
Behaviour of Zoo-Housed Red Pandas (Ailurus fulgens): A Case-Study Testing the Behavioural Variety Index


Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Area
2.2. Procedure and Data Collection
2.3. The Behavioural Variety Index (BVI)
2.4. Data Analysis
3. Results
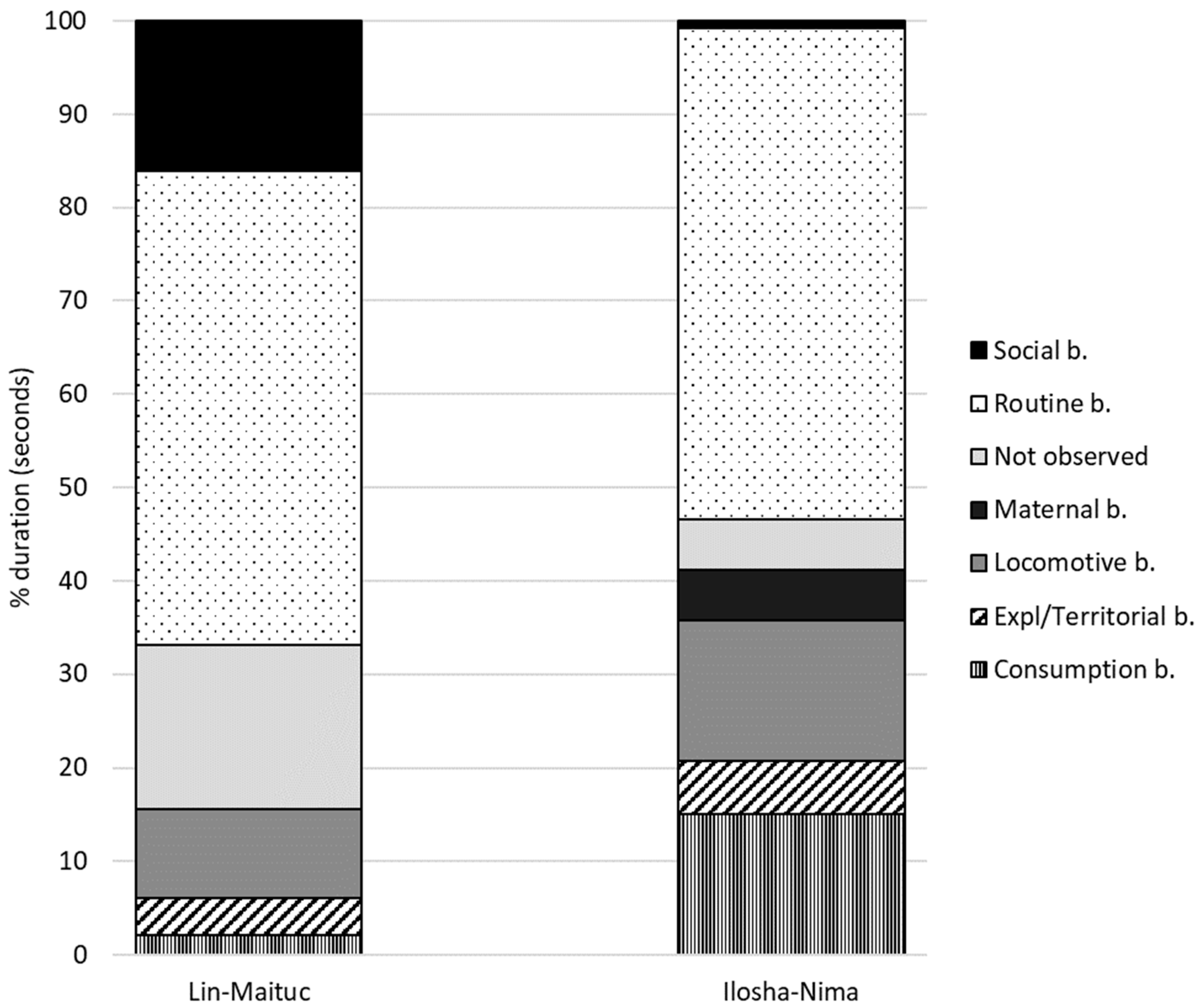
3.1. Lin and Maituc Pair
3.2. Ilosha and Ny’ma Pair
3.3. Morning vs. Afternoon Sessions
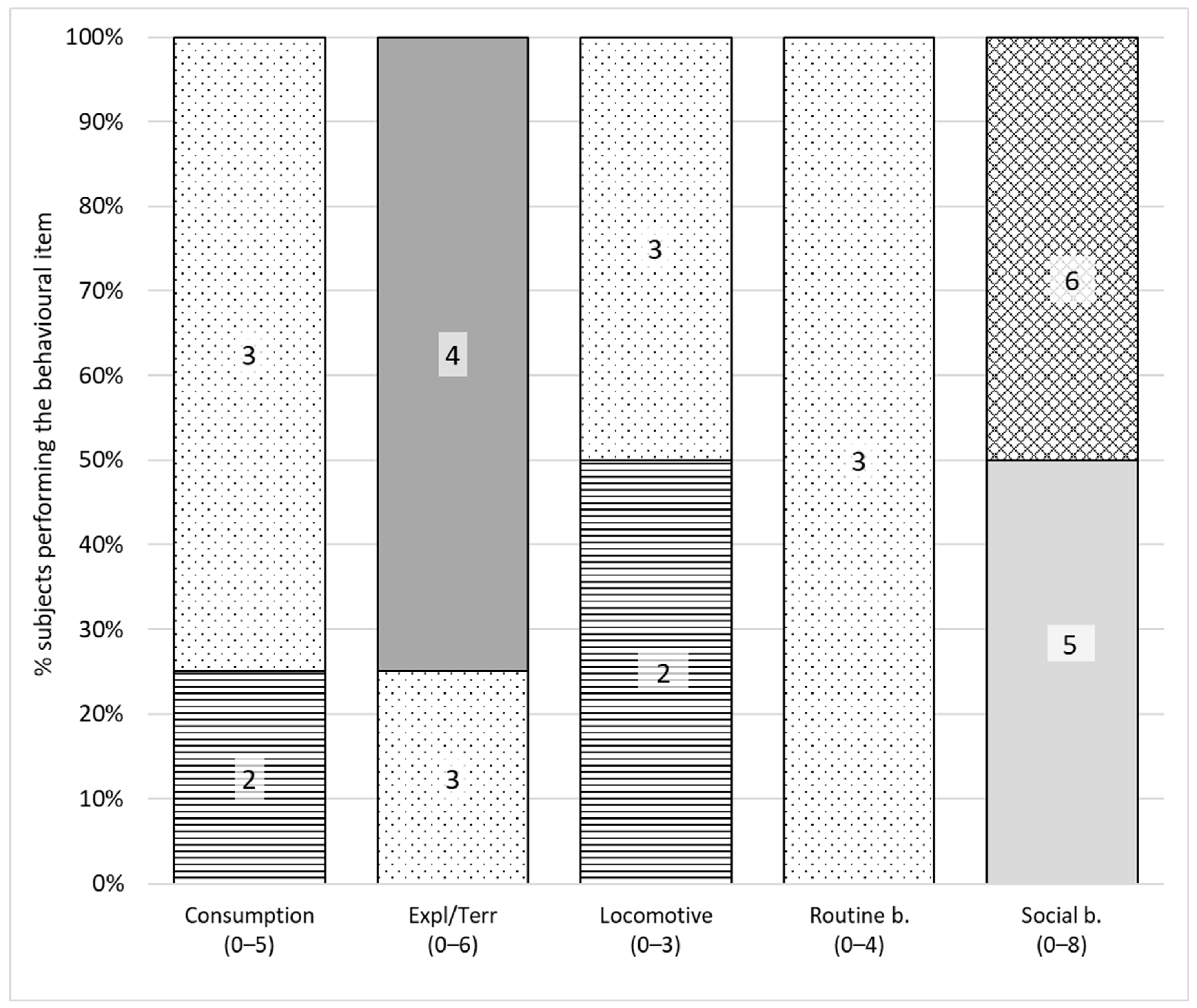
3.4. Behavioural Variety
3.5. Maternal Behaviour
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kappelhof, J.; Weerman, J. The development of the red panda Ailurus fulgens EEP: From a failing captive population to a stable population that provides effective support to in situ conservation. Int. Zoo Yearb. 2020, 54, 102–112. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, J.; Yang, J.; Li, M.; Wei, F. Food habits and space-use of red pandas Ailurus fulgens in the Fengtongzhai Nature Reserve, China: Food effects and behavioural responses. Acta Theriol. (Warsz.) 2009, 54, 225–234. [Google Scholar] [CrossRef]
- Hill, S.P.; Broom, D.M. Measuring zoo animal welfare: Theory and practice. Zoo Biol. 2009, 28, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Bacon, H. Behaviour-based husbandry—A holistic approach to the management of abnormal repetitive behaviors. Animals 2018, 8, 103. [Google Scholar] [CrossRef] [Green Version]
- Hosey, G.; Pankhurst, S.; Melfi, V. Zoo Animals: Behaviour, Management, and Welfare; Oxford University Press: New York, NY, USA, 2013. [Google Scholar]
- McCormick, W. Recognizing and Assessing Positive Welfare: Developing Positive Indicators for Use in Welfare Assessment. In Proceedings of the Measuring Behavior, Utrecht, The Netherlands, 28–31 August 2012. [Google Scholar]
- Spiezio, C.; Valsecchi, V.; Sandri, C.; Regaiolli, B. Investigating individual and social behaviour of the Northern bald ibis (Geronticus eremita): Behavioural variety and welfare. PeerJ 2018, 6, e5436. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.J.; Pisacane, C.B.; Vicino, G.A. Relationship between behavioural diversity and faecal glucocorticoid metabolites: A case study with cheetahs (Acinonyx jubatus). Anim. Welf. 2016, 25, 325–329. [Google Scholar] [CrossRef] [Green Version]
- Mason, G.J.; Latham, N.R. Can’t stop, won’t stop: Is stereotypy a reliable animal welfare indicator? Anim. Welf. 2004, 13, 57–69. [Google Scholar]
