
European Ground Squirrels at the Edge: Current Distribution Status and Anticipated Impact of Climate on Europe’s Southernmost Population

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
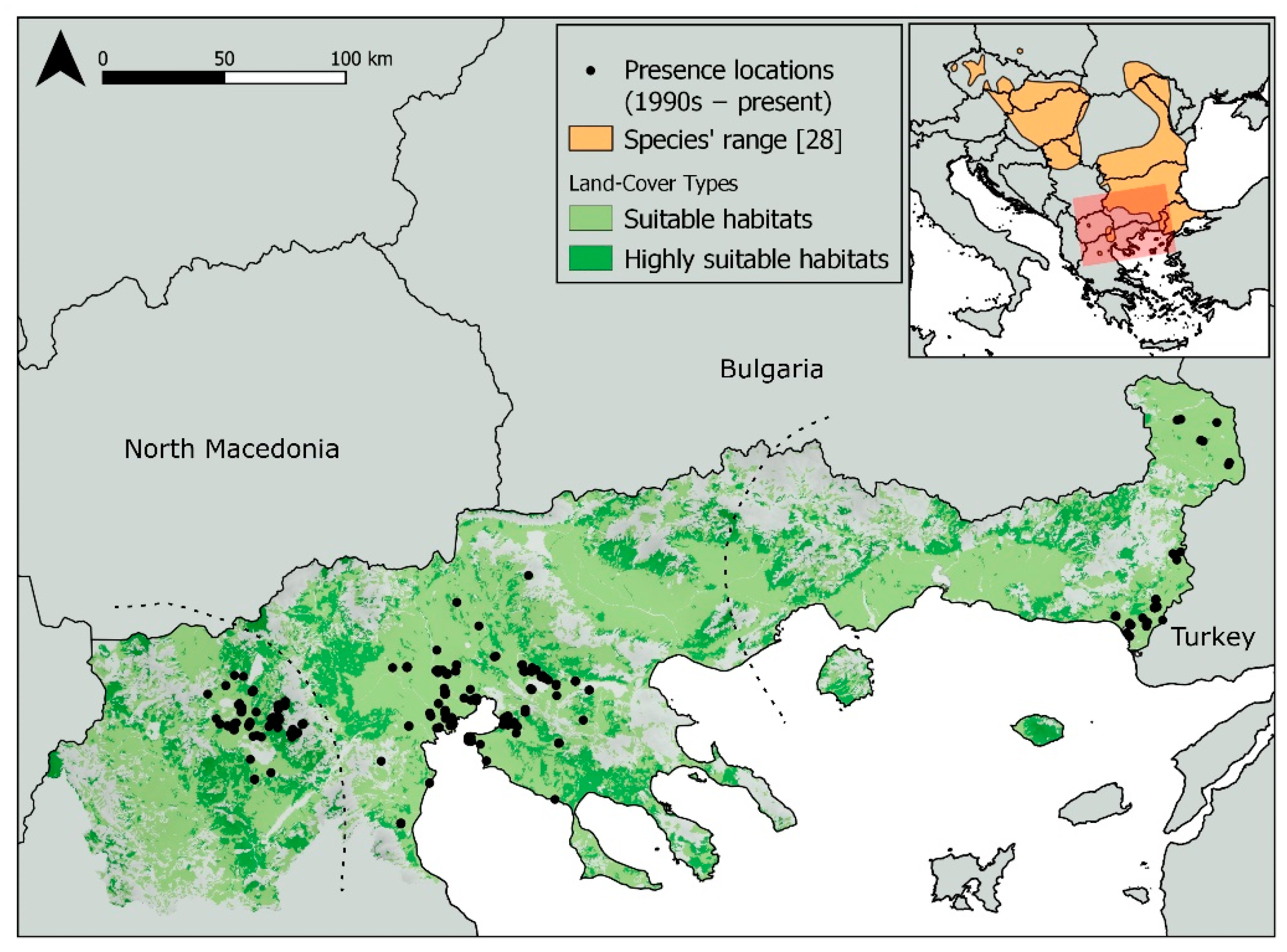
2.1. Study Area
2.2. Species Data
2.3. Ecological Niche Modeling Variables
2.4. Model Implementation and Processing
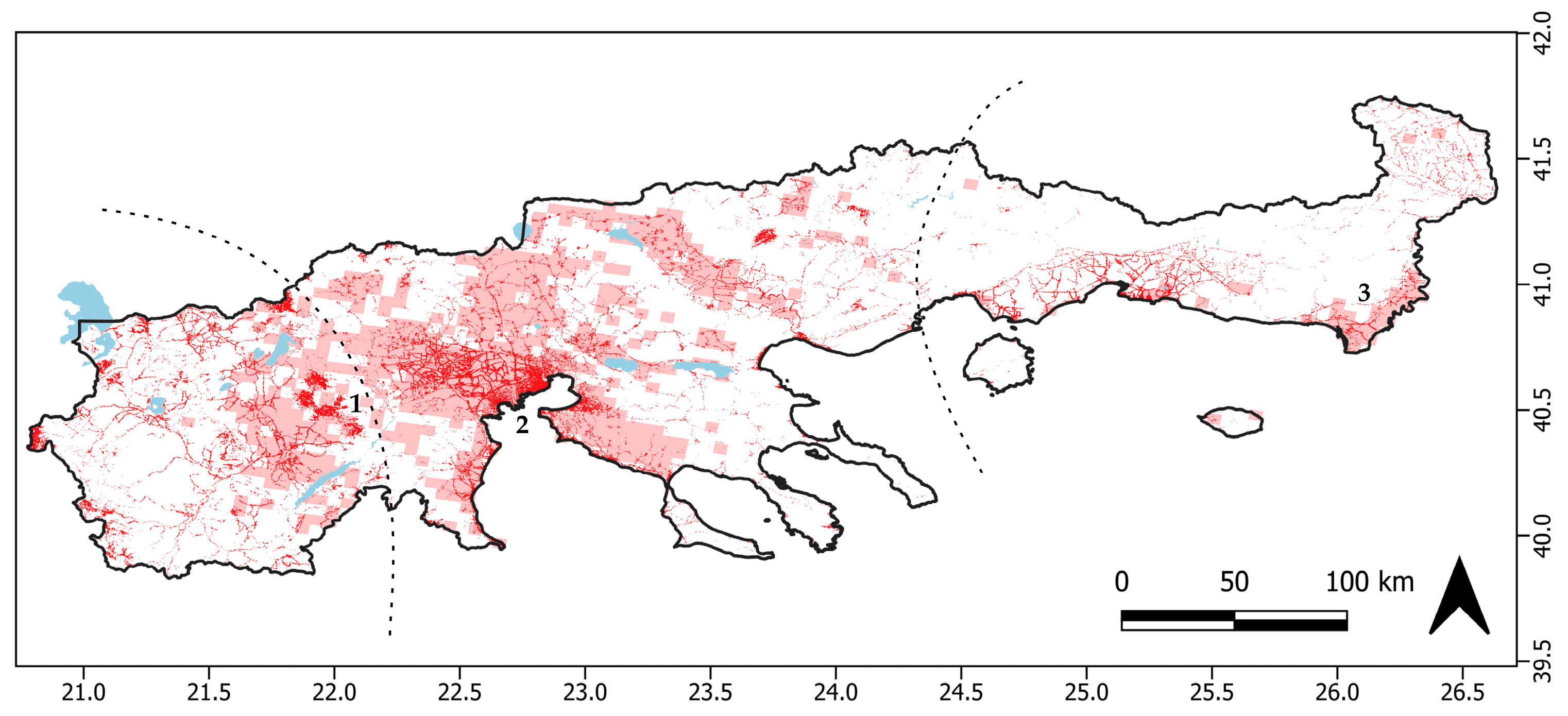
2.5. Drivers of Colony Extinction
3. Results
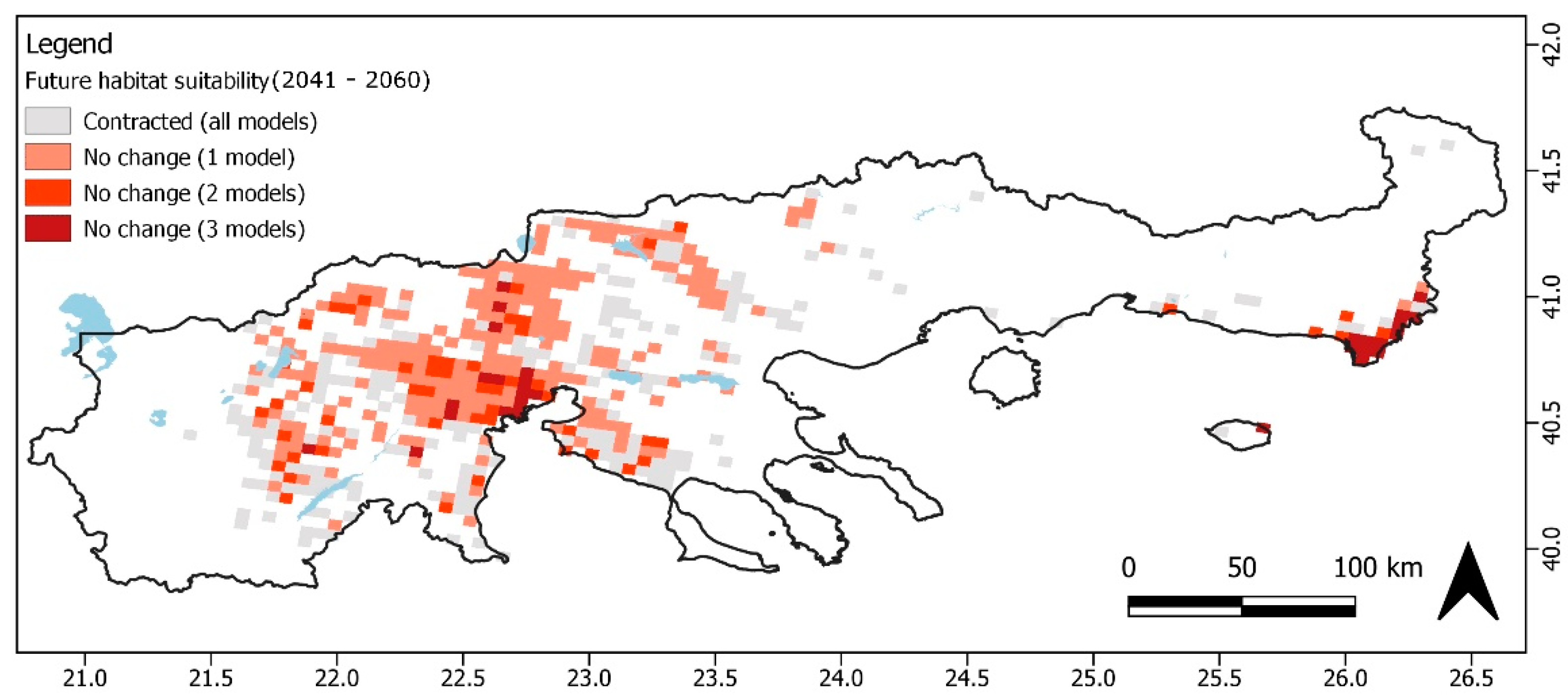
3.1. Ecological Niche Modeling
3.2. Drivers of Colony Extinction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antrop, M. Landscape change and the urbanization process in Europe. Landsc. Urban Plan. 2004, 67, 9–26. [Google Scholar] [CrossRef]
- Butchart, S.H.M.; Walpole, M.; Collen, B.; Van Strien, A.; Scharlemann, J.P.W.; Almond, R.E.A.; Baillie, J.E.M.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global biodiversity: Indicators of recent declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [Green Version]
- Geri, F.; Amici, V.; Rocchini, D. Human activity impact on the heterogeneity of a mediterranean landscape. Appl. Geogr. 2010, 30, 370–379. [Google Scholar] [CrossRef]
- Jongman, R.H.G. Homogenisation and fragmentation of the european landscape: Ecological consequences and solutions. Landsc. Urban Plan. 2002, 58, 211–221. [Google Scholar] [CrossRef]
- Leu, M.; Hanser, S.E.; Knick, S.T. The human footprint in the West: A large-scale analysus of anthropogenic impacts. Ecol. Appl. 2008, 18, 1119–1139. [Google Scholar] [CrossRef]
- Hadjigeorgiou, I. Past, present and future of pastoralism in Greece. Pastor. Res. Policy Pract. 2011, 1, 24. [Google Scholar] [CrossRef] [Green Version]
- Malandra, F.; Vitali, A.; Urbinati, C.; Garbarino, M. 70 Years of land use/land cover changes in the apennines (Italy): A meta-analysis. Forests 2018, 9, 551. [Google Scholar] [CrossRef] [Green Version]
- Sidiropoulou, A.; Karatassiou, M.; Galidaki, G.; Sklavou, P. Landscape pattern changes in response to transhumance abandonment on mountain vermio (North Greece). Sustainability 2015, 7, 15652–15673. [Google Scholar] [CrossRef] [Green Version]
