Potential Impact of MicroRNA Gene Polymorphisms in the Pathogenesis of Diabetes and Atherosclerotic Cardiovascular Disease


 ,
,
Abstract
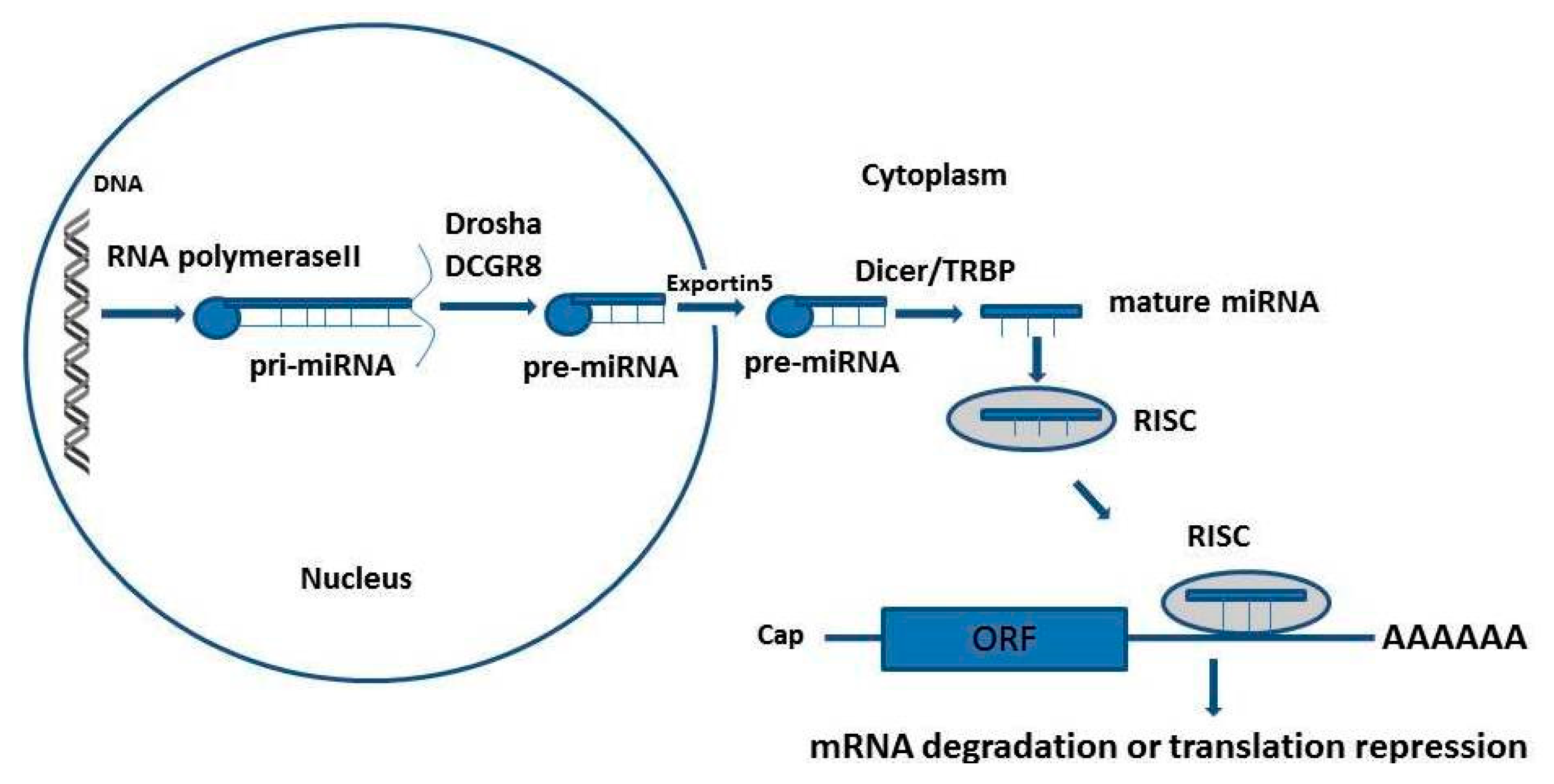
:1. Introduction
2. Micro-RNA Gene Polymorphisms in Diabetes and Diabetes Complications
2.1. MiR-124a rs531564 G>C
2.2. MiR-375
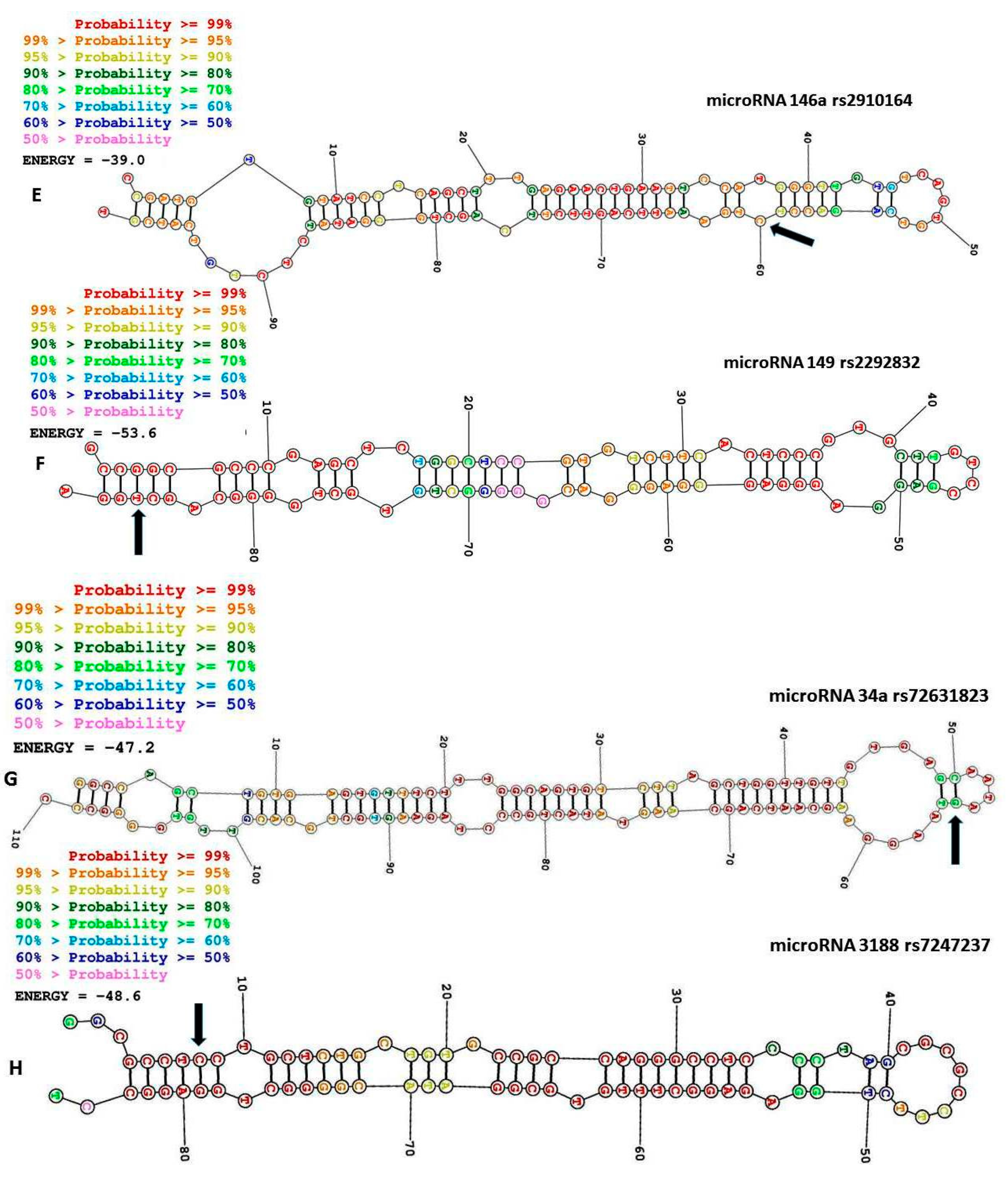
2.3. MiR-146a rs2910164 C>G
2.4. MiR-27a rs895819
2.5. MiR-34a rs72631823 G>A
2.6. MicroRNA Let-7a-2 (MIRLET-7A2 rs1143770)
2.7. MiR-155 rs767649 T>A
2.8. MiR128a rs11888095 C>T
2.9. MiR-3188 rs7247237 C>T
2.10. MiR-126 rs4636297 A>G
3. Micro-RNA Gene Polymorphisms in Atherosclerotic Cardiovascular Disease
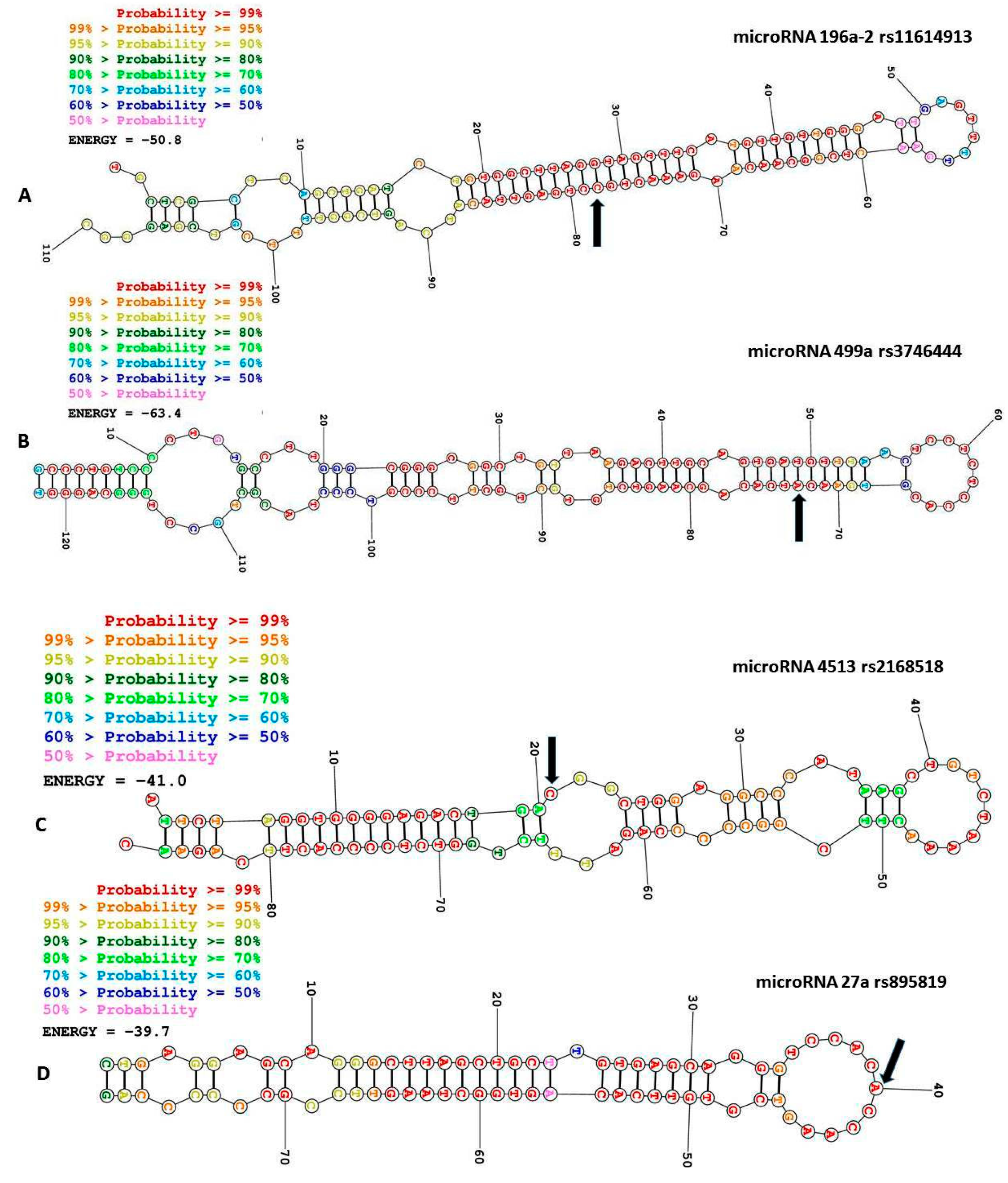
3.1. The miR-196a2 rs11614913 T>C
3.2. MiR-499 rs3746444 A>G
3.3. MiR-4513 rs2168518 C>T
3.4. The Pre-MiR-27a rs895819 A>T
3.5. MiR-146a rs2910164 G>C
3.6. MiR-149 rs2292832 T>C
3.7. MIRLET7 Family of microRNA
4. Conclusions and Future Perspective
Author Contributions
Funding
Conflicts of Interest
References
- Starega-Roslan, J.; Krol, J.; Koscianska, E.; Kozlowski, P.; Szlachcic, W.J.; Sobczak, K.; Krzyzosiak, W.J. Structural basis of microrna length variety. Nucleic Acids Res. 2011, 39, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by micrornas: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.M.; Tian, Y.; Tu, M.J.; Ho, P.Y.; Jilek, J.L. Microrna pharmacoepigenetics: Posttranscriptional regulation mechanisms behind variable drug disposition and strategy to develop more effective therapy. Drug Metab. Dispos. Biol. Fate Chem. 2016, 44, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Saunders, M.A.; Liang, H.; Li, W.H. Human polymorphism at micrornas and microrna target sites. Proc. Natl. Acad. Sci. USA 2007, 104, 3300–3305. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.Y.; Cheng, J.; Zhou, M.Y.; Liang, L.L.; Lian, S.M.; Xie, X.S.; Xu, S.; Liu, X.; Xiong, X.D. The association between pre-mir-27a rs895819 polymorphism and myocardial infarction risk in a chinese han population. Lipids Health Dis. 2018, 17, 7. [Google Scholar] [CrossRef]
- Ladomery, M.R.; Maddocks, D.G.; Wilson, I.D. Micrornas: Their discovery, biogenesis, function and potential use as biomarkers in non-invasive prenatal diagnostics. Int. J. Mol. Epidemiol. Genet. 2011, 2, 253–260. [Google Scholar]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. Microrna genes are transcribed by rna polymerase ii. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef]
- Graves, P.; Zeng, Y. Biogenesis of mammalian micrornas: A global view. Genom. Proteom. Bioinform. 2012, 10, 239–245. [Google Scholar] [CrossRef]
- Iwakawa, H.O.; Tomari, Y. The functions of micrornas: Mrna decay and translational repression. Trends Cell Biol. 2015, 25, 651–665. [Google Scholar] [CrossRef]
- Romaine, S.P.; Tomaszewski, M.; Condorelli, G.; Samani, N.J. Micrornas in cardiovascular disease: An introduction for clinicians. Heart 2015, 101, 921–928. [Google Scholar] [CrossRef]
- Curtis, H.J.; Sibley, C.R.; Wood, M.J. Mirtrons, an emerging class of atypical mirna. Wiley Interdiscip. Rev. RNA 2012, 3, 617–632. [Google Scholar] [CrossRef] [PubMed]
- Cole, C.; Sobala, A.; Lu, C.; Thatcher, S.R.; Bowman, A.; Brown, J.W.; Green, P.J.; Barton, G.J.; Hutvagner, G. Filtering of deep sequencing data reveals the existence of abundant dicer-dependent small rnas derived from trnas. RNA 2009, 15, 2147–2160. [Google Scholar] [CrossRef] [PubMed]
- Babiarz, J.E.; Hsu, R.; Melton, C.; Thomas, M.; Ullian, E.M.; Blelloch, R. A role for noncanonical micrornas in the mammalian brain revealed by phenotypic differences in dgcr8 versus dicer1 knockouts and small rna sequencing. RNA 2011, 17, 1489–1501. [Google Scholar] [CrossRef] [PubMed]
- Catalanotto, C.; Cogoni, C.; Zardo, G. Microrna in control of gene expression: An overview of nuclear functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Coller, J. What comes first: Translational repression or mrna degradation? The deepening mystery of microrna function. Cell Res. 2012, 22, 1322–1324. [Google Scholar] [CrossRef]
- Hammond, S.M. An overview of micrornas. Adv. Drug Deliv. Rev. 2015, 87, 3–14. [Google Scholar] [CrossRef]
- Mishra, P.J.; Bertino, J.R. Microrna polymorphisms: The future of pharmacogenomics, molecular epidemiology and individualized medicine. Pharmacogenomics 2009, 10, 399–416. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Akiyama, Y.; Yuasa, Y. Multiple-to-multiple relationships between micrornas and target genes in gastric cancer. PLoS ONE 2013, 8, e62589. [Google Scholar] [CrossRef]
- Shastry, B.S. Snps: Impact on gene function and phenotype. Methods Mol. Biol. 2009, 578, 3–22. [Google Scholar]
- Casto, A.M.; Feldman, M.W. Genome-wide association study snps in the human genome diversity project populations: Does selection affect unlinked snps with shared trait associations? PLoS Genet. 2011, 7, e1001266. [Google Scholar] [CrossRef]