- Duncan, I.J. Behavior and behavioral needs. Poult. Sci. 1998, 77, 1766–1772. [Google Scholar] [CrossRef] [PubMed]
- Andres-Bray, T.; Moller, P.; Powell, D.M. Preliminary model of personality structure in captive red pandas (Ailurus fulgens). JZAR 2020, 8, 29–36. [Google Scholar] [CrossRef]
- Conover, G.K.; Gittleman, J.L. Scent-marking in captive red pandas (Ailurus fulgens). Zoo Biol. 1989, 8, 193–205. [Google Scholar] [CrossRef]
- EAZA Small Carnivor Tag. EAZA Best Practice Guidelines Red Panda (Ailurus fulgens). EAZA 2013, 1, 1–42. [Google Scholar]
- Reid, D.G.; Jinchu, H.; Yan, H. Ecology of the red panda Ailurus fulgens in the Wolong Reserve, China. J. Zool. 1991, 225, 347–364. [Google Scholar] [CrossRef]
- Wei, F.; Zhang, Z. Red Panda Ecology. In Red Panda: Biology and Conservation of the First Panda; Glatston, A.R., Ed.; Academic Press: Cambridge, MA, USA, 2021; pp. 193–212. [Google Scholar]
- Glatston, A.R. Red Panda: Biology and Conservation of the First Panda; Academic Press: London, UK, 2021. [Google Scholar]
- Gittleman, J.L. Behavioral Energetics of Lactation in an Herbivorous Carnivore, the Red Panda (Ailurus fulgens). Ethology 1988, 79, 13–24. [Google Scholar] [CrossRef]
- Delfour, F.; Vaicekauskaite, R.; García-Párraga, D.; Pilenga, C.; Serres, A.; Brasseur, I.; Pascaud, A.; Perlado-Campos, E.; Sánchez-Contreras, G.J.; Baumgartner, K.; et al. Behavioural diversity study in bottlenose dolphin (Tursiops truncatus) groups and its implications for welfare assessments. Animals 2021, 11, 1715. [Google Scholar] [CrossRef]
- Princée, F.P.G.; Glatston, A.R. Influence of climate on the survivorship of neonatal red pandas in captivity. Zoo Biol. 2016, 35, 104–110. [Google Scholar] [CrossRef]
- Goswami, S.; Tyagi, P.C.; Malik, P.K.; Pandit, S.J.; Kadivar, R.F.; Fitzpatrick, M.; Mondol, S. Effects of personality and rearing-history on the welfare of captive Asiatic lions (Panthera leo persica). PeerJ 2020, 8, e8425. [Google Scholar] [CrossRef] [Green Version]
- Spiezio, C.; Vaglio, S.; Vandelle, C.; Sandri, C.; Regaiolli, B. Effects of rearing on the behaviour of zoo-housed chimpanzees (Pan troglodytes). Folia Primatol. 2021, 92, 91–102. [Google Scholar] [CrossRef]
- Altmann, J. Observational study of behaviour: Sampling methods. Behaviour 1974, 49, 227–266. [Google Scholar] [CrossRef] [Green Version]
- Jule, K.R. Effects of Captivity and Implications of Ex Situ Conservation: With Special Reference the Red Panda (Ailurus fulgens). Ph.D. Dissertation, University of Exeter, Exeter, UK, 2008. [Google Scholar]
