
Soil Organic Carbon Isotope Tracing in Sorghum under Ambient CO2 and Free-Air CO2 Enrichment (FACE)

 ,
, 
Abstract
:1. Introduction
2. Methods
3. Results and Discussion
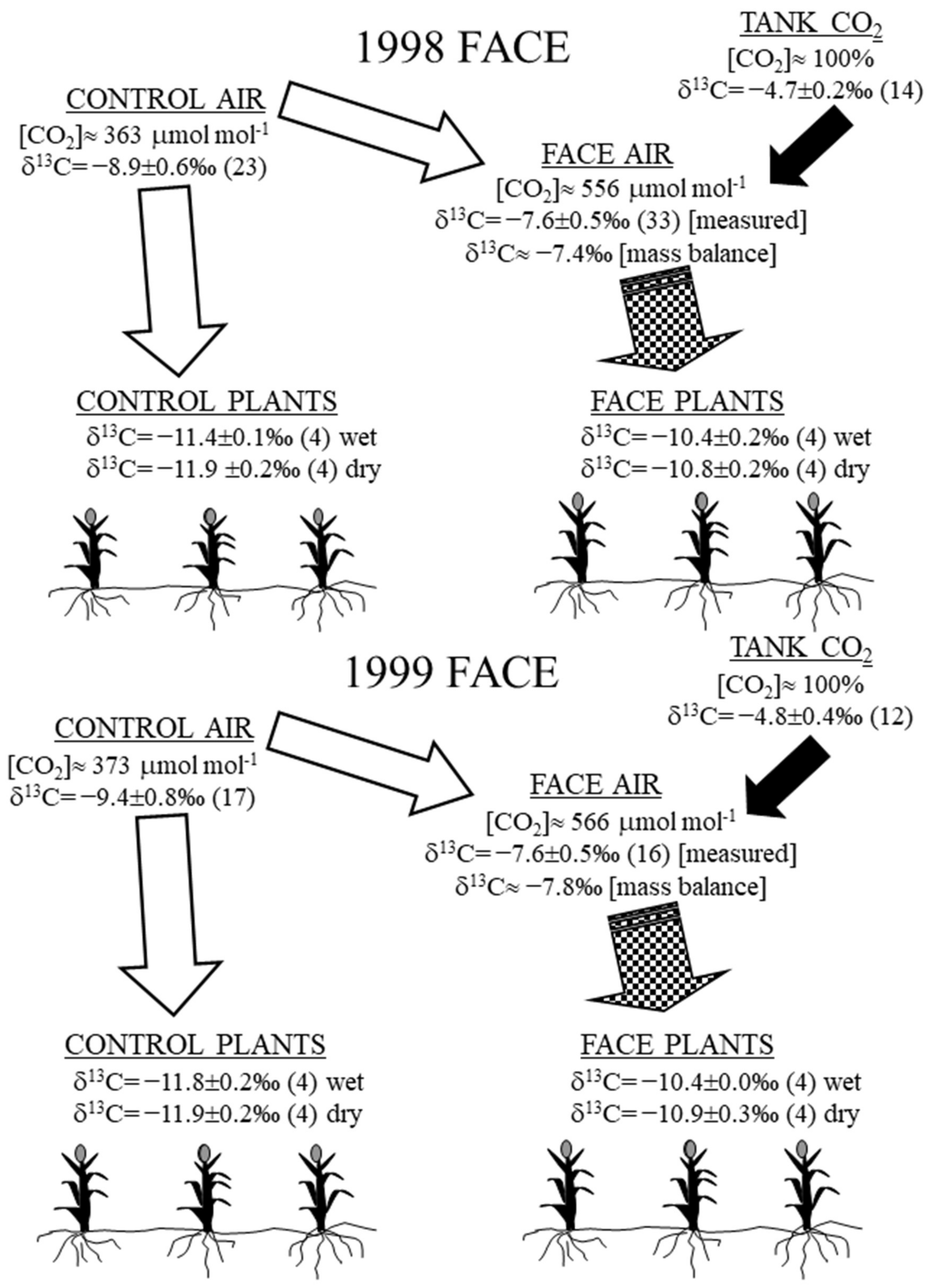
3.1. Air Isotopic Composition
3.2. Plant Isotopic Composition
