. Genes marked in yellow/red: CFTR.
Figure A1.
Sex-biased expressed genes in cystic fibrosis and their involvement in the cAMP signaling pathway (from KEGG Mapper). Pink indicates differentially expressed genes in CF vs. non-CF [3], yellow indicates sex-related differentially expressed genes in CF patients detected here (all MG, see also Table 2), and blue indicates potential targets for therapeutics (-2 adrenergic receptor agonists (bronchodilatation): ADRB2), as described in the main text. The CFTR gene (yellow/red) is the gene whose mutation causes CF. It appears as a male-biased gene but is also used as a target in therapeutics to restore activity. The double line represents the cell membrane. Genes marked in yellow: HCN4, GRIN2A, GRIN2B, GRIA1, GRIA2, ATP2B2, PDE10A. Genes marked in blue: ADRB2. Genes marked in pink: GHRL, PIK3R1, NFKBIA, NFKB1, SOX9. Genes marked in yellow/red: CFTR.

Appendix B. Additional Figure A2
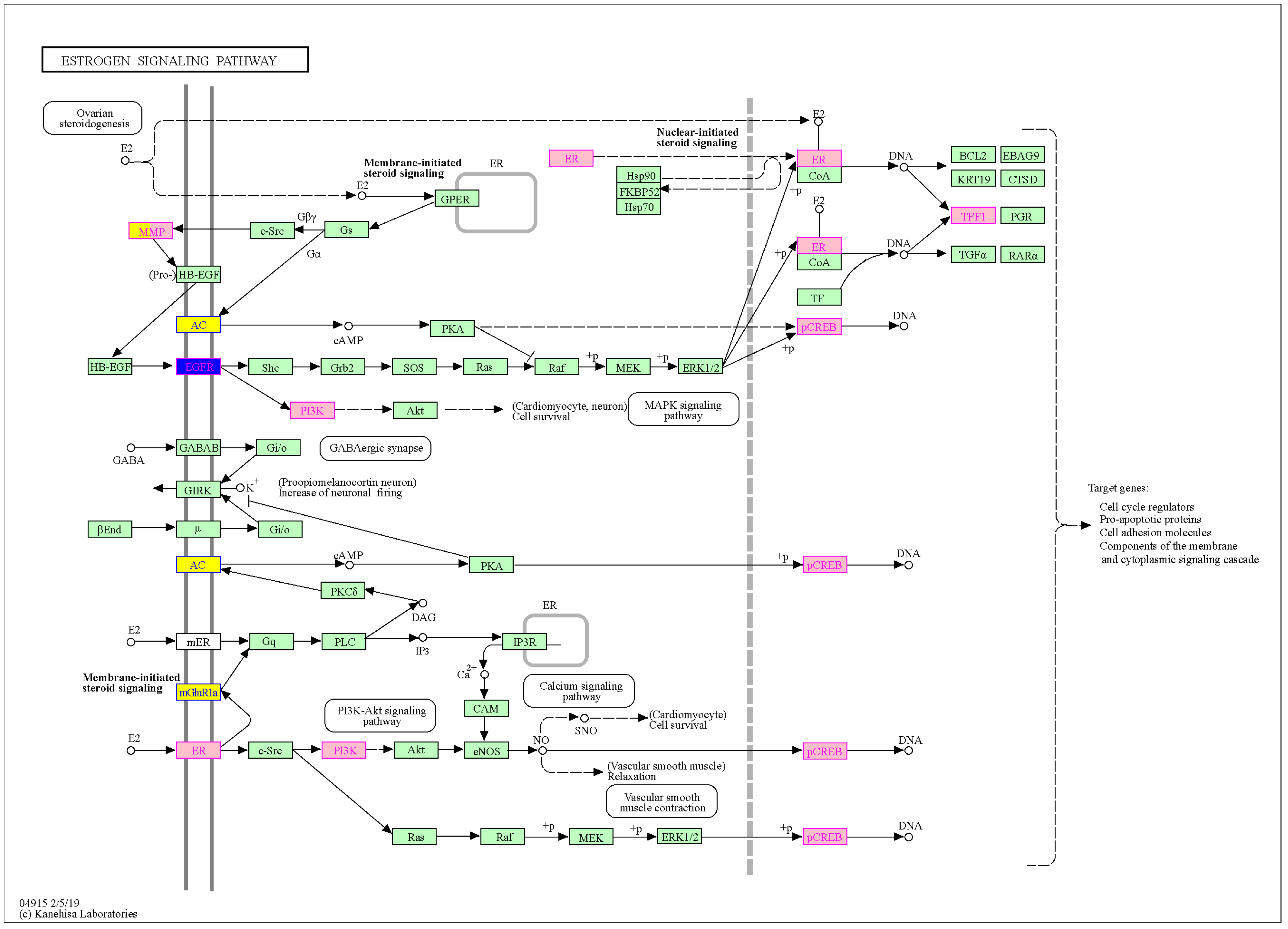
Figure A2.