- Elfaki, I.; Mir, R.; Almutairi, F.M.; Duhier, F.M.A. Cytochrome p450: Polymorphisms and roles in cancer, diabetes and atherosclerosis. Asian Pac. J. Cancer Prev. 2018, 19, 2057–2070. [Google Scholar] [PubMed]
- Elfaki, I.; Almutairi, F.M.; Mir, R.; Khan, R.; Abu-Duhier, F. Cytochrome p450 cyp1b1*2 gene and its association with t2d in tabuk population, northwestern region of saudi arabia. Asian J. Pharm. Clin. Res. 2018, 11, 55–59. [Google Scholar] [CrossRef]
- Elfaki, I.; Mir, R.; Abu-Duhier, F.M.; Khan, R.; Sakran, M. Phosphatidylinositol 3-kinase glu545lys and his1047tyr mutations are not associated with t2d. Curr. Diabetes Rev. 2019. [Google Scholar] [CrossRef]
- Almutairi, F.M.; Mir, R.; Abu-Duhier, F.; Khan, R.; Harby, K.; Elfaki, I. Slc2a2 gene (glucose transporter 2) variation is associated with an increased risk of developing t2d in an ethnic population of saudi arabia. Indian J. Public Health Res. Dev. 2019, 10, 600–605. [Google Scholar] [CrossRef]
- Xue, A.; Wu, Y.; Zhu, Z.; Zhang, F.; Kemper, K.E.; Zheng, Z.; Yengo, L.; Lloyd-Jones, L.R.; Sidorenko, J.; Wu, Y.; et al. Genome-wide association analyses identify 143 risk variants and putative regulatory mechanisms for type 2 diabetes. Nat. Commun. 2018, 9, 2941. [Google Scholar] [CrossRef]
- Jha, C.K.; Mir, R.; Elfaki, I.; Javid, J.; Babakr, A.T.; Banu, S.; Chahal, S.M.S. Evaluation of the association of omentin 1 rs2274907 a>t and rs2274908 g>a gene polymorphisms with coronary artery disease in indian population: A case control study. J. Pers Med. 2019, 9, 30. [Google Scholar] [CrossRef]
- Mir, R.; Jha, C.K.; Elfaki, I.; Javid, J.; Rehman, S.; Khullar, N.; Banu, S.; Chahal, S.M.S. Incidence of micror-4513c/t gene variability in coronary artery disease—A case-control study. Endocr. Metab. Immune Disord. Drug Targets 2019. [Google Scholar] [CrossRef]
- Jha, C.; Mir, R.; Elfaki, I.; Banu, S. Ldlr Gene Polymorphisms (Rs5925 and Rs1529729) Are Associated with Susceptibility to Coronary Artery Disease in a South Indian Population. Med. Sci. 2019, 7, 80. [Google Scholar] [CrossRef]
- Mir, R.; Jha, C.K.; Elfaki, I.; Rehman, S.; Javid, J.; Khullar, N.; Banu, S.; Chahal, S.M.S. Microrna-224 (rs188519172 a > g) gene variability is associated with a decreased susceptibility to coronary artery disease: A case-control study. MicroRNA 2018, 8, 198–205. [Google Scholar] [CrossRef]
- Jha, C.K.; Mir, R.; Elfaki, I.; Khullar, N.; Rehman, S.; Javid, J.; Banu, S.; Chahal, S.M.S. Potential impact of microrna-423 gene variability in coronary artery disease. Endocr. Metab. Immune Disord. Drug Targets 2019, 19, 67–74. [Google Scholar] [CrossRef]
- Jha, C.K.; Mir, R.; Banu, S.; Elfaki, I.; Chahal, S.M.S. Heterozosity in ldlr rs2228671 and ldlr rs72658855 are associated with increased risk of developing coronary artery disease in india—A case control study. Endocr. Metab. Immune Disord. Drug Targets 2019. [Google Scholar] [CrossRef]
- Baulina, N.M.; Kulakova, O.G.; Favorova, O.O. Micrornas: The role in autoimmune inflammation. Acta Nat. 2016, 8, 21–33. [Google Scholar] [CrossRef]
- Kwak, P.B.; Iwasaki, S.; Tomari, Y. The microrna pathway and cancer. Cancer Sci. 2010, 101, 2309–2315. [Google Scholar] [CrossRef] [PubMed]
- Nariman-Saleh-Fam, Z.; Bastami, M.; Somi, M.H.; Behjati, F.; Mansoori, Y.; Daraei, A.; Saadatian, Z.; Nariman-Saleh-Fam, L.; Mahmoodzadeh, H.; Makhdoumi, Y.; et al. Mirna-related polymorphisms in mir-423 (rs6505162) and pex6 (rs1129186) and risk of esophageal squamous cell carcinoma in an iranian cohort. Genet. Test. Mol. Biomark. 2017, 21, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Moszynska, A.; Gebert, M.; Collawn, J.F.; Bartoszewski, R. Snps in microrna target sites and their potential role in human disease. Open Biol. 2017, 7, 170019. [Google Scholar] [CrossRef]
- Parvin, M.; Jahan, F.; Sarkar, P.K.; Howlader, Z.H.; Nabi, A.; Hosen, M.I. Functional polymorphism located in the microrna binding site of the insulin receptor (insr) gene confers risk for type 2 diabetes mellitus in the bangladeshi population. Biochem. Genet. 2019, 57, 20–33. [Google Scholar] [CrossRef]
- Buraczynska, M.; Zukowski, P.; Wacinski, P.; Ksiazek, K.; Zaluska, W. Polymorphism in microrna-196a2 contributes to the risk of cardiovascular disease in type 2 diabetes patients. J. Diabetes Com. 2014, 28, 617–620. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, M.; Sun, M.; Tang, K.; Zhou, B. The relationship of mir-146a gene polymorphism with carotid atherosclerosis in chinese patients with type 2 diabetes mellitus. Thromb. Res. 2015, 136, 1149–1155. [Google Scholar] [CrossRef]
- Wu, Y.F.; Xu, Q.; He, C.Y.; Li, Y.; Liu, J.W.; Deng, N.; Sun, L.P.; Yuan, Y. Association of polymorphisms in three pri-mirnas that target pepsinogen c with the risk and prognosis of gastric cancer. Sci. Rep. 2017, 7, 39528. [Google Scholar] [CrossRef] [Green Version]