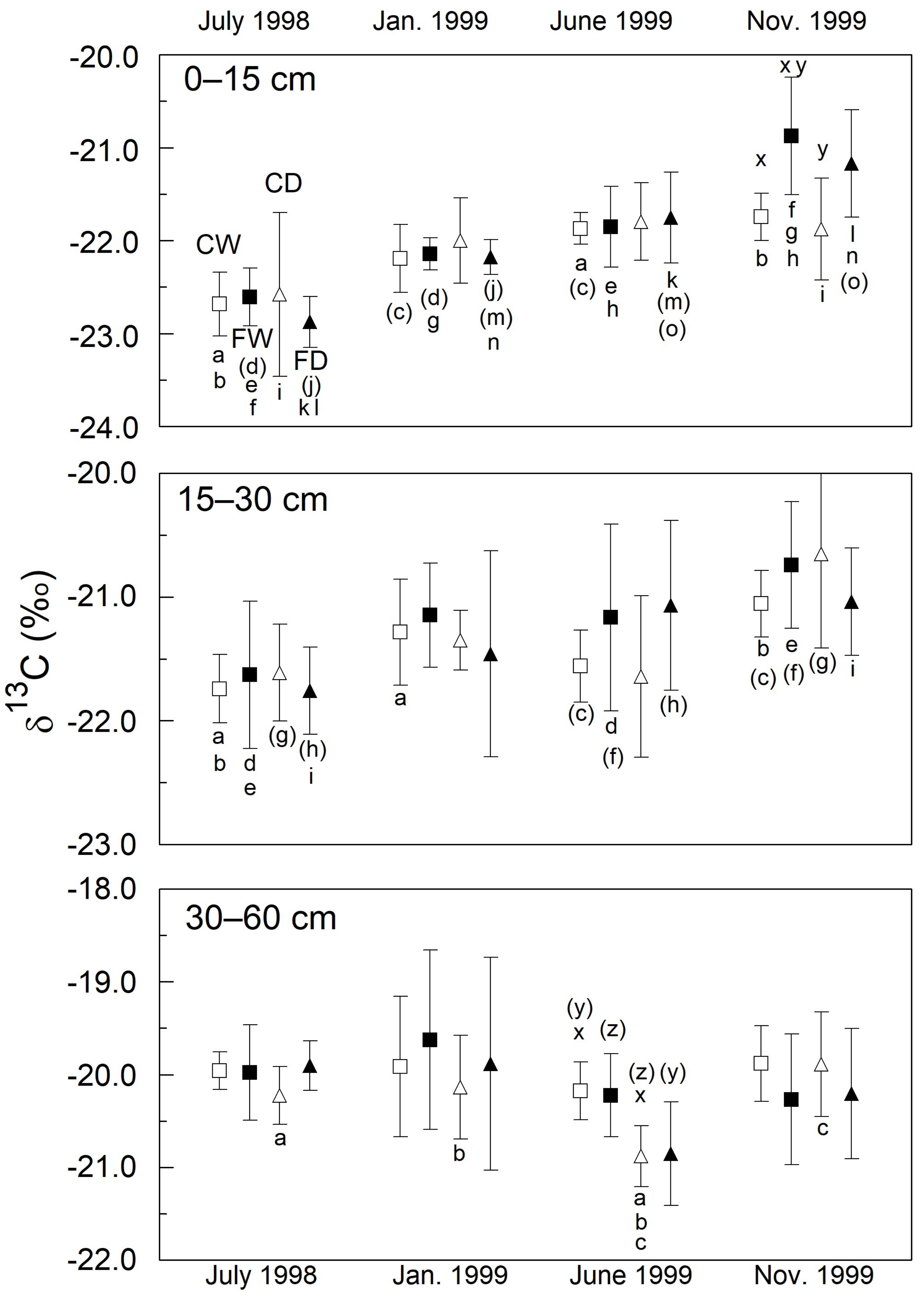
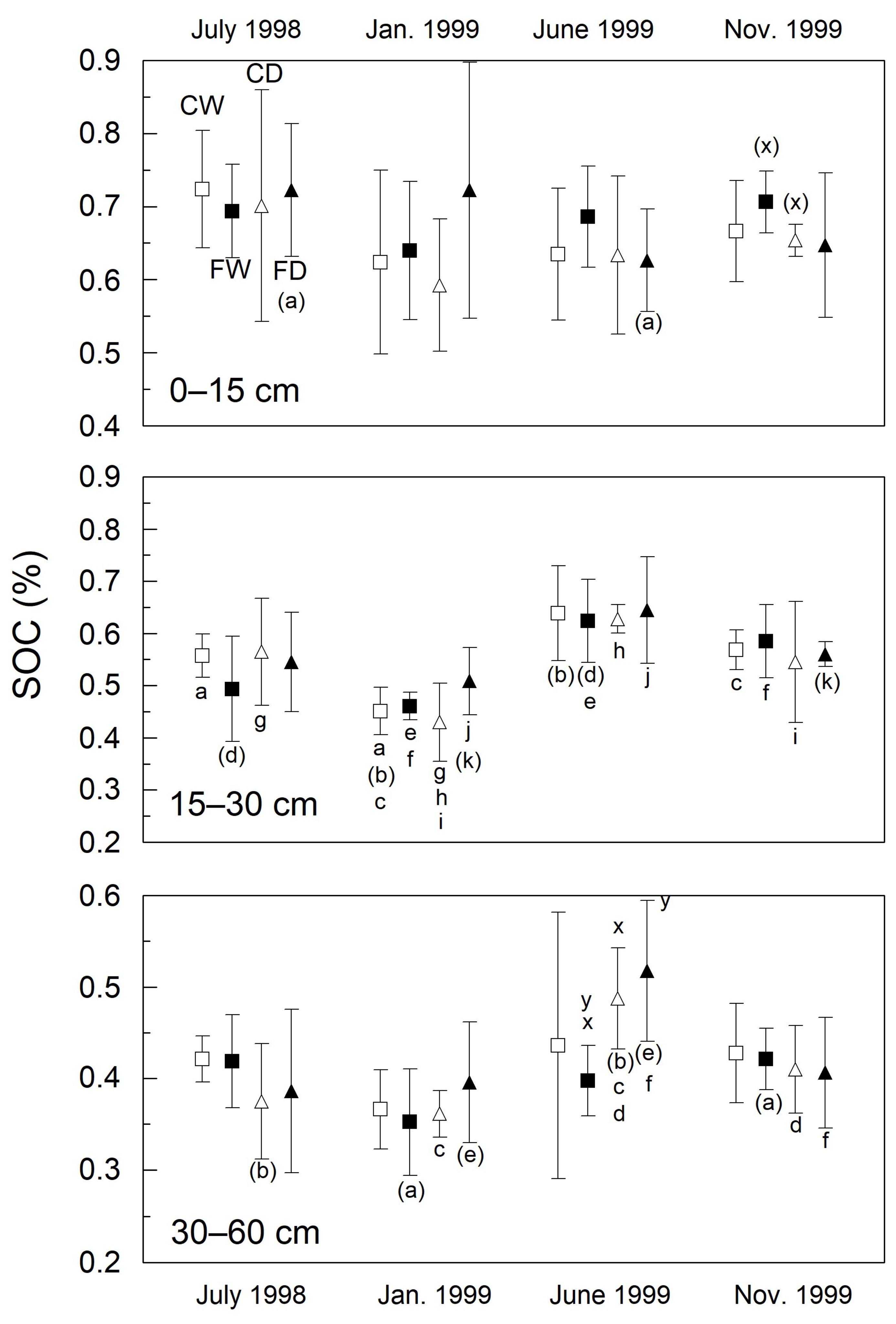
3.3. Soil Organic Carbon (%)
3.4. Isotopic Estimation of Carbon Input
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Statement
References
- Arias, P.A.; Bellouin, N.; Coppola, E.; Jones, R.G.; Krinner, G.; Marotzke, J.; Naik, V.; Palmer, M.D.; Plattner, G.-K.; Rogelj, J.; et al. Technical summary. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; Available online: https://www.ipcc.ch/report/ar6/wg1/downloads/report/IPCC_AR6_WGI_TS.pdf (accessed on 23 January 2022).