- Bray, T.C. Assessing Behavioral Syndromes in Captive Red Pandas (Ailurus fulgens) Using an Ethological Approach. CUNY Acad. Works 2017, 180, 1–46. Available online: https://academicworks.cuny.edu/hc_sas_etds/ (accessed on 30 October 2021).
- Roberts, B.M.S.; Gittleman, J.L. Ailurus fulgens. Mamm. Species 1984, 222, 1–8. [Google Scholar] [CrossRef]
- van de Bunte, W.; Weerman, J.; Hof, A.R. Potential effects of GPS collars on the behaviour of two red pandas (Ailurus fulgens) in Rotterdam Zoo. PLoS ONE 2021, 16, e0252456. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, J.; Han, Z.; Wei, F. Activity patterns of wild red pandas in Fengtongzhai Nature Reserve, China. Ital. J. Zool. 2011, 78, 398–404. [Google Scholar] [CrossRef]
- Sanders, K.; Fernandez, E.J. Behavioral implications of enrichment for golden lion tamarins: A tool for ex situ conservation. J. Appl. Anim. Welf. Sci. 2020, 25, 1–10. [Google Scholar] [CrossRef]
- Bateson, M.; Martin, P. Measuring Behaviour: An Introductory Guide, 4th ed.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar] [CrossRef]
- Rose, P.E.; Scales, J.S.; Brereton, J.E. Why the “Visitor Effect” is complicated. unravelling individual animal, visitor number, and climatic influences on behavior, space use and interactions with keepers—A Case Study on Captive Hornbills. Front. Vet. Sci. 2020, 7, 236. [Google Scholar] [CrossRef]
- Regaiolli, B.; Sandri, C.; Rose, P.E.; Vallarin, V.; Spiezio, C. Investigating parental care behaviour in same-sex pairing of zoo greater flamingo (Phoenicopterus roseus). PeerJ 2018, 6, e5227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.S.; Rai, U.; Chand, P.; Baskaran, N. Summer Activity and Feeding pattern of Captive Red panda (Ailurus fulgens fulgens) at Padmaja Naidu Himalayan Zoological Park, Darjeeling, India. Lab-2-Land Mag. 2019, 2, 8–14. [Google Scholar]
- Wei, F.; Feng, Z.; Wang, Z.; Hu, J. Habitat use and separation between the giant panda and the red panda. J. Mammal. 2000, 81, 448–455. [Google Scholar] [CrossRef]
- Wei, F.; Feng, Z.; Wang, Z.; Zhou, A.; Hu, J. Nutrient and energy requirements of red panda (Ailurus fulgens) during lactation. Mammalogy 1999, 63, 3–10. [Google Scholar] [CrossRef]
- Lawlor, T.E.; Kowalski, K. Mammals, an outline of theriology. (English translation of original 1971 Polish edition by M. Bibrich for the Smithsonian In-stitute and National Science Foundation). J. Mammal. 1979, 60, 240. [Google Scholar] [CrossRef]