- Rong, G.Q.; Zhang, X.M.; Chen, B.; Yang, X.D.; Wu, H.R.; Gong, W. Microrna gene polymorphisms and the risk of colorectal cancer. Oncol. Lett. 2017, 13, 3617–3623. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Bai, R.; Liu, C.; Ma, C.; Chen, X.; Yang, J.; Sun, D. Microrna single-nucleotide polymorphisms and diabetes mellitus: A comprehensive review. Clin. Genet. 2019, 95, 451–461. [Google Scholar] [CrossRef]
- LaPierre, M.P.; Stoffel, M. Micrornas as stress regulators in pancreatic beta cells and diabetes. Mol. Metab. 2017, 6, 1010–1023. [Google Scholar] [CrossRef] [PubMed]
- Poy, M.N.; Eliasson, L.; Krutzfeldt, J.; Kuwajima, S.; Ma, X.; Macdonald, P.E.; Pfeffer, S.; Tuschl, T.; Rajewsky, N.; Rorsman, P.; et al. A pancreatic islet-specific microrna regulates insulin secretion. Nature 2004, 432, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Baroukh, N.N.; Van Obberghen, E. Function of microrna-375 and microrna-124a in pancreas and brain. FEBS J. 2009, 276, 6509–6521. [Google Scholar] [CrossRef] [PubMed]
- Frost, R.J.; Olson, E.N. Control of glucose homeostasis and insulin sensitivity by the let-7 family of micrornas. Proc. Natl. Acad. Sci. USA 2011, 108, 21075–21080. [Google Scholar] [CrossRef] [Green Version]
- Cerf, M.E. Beta cell dysfunction and insulin resistance. Front. Endocrinol. (Lausanne) 2013, 4, 37. [Google Scholar] [CrossRef] [Green Version]
- Massart, J.; Sjogren, R.J.O.; Lundell, L.S.; Mudry, J.M.; Franck, N.; O’Gorman, D.J.; Egan, B.; Zierath, J.R.; Krook, A. Altered mir-29 expression in type 2 diabetes influences glucose and lipid metabolism in skeletal muscle. Diabetes 2017, 66, 1807–1818. [Google Scholar] [CrossRef] [Green Version]
- Guglielmi, V.; D’Adamo, M.; Menghini, R.; Cardellini, M.; Gentileschi, P.; Federici, M.; Sbraccia, P. Microrna 21 is up-regulated in adipose tissue of obese diabetic subjects. Nutr. Healthy Aging 2017, 4, 141–145. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; An, X.; Niu, L. Role of micrornas in cardiac development and disease. Exp. Ther. Med. 2017, 13, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Hernando, C.; Suarez, Y. Micrornas in endothelial cell homeostasis and vascular disease. Curr. Opin. Hematol. 2018, 25, 227–236. [Google Scholar] [CrossRef]
- Yang, L.; Wang, B.; Zhou, Q.; Wang, Y.; Liu, X.; Liu, Z.; Zhan, Z. Microrna-21 prevents excessive inflammation and cardiac dysfunction after myocardial infarction through targeting kbtbd7. Cell Death Dis. 2018, 9, 769. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.W.; Zhu, X.L.; Wang, B.C.; Hu, J.L.; Sun, J.J.; Wang, S.; Chen, X.J.; Meng, S.P.; Liu, L.; Cheng, Z.Y. Microrna-26b relieves inflammatory response and myocardial remodeling of mice with myocardial infarction by suppression of mapk pathway through binding to ptgs2. Int. J. Cardiol. 2019, 280, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Sethupathy, P.; Collins, F.S. Microrna target site polymorphisms and human disease. Trends Genet. TIG 2008, 24, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Y.; Li, X.; Shi, L.; Tao, W.; Shi, L.; Yang, M.; Wang, X.; Yang, Y.; Yao, Y. Association study of polymorphisms in mirnas with t2dm in chinese population. Int. J. Med. Sci. 2015, 12, 875–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciccacci, C.; Di Fusco, D.; Cacciotti, L.; Morganti, R.; D’Amato, C.; Greco, C.; Rufini, S.; Novelli, G.; Sangiuolo, F.; Spallone, V.; et al. Microrna genetic variations: Association with type 2 diabetes. Acta Diabetol. 2013, 50, 867–872. [Google Scholar] [CrossRef] [PubMed]
- Sebastiani, G.; Po, A.; Miele, E.; Ventriglia, G.; Ceccarelli, E.; Bugliani, M.; Marselli, L.; Marchetti, P.; Gulino, A.; Ferretti, E.; et al. Microrna-124a is hyperexpressed in type 2 diabetic human pancreatic islets and negatively regulates insulin secretion. Acta Diabetol. 2015, 52, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, N.; Tanaka, T. Role of mirnas in the pathogenesis and susceptibility of diabetes mellitus. J. Hum. Genet. 2017, 62, 141–150. [Google Scholar] [CrossRef]
- Erener, S.; Mojibian, M.; Fox, J.K.; Denroche, H.C.; Kieffer, T.J. Circulating mir-375 as a biomarker of beta-cell death and diabetes in mice. Endocrinology 2013, 154, 603–608. [Google Scholar] [CrossRef]
- Jazdzewski, K.; Murray, E.L.; Franssila, K.; Jarzab, B.; Schoenberg, D.R.; de la Chapelle, A. Common snp in pre-mir-146a decreases mature mir expression and predisposes to papillary thyroid carcinoma. Proc. Natl. Acad. Sci. USA 2008, 105, 7269–7274. [Google Scholar] [CrossRef] [Green Version]
- Mann, M.; Mehta, A.; Zhao, J.L.; Lee, K.; Marinov, G.K.; Garcia-Flores, Y.; Baltimore, D. An nf-kappab-microrna regulatory network tunes macrophage inflammatory responses. Nat. Commun. 2017, 8, 851. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.; Santani, D. Role of nf-kappa b in the pathogenesis of diabetes and its associated complications. Pharmacol. Rep. Pr. 2009, 61, 595–603. [Google Scholar] [CrossRef]