Sex-biased expressed genes in cystic fibrosis and their involvement in the estrogen signaling pathway (from KEGG Mapper [31]). Estrogen acts either via estrogen receptors 1 and 2 (mER, ER) or via the G protein-coupled estrogen receptor (GPER). Yellow indicates differentially expressed genes regarding CF patient sex, which are all MG (see also Table 2). Pink indicates differentially expressed genes in CF vs. non-CF conditions [3]. Blue indicates EGFR, which is a potential target for therapeutics and is also regarded as a male-biased gene in our data (see also Table 2 and the main text for more details). The gene marked in yellow/pink (MMP9) is a male-biased gene, more highly expressed in CF vs. non-CF, and occurs in the IL-17 signaling pathway. Genes marked in pink: ESR1, TFF1, PIK3R1, ATF2, ATF4, ATF6B. Genes marked in yellow/pink: MMP9. Genes marked in blue: EGFR. Genes marked in yellow: ADCY2, ADCY8, GRM1, (EGFR).
Figure A2.
Sex-biased expressed genes in cystic fibrosis and their involvement in the estrogen signaling pathway (from KEGG Mapper [31]). Estrogen acts either via estrogen receptors 1 and 2 (mER, ER) or via the G protein-coupled estrogen receptor (GPER). Yellow indicates differentially expressed genes regarding CF patient sex, which are all MG (see also Table 2). Pink indicates differentially expressed genes in CF vs. non-CF conditions [3]. Blue indicates EGFR, which is a potential target for therapeutics and is also regarded as a male-biased gene in our data (see also Table 2 and the main text for more details). The gene marked in yellow/pink (MMP9) is a male-biased gene, more highly expressed in CF vs. non-CF, and occurs in the IL-17 signaling pathway. Genes marked in pink: ESR1, TFF1, PIK3R1, ATF2, ATF4, ATF6B. Genes marked in yellow/pink: MMP9. Genes marked in blue: EGFR. Genes marked in yellow: ADCY2, ADCY8, GRM1, (EGFR).

Appendix C. Overlap with StringDB

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Male-biased genes (MG) in CF found in StringDB associated with various cellular functions.
Table A1.
Male-biased genes (MG) in CF found in StringDB associated with various cellular functions.
| Gene Name | Term Description | Term ID | Gene Name | Term Description | Term ID |
|---|---|---|---|---|---|
| NALCN | Voltage-gated ion channel activity | GO:0005244 | ANO3 | Transporter activity | GO:0005215 |
| ABCC9 | Signaling receptor activity | GO:0038023 | GABRA4 | Signaling receptor activity | GO:0038023 |
| GABRA6 | Signaling receptor activity | GO:0038023 | KCNE4 | Voltage-gated ion channel activity | GO:0005244 |
| KCNT2 | Voltage-gated ion channel activity | GO:0005244 | TMC1 | Voltage-gated ion channel activity | GO:0005244 |
| CACNG2 | Voltage-gated ion channel activity | GO:0005244 | KCNH5 | Voltage-gated ion channel activity | GO:0005244 |
| SLC24A2 | Calcium channel activity | GO:0005262 | SCNN1B | Ligand-gated ion channel activity | GO:0015276 |
| CACNA2D1 | Voltage-gated ion channel activity | GO:0005244 | TRPC4 | Calcium channel activity | GO:0005262 |
| KCNU1 | Voltage-gated ion channel activity | GO:0005244 | SCN10A | Voltage-gated ion channel activity | GO:0005244 |
| PIEZO2 | Transporter activity | GO:0005215 | TRPC7 | Calcium channel activity | GO:0005262 |
| CACNG5 | Voltage-gated ion channel activity | GO:0005244 | TMEM120A | Transporter activity | GO:0005215 |
| GABRG3 | Signaling receptor activity | GO:0038023 | SLC10A2 | Transporter activity | GO:0005215 |
| SLC6A15 | Transporter activity | GO:0005215 | SLC28A1 | Transporter activity | GO:0005215 |
| TUSC3 | Transporter activity | GO:0005215 | SLC6A5 | Transporter activity | GO:0005215 |
| SLCO6A1 | Transporter activity | GO:0005215 | GRM5 | Neurotransmitter receptor activity | GO:0099583 |
| GRM1 | Neurotransmitter receptor activity | GO:0099583 | RANBP17 | Transporter activity | GO:0005215 |
| UNC13C | Calcium ion binding | GO:0005509 | GUCA1C | Calcium ion binding | GO:0005509 |
| CASQ2 | Calcium ion binding | GO:0005509 | CDH19 | Calcium ion binding | GO:0005509 |
| LRP2 | Calcium ion binding | GO:0005509 | CDH10 | Calcium ion binding | GO:0005509 |
| FSTL5 | Calcium ion binding | GO:0005509 | SLIT3 | Calcium ion binding | GO:0005509 |
| EML1 | Calcium ion binding | GO:0005509 | PCLO | Calcium ion binding | GO:0005509 |
| DNER | Signaling receptor activity | GO:0038023 | SMOC2 | Calcium ion binding | GO:0005509 |
| EGFLAM | Calcium ion binding | GO:0005509 | SPOCK3 | Calcium ion binding | GO:0005509 |
| PCDH15 | Calcium ion binding | GO:0005509 | TGM3 | Calcium ion binding | GO:0005509 |
| LRP1B | Calcium ion binding | GO:0005509 | CALN1 | Calcium ion binding | GO:0005509 |
| SVEP1 | Calcium ion binding | GO:0005509 | DGKB | Calcium ion binding | GO:0005509 |
| CLSTN2 | Calcium ion binding | GO:0005509 | CCBE1 | Calcium ion binding | GO:0005509 |
| SLIT2 | Calcium ion binding | GO:0005509 | CDH18 | Calcium ion binding | GO:0005509 |
| TENM2 | Calcium ion binding | GO:0005509 | FAT3 | Calcium ion binding | GO:0005509 |
| PCDH7 | Calcium ion binding | GO:0005509 | CDH8 | Calcium ion binding | GO:0005509 |
| PLA2G4C | Calcium ion binding | GO:0005509 | HMCN2 | Calcium ion binding | GO:0005509 |
| NOD2 | Signaling receptor activity | GO:0038023 | PKHD1 | Signaling receptor activity | GO:0038023 |
| PAQR5 | Signaling receptor activity | GO:0038023 | HNF4G | Signaling receptor activity | GO:0038023 |
| ADAMTS19 | Metalloendopeptidase activity | GO:0004222 | ADAMTS18 | Metalloendopeptidase activity | GO:0004222 |
| MMP16 | Metalloendopeptidase activity | GO:0004222 | ADAMTS3 | Metalloendopeptidase activity | GO:0004222 |
| MMP9 | Metalloendopeptidase activity | GO:0004222 | ADAM32 | Metalloendopeptidase activity | GO:0004222 |
| ADAMTS7 | Metalloendopeptidase activity | GO:0004222 | ADAMTS20 | Metalloendopeptidase activity | GO:0004222 |
| ADAMTS12 | Metalloendopeptidase activity | GO:0004222 | SHISA6 | PDZ domain binding | GO:0030165 |
Table A2.