- Farina, A. Recent changes of the mosaic patterns in a montane landscape (North Italy) and consequences on vertebrate fauna. Options Méditerr. Série Sémin. 1991, 15, 121–134. [Google Scholar]
- Francis, R.A.; Chadwick, M.A. What makes a species synurbic? Appl. Geogr. 2012, 32, 514–521. [Google Scholar] [CrossRef]
- Robledano, F.; Esteve, M.A.; Farinós, P.; Carreño, M.F.; Martínez-Fernández, J. Terrestrial birds as indicators of agricultural-induced changes and associated loss in conservation value of mediterranean wetlands. Ecol. Indic. 2010, 10, 274–286. [Google Scholar] [CrossRef]
- Russo, D.; Ancillotto, L. Sensitivity of bats to urbanization: A Review. Mamm. Biol. 2015, 80, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.; Lindenmayer, D.B. Landscape modification and habitat fragmentation: A synthesis. Glob. Ecol. Biogeogr. 2007, 16, 265–280. [Google Scholar] [CrossRef]
- Schloss, C.A.; Nuñez, T.A.; Lawler, J.J. Dispersal will limit ability of mammals to track climate change in the western hemisphere. Proc. Natl. Acad. Sci. USA 2012, 109, 8606–8611. [Google Scholar] [CrossRef] [Green Version]
- Broennimann, O.; Thuiller, W.; Hughes, G.; Midgley, G.F.; Alkemade, J.M.R.; Guisan, A. Do geographic distribution, niche property and life form explain plants’ vulnerability to global change? Glob. Chang. Biol. 2006, 12, 1079–1093. [Google Scholar] [CrossRef]
- Coroiu, C.; Kryštufek, B.; Vohralík, V.; Zagorodnyuk, I. Spermophilus Citellus. In IUCN 2012, IUCN Red List of Threatened Species; Version 2012.1; IUCN: Gland, Switzerland, 2008; Available online: https://www.iucnredlist.org/species/20472/9204055 (accessed on 10 January 2022).
- Hoffmann, I.E.; Millesi, E.; Pieta, K.; Dittami, J.P. anthropogenic effects on the population ecology of european ground squirrels (Spermophilus citellus) at the periphery of their geographic range. Mamm. Biol. 2003, 68, 205–213. [Google Scholar] [CrossRef]