- Spiezio, C.; Manicardi, S.; Vallisneri, M.; Regaiolli, B. Don’t leave me alone! Housing solitary species in pairs: The case of the lowland tapir (Tapirus terrestris). Int. Zoo News 2016, 63, 244–254. [Google Scholar]
- Clauss, M.; Müller, D.; Steinmetz, H.; Hatt, J.M. The more the merrier or happy when alone? Hypothesis on stress susceptibility in captive individuals of solitary species. Second Int. Conf. Dis. Zoo Wild Anim. 2010, 12, 92–95. [Google Scholar]
- Macri, A.M.; Patterson-Kane, E. Behavioural analysis of solitary versus socially housed snow leopards (Panthera uncia), with the provision of simulated social contact. Appl. Anim. Behav. Sci. 2011, 130, 115–123. [Google Scholar] [CrossRef]
- Pitsko, L.E. Wild Tigers in Captivity: A Study of the Effects of the Captive Environment on Tiger Behavior. Master’s Thesis, Virginia Polytechnic Institute and State University, Blacksburg, VA, USA, 2003. [Google Scholar]
- Swaisgood, R.R.; Shepherdson, D.J. Scientific approaches to enrichment and stereotypies in zoo animals: What’s been done and where should we go next? Zoo Biol. 2005, 24, 499–518. [Google Scholar] [CrossRef]
- Mason, R.T.; LeMaster, M.P.; Müller-Schwarze, D. Chemical Signals in Vertebrates; Springer Press: New York, NY, USA, 2005; Volume 10. [Google Scholar]
- Brereton, J.E.; Fernandez, E.J. Which index should I use? A comparison of indices for enclosure use studies. Anim. Behav. Cogn. 2022, 9, 119–132. [Google Scholar] [CrossRef]
- Fernandez, E.J. Training as enrichment: A critical review. Anim. Welf. 2022, 31, 1–12. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Behavioural Class | Behavioural Category | Definition |
|---|---|---|
| Exploratory/territorial b. (Expl/terr) | Digging | Digging in ground. |
| Human-directed b. | Observing visitors, zookeepers or other humans, following them with attention. | |
| Licking | Olfactory investigation by licking any object (e.g., branches, leaves, enclosure furnishing) or substrate in the enclosure. | |
| Individual-play/Manipulation | Tactile investigation of objects in the enclosure, biting or chewing objects, interaction with environmental enrichment devices, carrying objects while moving or rolling on the back. | |
| Interspecific b. | Watching non-conspecifics, following them with attention (e.g., muntjac, bird). | |
| Scent-marking | Rubbing genitals on ground, objects, or enclosure furnishing. | |