Female-biased genes (FG) in CF found in StringDB associated with various cellular functions.
Table A2.
Female-biased genes (FG) in CF found in StringDB associated with various cellular functions.
| Gene Name | Term Description | Term ID | Gene Name | Term Description | Term ID |
|---|---|---|---|---|---|
| MCOLN2 | Ligand-gated ion channel activity | GO:0015276 | CD69 | Signaling receptor activity | GO:0038023 |
| DSC1 | Calcium ion binding | GO:0005509 | TNNT3 | Calcium ion binding | GO:0005509 |
| KRT1 | Signaling receptor activity | GO:0038023 | DLG3 | PDZ domain binding | GO:0030165 |
Table A3.
Male-biased genes (MG) in CF found in StringDB associated with different cellular compartments.
Table A3.
Male-biased genes (MG) in CF found in StringDB associated with different cellular compartments.
| Gene Name | Term Description | Term ID | Gene Name | Term Description | Term ID |
|---|---|---|---|---|---|
| NTRK2 | Intrinsic component of membrane | GOCC:0031224 | LHFPL4 | Membrane | GOCC:0016020 |
| GRIA2 | Intrinsic component of membrane | GOCC:0031224 | PCLO | Cytoskeleton of presynaptic active zone | GOCC:0048788 |
| ERBB4 | Plasma membrane protein complex | GOCC:0098797 | NGF | Somatodendritic compartment | GOCC:0036477 |
| RAB3B | Intrinsic component of membrane | GOCC:0031224 | PCDH15 | Plasma membrane-bounded cell projection | GOCC:0120025 |
| SYNDIG1 | Membrane | GOCC:0016020 | ELAVL2 | Presynapse | GOCC:0098793 |
| NLGN1 | Glutamatergic synapse | GOCC:0098978 | SNAP91 | Presynaptic membrane | GOCC:0042734 |
| APBB2 | Membrane | GOCC:0016020 | GRIA1 | Glutamatergic synapse | GOCC:0098978 |
| SLC6A5 | Membrane | GOCC:0016020 | PPFIA2 | Dendritic spine | GOCC:0043197 |
| SHANK2 | Dendritic spine | GOCC:0043197 | CNTNAP4 | Presynaptic membrane | GOCC:0042734 |
| GRHL2 | Cell junction | GOCC:0030054 | EGFR | Intrinsic component of membrane | GOCC:0031224 |
| DLC1 | Membrane | GOCC:0016020 | SHROOM3 | Cell junction | GOCC:0030054 |
| NRAP | Cell junction | GOCC:0030054 | FRMPD2 | Membrane | GOCC:0016020 |
| MPP4 | Cell junction | GOCC:0030054 | CTNNA3 | Cell junction | GOCC:0030054 |
| CFTR | Intrinsic component of membrane | GOCC:0031224 | GPRC5A | Membrane | GOCC:0016020 |
| GNGT1 | Plasma membrane protein complex | GOCC:0098797 | NALCN | Membrane | GOCC:0016020 |
| FOLH1 | Intrinsic component of membrane | GOCC:0031224 | MEOX2 | Cell periphery | GOCC:0071944 |
| CDH19 | Membrane | GOCC:0016020 | LRP2 | Membrane | GOCC:0016020 |
| SLC6A15 | Intrinsic component of membrane | GOCC:0031224 | MMP16 | Intrinsic component of membrane | GOCC:0031224 |
| KCNT2 | Membrane | GOCC:0016020 | CSMD3 | Membrane | GOCC:0016020 |
| USH2A | Membrane | GOCC:0016020 | ADRA1B | Membrane | GOCC:0016020 |
| SLCO2A1 | Membrane | GOCC:0016020 | OR4N2 | Membrane | GOCC:0016020 |
| KCNH5 | Membrane | GOCC:0016020 | KRT7 | Cell periphery | GOCC:0071944 |
| EPHA3 | Intrinsic component of membrane | GOCC:0031224 | GFRAL | Membrane | GOCC:0016020 |
| SLC24A2 | Membrane | GOCC:0016020 | SCNN1B | Intrinsic component of membrane | GOCC:0031224 |
| SMOC2 | Cell periphery | GOCC:0071944 | ANO1 | Intrinsic component of membrane | GOCC:0031224 |
| GFRA1 | Membrane | GOCC:0016020 | TAAR2 | Membrane | GOCC:0016020 |
| SLC16A12 | Intrinsic component of membrane | GOCC:0031224 | NRG3 | Intrinsic component of membrane | GOCC:0031224 |
| PTPRT | Intrinsic component of membrane | GOCC:0031224 | ITGBL1 | Membrane | GOCC:0016020 |
