Exploring the Use of Sentinel-2 Data to Monitor Heterogeneous Effects of Contextual Drought and Heatwaves on Mediterranean Forests

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
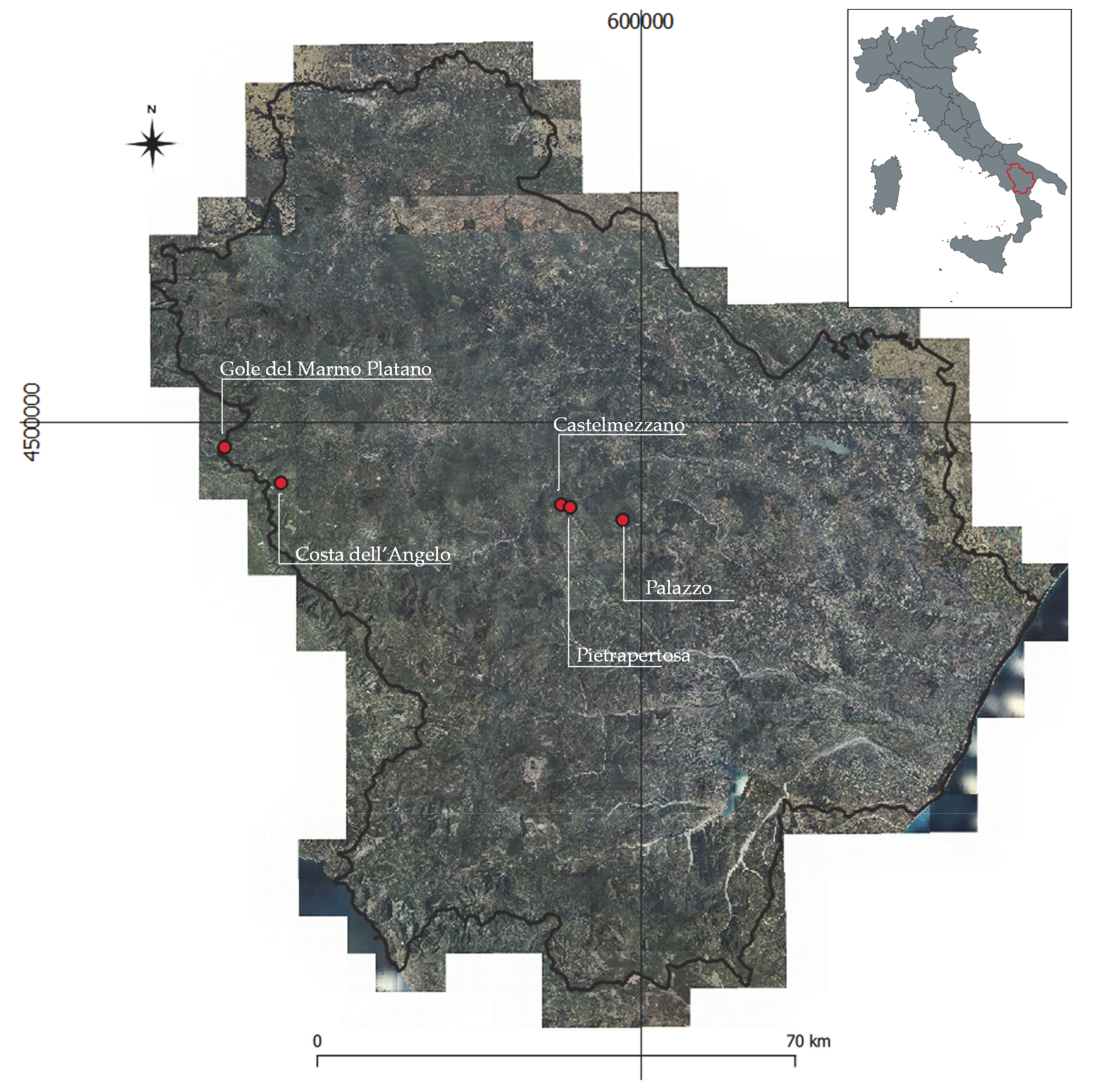
2. Study Area