- Janák, M.; Marhoul, P.; Matějů, J. Action Plan for the Conservation of the European Ground Squirrel Spermophilus Citellus in the European Union; European Commission: Brussels, Belgium, 2013; pp. 5–8. [Google Scholar]
- Kryštufek, B.; Vohralík, V. Mammals of Turkey and Cyprus. Rodentia I: Sciuridae, Dipodidae, Gliridae, Arvicolinae; Zgodovinsko društvo za južno Primorsko: Ljubljana, Slovenia, 2005; pp. 42–43. [Google Scholar]
- Kryštufek, B.; Glasnović, P.; Petkovski, S. The status of a rare phylogeographic lineage of the vulnerable european souslik spermophilus citellus, endemic to central macedonia. Oryx 2012, 46, 442–445. [Google Scholar] [CrossRef] [Green Version]
- Grulich, I. Sysel obecný citellus citellus, L. v ČSSR. Práce Brněn. Základny ČSAV 1960, 32, 473–563. [Google Scholar]
- Youlatos, D.; Boutsis, Y.; Pantis, J.D.; Hadjicharalambous, H. Activity patterns of european ground squirrels (Spermophilus citellus) in a cultivated field in Northern Greece. Mammalia 2007, 71, 183–186. [Google Scholar] [CrossRef]
- Davidson, A.D.; Lightfoot, D.C. Interactive effects of keystone rodents on the structure of desert grassland arthropod communities. Ecography 2007, 30, 515–525. [Google Scholar] [CrossRef]
- Lindtner, P.; Gömöryová, E.; Gömöry, D.; Stašiov, S.; Kubovčík, V. Development of physico-chemical and biological soil properties on the european ground squirrel mounds. Geoderma 2019, 339, 85–93. [Google Scholar] [CrossRef]
- Lindtner, P.; Svitok, M.; Ujházy, K.; Kubovčík, V. Disturbances by the European ground squirrel enhance diversity and spatial heterogeneity of plant communities in temperate grassland. Biodivers. Conserv. 2020, 29, 853–867. [Google Scholar] [CrossRef]
- Ramos-Lara, N.; Koprowski, J.L.; Kryštufek, B.; Hoffmann, I.E. Spermophilus citellus (Rodentia: Sciuridae). Mamm. Species 2014, 46, 71–87. [Google Scholar] [CrossRef] [Green Version]
- Hegyeli, Z. Spermophilus Citellusi, IUCN Red List Threatment Species. 2020. Available online: https://www.iucnredlist.org/species/20472/91282380 (accessed on 10 January 2022).