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 2004, 165, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Long, S.P. 30 years of free-air carbon dioxide enrichment (FACE): What have we learned about future crop productivity and its potential for adaptation? Glob. Chang. Biol. 2021, 27, 27–49. [Google Scholar] [CrossRef] [PubMed]
- Kimball, B.; Kobayashi, K.; Bindi, M. Responses of agricultural crops to free-air CO2 enrichment. Adv. Agron. 2002, 77, 293–368. [Google Scholar]
- Kimball, B. Crop responses to elevated CO2 and interactions with H2O, N, and temperature. Curr. Opin. Plant Biol. 2016, 31, 36–43. [Google Scholar] [CrossRef] [PubMed]
- U.S. DOE. U.S. Department of Energy Free-Air CO2 Enrichment Experiments: FACE Results, Lessons, and Legacy; DOE/SC–0202; U.S. Department of Energy Office of Science: Washington, DC, USA, 2020. [Google Scholar] [CrossRef]
- Shukla, P.R.; Skea, J.; Calvo Buendia, E.; Masson-Delmotte, V.; Pörtner, H.-O.; Roberts, D.C.; Zhai, P.; Slade, R.; Connors, S.; van Diemen, R.; et al. Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; The Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2019; Available online: https://www.ipcc.ch/site/assets/uploads/2019/11/SRCCL-Full-Report-Compiled-191128.pdf (accessed on 26 December 2021).
- Leavitt, S.W.; Paul, E.A.; Kimball, B.A.; Hendrey, G.R.; Mauney, J.R.; Rauschkolb, R.; Rogers, H.H., Jr.; Lewin, K.F.; Nagy, J.; Pinter, P.J., Jr.; et al. Carbon isotope dynamics of CO2-enriched FACE cotton and soils. Agric. For. Meteorol. 1994, 70, 87–101. [Google Scholar] [CrossRef]
- Leavitt, S.W.; Paul, E.A.; Galadima, A.; Nakayama, F.S.; Danzer, S.R.; Johnson, H.; Kimball, B.A. Carbon isotopes and carbon turnover in cotton and wheat FACE experiments. Plant Soil 1996, 187, 147–155. [Google Scholar] [CrossRef]
- Leavitt, S.W.; Pendall, E.; Paul, E.A.; Brooks, T.; Kimball, B.A.; Pinter, P.J., Jr.; Johnson, H.B.; Matthias, A.; Wall, G.W.; LaMorte, R.L. Stable-carbon isotopes and soil organic carbon in wheat under CO2 enrichment. New Phytol. 2001, 150, 305–314. [Google Scholar] [CrossRef]
- Nitschelm, J.J.; Luscher, A.; Hartwig, U.A.; Van Kessel, C. Using stable isotopes to determine soil carbon input differences under ambient and elevated atmospheric CO2 conditions. Glob. Chang. Biol. 1997, 3, 411–416. [Google Scholar] [CrossRef]
- Van Kessel, C.; Horwath, W.R.; Hartwig, U.; Harris, D.; Luscher, A. Net soil carbon input under ambient and elevated CO2 concentrations: Isotopic evidence after 4 years. Glob. Chang. Biol. 2000, 6, 435–444. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Lichter, J. Limited carbon storage in soil and litter of experimental forest plots under increased atmospheric CO2. Nature 2001, 411, 466–469. [Google Scholar] [CrossRef]
- Hungate, B.A.; Holland, E.A.; Jackson, R.B.; Chapin, F.S., III; Mooney, H.A.; Field, C.B. The fate of carbon under carbon dioxide enrichment. Nature 1997, 388, 576–579. [Google Scholar] [CrossRef]