| Sniffing | Sniffing any object (e.g., branches, leaves, enclosure furnishing) or substrate in the enclosure. | |
| Locomotive b. | Arboreal locomotion | Climbing on trees, walking, or running on tree branches or in the canopy. |
| Ground locomotion | Walking or running on the ground. | |
| Standing | Standing on back two paws. | |
| Routine b. | Hunt/stalk | Hunting, harassing, following a non-conspecific (e.g., birds). |
| Individual resting | Lying sleeping, curled in ball, or flat out. | |
| Comfort | Self-cleaning of the fur, scratching, and stretching. | |
| Vigilance | Being watchful, alert, while observing the surroundings. Lying, sitting, or standing with head up and eyes open, head or ears moving. | |
| Consumption b. | Eating | Eating food from the bowls (fruit and vegetables) or eating bamboo from the bamboo feeding point. |
| Foraging | Looking for food in the enclosure, searching in the grass, digging on the ground, or eating when browsing. | |
| Maintenance | Drinking at the water pool of the enclosure; urinating or defecating. | |
| Social b. | Aggression | Hitting a conspecific with paws, biting a conspecific. Initiating or receiving an aggression. |
| Chasing/display | Agonistic behaviours without physical contact: chase, threat (arching the back and the tail, moving the head up and down) or displacing a conspecific. Receiving an agonistic behaviour without physical contact. | |
| Grooming | Cleaning the fur by licking a conspecific. Grooming could be mutual, received, or done actively. | |
| Observing conspecific | Watching a conspecific or being watched by a conspecific. | |
| Sexual behaviour | Courtship (the female moves, marking heavily while the male follows her, marking over her marks) and mating behaviour. | |
| Sniffing conspecific | Sniffing a conspecific or being sniffed by a conspecific. | |
| Social play | Playing with another individual by lunging, wrestling, biting softly. | |
| Social resting | Resting in contact with a conspecific. | |
| Out of sight | Not observed | The individual is not visible. |
| Abnormal b. | Abnormal b. | Behaviours such as purposeless locomotion, repetitive route in the enclosure, excessive mouth movements (e.g., tongue flicking). |
| Behavioural Category | Definition |
|---|---|
| Antagonistic b. | The mother displaces, hits, or bites the cub. |
| Den | The mother and the cubs are not visible, hiding in the artificial nest. |
| Grooming cubs | Grooming of the cubs. |