- Youlatos, D. Spermophilus Citellus (Linnaeus, 1766). In The Red Book of Endangered Animals of Greece; Legakis, A., Maragou, P., Eds.; Hellenic Zoological Society: Athens, Greece, 2009; p. 528. [Google Scholar]
- Rammou, D.-L.; Kavroudakis, D.; Youlatos, D. Distribution, population size, and habitat characteristics of the endangered european ground squirrel (Spermophilus citellus, Rodentia, Mammalia) in its southernmost range. Sustainability 2021, 13, 8411. [Google Scholar] [CrossRef]
- Giannakopoulos, C.; Le Sager, P.; Bindi, M.; Moriondo, M.; Kostopoulou, E.; Goodess, C.M. Climatic changes and associated impacts in the mediterranean resulting from a 2 °C global warming. Glob. Planet. Chang. 2009, 68, 209–224. [Google Scholar] [CrossRef]
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Goldberg, A.R.; Conway, C.J. Hibernation behavior of a federally threatened ground squirrel: Climate change and habitat selection implications. J. Mammal. 2021, 102, 574–587. [Google Scholar] [CrossRef]
- Németh, I.; Nyitrai, V.; Altbäcker, V. Ambient temperature and annual timing affect torpor bouts and euthermic phases of hibernating european ground squirrels (Spermophilus citellus). Can. J. Zool. 2009, 87, 204–210. [Google Scholar] [CrossRef]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef]
- Austin, M.P. Spatial prediction of species distribution: An interface between ecological theory and statistical modelling. Ecol. Model. 2002, 157, 101–118. [Google Scholar] [CrossRef] [Green Version]
- Cianfrani, C.; Broennimann, O.; Loy, A.; Guisan, A. More than range exposure: Global otter vulnerability to climate change. Biol. Conserv. 2018, 221, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. A maximum entropy approach to species distribution modeling. In Proceedings of the Twenty-First International Conference On Machine Learning, Banff, AB, Canada, 4−8 July 2004; pp. 655–662. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Demirtaş, S. Estimation of the climate preference between two lineages of europe-an ground squirrel using maximum entropy modeling. J. Adv. Res. Nat. Appl. Sci. 2020, 6, 328–341. [Google Scholar] [CrossRef]
- Gür, H. The future impact of climate and land-use changes on anatolian ground squirrels under different scenarios. bioRxiv 2021, 460244. [Google Scholar] [CrossRef]
- Dilts, T.E.; Weisberg, P.J.; Leitner, P.; Matocq, M.D.; Inman, R.D.; Nussear, K.E.; Esque, T.C. Multiscale connectivity and graph theory highlight critical areas for conservation under climate change. Ecol. Appl. 2016, 26, 1223–1237. [Google Scholar] [CrossRef]
- Holt, A.C.; Salkeld, D.J.; Fritz, C.L.; Tucker, J.R.; Gong, P. Spatial analysis of plague in California: Niche modeling predictions of the current distribution and potential response to climate change. Int. J. Health Geogr. 2009, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Inman, R.D.; Esque, T.C.; Nussear, K.E.; Leitner, P.; Matocq, M.D.; Weisberg, P.J.; Dilts, T.E.; Vandergast, A.G. Is there room for all of us? Renewable energy and Xerospermophilus Mohavensis. Endanger. Species Res. 2013, 20, 1–18. [Google Scholar] [CrossRef]
- Kryštufek, B.; Stanciu, C.; Ivajnšič, D.; Cherkaoui, S.I.; Janžekovič, F. Facts and misconceptions on the palaearctic existence of the striped ground squirrel. Mammalia 2018, 82, 248–255. [Google Scholar] [CrossRef]
- Tian, L. Relationship between environmental factors and the spatial distribution of Spermophilus dauricus during 2000–2015 in China. Int. J. Biometeorol. 2018, 62, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Tzvetkov, J.; Koshev, Y. GIS habitat model of potential distribution of european ground squirrel (Spermophilus citellus) in Bulgaria. In Proceedings of the 6th European Ground Squirrel Meeting, Belgrade, Serbia, 4−6 November 2016. [Google Scholar]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Pearson, R.G.; Thuiller, W.; Araújo, M.B.; Martinez-Meyer, E.; Brotons, L.; McClean, C.; Miles, L.; Segurado, P.; Dawson, T.P.; Lees, D.C. Model-based uncertainty in species range prediction. J. Biogeogr. 2006, 33, 1704–1711. [Google Scholar] [CrossRef]
- Rebelo, H.; Jones, G. Ground validation of presence-only modelling with rare species: A case study on Barbastelles Barbastella Barbastellus (Chiroptera: Vespertilionidae). J. Appl. Ecol. 2010, 47, 410–420. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A.; NCEAS Predicting Species Distributions Working Group. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future köppen-geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [Green Version]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Feng, X.; Park, D.S.; Liang, Y.; Pandey, R.; Papeş, M. Collinearity in ecological niche modeling: Confusions and challenges. Ecol. Evol. 2019, 9, 10365–10376. [Google Scholar] [CrossRef] [Green Version]
- QGIS Development Team QGIS Geographic Information System. 2021. Available online: http://www.qgis.org (accessed on 10 January 2022).