- Ndisang, J.F. Role of heme oxygenase in inflammation, insulin-signalling, diabetes and obesity. Mediat. Inflamm. 2010, 2010, 359732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suryavanshi, S.V.; Kulkarni, Y.A. Nf-kappabeta: A potential target in the management of vascular complications of diabetes. Front. Pharmacol. 2017, 8, 798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovis, P.; Roggli, E.; Laybutt, D.R.; Gattesco, S.; Yang, J.Y.; Widmann, C.; Abderrahmani, A.; Regazzi, R. Alterations in microrna expression contribute to fatty acid-induced pancreatic beta-cell dysfunction. Diabetes 2008, 57, 2728–2736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaidonis, G.; Gillies, M.C.; Abhary, S.; Liu, E.; Essex, R.W.; Chang, J.H.; Pal, B.; Sivaprasad, S.; Pefkianaki, M.; Daniell, M.; et al. A single-nucleotide polymorphism in the microrna-146a gene is associated with diabetic nephropathy and sight-threatening diabetic retinopathy in caucasian patients. Acta Diabetol. 2016, 53, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Assmann, T.S.; Duarte, G.C.K.; Brondani, L.A.; de Freitas, P.H.O.; Martins, E.M.; Canani, L.H.; Crispim, D. Polymorphisms in genes encoding mir-155 and mir-146a are associated with protection to type 1 diabetes mellitus. Acta Diabetol. 2017, 54, 433–441. [Google Scholar] [CrossRef]
- Ciccacci, C.; Morganti, R.; Di Fusco, D.; D’Amato, C.; Cacciotti, L.; Greco, C.; Rufini, S.; Novelli, G.; Sangiuolo, F.; Marfia, G.A.; et al. Common polymorphisms in mir146a, mir128a and mir27a genes contribute to neuropathy susceptibility in type 2 diabetes. Acta Diabetol. 2014, 51, 663–671. [Google Scholar] [CrossRef]
- Abo-Elmatty, D.M.; Mehanna, E.T. Mir146a rs2910164 (g/c) polymorphism is associated with incidence of preeclampsia in gestational diabetes patients. Biochem. Genet. 2019, 57, 222–233. [Google Scholar] [CrossRef]
- Dziedzic, M.; Powrozek, T.; Orlowska, E.; Koch, W.; Kukula-Koch, W.; Gawel, K.; Bednarek-Skublewska, A.; Malecka-Massalska, T.; Milanowski, J.; Petkowicz, B.; et al. Relationship between microrna-146a expression and plasma renalase levels in hemodialyzed patients. PLoS ONE 2017, 12, e0179218. [Google Scholar] [CrossRef]
- Desir, G.V. Role of renalase in the regulation of blood pressure and the renal dopamine system. Curr. Opin. Nephrol. Hypertens. 2011, 20, 31–36. [Google Scholar] [CrossRef]
- Ghaedi, H.; Tabasinezhad, M.; Alipoor, B.; Shokri, F.; Movafagh, A.; Mirfakhraie, R.; Omrani, M.D.; Masotti, A. The pre-mir-27a variant rs895819 may contribute to type 2 diabetes mellitus susceptibility in an iranian cohort. J. Endocrinol. Investig. 2016, 39, 1187–1193. [Google Scholar] [CrossRef]
- Offer, S.M.; Butterfield, G.L.; Jerde, C.R.; Fossum, C.C.; Wegner, N.J.; Diasio, R.B. Micrornas mir-27a and mir-27b directly regulate liver dihydropyrimidine dehydrogenase expression through two conserved binding sites. Mol. Cancer Ther. 2014, 13, 742–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.; Gu, H.; Zeng, Y.; Xia, Y.; Wang, Y.; Jing, Y.; Yang, L.; Wang, B. Hsa-mir-27a genetic variant contributes to gastric cancer susceptibility through affecting mir-27a and target gene expression. Cancer Sci. 2010, 101, 2241–2247. [Google Scholar] [CrossRef]
- Qin, L.; Chen, Y.; Niu, Y.; Chen, W.; Wang, Q.; Xiao, S.; Li, A.; Xie, Y.; Li, J.; Zhao, X.; et al. A deep investigation into the adipogenesis mechanism: Profile of micrornas regulating adipogenesis by modulating the canonical wnt/beta-catenin signaling pathway. BMC Genom. 2010, 11, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Kim, A.Y.; Lee, H.W.; Son, Y.H.; Lee, G.Y.; Lee, J.W.; Lee, Y.S.; Kim, J.B. Mir-27a is a negative regulator of adipocyte differentiation via suppressing ppargamma expression. Biochem. Biophys. Res. Commun. 2010, 392, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Ye, J. Mechanisms of insulin resistance in obesity. Front. Med. 2013, 7, 14–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.T.; Chen, Y.J.; Sun, L.L.; Zhang, S.J.; Zhou, Z.Y.; Qiao, H. Affection of single-nucleotide polymorphisms in mir-27a, mir-124a, and mir-146a on susceptibility to type 2 diabetes mellitus in chinese han people. Chin. Med. J. 2015, 128, 533–539. [Google Scholar] [CrossRef]
- Locke, J.M.; Lango Allen, H.; Harries, L.W. A rare snp in pre-mir-34a is associated with increased levels of mir-34a in pancreatic beta cells. Acta Diabetol. 2014, 51, 325–329. [Google Scholar] [CrossRef] [Green Version]
- Regazzi, R.; Sadoul, K.; Meda, P.; Kelly, R.B.; Halban, P.A.; Wollheim, C.B. Mutational analysis of vamp domains implicated in ca2+-induced insulin exocytosis. EMBO J. 1996, 15, 6951–6959. [Google Scholar] [CrossRef] [Green Version]
- Hastoy, B.; Scotti, P.A.; Milochau, A.; Fezoua-Boubegtiten, Z.; Rodas, J.; Megret, R.; Desbat, B.; Laguerre, M.; Castano, S.; Perrais, D.; et al. A central small amino acid in the vamp2 transmembrane domain regulates the fusion pore in exocytosis. Sci. Rep. 2017, 7, 2835. [Google Scholar] [CrossRef] [Green Version]