| GPR158 | Membrane | GOCC:0016020 | TRPM3 | Membrane | GOCC:0016020 |
| TGM3 | Membrane | GOCC:0016020 | NLRP5 | Cell periphery | GOCC:0071944 |
| DPP10 | Membrane | GOCC:0016020 | PTPRZ1 | Intrinsic component of membrane | GOCC:0031224 |
| COL25A1 | Intrinsic component of membrane | GOCC:0031224 | MDGA2 | Membrane | GOCC:0016020 |
| KCNU1 | Membrane | GOCC:0016020 | LRRC32 | Intrinsic component of membrane | GOCC:0031224 |
| UNC80 | Membrane | GOCC:0016020 | BMPR1B | Intrinsic component of membrane | GOCC:0031224 |
| RXFP1 | Membrane | GOCC:0016020 | LPAR4 | Membrane | GOCC:0016020 |
| PEX5L | Intrinsic component of membrane | GOCC:0031224 | SLCO6A1 | Membrane | GOCC:0016020 |
| TUSC3 | Intrinsic component of membrane | GOCC:0031224 | CDH18 | Membrane | GOCC:0016020 |
| TRPC7 | Membrane | GOCC:0016020 | SNTG1 | Plasma membrane protein complex | GOCC:0098797 |
| PCDH7 | Membrane | GOCC:0016020 | GPR17 | Membrane | GOCC:0016020 |
| CNTN1 | Membrane | GOCC:0016020 | NRXN3 | Intrinsic component of membrane | GOCC:0031224 |
| ABCA8 | Membrane | GOCC:0016020 | HCRTR2 | Intrinsic component of membrane | GOCC:0031224 |
| FGF1 | Cell periphery | GOCC:0071944 | MLPH | Somatodendritic compartment | GOCC:0036477 |
| TMC1 | Plasma membrane-bounded cell projection | GOCC:0120025 | OLFM1 | Somatodendritic compartment | GOCC:0036477 |
| GRXCR2 | Plasma membrane-bounded cell projection | GOCC:0120025 | MYO3B | Plasma membrane-bounded cell projection | GOCC:0120025 |
| CASQ2 | Intrinsic component of membrane | GOCC:0031224 | ARNT2 | Receptor complex | GOCC:0043235 |
| LRP1B | Receptor complex | GOCC:0043235 | ZPBP | Membrane | GOCC:0016020 |
| ART1 | Membrane | GOCC:0016020 | SORCS1 | Membrane | GOCC:0016020 |
| TYR | Membrane | GOCC:0016020 | ABCA12 | Intrinsic component of membrane | GOCC:0031224 |
| STXBP5L | Membrane | GOCC:0016020 | LRRC4C | Intrinsic component of membrane | GOCC:0031224 |
| FUT9 | Membrane | GOCC:0016020 | FUT7 | Membrane | GOCC:0016020 |
| IGDCC3 | Membrane | GOCC:0016020 | ROS1 | Membrane | GOCC:0016020 |
| GPC5 | Intrinsic component of membrane | GOCC:0031224 | SEL1L2 | Membrane | GOCC:0016020 |
| CADPS | Membrane | GOCC:0016020 | TSPAN8 | Membrane | GOCC:0016020 |
| CALN1 | Membrane | GOCC:0016020 | SLIT2 | Membrane | GOCC:0016020 |
| REEP1 | Membrane | GOCC:0016020 | PIK3C2G | Membrane | GOCC:0016020 |
| PLA2G4C | Membrane | GOCC:0016020 |
Table A4.
Female-biased genes (FG) in CF found in StringDB associated with different cellular compartments.
Table A4.
Female-biased genes (FG) in CF found in StringDB associated with different cellular compartments.
| Gene Name | Term Description | Term ID | Gene Name | Term Description | Term ID |
|---|---|---|---|---|---|
| CD69 | Intrinsic component of membrane | GOCC:0031224 | TAS2R5 | Membrane | GOCC:0016020 |
| KRT1 | Membrane | GOCC:0016020 | GPR174 | Membrane | GOCC:0016020 |
| TRAT1 | Plasma membrane protein complex | GOCC:0098797 | ACVR2B | Intrinsic component of membrane | GOCC:0031224 |
| NCR3LG1 | Membrane | GOCC:0016020 | FPR3 | Membrane | GOCC:0016020 |
| MCOLN2 | Membrane | GOCC:0016020 | GPR183 | Membrane | GOCC:0016020 |
| CCDC136 | Membrane | GOCC:0016020 | KLRC4 | Membrane | GOCC:0016020 |
| ABCD2 | Membrane | GOCC:0016020 | ELOVL2 | Membrane | GOCC:0016020 |
| SPTSSB | Membrane | GOCC:0016020 | TXLNG | Membrane | GOCC:0016020 |
| KLRC3 | Membrane | GOCC:0016020 | SLFN12L | Membrane | GOCC:0016020 |
Table A5.