- Riahi, K.; Van Vuuren, D.P.; Kriegler, E.; Edmonds, J.; O’Neill, B.C.; Fujimori, S.; Bauer, N.; Calvin, K.; Dellink, R.; Fricko, O.; et al. The shared socioeconomic pathways and their energy, land use, and greenhouse gas emissions implications: An overview. Glob. Environ. Chang. 2017, 42, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Kaky, E.; Nolan, V.; Alatawi, A.; Gilbert, F. A comparison between ensemble and MaxEnt species distribution modelling approaches for conservation: A case study with egyptian medicinal plants. Ecol. Inform. 2020, 60, 101150. [Google Scholar] [CrossRef]
- Morales, N.S.; Fernández, I.C.; Baca-González, V. MaxEnt’s parameter configuration and small samples: Are we paying attention to recommendations? A systematic review. PeerJ 2017, 5, e3093. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for maxent ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference. In A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Boyce, M.S.; Vernier, P.R.; Nielsen, S.E.; Schmiegelow, F.K.A. Evaluating resource selection functions. Ecol. Model. 2002, 157, 281–300. [Google Scholar] [CrossRef] [Green Version]
- Hirzel, A.H.; Le Lay, G.; Helfer, V.; Randin, C.; Guisan, A. Evaluating the ability of habitat suitability models to predict species presences. Predict. Species Distrib. 2006, 199, 142–152. [Google Scholar] [CrossRef]
- Araújo, M.B.; Pearson, R.G.; Thuiller, W.; Erhard, M. Validation of species–climate impact models under climate change. Glob. Chang. Biol. 2005, 11, 1504–1513. [Google Scholar] [CrossRef] [Green Version]
- Lobo, J.M.; Jiménez-Valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Sun, X.; Long, Z.; Jia, J. A multi-scale maxent approach to model habitat suitability for the giant pandas in the Qionglai mountain, China. Glob. Ecol. Conserv. 2021, 30, e01766. [Google Scholar] [CrossRef]
- Ashcroft, M.B. Identifying refugia from climate change. J. Biogeogr. 2010, 37, 1407–1413. [Google Scholar] [CrossRef]
- Armitage, K.B. Sociality as a life-history tactic of ground squirrels. Oecologia 1981, 48, 36–49. [Google Scholar] [CrossRef]
- Koshev, Y.S. Distribution and status of the european ground squirrel (Spermophilus citellus) in Bulgaria. Lynx 2008, 39, 251–261. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foudation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Matějů, J.; Hulová, Š.; Nová, P.; Cepáková, E.; Marhoul, P.; Uhlíková, J. Action Plan for the European Ground Squirrel (Spermophilus citellus) in the Czech Republic; Charles University and Agency for Nature and Landscape Protection of the Czech Republic: Prague, Czech Republic, 2010; p. 80. [Google Scholar]
- Kati, V.; Kassara, C.; Psaralexi, M.; Tzortzakaki, O.; Petridou, M.; Galani, A.; Hoffmann, M.T. Conservation policy under a roadless perspective: Minimizing fragmentation in Greece. Biol. Conserv. 2020, 252, 108828. [Google Scholar] [CrossRef]
- Brown, J.H.; Kodricbrown, A. Turnover rates in insular biogeography—Effect of immigration on extinction. Ecology 1977, 58, 445–449. [Google Scholar] [CrossRef] [Green Version]
- Pulliam, H.R. On the relationship between niche and distribution. Ecol. Lett. 2000, 3, 349–361. [Google Scholar] [CrossRef]
- Hubbell, S.P. The Unified Neutral Theory of Biodiversity and Biogeography; Princeton University Press: Princeton, NJ, USA, 2001; pp. 152–200. [Google Scholar]
- Říčanová, Š.; Bryja, J.; Cosson, J.-F.; Gedeon, C.; Choleva, L.; Ambros, M.; Sedláček, F. Depleted genetic variation of the european ground squirrel in Central Europe in both microsatellites and the major histocompatibility complex gene: Implications for conservation. Conserv. Genet. 2011, 12, 1115–1129. [Google Scholar] [CrossRef]
- Davidson, A.D.; Ponce, E.; Lightfoot, D.C.; Fredrickson, E.L.; Brown, J.H.; Cruzado, J.; Brantley, S.L.; Sierra-Corona, R.; List, R.; Toledo, D.; et al. Rapid response of a grassland ecosystem to an experimental manipulation of a keystone rodent and domestic livestock. Ecology 2010, 91, 3189–3200. [Google Scholar] [CrossRef]
- Luoto, M.; Pykälä, J.; Kuussaari, M. Decline of landscape-scale habitat and species diversity after the end of cattle grazing. J. Nat. Conserv. 2003, 11, 171–178. [Google Scholar] [CrossRef]
- Tzanopoulos, J.; Mitchley, J.; Pantis, J.D. Vegetation dynamics in abandoned crop fields on a Mediterranean island: Development of succession model and estimation of disturbance thresholds. Agric. Ecosyst. Environ. 2007, 120, 370–376. [Google Scholar] [CrossRef]