- Roggli, E.; Gattesco, S.; Caille, D.; Briet, C.; Boitard, C.; Meda, P.; Regazzi, R. Changes in microrna expression contribute to pancreatic beta-cell dysfunction in prediabetic nod mice. Diabetes 2012, 61, 1742–1751. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Peng, R.; Li, T.; Luo, X.; Peng, H.; Zha, H.; Yin, P.; Wen, L.; Zhang, Z. A potentially functional polymorphism in the regulatory region of let-7a-2 is associated with an increased risk for diabetic nephropathy. Gene 2013, 527, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, A.P.; Sharma, K. Transcription factors in the pathogenesis of diabetic nephropathy. Expert Rev. Mol. Med. 2009, 11, e13. [Google Scholar] [CrossRef] [PubMed]
- Faraoni, I.; Antonetti, F.R.; Cardone, J.; Bonmassar, E. Mir-155 gene: A typical multifunctional microrna. Biochim. Biophys. Acta 2009, 1792, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Becker Buscaglia, L.E.; Barker, J.R.; Li, Y. Micrornas in nf-kappab signaling. J. Mol. Cell Biol. 2011, 3, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Caruso, M.; Ma, D.; Msallaty, Z.; Lewis, M.; Seyoum, B.; Al-janabi, W.; Diamond, M.; Abou-Samra, A.B.; Hojlund, K.; Tagett, R.; et al. Increased interaction with insulin receptor substrate 1, a novel abnormality in insulin resistance and type 2 diabetes. Diabetes 2014, 63, 1933–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derosa, G.; Maffioli, P. Peroxisome proliferator-activated receptor-gamma (ppar-gamma) agonists on glycemic control, lipid profile and cardiovascular risk. Curr Mol. Pharm. 2012, 5, 272–281. [Google Scholar] [CrossRef]
- Wu, B.; Liu, G.; He, F.; Liu, R.; Wang, Z.; Wang, Y.; Zhou, H.; Zhang, W. Mir-3188 (rs7247237-c>t) single-nucleotide polymorphism is associated with the incidence of vascular complications in chinese patients with type 2 diabetes. J. Cardiovasc. Pharmacol. 2019, 74, 62–70. [Google Scholar] [CrossRef]
- Sharma, A.; Sellers, S.; Stefanovic, N.; Leung, C.; Tan, S.M.; Huet, O.; Granville, D.J.; Cooper, M.E.; de Haan, J.B.; Bernatchez, P. Direct endothelial nitric oxide synthase activation provides atheroprotection in diabetes-accelerated atherosclerosis. Diabetes 2015, 64, 3937–3950. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Liu, E.; Li, W.; Cui, J.; Li, T. Mir-3188 inhibits non-small cell lung cancer cell proliferation through foxo1-mediated mtor-p-pi3k/akt-c-jun signaling pathway. Front. Pharmacol. 2018, 9, 1362. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Liu, G.; Guo, J.; Su, Z. The pi3k/akt pathway in obesity and type 2 diabetes. Int. J. Biol. Sci. 2018, 14, 1483–1496. [Google Scholar] [CrossRef] [Green Version]
- McAuley, A.K.; Dirani, M.; Wang, J.J.; Connell, P.P.; Lamoureux, E.L.; Hewitt, A.W. A genetic variant regulating mir-126 is associated with sight threatening diabetic retinopathy. Diabetes Vasc. Dis. Res. 2015, 12, 133–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fish, J.E.; Santoro, M.M.; Morton, S.U.; Yu, S.; Yeh, R.F.; Wythe, J.D.; Ivey, K.N.; Bruneau, B.G.; Stainier, D.Y.; Srivastava, D. Mir-126 regulates angiogenic signaling and vascular integrity. Dev. Cell 2008, 15, 272–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaman, S.; Leppanen, V.M.; Alitalo, K. Vascular endothelial growth factor signaling in development and disease. Development 2018, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, Y.; Guo, J.; Peng, Y.F.; Tan, T.; Huang, H.T.; Luo, H.C.; Wei, Y.S. Association of mir-21, mir-126 and mir-605 gene polymorphisms with ischemic stroke risk. Oncotarget 2017, 8, 95755–95763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sessa, R.; Seano, G.; di Blasio, L.; Gagliardi, P.A.; Isella, C.; Medico, E.; Cotelli, F.; Bussolino, F.; Primo, L. The mir-126 regulates angiopoietin-1 signaling and vessel maturation by targeting p85beta. Biochim. Biophys. Acta 2012, 1823, 1925–1935. [Google Scholar] [CrossRef] [Green Version]
- Fragoso, J.M.; Ramirez-Bello, J.; Martinez-Rios, M.A.; Pena-Duque, M.A.; Posadas-Sanchez, R.; Delgadillo-Rodriguez, H.; Jimenez-Morales, M.; Posadas-Romero, C.; Vargas-Alarcon, G. Mir-196a2 (rs11614913) polymorphism is associated with coronary artery disease, but not with in-stent coronary restenosis. Inflamm. Res. J. Eur. Histamine Res. Soc. 2019, 68, 215–221. [Google Scholar] [CrossRef]
- Hoffman, A.E.; Zheng, T.; Yi, C.; Leaderer, D.; Weidhaas, J.; Slack, F.; Zhang, Y.; Paranjape, T.; Zhu, Y. Microrna mir-196a-2 and breast cancer: A genetic and epigenetic association study and functional analysis. Cancer Res. 2009, 69, 5970–5977. [Google Scholar] [CrossRef]