| Nest building | Building a nest with twigs and grass. |
| Observing cubs | The mother observes and monitor the cubs. |
| Play with cubs | The mother plays with the cubs by lunging, wrestling, biting softly. |
| Rest and sleep with cubs | Lying, sleeping with the cubs. |
| Transport | The mother carries the cubs in the mouth while moving in the enclosure. |
| Consumption (CO) | Exploratory/Territorial (EXPL/TERR) | Locomotive (LOC) | Routine b. (R) | Social (S) | Maternal b. (MAT) | |
|---|---|---|---|---|---|---|
| Eating | Digging | Arboreal locomotion | Hunting/stalking | Aggression | Antagonistic b. | |
| Foraging | Licking | Ground locomotion | Resting | Chasing/displac-ing | Den | |
| Maintenance | Individual play/manipulation/carrying | Standing bipedal | Comfort | Grooming | Grooming cubs | |
| Scent-marking | Vigilance | Observe consp | Nest building | |||
| Sniffing | Sexual b. | Observe cubs | ||||
| Interspecific b. | Sniffing consp | Play with cubs | ||||
| Social play | Rest and sleep with cubs | |||||
| Social resting | Transport | |||||
| Score | 0–3 | 0–6 | 0–3 | 0–4 | 0–8 | 0–8 |
| Ilosha | Ny’ma | Lin | Maituc | ||
|---|---|---|---|---|---|
| CONSUMPTION | Eating | 568.5 (1161.3) | 0 (411.3) | 0 (278.5) | 0 (7.5) |
| Foraging | 0 (86.5) | 0 (0) | 0 (0) | 0 (0) | |
| Maintenance | 0 (13) | 0 (16.3) | 0 (0) | 0 (0) | |
| EXPL/TERR | Human-dir behav. | 18 (44.3) | 49 (117.3) | 40.5 (103.8) | 32 (91) |
| Ind play/manip | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| Interspecific b. | 0 (0) | 0 (2.3) | 11.5 (37.3) | 0 (13) | |
| Scent-marking | 0 (8.3) | 1.5 (38.3) | 0 (11.5) | 15.5 (76.8) | |
| Sniff/dig/lick | 34.5 (232.5) | 0 (14.8) | 0 (43.8) | 0 (14) | |
| LOCOMOTIVE | Arboreal loc | 115 (216.5) | 320 (684.8) | 184.5 (330) | 278.5 (442.3) |
| Ground loc | 95 (109.5) | 81 (525.5) | 0 (13.8) | 0 (278.8) | |
| Standing | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| ROUTINE B. | Comfort | 21.5 (226.3) | 136.5 (490) | 359 (789.8) | 253 (717.8) |
| Resting ind | 811.5 (1783) | 1755 (1508.8) | 258.5 (1637.3) | 1222.5 (2071.5) | |
| Vigilance | 96.5 (270) | 191 (253) | 197.5 (292.3) | 242.5 (440.3) | |
| SOCIAL B. | Aggression | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Chase/displace | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| Grooming | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| Obs consp | 0 (2.3) | 19.5 (64.8) | 0 (33) | 7.5 (35) | |