- Torbert, H.A.; Rogers, H.H.; Prior, S.A.; Schlesinger, W.H.; Runion, G.B. Effects of elevated atmospheric CO2 in agro-ecosystems on soil carbon storage. Glob. Chang. Biol. 1997, 3, 513–521. [Google Scholar] [CrossRef] [Green Version]
- Ineson, P.; Cortufo, M.F.; Bol, R.; Harkness, D.D.; Blum, H. Quantification of soil carbon inputs under elevated CO2: C3 plants in a C4 soil. Plant Soil 1996, 187, 345–350. [Google Scholar] [CrossRef]
- Balesdent, J.; Balabane, M. Maize root-derived soil organic carbon estimated by natural 13C abundance. Soil Biol. Biochem. 1992, 24, 97–101. [Google Scholar] [CrossRef]
- Balesdent, J.; Mariotti, A.; Guillet, B. Natural 13C abundance as a tracer for studies of soil organic matter. Soil Biol. Biochem. 1987, 19, 25–30. [Google Scholar] [CrossRef]
- Balesdent, J.; Wagner, G.H.; Mariotti, A. Soil organic matter turnover in long-term field experiments as revealed by carbon-13 natural abundance. Soil Sci. Soc. Am. J. 1988, 52, 118–124. [Google Scholar] [CrossRef]
- Tissue, D.T.; Griffin, K.L.; Thomas, R.B.; Strain, B. Effects of low and elevated CO2 on C3 and C4 annuals II: Photosynthesis and leaf biochemistry. Oecologia 1995, 101, 21–28. [Google Scholar] [CrossRef]
- Poorter, H.; Roument, C.; Campbell, B.D. Interspecific variation in the growth response of plants to elevated CO2: A search for functional types. In Carbon Dioxide, Populations, and Communities; Körner, C., Bazzaz, F.A., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 375–411. [Google Scholar]
- Owensby, C.E.; Coyne, P.I.; Ham, J.M.; Auen, L.A.; Knapp, A.K. Biomass production in a tallgrass prairie system exposed to ambient and elevated CO2. Ecol. Appl. 1993, 3, 644–653. [Google Scholar] [CrossRef] [Green Version]
- Hamerlynck, E.P.; McAllister, C.A.; Knapp, A.K.; Ham, J.M.; Owensby, C.E. Photosynthetic gas exchange and water relation responses of three tallgrass prairie species to elevated carbon dioxide and moderate drought. Int. J. Plant Sci. 1997, 158, 608–616. [Google Scholar] [CrossRef]
- Ottman, M.J.; Kimball, B.A.; Pinter, P.J., Jr.; Wall, G.W.; Vanderlip, R.L.; Leavitt, S.W.; LaMorte, R.L.; Matthias, A.D.; Brooks, T.J. Elevated increases sorghum biomass under drought conditions. New Phytol. 2001, 150, 261–273. [Google Scholar] [CrossRef]
- Wall, G.W.; Kimball, B. Biological databases derived from Free-air carbon dioxide enrichment experiments. In Design and Execution of Experiments on CO2 Enrichment; Schulze, E.-D., Mooney, H.A., Eds.; Ecosystem Research Report 6; Commission of the European Communities: Brussels, Belgium, 1993; pp. 329–351. [Google Scholar]
- Triggs, J.M.; Kimball, B.A.; Pinter, P.J., Jr.; Wall, G.W.; Conley, M.M.; Brooks, T.J.; LaMorte, R.L.; Adam, N.R.; Ottman, M.J.; Matthias, A.D.; et al. Free-air carbon dioxide enrichment effects on energy balance and evapotranspiration of sorghum. Agric. For. Meteorol. 2004, 124, 63–79. [Google Scholar] [CrossRef]