Male-biased genes (MG) in CF found in StringDB associated with alternative splicing (StringDB term KW-0025).
Table A5.
Male-biased genes (MG) in CF found in StringDB associated with alternative splicing (StringDB term KW-0025).
| Gene Name | Gene Name |
|---|---|
| GRHL2 | DOK5 |
| TYR | RAPGEFL1 |
| DLC1 | ARNT2 |
| RBFOX1 | SH3GL3 |
| MYO18B | ADCY2 |
| NRAP | KAZN |
| POU6F2 | DGKB |
| SHISA6 | DDC |
| CA10 | APBB2 |
| TRDN | PLA2G4C |
| POU2F3 | NOS1AP |
| ANO3 | SLC7A13 |
| UNC80 | CADPS2 |
| SLCO6A1 | CADPS |
| STXBP5L | ABCA8 |
| SNAP91 | DPP10 |
| MLPH | NKAIN1 |
| EPB41L4B | KSR2 |
| KIAA1549L | SYT14 |
| MS4A13 | REEP1 |
| NXPE2 | GALNT13 |
| MGAT4C | CGNL1 |
| ZBBX | SNX7 |
| NPAS3 | SNX31 |
| ELAVL2 | IQCJ |
| LIMCH1 | LRRIQ1 |
| LRRC49 | MIPOL1 |
| SERPINB11 | RGL3 |
| LRRC75B | LRRC31 |
| CDC20B | ANKRD62 |
Table A6.
Female-biased genes (FG) in CF found in StringDB associated with alternative splicing (StringDB term KW-0025).
Table A6.
Female-biased genes (FG) in CF found in StringDB associated with alternative splicing (StringDB term KW-0025).
| Gene Name | Gene Name |
|---|---|
| AIRE | SLFN12L |
| TXLNG | ZNF711 |
Table A7.
Male-biased (MG) alternatively spliced genes in CF patients.
| Ensembl ID | Gene Name |
|---|---|
| ENSG00000122783 | C7orf49 |
| ENSG00000073578 | SDHA |
| ENSG00000198034 | RPS4X |
| ENSG00000031698 | SARS |
| ENSG00000122783 | C7orf49 |
| ENSG00000175567 | UCP2 |
| ENSG00000115464 | USP34 |
| ENSG00000159140 | SON |
| ENSG00000134905 | CARS2 |
| ENSG00000115760 | BIRC6 |
| ENSG00000168028 | RPSA |
| ENSG00000236213 | AC006369.1 |
| ENSG00000162402 | USP24 |
| ENSG00000107290 | SETX |
| ENSG00000228655 | AC096558.1 |
| ENSG00000141469 | SLC14A1 |
Table A8.
Female-biased (FG) alternatively spliced genes in CF patients.
| Ensembl ID | Gene Name |
|---|---|
| ENSG00000058272 | PPP1R12A |
| ENSG00000107099 | DOCK8 |
| ENSG00000055609 | KMT2C |
| ENSG00000137642 | SORL1 |
| ENSG00000105851 | PIK3CG |
| ENSG00000124942 | AHNAK |
| ENSG00000197852 | FAM212B |
| ENSG00000145416 | MARCH1 |
| ENSG00000183486 | MX2 |
| ENSG00000114331 | ACAP2 |
| ENSG00000160255 | ITGB2 |
| ENSG00000139083 | ETV6 |
| ENSG00000164631 | ZNF12 |
| ENSG00000118058 | KMT2A |
| ENSG00000069667 | RORA |
| ENSG00000101596 | SMCHD1 |
| ENSG00000197329 | PELI1 |
| ENSG00000106714 | CNTNAP3 |
References
- Burgel, P.R.; Bellis, G.; Olesen, H.V.; Viviani, L.; Zolin, A.; Blasi, F.; Elborn, J.S. Future trends in cystic fibrosis demography in 34 European countries. Eur. Respir. J. 2015, 46, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Lam, G.Y.; Goodwin, J.; Wilcox, P.G.; Quon, B.S. Sex disparities in cystic fibrosis: Review on the effect of female sex hormones on lung pathophysiology and outcomes. ERJ Open Res. 2021, 7, 00475–2020. [Google Scholar] [CrossRef] [PubMed]
- Deny, M.; Popotas, A.; Hanssens, L.; Lefèvre, N.; Nuñez, L.A.A.; Ouafo, G.S.; Corazza, F.; Casimir, G.; Chamekh, M. Sex-biased expression of selected chromosome x-linked microRNAs with potent regulatory effect on the inflammatory response in children with cystic fibrosis: A preliminary pilot investigation. Front. Immunol. 2023, 14, 1114239. [Google Scholar] [CrossRef]
- Harness-Brumley, C.L.; Elliott, A.C.; Rosenbluth, D.B.; Raghavan, D.; Jain, R. Gender differences in outcomes of patients with cystic fibrosis. J. Women’s Health 2014, 23, 1012–1020. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.J.; Tildesley, H.D.; Wilcox, P.G.; Zhang, H.; Kreisman, S.H. Sex disparities in effects of cystic fibrosis-related diabetes on clinical outcomes: A matched study. Can. Respir. J. 2008, 15, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, A.L.; Tom, M.; Berthiaume, Y.; Singer, L.G.; Aaron, S.D.; Whitmore, G.; Stanojevic, S. A contemporary survival analysis of individuals with cystic fibrosis: A cohort study. Eur. Respir. J. 2015, 45, 670–679. [Google Scholar] [CrossRef]
- Cystic Fibrosis Canada. The Canadian Cystic Fibrosis Registry. 2017 Annual Data Report. Available online: www.cysticfibrosis.ca/blog/2017-registry-annual-data-report (accessed on 20 April 2023).