| Sexual b. | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| Sniffing consp | 0 (0) | 0 (0) | 0 (0) | 0 (10.8) | |
| Social resting | 0 (0) | 0 (0) | 0 (1643.5) | 0 (89.3) | |
| OoS | Not observed | 73 (350.5) | 0 (19) | 130.5 (616.5) | 36 (631.5) |
| CO | EXPL/TERR | LOC | OOS | R | S | MAT | ||
|---|---|---|---|---|---|---|---|---|
| Ilosha | AM | 100.5 (705) | 1 (83.8) | 68.5 (255.8) | 0 (46.3) | 158 (1442.3) | 0 (10.5) | 24 (498.8) |
| PM | 176 (1053.8) | 31 (113) | 104 (232.8) | 45 (152.5) | 676.5 (1557) | 0 (10) | 0 (19.5) | |
| V = 96; p = 0.322 | V = 87; p = 0.948 | V = 84; p = 0.274 | V = 64.5; p = 0.570 | V = 88; p = 0.339 | V = 40; p = 0.937 | V = 102; p = 0.079 | ||
| Ny’ma | AM | 0 (195.3) | 36 (144.5) | 191.5 (1131) | 0 (0) | 1301.5 (1579.5) | 0 (21.5) | |
| PM | 0 (139.5) | 40.5 (85.8) | 75 (285) | 0 (0) | 1471.5 (819.5) | 5.5 (27) | ||
| V = 55; p = 0.776 | V = 126; p = 0.715 | V = 183.5; p = 0.064 | V = 13.5; p = 0.933 | V = 83; p = 0.158 | V = 66.5; p = 0.636 | |||
| Lin | AM | 0 (0) | 8.5 (106) | 9 (194.8) | 0 (351.8) | 524.5 (1433.3) | 28.5 (1652) | |
| PM | 0 (258.3) | 30 (146.8) | 22.5 (236) | 0 (192.8) | 767.5 (1232.8) | 0 (388.8) | ||
| V = 12; p = 0.062 | V = 88.5; p = 0.538 | V = 103; p = 0.940 | V = 38; p = 0.657 | V = 88; p = 0.211 | V = 80; p = 0.084 | |||
| Maituc | AM | 0 (0) | 13.5 (241) | 6 (581.8) | 27.5 (412) | 437.5 (1170) | 10.5 (226.8) | |
| PM | 0 (0) | 0 (41.8) | 0 (219.5) | 0 (0) | 1662 (1085.5) | 0 (43.3) | ||
| V = 10; p = 0.499 | V = 136; p = 0.028 * | V = 107; p = 0.044 * | V = 59; p = 0.117 | V = 30; p = 0.005 * | V= 92; p = 0.069 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spiezio, C.; Altamura, M.; Weerman, J.; Regaiolli, B. Behaviour of Zoo-Housed Red Pandas (Ailurus fulgens): A Case-Study Testing the Behavioural Variety Index. J. Zool. Bot. Gard. 2022, 3, 223-237. https://0-doi-org.brum.beds.ac.uk/10.3390/jzbg3020018
Spiezio C, Altamura M, Weerman J, Regaiolli B. Behaviour of Zoo-Housed Red Pandas (Ailurus fulgens): A Case-Study Testing the Behavioural Variety Index. Journal of Zoological and Botanical Gardens. 2022; 3(2):223-237. https://0-doi-org.brum.beds.ac.uk/10.3390/jzbg3020018
Chicago/Turabian StyleSpiezio, Caterina, Mariangela Altamura, Janno Weerman, and Barbara Regaiolli. 2022. "Behaviour of Zoo-Housed Red Pandas (Ailurus fulgens): A Case-Study Testing the Behavioural Variety Index" Journal of Zoological and Botanical Gardens 3, no. 2: 223-237. https://0-doi-org.brum.beds.ac.uk/10.3390/jzbg3020018