- Conley, M.M.; Kimball, B.A.; Brooks, T.J.; Pinter, P.J.; Hunsaker, D.J.; Wall, G.W.; Adam, N.R.; LaMorte, R.L.; Matthias, A.D.; Thompson, T.L.; et al. CO2 enrichment increases water-use efficiency in sorghum. New Phytol. 2001, 151, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.Z.; van Loocke, A.; Siebers, M.H.; Ruiz-Vera, U.M.; Markelz, R.J.C.; Leakey, A.D.B.; Ort, D.R.; Bernacchi, C.J. Future carbon dioxide concentration decreases canopy evapotranspiration and soil water depletion by field-grown maize. Glob. Chang. Biol. 2013, 19, 1572–1584. [Google Scholar] [CrossRef] [PubMed]
- Manderscheid, R.; Erbs, M.; Weigel, H.-J. Interactive effects of free-air CO2 enrichment and drought stress on maize growth. Eur. J. Agron. 2014, 52, 11–21. [Google Scholar] [CrossRef]
- Cousins, A.B.; Adam, N.R.; Wall, G.W.; Kimball, B.A.; Pinter, P.J., Jr.; Leavitt, S.W.; LaMorte, R.L.; Matthias, A.D.; Ottman, M.J.; Thompson, T.L.; et al. Reduced photorespiration and increased energy-use efficiency in young CO2-enriched sorghum leaves. New Phytol. 2001, 150, 275–284. [Google Scholar] [CrossRef]
- Williams, D.G.; Gempko, V.; Fravolini, A.; Leavitt, S.W.; Wall, G.W.; Kimball, B.A.; Pinter, P.J., Jr.; LaMorte, R.L.; Ottman, M. Carbon isotope discrimination by Sorghum bicolor under CO2 enrichment and drought. New Phytol. 2001, 150, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Kimball, B.A.; Ottman, M.J.; Pinter, P.J., Jr.; Wall, G.W.; Leavitt, S.W.; Cheng, L.; Conley, M.M.; LaMorte, R.L.; Triggs, J.M.; Gleadow, R. Data from the Arizona FACE (Free-Air CO2 Enrichment) experiments on sorghum at ample and limiting levels of water. Open Data J. Agric. Res. 2021, 7, 1–10. [Google Scholar]
- Post, D.F.; Mack, C.; Camp, P.D.; Suliman, A.S. Mapping and characterization of the soils on the University of Arizona Maricopa Agricultural Center. Hydrol. Water Resour. Ariz. 1988, 18, 49–60. [Google Scholar]
- Kimball, B.A.; LaMorte, R.L.; Peresta, G.J.; Mauney, J.R.; Lewin, K.F.; Hendrey, G.R. Appendices: Weather, soils, cultural practices, and cotton growth data from the 1989 FACE experiment in IBSNAT format. CRC Crit. Rev. Plant Sci. 1992, 11, 271–308. [Google Scholar] [CrossRef]
- Nagy, J.; Lewin, K.F.; Hendrey, G.R.; Hassinger, E.; LaMorte, R. FACE facility CO2 concentration control and CO2 use in 1990 and 1991. Agric. For. Meteorol. 1994, 70, 31–48. [Google Scholar] [CrossRef]
- Pinter, P.J., Jr.; Kimball, B.A.; Wall, G.W.; LaMorte, R.L.; Hunsaker, D.J.; Adamsen, F.J.; Frumau, K.F.A.; Vugts, H.F.; Hendrey, G.R.; Lewin, K.F.; et al. Free-air CO2 enrichment (FACE): Blower effects on wheat canopy microclimate and plant development. Agric. For. Meteorol. 2000, 103, 319–332. [Google Scholar] [CrossRef]
- Hendrey, G.R. (Ed.) Free-Air Carbon Dioxide Enrichment for Plant Research in the Field; C.K. Smoley: Boca Raton, FL, USA, 1993; p. 308. [Google Scholar]
- Hendrey, G.; Lewin, K.; Nagy, J. Control of carbon dioxide in unconfined field plots. In Design and Execution of Experiments on CO2 Enrichment; Schulze, E.-D., Mooney, H.A., Eds.; Commission of the European Communities: Brussels, Belgium, 1993; pp. 309–327. [Google Scholar]