- Zomeni, M.; Tzanopoulos, J.; Pantis, J.D. Historical analysis of landscape change using remote sensing techniques: An explanatory tool for agricultural transformation in Greek rural areas. Landsc. Urban Plan. 2008, 86, 38–46. [Google Scholar] [CrossRef]
- Zakkak, S.; Kakalis, E.; Radović, A.; Halley, J.M.; Kati, V. The impact of forest encroachment after agricultural land abandonment on passerine bird communities: The case of Greece. J. Nat. Conserv. 2014, 22, 157–165. [Google Scholar] [CrossRef]
- Zakkak, S.; Halley, J.M.; Akriotis, T.; Kati, V. Lizards along an agricultural land abandonment gradient in pindos mountains, Greece. Amphib. Reptil. 2015, 36, 253–264. [Google Scholar] [CrossRef]
- Blüthgen, N.; Dormann, C.F.; Prati, D.; Klaus, V.H.; Kleinebecker, T.; Hölzel, N.; Alt, F.; Boch, S.; Gockel, S.; Hemp, A.; et al. A quantitative index of land-use intensity in grasslands: Integrating mowing, grazing and fertilization. Basic Appl. Ecol. 2012, 13, 207–220. [Google Scholar] [CrossRef]
- Petluš, P.; Petlušová, V.; Baláž, I.; Ševčík, M.; Lešová, A.; Hapl, E. Impact of management measures on the european ground squirrel population development. Folia Oecol. 2021, 48, 169–179. [Google Scholar] [CrossRef]
- European Environment Agency. High Nature Value Farmland: Characteristics, Trends, and Policy Challenges; Institute European Environmental Policy: Copenhagen, Denmark, 2004. [Google Scholar]
- Herzon, I.; Birge, T.; Allen, B.; Povellato, A.; Vanni, F.; Hart, K.; Radley, G.; Tucker, G.; Keenleyside, C.; Oppermann, R.; et al. Time to look for evidence: Results-based approach to biodiversity conservation on farmland in Europe. Land Use Policy 2018, 71, 347–354. [Google Scholar] [CrossRef]
- Kis, J.; Váczi, O.; Katona, K.; Altbäcker, V. A növényzet magasságának hatása a cinegési ürgék élőhelyválasztására. The effect of vegetation height to habitat selection of ground squirrels in cinegés. Termvéd. Közlemények 1998, 7, 117–123. [Google Scholar]
- Matějů, J.; Nová, P.; Uhlíkovà, J.; Hulová, Š.; Cepáková, E. Distribution of the European ground squirrel (Spermophilus citellus) in the Czech Republic in 2002–2008. Lynx 2008, 39, 277–294. [Google Scholar]
- Ricankova, V.; Fric, Z.; Chlachula, J.; Stastna, P.; Faltynkova, A.; Zemek, F. Habitat requirements of the long-tailed ground squirrel (Spermophilus undulatus) in the Southern Altai. J. Zool. 2006, 270, 1–8. [Google Scholar] [CrossRef]
- European Commission. Agri-Environment Measures—Overview on General Principles, Types of Measures, and Application; European Commission: Brussels, Belgium, 2005. [Google Scholar]
- Kleijn, D.; Baquero, R.A.; Clough, Y.; Díaz, M.; De Esteban, J.; Fernández, F.; Gabriel, D.; Herzog, F.; Holzschuh, A.; Jöhl, R.; et al. Mixed biodiversity benefits of agri-environment schemes in five european countries. Ecol. Lett. 2006, 9, 243–254. [Google Scholar] [CrossRef]
- Walker, L.K.; Morris, A.J.; Cristinacce, A.; Dadam, D.; Grice, P.V.; Peach, W.J. Effects of higher-tier agri-environment scheme on the abundance of priority farmland birds. Anim. Conserv. 2018, 21, 183–192. [Google Scholar] [CrossRef]
- Sokos, C.K.; Mamolos, A.P.; Kalburtji, K.L.; Birtsas, P.K. Farming and wildlife in mediterranean agroecosystems. J. Nat. Conserv. 2013, 21, 81–92. [Google Scholar] [CrossRef]
- Bald, V.; Boetzl, F.A.; Krauss, J. Where do hamsters go after cereal harvest? A case study. Basic Appl. Ecol. 2021, 54, 98–107. [Google Scholar] [CrossRef]
- Tissier, M.L.; Kletty, F.; Robin, J.-P.; Habold, C. Sustainable agriculture: Nutritional benefits of wheat–soybean and maize–sunflower associations for hibernation and reproduction of endangered common hamsters. Sustainability 2021, 13, 1352. [Google Scholar] [CrossRef]
- Liordos, V.; Kontsiotis, V.J.; Anastasiadou, M.; Karavasias, E. Effects of attitudes and demography on public support for endangered species conservation. Sci. Total Environ. 2017, 595, 25–34. [Google Scholar] [CrossRef]
- Clevenger, A.P.; Chruszcz, B.; Gunson, K.E. Spatial patterns and factors influencing small vertebrate fauna road-kill aggregations. Biol. Conserv. 2002, 109, 15–26. [Google Scholar] [CrossRef]
- D’Amico, M.; Román, J.; De los Reyes, L.; Revilla, E. Vertebrate road-kill patterns in mediterranean habitats: Who, when and where. Biol. Conserv. 2015, 191, 234–242. [Google Scholar] [CrossRef]
- Kenyeres, Z.; Bauer, N.; Nagy, L.; Szabó, S. Enhancement of a declining european ground squirrel (Spermophilus citellus) population with habitat restoration. J. Nat. Conserv. 2018, 45, 98–106. [Google Scholar] [CrossRef]