- Raghavan, D.; Jain, R. Increasing awareness of sex differences in airway diseases. Respirology 2016, 21, 449–459. [Google Scholar] [CrossRef] [PubMed]
- LoMauro, A.; Aliverti, A. Sex differences in respiratory function. Breathe 2018, 14, 131–140. [Google Scholar] [CrossRef]
- Viviani, L.; Bossi, A.; Assael, B.M.; Italian Registry for Cystic Fibrosis Collaborative Group. Absence of a gender gap in survival. An analysis of the Italian registry for cystic fibrosis in the paediatric age. J. Cyst. Fibros. 2011, 10, 313–317. [Google Scholar] [CrossRef]
- Sutton, S.; Rosenbluth, D.; Raghavan, D.; Zheng, J.; Jain, R. Effects of puberty on cystic fibrosis related pulmonary exacerbations in women versus men. Pediatr. Pulmonol. 2014, 49, 28–35. [Google Scholar] [CrossRef]
- Arterburn, J.B.; Prossnitz, E.R. G Protein–Coupled Estrogen Receptor GPER: Molecular Pharmacology and Therapeutic Applications. Annu. Rev. Pharmacol. Toxicol. 2023, 63, 295–320. [Google Scholar] [CrossRef]
- Clarke, L.A.; Sousa, L.; Barreto, C.; Amaral, M.D. Changes in transcriptome of native nasal epithelium expressing F508del-CFTR and intersecting data from comparable studies. Respir. Res. 2013, 14, 38. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cela, E.; Gagnon, S.; Sweezey, N.B. Estrogen aggravates inflammation in Pseudomonas aeruginosa pneumonia in cystic fibrosis mice. Respir. Res. 2010, 11, 166. [Google Scholar] [CrossRef] [PubMed]
- Tyrrell, J.; Harvey, B.J. Sexual dimorphism in the microbiology of the CF ‘Gender Gap’: Estrogen modulation of Pseudomonas aeruginosa virulence. Steroids 2020, 156, 108575. [Google Scholar] [CrossRef]
- Chotirmall, S.H.; Smith, S.G.; Gunaratnam, C.; Cosgrove, S.; Dimitrov, B.D.; O’Neill, S.J.; Harvey, B.J.; Greene, C.M.; McElvaney, N.G. Effect of estrogen on pseudomonas mucoidy and exacerbations in cystic fibrosis. N. Engl. J. Med. 2012, 366, 1978–1986. [Google Scholar] [CrossRef]
- Secunda, K.E.; Guimbellot, J.S.; Jovanovic, B.; Heltshe, S.L.; Sagel, S.D.; Rowe, S.M.; Jain, M. Females with cystic fibrosis demonstrate a differential response profile to ivacaftor compared with males. Am. J. Respir. Crit. Care Med. 2020, 201, 996–998. [Google Scholar] [CrossRef]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Prieto, M.D.; Jang, J.; Franciosi, A.N.; Av-Gay, Y.; Bach, H.; Tebbutt, S.J.; Quon, B.S. Whole blood RNA-seq demonstrates an increased host immune response in individuals with cystic fibrosis who develop nontuberculous mycobacterial pulmonary disease. PLoS ONE 2022, 17, e0278296. [Google Scholar] [CrossRef]
- Fallmann, J.; Goldmann, R.; Stadler, P.F. MONSDA: Modular Organizer of Nextflow and Snakemake Driven HTS Data Analysis. OSF Prepr. 2022, 8. [Google Scholar] [CrossRef]
- SRA Toolkit Development Team SRA Toolkit. Available online: https://github.com/ncbi/sra-tools (accessed on 26 May 2023).
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Babraham Bioinformatics—FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 26 May 2023).