- Dugas, W.A.; Pinter, P.J., Jr. (Eds.) Free-Air Carbon Dioxide Enrichment (Face) Cotton Project: A New Field Approach to Assess the Biological Consequences of Global Change. Agric. For. Meteorol. 1994, 70, 1–342. [Google Scholar] [CrossRef]
- Fox, F.A., Jr.; Scherer, T.; Slack, D.C.; Clark, L.J. AriZona Irrigation SCHEDuling User’s Manual; Cooperative Extension, Agricultural and Biosystems Engineering, University of Arizona: Tucson, AZ, USA, 1992; p. 36. [Google Scholar]
- Cambardella, C.A.; Elliott, E.T. Particulate soil organic-matter changes across a grassland cultivation sequence. Soil Sci. Soc. Am. J. 1992, 56, 777–783. [Google Scholar] [CrossRef]
- Boutton, T.W. Stable carbon isotope ratios of natural materials: I. Sample preparation and mass spectrometric analysis. In Carbon Isotope Techniques; Coleman, D.C., Fry, B., Eds.; Academic Press: San Diego, CA, USA, 1991; pp. 173–175. [Google Scholar]
- Craig, H. Isotopic standards for carbon and oxygen and correction factors for mass-spectrometric analysis of CO2. Geochim. Cosmochim. Acta 1957, 12, 133–149. [Google Scholar] [CrossRef]
- Coplen, T.B. Discontinuance of SMOW and PDB. Nature 1995, 375, 285. [Google Scholar] [CrossRef]
- Henderson, S.; von Caemmerer, S.; Farquhar, G.D.; Wade, L.; Hammer, G. Correlation between carbon isotope discrimination and transpiration efficiency in lines of the C4 species Sorghum bicolor in the glasshouse and the field. Aust. J. Plant Physiol. 1998, 25, 111–123. [Google Scholar] [CrossRef]
- Leavitt, S.W.; Paul, E.A.; Pendall, E.; Pinter, P.J., Jr.; Kimball, B. Field variability of carbon isotopes in soil organic carbon. Nucl. Instrum. Methods Phys. Res. B 1997, 123, 451–454. [Google Scholar] [CrossRef]
- Cernusak, L.A.; Tcherkez, G.; Keitel, C.; Cornwell, W.K.; Santiago, L.S.; Knohl, A.; Barbour, M.M.; Williams, D.G.; Reich, P.B.; Ellsworth, D.S.; et al. Why are non-photosynthetic tissues generally 13C enriched compared with leaves in C3 plants? Review and synthesis of current hypotheses. Funct. Plant Biol. 2009, 36, 199–213. [Google Scholar] [CrossRef]
- Wall, G.W.; Brooks, T.J.; Adam, N.R.; Cousins, A.B.; Kimball, B.A.; Pinter, P.J., Jr.; LaMorte, R.L.; Triggs, J.; Ottman, M.J.; Leavitt, S.W.; et al. Elevated atmospheric CO2 improved sorghum plant water status by ameliorating the adverse effects of drought. New Phytol. 2001, 152, 231–248. [Google Scholar] [CrossRef]
- Jenkinson, D.S.; Fox, R.H.; Rayner, J.H. Interactions between fertilizer nitrogen and soil nitrogen—The so-called ‘priming’ effect. Eur. J. Soil Sci. 1985, 36, 425–444. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Friedel, J.K.; Stahr, K. Review of mechanisms and quantification of priming effects. Soil Biol. Biochem. 2000, 32, 1485–1498. [Google Scholar] [CrossRef]
- Pendall, E.; Leavitt, S.W.; Brooks, T.; Kimball, B.A.; Pinter, P.J., Jr.; Wall, G.W.; LaMorte, R.L.; Wechsung, G.; Wechsung, F.; Adamsen, F.; et al. Elevated CO2 stimulates soil respiration in a FACE wheat field. Basic Appl. Ecol. 2001, 2, 193–201. [Google Scholar] [CrossRef]