- Krigas, N.; Tsiafouli, M.A.; Katsoulis, G.; Votsi, N.-E.; Van Kleunen, M. Investigating the invasion pattern of the alien plant solanum elaeagnifolium cav. (Silverleaf Nightshade): Environmental and human-induced drivers. Plants 2021, 10, 805. [Google Scholar] [CrossRef]
- Krigas, N.; Kokkini, S. A survey of the alien vascular flora of the urban and suburban area of thessaloniki, N Greece. Willdenowia 2004, 34, 81–99. [Google Scholar] [CrossRef]
- Kajzer-Bonk, J.; Szpiłyk, D.; Woyciechowski, M. Invasive goldenrods affect abundance and diversity of grassland ant communities (Hymenoptera: Formicidae). J. Insect Conserv. 2016, 20, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Dueñas, M.-A.; Hemming, D.J.; Roberts, A.; Diaz-Soltero, H. The threat of invasive species to IUCN-listed critically endangered species: A systematic review. Glob. Ecol. Conserv. 2021, 26, e01476. [Google Scholar] [CrossRef]
- Inman, R.D.; Esque, T.C.; Nussear, K.E.; Leitner, P.; Matocq, M.D.; Weisberg, P.J.; Dilts, T.E. Impacts of climate change and renewable energy development on habitat of an endemic squirrel, Xerospermophilus mohavensis, in the Mojave Desert, USA. Biol. Conserv. 2016, 200, 112–121. [Google Scholar] [CrossRef]
- Łopucki, R.; Łopucki, R.; Klich, D.; Ścibior, A.; Gołębiowska, D.; Perzanowski, K. Living in habitats affected by wind turbines may result in an increase in corticosterone levels in ground dwelling animals. Ecol. Indic. 2018, 84, 165. [Google Scholar] [CrossRef]
- Lovich, J.E.; Ennen, J.R. Assessing the state of knowledge of utility-scale wind energy development and operation on non-volant terrestrial and marine wildlife. Appl. Eng. 2013, 103, 52–60. [Google Scholar] [CrossRef]
- Kati, V.; Kassara, C.; Vrontisi, Z.; Moustakas, A. The biodiversity-wind energy-land use nexus in a global biodiversity hotspot. Sci. Total Environ. 2021, 768, 144474. [Google Scholar] [CrossRef]
- Koshev, Y.; Kachamakova, M.; Arangelov, S.; Ragyov, D. Translocations of european ground squirrel (Spermophilus citellus) along altitudinal gradient in Bulgaria—An Overview. Nat. Conserv. 2019, 35, 63–95. [Google Scholar] [CrossRef] [Green Version]
- Balaz, I.; Jancova, A.; Ambros, M. Restitution of the european ground squirrel (Spermophilus citellus) in Slovakia. Lynx 2008, 39, 235–240. [Google Scholar]
- Gedeon, C.I.; Váczi, O.; Koósz, B.; Altbäcker, V. Morning release into artificial burrows with retention caps facilitates success of European ground squirrel (Spermophilus citellus) translocations. Eur. J. Wildl. Res. 2011, 57, 1101–1105. [Google Scholar] [CrossRef]
- Kachamakova, M.; Koshev, Y. Post-release settlement and survival of endangered European ground squirrel after conservation reinforcement. J. Nat. Conserv. 2021, 63, 126048. [Google Scholar] [CrossRef]
- Löbbová, D.; Hapl, E. Conservation of European ground squirrel (Mammalia: Rodentia) in Slovakia: Results of current reintroduction programme. Slovak Raptor J. 2014, 8, 105. [Google Scholar] [CrossRef] [Green Version]
- Matějů, J.; Říčanová, Š.; Poláková, S.; Ambros, M.; Kala, B.; Matějů, K.; Kratochvíl, L. Method of releasing and number of animals are determinants for the success of European ground squirrel (Spermophilus citellus) reintroductions. Eur. J. Wildl. Res. 2012, 58, 473–482. [Google Scholar] [CrossRef]
- Bottrill, M.C.; Joseph, L.N.; Carwardine, J.; Bode, M.; Cook, C.; Game, E.T.; Grantham, H.; Kark, S.; Linke, S.; McDonald-Madden, E.; et al. Is conservation triage just smart decision making? Trends Ecol. Evol. 2008, 23, 649–654. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Model | Category | Variable | Type | Source | Initial Resolution |
|---|---|---|---|---|---|
| 1 * | Environmental—Abiotic | Elevation | Continuous | https://land.copernicus.eu (EU-DEM v1.1) | 25 m |
| 1 *,2 * | Environmental—Abiotic | Slope | Continuous | Developed using the EU-DEM v1.1 layer and the QGIS Slope function | 25 m |
| 1 * | Environmental—Abiotic | Aspect | Continuous | Developed using the EU-DEM v1.1 layer and the QGIS Aspect function | 25 m |
| 1 * | Environmental—Abiotic | Tree cover density | Continuous | https://land.copernicus.eu (Tree Cover Density 2018) | 10 m |
| 1 * | Environmental—Abiotic | EGS suitable land-cover | Categorical | https://land.copernicus.eu (Corine Land Cover 2018) (Reclassified/suitable: 2–4, 9, 12–14, 19–22, 35, 37, 38; highly suitable: 6, 10, 11, 15–18, 26, 28, 32) | 100 m |