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize Analysis Results for Multiple Tools and Samples in a Single Report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Guo, S.; Zhou, Y.; Zeng, P.; Xu, G.; Wang, G.; Cui, Q. Identification and analysis of the human sex-biased genes. Briefings Bioinform. 2018, 19, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2007, 36, D480–D484. [Google Scholar] [CrossRef]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO summarizes and visualizes long lists of gene ontology terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef]
- Ideozu, J.E.; Zhang, X.; McColley, S.; Levy, H. Transcriptome profiling and molecular therapeutic advances in cystic fibrosis: Recent insights. Genes 2019, 10, 180. [Google Scholar] [CrossRef]
- Talebizadeh, Z.; Simon, S.D.; Butler, M.G. X Chromosome Gene Expression in Human Tissues: Male and Female Comparisons. Genomics 2006, 88, 675–681. [Google Scholar] [CrossRef]
- Ogilvie, V.; Passmore, M.; Hyndman, L.; Jones, L.; Stevenson, B.; Wilson, A.; Davidson, H.; Kitchen, R.R.; Gray, R.D.; Shah, P.; et al. Differential Global Gene Expression in Cystic Fibrosis Nasal and Bronchial Epithelium. Genomics 2011, 98, 327–336. [Google Scholar] [CrossRef]
- Wright, J.M.; Merlo, C.A.; Reynolds, J.B.; Zeitlin, P.L.; Garcia, J.G.; Guggino, W.B.; Boyle, M.P. Respiratory epithelial gene expression in patients with mild and severe cystic fibrosis lung disease. Am. J. Respir. Cell Mol. Biol. 2006, 35, 327–336. [Google Scholar] [CrossRef]
- Trivedi, T.S.; Bhadresha, K.P.; Patel, M.P.; Mankad, A.U.; Rawal, R.M.; Patel, S.K. Identification of hub genes associated with human cystic fibrosis: A Meta-analysis approach. Hum. Gene 2023, 35, 201139. [Google Scholar] [CrossRef]
- Vermeer, P.D.; Denker, J.; Estin, M.; Moninger, T.O.; Keshavjee, S.; Karp, P.; Kline, J.N.; Zabner, J. MMP9 modulates tight junction integrity and cell viability in human airway epithelia. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2009, 296, L751–L762. [Google Scholar] [CrossRef] [PubMed]
- Devereux, G.; Steele, S.; Jagelman, T.; Fielding, S.; Muirhead, R.; Brady, J.; Grierson, C.; Brooker, R.; Winter, J.; Fardon, T.; et al. An observational study of matrix metalloproteinase (MMP)-9 in cystic fibrosis. J. Cyst. Fibros. 2014, 13, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Esposito, R.; Mirra, D.; Spaziano, G.; Panico, F.; Gallelli, L.; D’Agostino, B. The Role of MMPs in the Era of CFTR Modulators: An Additional Target for Cystic Fibrosis Patients? Biomolecules 2023, 13, 350. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Huang, M.; Yao, Y.m. Biology of interleukin-17 and its pathophysiological significance in sepsis. Front. Immunol. 2020, 11, 1558. [Google Scholar] [CrossRef]
- Eyerich, S.; Eyerich, K.; Cavani, A.; Schmidt-Weber, C. IL-17 and IL-22: Siblings, not twins. Trends Immunol. 2010, 31, 354–361. [Google Scholar] [CrossRef]
- Deutsche Gesellschaft für Pneumologie und Beatmungsmedizin e.V. (DGP); Gesellschaft für Pädiatrische Pneumologie e.V. (GPP). S3-Leitlinie Lungenerkrankung bei Mukoviszidose: Pseudomonas aeruginosa, V. 2.0. Available online: https://www.awmf.org/service/awmf-aktuell/lungenerkrankung-bei-mukoviszidose-pseudomonas-aeruginosa (accessed on 29 April 2023).
- Schmid, A.; Sutto, Z.; Schmid, N.; Novak, L.; Ivonnet, P.; Horvath, G.; Conner, G.; Fregien, N.; Salathe, M. Decreased Soluble Adenylyl Cyclase Activity in Cystic Fibrosis Is Related to Defective Apical Bicarbonate Exchange and Affects Ciliary Beat Frequency Regulation. J. Biol. Chem. 2010, 285, 29998–30007. [Google Scholar] [CrossRef]
- Turner, M.J.; Abbott-Banner, K.; Thomas, D.Y.; Hanrahan, J.W. Cyclic nucleotide phosphodiesterase inhibitors as therapeutic interventions for cystic fibrosis. Pharmacol. Ther. 2021, 224, 107826. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Rommens, J.M.; Corvol, H.; Li, W.; Li, X.; Chiang, T.A.; Lin, F.; Dorfman, R.; Busson, P.F.; Parekh, R.V.; et al. Multiple apical plasma membrane constituents are associated with susceptibility to meconium ileus in individuals with cystic fibrosis. Nat. Genet. 2012, 44, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Lorenowicz, M.J.; Fernandez-Borja, M.; Hordijk, P.L. cAMP signaling in leukocyte transendothelial migration. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Beyer, B.A.; Lewis, C.; Nadel, J.A. Normal CFTR inhibits epidermal growth factor receptor-dependent pro-inflammatory chemokine production in human airway epithelial cells. PLoS ONE 2013, 8, e72981. [Google Scholar] [CrossRef]
- Pierdominici, M.; Maselli, A.; Colasanti, T.; Giammarioli, A.M.; Delunardo, F.; Vacirca, D.; Sanchez, M.; Giovannetti, A.; Malorni, W.; Ortona, E. Estrogen receptor profiles in human peripheral blood lymphocytes. Immunol. Lett. 2010, 132, 79–85. [Google Scholar] [CrossRef]