- Ross, R.L.; Burns, J.E.; Taylor, C.F.; Mellor, P.; Anderson, D.H.; Knowles, M.A. Identification of mutations in distinct regions of p85 alpha in urothelial cancer. PLoS ONE 2013, 8, e84411. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; You, L.; Zhou, R.; Zhang, J. Significant association between functional microrna polymorphisms and coronary heart disease susceptibility: A comprehensive meta-analysis involving 16484 subjects. Oncotarget 2017, 8, 5692–5702. [Google Scholar] [CrossRef] [Green Version]
- Pin, A.L.; Houle, F.; Fournier, P.; Guillonneau, M.; Paquet, E.R.; Simard, M.J.; Royal, I.; Huot, J. Annexin-1-mediated endothelial cell migration and angiogenesis are regulated by vascular endothelial growth factor (vegf)-induced inhibition of mir-196a expression. J. Biol. Chem. 2012, 287, 30541–30551. [Google Scholar] [CrossRef] [Green Version]
- Purvis, G.S.D.; Chiazza, F.; Chen, J.; Azevedo-Loiola, R.; Martin, L.; Kusters, D.H.M.; Reutelingsperger, C.; Fountoulakis, N.; Gnudi, L.; Yaqoob, M.M.; et al. Annexin a1 attenuates microvascular complications through restoration of akt signalling in a murine model of type 1 diabetes. Diabetologia 2018, 61, 482–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jong, R.; Leoni, G.; Drechsler, M.; Soehnlein, O. The advantageous role of annexin a1 in cardiovascular disease. Cell Adhes. Migr. 2017, 11, 261–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tousoulis, D.; Oikonomou, E.; Economou, E.K.; Crea, F.; Kaski, J.C. Inflammatory cytokines in atherosclerosis: Current therapeutic approaches. Eur. Heart J. 2016, 37, 1723–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Chen, L.; Chen, D.; Wu, X.; Chen, M. Influence of microrna-related polymorphisms on clinical outcomes in coronary artery disease. Am. J. Transl. Res. 2015, 7, 393–400. [Google Scholar]
- Fawzy, M.S.; Toraih, E.A.; Hamed, E.O.; Hussein, M.H.; Ismail, H.M. Association of mir-499a expression and seed region variant (rs3746444) with cardiovascular disease in egyptian patients. Acta Cardiol. 2018, 73, 131–140. [Google Scholar] [CrossRef]
- Chen, W.; Shao, D.; Gu, H.; Gong, J.; Zhang, J. Hsa-mir-499 rs3746444 t/c polymorphism is associated with increased risk of coronary artery disease in a chinese population. Acta Cardiol. Sin. 2017, 33, 34–40. [Google Scholar]
- Xin, Y.; Yang, C.; Han, Z. Circulating mir-499 as a potential biomarker for acute myocardial infarction. Ann. Transl. Med. 2016, 4, 135. [Google Scholar] [CrossRef] [Green Version]
- Wilson, K.D.; Hu, S.; Venkatasubrahmanyam, S.; Fu, J.D.; Sun, N.; Abilez, O.J.; Baugh, J.J.; Jia, F.; Ghosh, Z.; Li, R.A.; et al. Dynamic microrna expression programs during cardiac differentiation of human embryonic stem cells: Role for mir-499. Circ. Cardiovasc. Genet. 2010, 3, 426–435. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Jia, Z.; Zhang, C.; Sun, M.; Wang, W.; Chen, P.; Ma, K.; Zhang, Y.; Li, X.; Zhou, C. Mir-499 protects cardiomyocytes from h 2o 2-induced apoptosis via its effects on pdcd4 and pacs2. RNA Biol. 2014, 11, 339–350. [Google Scholar] [CrossRef] [Green Version]
- Hosoda, T.; Zheng, H.; Cabral-da-Silva, M.; Sanada, F.; Ide-Iwata, N.; Ogorek, B.; Ferreira-Martins, J.; Arranto, C.; D’Amario, D.; del Monte, F.; et al. Human cardiac stem cell differentiation is regulated by a mircrine mechanism. Circulation 2011, 123, 1287–1296. [Google Scholar] [CrossRef] [Green Version]
- Hashemi, M.; Eskandari-Nasab, E.; Zakeri, Z.; Atabaki, M.; Bahari, G.; Jahantigh, M.; Taheri, M.; Ghavami, S. Association of pre-mirna-146a rs2910164 and premirna-499 rs3746444 polymorphisms and susceptibility to rheumatoid arthritis. Mol. Med. Rep. 2013, 7, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.X.; Jiao, J.Q.; Li, Q.; Long, B.; Wang, K.; Liu, J.P.; Li, Y.R.; Li, P.F. Mir-499 regulates mitochondrial dynamics by targeting calcineurin and dynamin-related protein-1. Nat. Med. 2011, 17, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Fox, R.J.; Burwell, L.S.; Yoon, Y. Regulation of mitochondrial fission and apoptosis by the mitochondrial outer membrane protein hfis1. J. Cell Sci. 2005, 118, 4141–4151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.X.; Weng, H.; Zheng, J.; Feng, Z.H.; Ou, J.L.; Liao, W.J. Association between micrornas polymorphisms and risk of ischemic stroke: A meta-analysis in chinese individuals. Front. Aging Neurosci. 2018, 10, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madjid, M.; Willerson, J.T. Inflammatory markers in coronary heart disease. Br. Med. Bull. 2011, 100, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Li, X.; Zhou, B. A meta-analysis of mir-499 rs3746444 polymorphism for cancer risk of different systems: Evidence from 65 case-control studies. Front. Physiol. 2018, 9, 737. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Pan, G. Association of rs2910164 polymorphism in mirna-146 and rs3746444 polymorphism in mirna-499 with inflammatory arthritis: A meta-analysis. Biomed. Res. Int. 2019, 2019, 7305750. [Google Scholar] [CrossRef] [Green Version]