| 1 * | Anthropogenic—Abiotic | Soil imperviousness (soil sealing) | Continuous | https://land.copernicus.eu | 10 m |
| 1 * | Anthropogenic—Abiotic | Road density | Continuous | https://geodata.gov.gr; converted tarred roads to 50 m interval point layer, and applied heat map/kernel density estimation function in QGIS (200 m radius/quartic kernel shape) | Vector |
| 2 | Environmental—Biotic | Normalized difference vegetation index (NDVI)—20-year mean (1999−2019) | Continuous | https://land.copernicus.eu | 1000 m |
| 2 | Bioclimatic—Abiotic | (Bio1) Annual mean temperature | Continuous | https://www.worldclim.org/ | 4500 m |
| 2 | Bioclimatic—Abiotic | (Bio2) Annual mean diurnal range | Continuous | https://www.worldclim.org/ | 4500 m |
| 2 * | Bioclimatic—Abiotic | (Bio4) Temperature seasonality SD | Continuous | https://www.worldclim.org/ | 4500 m |
| 2 | Bioclimatic—Abiotic | (Bio7) Annual temp range | Continuous | https://www.worldclim.org/ | 4500 m |
| 2 * | Bioclimatic—Abiotic | (Bio8) Mean temp of wettest quarter | Continuous | https://www.worldclim.org/ | 4500 m |
| 2 | Bioclimatic—Abiotic | (Bio9) Mean temp of driest quarter | Continuous | https://www.worldclim.org/ | 4500 m |
| 2 | Bioclimatic—Abiotic | (Bio10) Mean temp of warmest quarter | Continuous | https://www.worldclim.org/ | 4500 m |
| 2 | Bioclimatic—Abiotic | (Bio11) Mean temp of coldest quarter | Continuous | https://www.worldclim.org/ | 4500 m |
| 2 | Bioclimatic—Abiotic | (Bio12) Annual precipitation | Continuous | https://www.worldclim.org/ | 4500 m |
| 2 * | Bioclimatic—Abiotic | (Bio15) Precipitation seasonality (CV) | Continuous | https://www.worldclim.org/ | 4500 m |
| 2 | Bioclimatic—Abiotic | (Bio16) Precipitation of wettest quarter | Continuous | https://www.worldclim.org/ | 4500 m |
| 2 | Bioclimatic—Abiotic | (Bio17) Precipitation of driest quarter | Continuous | https://www.worldclim.org/ | 4500 m |
| 2 * | Bioclimatic—Abiotic | (Bio18) Precipitation of warmest quarter | Continuous | https://www.worldclim.org/ | 4500 m |
| 2 | Bioclimatic—Abiotic | (Bio19) Precipitation of coldest quarter | Continuous | https://www.worldclim.org/ | 4500 m |
| 2 | Anthropogenic—Biotic | Population density | Continuous | https://ec.europa.eu/eurostat | 1000 m |
| 2 * | Environmental—Abiotic | Soil bulk density | Continuous | https://esdac.jrc.ec.europa.eu (LUCAS Database) | 500 m |
| 2 * | Environmental—Abiotic | Soil texture (USDA classification) | Categorical | https://esdac.jrc.ec.europa.eu (LUCAS Database) | 500 m |
| Variables | Model 1: Habitat Availability | Model 2: Habitat Suitability | ||
|---|---|---|---|---|
| Pc (%) | Pi (%) | Pc (%) | Pi (%) | |
| Road density | 30.9 | 26.7 | − | − |
| Elevation | 29.1 | 25.7 | − | − |
| EGS suitable land-cover | 14 | 16 | − | − |
| Tree cover density | 12.7 | 9.5 | − | − |
| Slope | 8.8 | 18.4 | 52.5 | 29.8 |
| Aspect | 2.4 | 1.5 | − | − |
| Soil imperviousness | 2.1 | 2.3 | − | − |
| Precipitation seasonality (Bio15) | − | − | 22.4 | 34.7 |
| Soil texture | − | − | 11.1 | 6.1 |
| Soil bulk density | − | − | 7.2 | 3.6 |
| Precipitation of warmest quarter (Bio18) | − | − | 4.4 | 18.7 |
| Temperature seasonality SD (Bio4) | − | − | 1.5 | 3.3 |
| Mean temp of wettest quarter (Bio8) | − | − | 0.9 | 3.7 |
| Variables | Estimate | SE | z-Values | Pr (>│z│) |
|---|---|---|---|---|
| Intercept (β) | 1.555 | 0.337 | 4.613 | <0.0001 |
| Soil imperviousness | −0.0813 | 0.029 | −2.754 | <0.001 |
| High suitability habitats | −3.639 | 1.260 | −2.887 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rammou, D.-L.; Astaras, C.; Migli, D.; Boutsis, G.; Galanaki, A.; Kominos, T.; Youlatos, D. European Ground Squirrels at the Edge: Current Distribution Status and Anticipated Impact of Climate on Europe’s Southernmost Population. Land 2022, 11, 301. https://0-doi-org.brum.beds.ac.uk/10.3390/land11020301
Rammou D-L, Astaras C, Migli D, Boutsis G, Galanaki A, Kominos T, Youlatos D. European Ground Squirrels at the Edge: Current Distribution Status and Anticipated Impact of Climate on Europe’s Southernmost Population. Land. 2022; 11(2):301. https://0-doi-org.brum.beds.ac.uk/10.3390/land11020301
Chicago/Turabian StyleRammou, Dimitra-Lida, Christos Astaras, Despina Migli, George Boutsis, Antonia Galanaki, Theodoros Kominos, and Dionisios Youlatos. 2022. "European Ground Squirrels at the Edge: Current Distribution Status and Anticipated Impact of Climate on Europe’s Southernmost Population" Land 11, no. 2: 301. https://0-doi-org.brum.beds.ac.uk/10.3390/land11020301