- Nowak, K.; Jabłońska, E.; Ratajczak-Wrona, W. Neutrophils life under estrogenic and xenoestrogenic control. J. Steroid Biochem. Mol. Biol. 2019, 186, 203–211. [Google Scholar] [CrossRef]
- Kumar, R.S.; Goyal, N. Estrogens as regulator of hematopoietic stem cell, immune cells and bone biology. Life Sci. 2021, 269, 119091. [Google Scholar] [CrossRef]
- Vaeth, M.; Kahlfuss, S.; Feske, S. CRAC channels and calcium signaling in T cell-mediated immunity. Trends Immunol. 2020, 41, 878–901. [Google Scholar] [CrossRef]
- Quesada, R.; Dutzler, R. Alternative chloride transport pathways as pharmacological targets for the treatment of cystic fibrosis. J. Cyst. Fibros. 2020, 19, S37–S41. [Google Scholar] [CrossRef]
- Galietta, L.J. TMEM16A (ANO1) as a therapeutic target in cystic fibrosis. Curr. Opin. Pharmacol. 2022, 64, 102206. [Google Scholar] [CrossRef]
- Basu, M.; Wang, K.; Ruppin, E.; Hannenhalli, S. Predicting tissue-specific gene expression from whole blood transcriptome. Sci. Adv. 2021, 7, eabd6991. [Google Scholar] [CrossRef] [PubMed]
- Halloran, J.W.; Zhu, D.; Qian, D.C.; Byun, J.; Gorlova, O.Y.; Amos, C.I.; Gorlov, I.P. Prediction of the gene expression in normal lung tissue by the gene expression in blood. BMC Med. Genom. 2015, 8, 77. [Google Scholar] [CrossRef] [PubMed]
- Rotunno, M.; Hu, N.; Su, H.; Wang, C.; Goldstein, A.M.; Bergen, A.W.; Consonni, D.; Pesatori, A.C.; Bertazzi, P.A.; Wacholder, S.; et al. A Gene Expression Signature from Peripheral Whole Blood for Stage I Lung AdenocarcinomaGene Expression in Blood and Tissue in Stage I Lung Cancer. Cancer Prev. Res. 2011, 4, 1599–1608. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.V.; Luna, L.G.; Cotter, J.; Talbert, J.; Leach, S.M.; Kidd, R.; Turner, J.; Kummer, N.; Kervitsky, D.; Brown, K.K.; et al. The peripheral blood transcriptome identifies the presence and extent of disease in idiopathic pulmonary fibrosis. PLoS ONE 2012, 7, e37708. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chromosomal distribution of male-biased and female-biased upregulated genes in cystic fibrosis patients.
Figure 1.
Chromosomal distribution of male-biased and female-biased upregulated genes in cystic fibrosis patients.

Figure 2.
Sex-biased expressed genes in cystic fibrosis and their distribution across cellular compartments (selection, multiple assignments per gene possible). For detailed gene lists, see Table A3 and Table A4.

Table 1.
Baseline characteristics of males and females in the dataset used. Results are presented as medians (interquartile range). There were no significant differences observed in the baseline characteristics using Wilcoxon’s rank sum test (). FEV1: forced expiratory volume in one second; FVC: forced vital capacity.
Table 1.
Baseline characteristics of males and females in the dataset used. Results are presented as medians (interquartile range). There were no significant differences observed in the baseline characteristics using Wilcoxon’s rank sum test (). FEV1: forced expiratory volume in one second; FVC: forced vital capacity.
| Females | Males | |
|---|---|---|
| Patients (n) | 5 | 5 |
| Age (years) | 32 (30; 38) | 29 (26; 33) |
| FEV1 (% predicted) | 89.1 (80.6; 94.6) | 63.7 (45.6; 83.0) |
| FEV1/FVC | 0.79 (0.78; 0.8) | 0.65 (0.56; 0.76) |
Table 2.
Genes and corresponding KEGG pathways exhibiting sex-biased expression in CF.
| Pathway/Function | MG | FG |
|---|---|---|
| hsa04657 IL-17 signaling pathway | MAPK4, MMP9 | CXCL5 |
| hsa04915 Estrogen signaling pathway | MMP9, ADCY2, ADCY8, EGFR, GRM1 | – |
| hsa04152 AMPK signaling pathway | PPP2R2C, CFTR | – |
| hsa04024 cAMP signaling pathway | FSHR, ADCY2, ADCY8, HCN4, GRIN2A, GRIN2B, GRIA1, GRIA2, CFTR, ATP2B2, PDE10A | – |
| hsa04020 Calcium signaling pathway | ATP2B2, ADCY2, ADCY8, GRIN2A, TRDN, CASQ2, ADRA1B, GRM1, GRM5, FGF1, NGF, EGFR, ERBB4, NTRK2, PDE1A, PDE1C | MCOLN2 (TRPML) |
| hsa04530 Tight junction | CLDN14, MPP4, PPP2R2C, CGNL1, CFTR, EPB41L4B, MAGI1 | CLDN20, DLG3 |
| hsa5173214 O-glycosylation of TSR domain-containing proteins | ADAMTS18/19/20, ADAMTS3, ADAMTSL1, ADAMTS7, ADAMTS12 | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gärtner, C.; Fallmann, J.; Stadler, P.F.; Kaiser, T.; Berkemer, S.J. Toward a Systematic Assessment of Sex Differences in Cystic Fibrosis. J. Pers. Med. 2023, 13, 924. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm13060924
AMA Style
Gärtner C, Fallmann J, Stadler PF, Kaiser T, Berkemer SJ. Toward a Systematic Assessment of Sex Differences in Cystic Fibrosis. Journal of Personalized Medicine. 2023; 13(6):924. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm13060924
Chicago/Turabian StyleGärtner, Christiane, Jörg Fallmann, Peter F. Stadler, Thorsten Kaiser, and Sarah J. Berkemer. 2023. "Toward a Systematic Assessment of Sex Differences in Cystic Fibrosis" Journal of Personalized Medicine 13, no. 6: 924. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm13060924
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.