- Ghanbari, M.; de Vries, P.S.; de Looper, H.; Peters, M.J.; Schurmann, C.; Yaghootkar, H.; Dorr, M.; Frayling, T.M.; Uitterlinden, A.G.; Hofman, A.; et al. A genetic variant in the seed region of mir-4513 shows pleiotropic effects on lipid and glucose homeostasis, blood pressure, and coronary artery disease. Hum. Mutat. 2014, 35, 1524–1531. [Google Scholar] [CrossRef]
- Heni, M.; Haupt, A.; Schafer, S.A.; Ketterer, C.; Thamer, C.; Machicao, F.; Stefan, N.; Staiger, H.; Haring, H.U.; Fritsche, A. Association of obesity risk snps in pcsk1 with insulin sensitivity and proinsulin conversion. BMC Med. Genet. 2010, 11, 86. [Google Scholar] [CrossRef] [Green Version]
- Pan, S.; Guan, G.C.; Lv, Y.; Liu, Z.W.; Liu, F.Q.; Zhang, Y.; Zhu, S.M.; Zhang, R.H.; Zhao, N.; Shi, S.; et al. G-t haplotype established by rs3785889-rs16941382 in gosr2 gene is associated with coronary artery disease in chinese han population. Oncotarget 2017, 8, 82165–82173. [Google Scholar] [CrossRef] [Green Version]
- Pan, S.; Nakayama, T.; Sato, N.; Izumi, Y.; Soma, M.; Aoi, N.; Ma, Y.; Hinohara, S.; Doba, N. A haplotype of the gosr2 gene is associated with myocardial infarction in japanese men. Genet. Test. Mol. Biomark. 2013, 17, 481–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Cappello, T.; Wang, L. Emerging role of micrornas in lipid metabolism. Acta Pharm. Sin. B 2015, 5, 145–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Lv, Z.; Deng, Q.; Li, L.; Yang, B.; Feng, J.; Wu, T.; Zhang, X.; Cheng, J. A genetic variant in pre-mir-146a (rs2910164 c>g) is associated with the decreased risk of acute coronary syndrome in a chinese population. Tohoku J. Exp. Med. 2015, 237, 227–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramkaran, P.; Khan, S.; Phulukdaree, A.; Moodley, D.; Chuturgoon, A.A. Mir-146a polymorphism influences levels of mir-146a, irak-1, and traf-6 in young patients with coronary artery disease. Cell Biochem. Biophys. 2014, 68, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.H.; Kim, S.H.; Yang, W.I.; Kim, W.J.; Moon, J.Y.; Kim, I.J.; Cha, D.H.; Cho, S.Y.; Kim, J.O.; Kim, K.A.; et al. Mirna polymorphisms (mir146a, mir149, mir196a2 and mir499) are associated with the risk of coronary artery disease. Mol. Med. Rep. 2016, 14, 2328–2342. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.S.; Sivachandran, N.; Lau, A.; Boudreau, E.; Zhao, J.L.; Baltimore, D.; Delgado-Olguin, P.; Cybulsky, M.I.; Fish, J.E. Microrna-146 represses endothelial activation by inhibiting pro-inflammatory pathways. Embo Mol. Med. 2013, 5, 1017–1034. [Google Scholar] [CrossRef]
- Saba, R.; Sorensen, D.L.; Booth, S.A. Microrna-146a: A dominant, negative regulator of the innate immune response. Front. Immunol. 2014, 5, 578. [Google Scholar] [CrossRef]
- Hou, J.; Wang, P.; Lin, L.; Liu, X.; Ma, F.; An, H.; Wang, Z.; Cao, X. Microrna-146a feedback inhibits rig-i-dependent type i ifn production in macrophages by targeting traf6, irak1, and irak2. J. Immunol. 2009, 183, 2150–2158. [Google Scholar] [CrossRef] [Green Version]
- Fioranelli, M.; Bottaccioli, A.G.; Bottaccioli, F.; Bianchi, M.; Rovesti, M.; Roccia, M.G. Stress and inflammation in coronary artery disease: A review psychoneuroendocrineimmunology-based. Front. Immunol. 2018, 9, 2031. [Google Scholar] [CrossRef]
- Pamukcu, B.; Lip, G.Y.; Shantsila, E. The nuclear factor--kappa b pathway in atherosclerosis: A potential therapeutic target for atherothrombotic vascular disease. Thromb. Res. 2011, 128, 117–123. [Google Scholar] [CrossRef]
- Qu, J.Y.; Xi, J.; Zhang, Y.H.; Zhang, C.N.; Song, L.; Song, Y.; Hui, R.T.; Chen, J.Z. Association of the microrna-146a snp rs2910164 with ischemic stroke incidence and prognosis in a chinese population. Int. J. Mol. Sci. 2016, 17, 660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaffarzadeh, M.; Ghaedi, H.; Alipoor, B.; Omrani, M.D.; Kazerouni, F.; Shanaki, M.; Labbaf, A.; Pashaiefar, H.; Rahimipour, A. Association of mir-149 (rs2292832) variant with the risk of coronary artery disease. J. Med. Biochem. 2017, 36, 251–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, S.L.; Wang, J.X.; Jiao, J.Q.; Tu, X.; Wang, Q.; Liu, F.; Li, Q.; Gao, J.; Zhou, Q.Y.; Gu, D.F.; et al. A pre-microrna-149 (mir-149) genetic variation affects mir-149 maturation and its ability to regulate the puma protein in apoptosis. J. Biol. Chem. 2013, 288, 26865–26877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Rooij, E.; Sutherland, L.B.; Thatcher, J.E.; DiMaio, J.M.; Naseem, R.H.; Marshall, W.S.; Hill, J.A.; Olson, E.N. Dysregulation of micrornas after myocardial infarction reveals a role of mir-29 in cardiac fibrosis. Proc. Natl. Acad. Sci. USA 2008, 105, 13027–13032. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Zhang, Z.; Xing, Y.; Wei, J.; Ge, Z.; Liu, X.; Zhang, Y.; Huang, X. Microrna-149 negatively regulates tlr-triggered inflammatory response in macrophages by targeting myd88. J. Cell Biochem. 2014, 115, 919–927. [Google Scholar] [CrossRef]