- Carney, K.M.; Hungate, B.A.; Drake, B.G.; Megonigal, J.P. Altered soil microbial community at elevated CO2 leads to loss of soil carbon. Proc. Natl. Acad. Sci. USA 2007, 104, 4990–4995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, L. Elevated Atmospheric CO2 Impacts Carbon Dynamics in a C4-Sorghum-Soil Agroecosystem—An Application of Stable Carbon Isotopes (δ13C) in Tracing the Fate of Carbon in the Atmosphere-Plant-Soil Ecosystem. Ph.D. Thesis, University of Arizona, Tucson, AZ, USA, 2005. [Google Scholar]
- Cheng, L.; Leavitt, S.W.; Kimball, B.A.; Pinter, P.J., Jr.; Ottman, M.J.; Matthias, A.; Wall, G.W.; Brooks, T.; Williams, D.G.; Thompson, T.L. Dynamics of labile and recalcitrant soil carbon pools in a sorghum free-air CO2 enrichment (FACE) agroecosystem. Soil Biol. Biochem. 2007, 39, 2250–2263. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| 1998 | FACE–Wet | Control–Wet | FACE–Dry | Control–Dry |
| mean | 2.8 | 2.5 | 3.2 | 3.0 |
| std dev | 0.2 | 0.0 | 0.2 | 0.2 |
| t-signif | p < 0.05 | ns | ||
| 1999 | FACE–Wet | Control–Wet | FACE–Dry | Control–Dry |
| mean | 2.8 | 2.4 | 3.6 | 2.5 |
| std dev | 0.0 | 0.2 | 0.3 | 0.1 |
| t-signif | p < 0.01 | p < 0.01 | ||
| 0–15 cm Depth | 15–30 cm Depth | |||||||
|---|---|---|---|---|---|---|---|---|
| Dry | Wet | Dry | Wet | |||||
| Control | FACE | Control | FACE | Control | FACE | Control | FACE | |
| δ13Cplant | −11.9 ± 0.2‰ | −10.8 ± 0.3 | −11.6 ± 0.3 | −10.4 ± 0.1 | −11.9 ± 0.2 | −10.8 ± 0.3 | −11.6 ± 0.3 | −10.4 ± 0.1 |
| δ13Csoil original (July 1998) | −22.6 ± 0.9‰ | −22.9 ± 0.3 | −22.7 ± 0.4 | −22.6 ± 0.3 | −21.6 ± 0.4 | −21.8 ± 0.4 | −21.7 ± 0.3 | −21.6 ± 0.6 |
| δ13Csoil (November 1999) | −21.9 ± 0.6‰ | −21.2 ± 0.6 | −21.8 ± 0.2 | −20.9 ± 0.6 | −20.6 ± 0.8 | −21.0 ± 0.4 | −21.0 ± 0.3 | −20.7 ± 0.5 |
| finput new-C fraction | 6.5% | 14.2 | 8.4 | 14.2 | 9.9 | 6.6 | 6.8 | 7.9 |
| Net CO2 effect (FACE-Control) | 7.7% | 5.8 | −3.3 | 1.1 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leavitt, S.W.; Cheng, L.; Williams, D.G.; Brooks, T.; Kimball, B.A.; Pinter, P.J.; Wall, G.W.; Ottman, M.J.; Matthias, A.D.; Paul, E.A.; et al. Soil Organic Carbon Isotope Tracing in Sorghum under Ambient CO2 and Free-Air CO2 Enrichment (FACE). Land 2022, 11, 309. https://0-doi-org.brum.beds.ac.uk/10.3390/land11020309
Leavitt SW, Cheng L, Williams DG, Brooks T, Kimball BA, Pinter PJ, Wall GW, Ottman MJ, Matthias AD, Paul EA, et al. Soil Organic Carbon Isotope Tracing in Sorghum under Ambient CO2 and Free-Air CO2 Enrichment (FACE). Land. 2022; 11(2):309. https://0-doi-org.brum.beds.ac.uk/10.3390/land11020309
Chicago/Turabian StyleLeavitt, Steven W., Li Cheng, David G. Williams, Talbot Brooks, Bruce A. Kimball, Paul J. Pinter, Gerard W. Wall, Michael J. Ottman, Allan D. Matthias, Eldor A. Paul, and et al. 2022. "Soil Organic Carbon Isotope Tracing in Sorghum under Ambient CO2 and Free-Air CO2 Enrichment (FACE)" Land 11, no. 2: 309. https://0-doi-org.brum.beds.ac.uk/10.3390/land11020309