- Kuppusamy, K.T.; Jones, D.C.; Sperber, H.; Madan, A.; Fischer, K.A.; Rodriguez, M.L.; Pabon, L.; Zhu, W.Z.; Tulloch, N.L.; Yang, X.; et al. Let-7 family of microrna is required for maturation and adult-like metabolism in stem cell-derived cardiomyocytes. Proc. Natl. Acad. Sci. USA 2015, 112, 2785–2794. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Hong, H.; Chen, L.; Shi, X.; Chen, Y.; Weng, Q. Association of microrna polymorphisms with the risk of myocardial infarction in a chinese population. Tohoku J. Exp. Med. 2014, 233, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Bastami, M.; Ghaderian, S.M.; Omrani, M.D.; Mirfakhraie, R.; Vakili, H.; Parsa, S.A.; Nariman-Saleh-Fam, Z.; Masotti, A. Mirna-related polymorphisms in mir-146a and tcf21 are associated with increased susceptibility to coronary artery disease in an iranian population. Genet. Test. Mol. Biomark. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Feng, J.; Xing, W.; Xie, L. Regulatory roles of micrornas in diabetes. Int. J. Mol. Sci. 2016, 17, 1729. [Google Scholar] [CrossRef] [PubMed]
- Moeez, S.; Riaz, S.; Masood, N.; Kanwal, N.; Arif, M.A.; Niazi, R.; Khalid, S. Evaluation of the rs3088442 g > a slc22a3 gene polymorphism and the role of microrna 147 in groups of adult pakistani populations with type 2 diabetes in response to metformin. Can. J. Diabetes 2019, 43, 128–135. [Google Scholar] [CrossRef]
- Chen, L.; Hong, C.; Chen, E.C.; Yee, S.W.; Xu, L.; Almof, E.U.; Wen, C.; Fujii, K.; Johns, S.J.; Stryke, D.; et al. Genetic and epigenetic regulation of the organic cation transporter 3, slc22a3. Pharm. J. 2013, 13, 110–120. [Google Scholar]
- O’Connell, R.M.; Rao, D.S.; Baltimore, D. Microrna regulation of inflammatory responses. Annu. Rev. Immunol. 2012, 30, 295–312. [Google Scholar] [CrossRef] [PubMed]
- Bush, W.S.; Moore, J.H. Chapter 11: Genome-wide association studies. PLoS Comput. Biol. 2012, 8, e1002822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, N.; Shi, J.; Garcia-Closas, M. Developing and evaluating polygenic risk prediction models for stratified disease prevention. Nat. Rev. Genet. 2016, 17, 392–406. [Google Scholar] [CrossRef]
- Lyssenko, V.; Laakso, M. Genetic screening for the risk of type 2 diabetes: Worthless or valuable? Diabetes Care 2013, 36 (Suppl. 2), S120–S126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knowles, J.W.; Ashley, E.A. Cardiovascular disease: The rise of the genetic risk score. PLoS Med. 2018, 15, e1002546. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| miRNA | Rs Number | Disease | No of Control/Patients | Population | Ref |
|---|---|---|---|---|---|
| 1-MiR-124a | rs531564 | T2DM | 610/738 | Chinese | [54,55] |
| T2DM | 185/163 | Italian | |||
| 2-MiR-146a | rs2910164 | T2DM | 610/738 | Chinese | [54,65,67] |
| T1DM (DN) and T2DM (DME) | −/2948 | Caucasian | |||
| T2DM (DPN) | −/132 | Italian | |||
| 3-MiR-27a | rs895819 | T2DM | 185/163 | Italian | [55,71,77] |
| T2DM | 209/204 | Iranian | |||
| T2DM | 967/995 | Chinese Han | |||
| 4-MiR-34a | rs72631823 | T2DM | - | INS-1, MIN6 and Hela cell lines | [78] |
| 5-MIRLET7A2 | rs1143770 | T2DM/DN | 62/212 | Chinese | [82] |
| 6-MiR128a | rs11888095 | DPN | −/132 | Italian | [67] |
| MiRNA | rs Number | Disease | No. of Control/Patients | Population | Ref |
|---|---|---|---|---|---|
| 1-MiR-196a | rs11614913 | CVD | 834/920 | Polish (With T2DM) | [37] |
| 2-MiR-499 | rs3746444 | CAD | −/1004 | Chinese | [105,107,115,138] |
| IS | Meta-analysis | Chinese | |||
| CAD | 480/435 | Chinese | |||
| MI | 889/919 | Chinese | |||
| 3-MiR-4513 | rs2168518 | CAD | −/1004 | Chinese | [105] |
| 4-Pre-MiR-27a | rs895819 | MI | 287/646 | Chinese Han population | [5] |
| 5-Pre-MiR-146a | rs2910164 | ACS | 721/722 | Chinese | [100,124,126,139] |
| CAD | 535/522 | Korean | |||
| CAD | 300/300 | Iranian | |||
| 6-MiR-149 | rs2292832 | CAD | 535/522 | Korean | [126] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elfaki, I.; Mir, R.; Mir, M.M.; AbuDuhier, F.M.; Babakr, A.T.; Barnawi, J. Potential Impact of MicroRNA Gene Polymorphisms in the Pathogenesis of Diabetes and Atherosclerotic Cardiovascular Disease. J. Pers. Med. 2019, 9, 51. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm9040051
Elfaki I, Mir R, Mir MM, AbuDuhier FM, Babakr AT, Barnawi J. Potential Impact of MicroRNA Gene Polymorphisms in the Pathogenesis of Diabetes and Atherosclerotic Cardiovascular Disease. Journal of Personalized Medicine. 2019; 9(4):51. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm9040051
Chicago/Turabian StyleElfaki, Imadeldin, Rashid Mir, Mohammad Muzaffar Mir, Faisel M AbuDuhier, Abdullatif Taha Babakr, and Jameel Barnawi. 2019. "Potential Impact of MicroRNA Gene Polymorphisms in the Pathogenesis of Diabetes and Atherosclerotic Cardiovascular Disease" Journal of Personalized Medicine 9, no. 4: 51. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm9040051