3. Materials and Methods
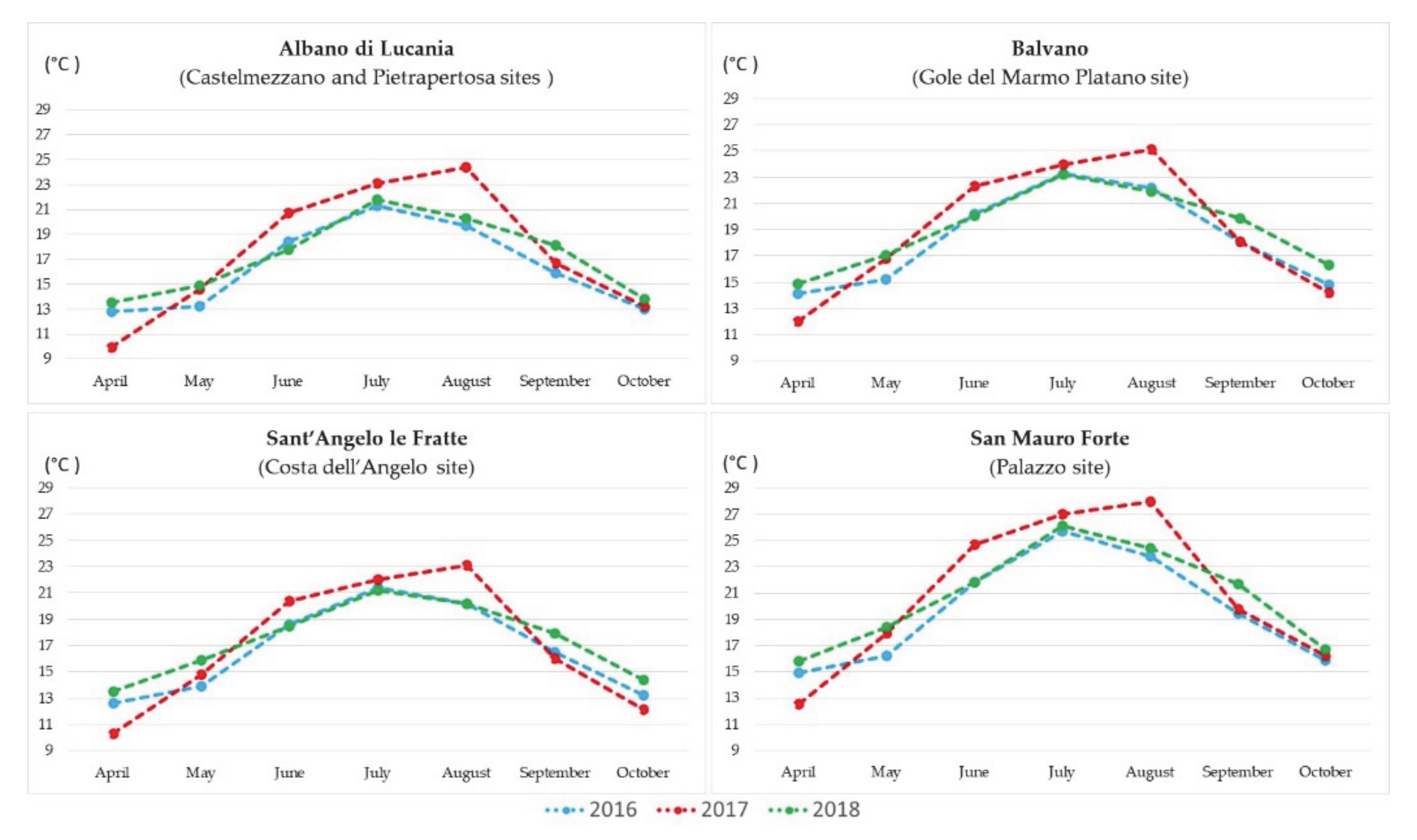
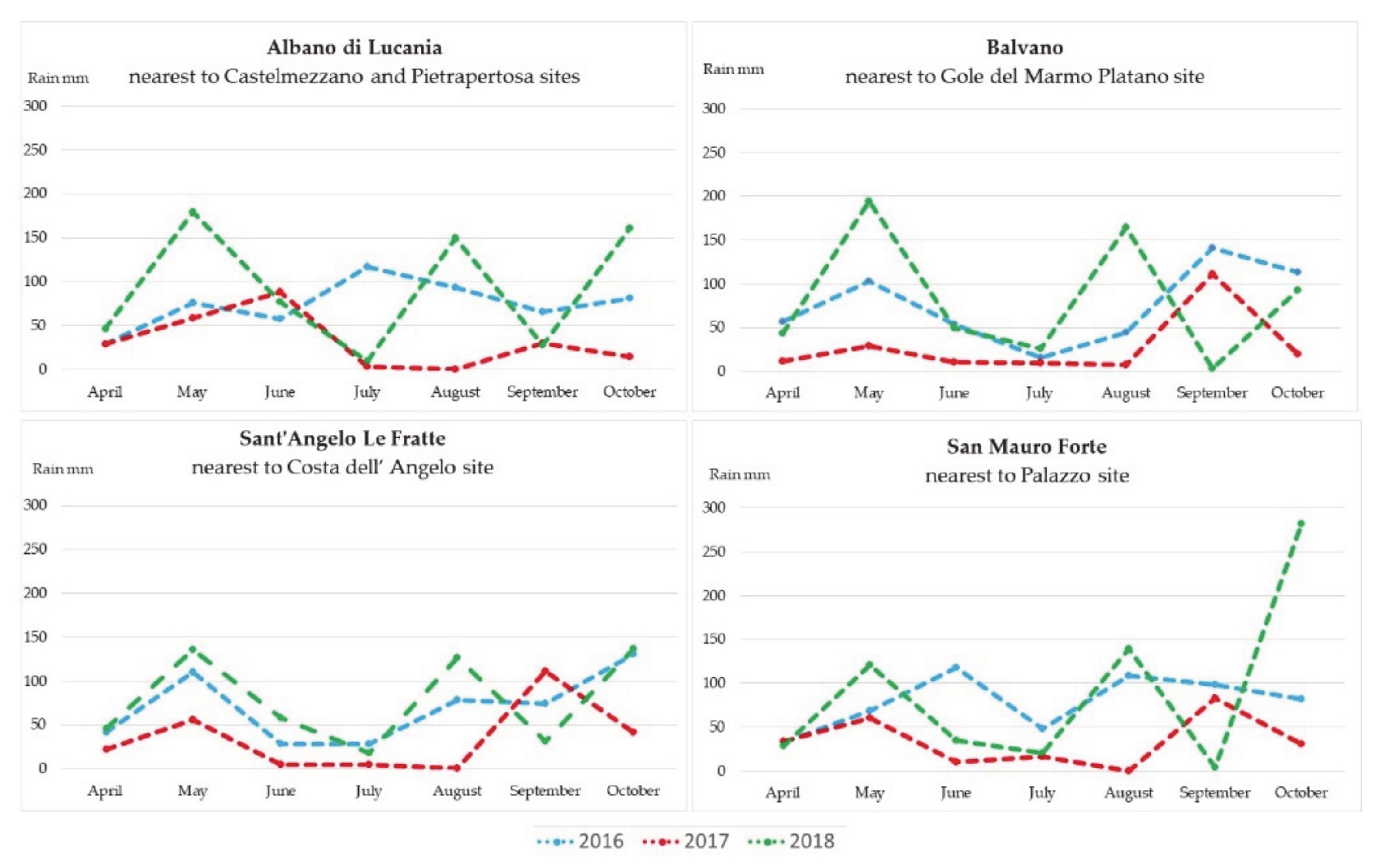
3.1. Meteorological Data
- Balvano station is closest to the Gole del Marmo Platano site;
- Sant’Angelo Le Fratte - Fiume Melandro station is closest to the Costa dell’Angelo site;
- Albano di Lucania station is closest to the Castelmezzano and Pietrapertosa sites;
- San Mauro Forte station is closest to the Palazzo site.
3.2. Satellite Data
3.3. In Situ Measurements
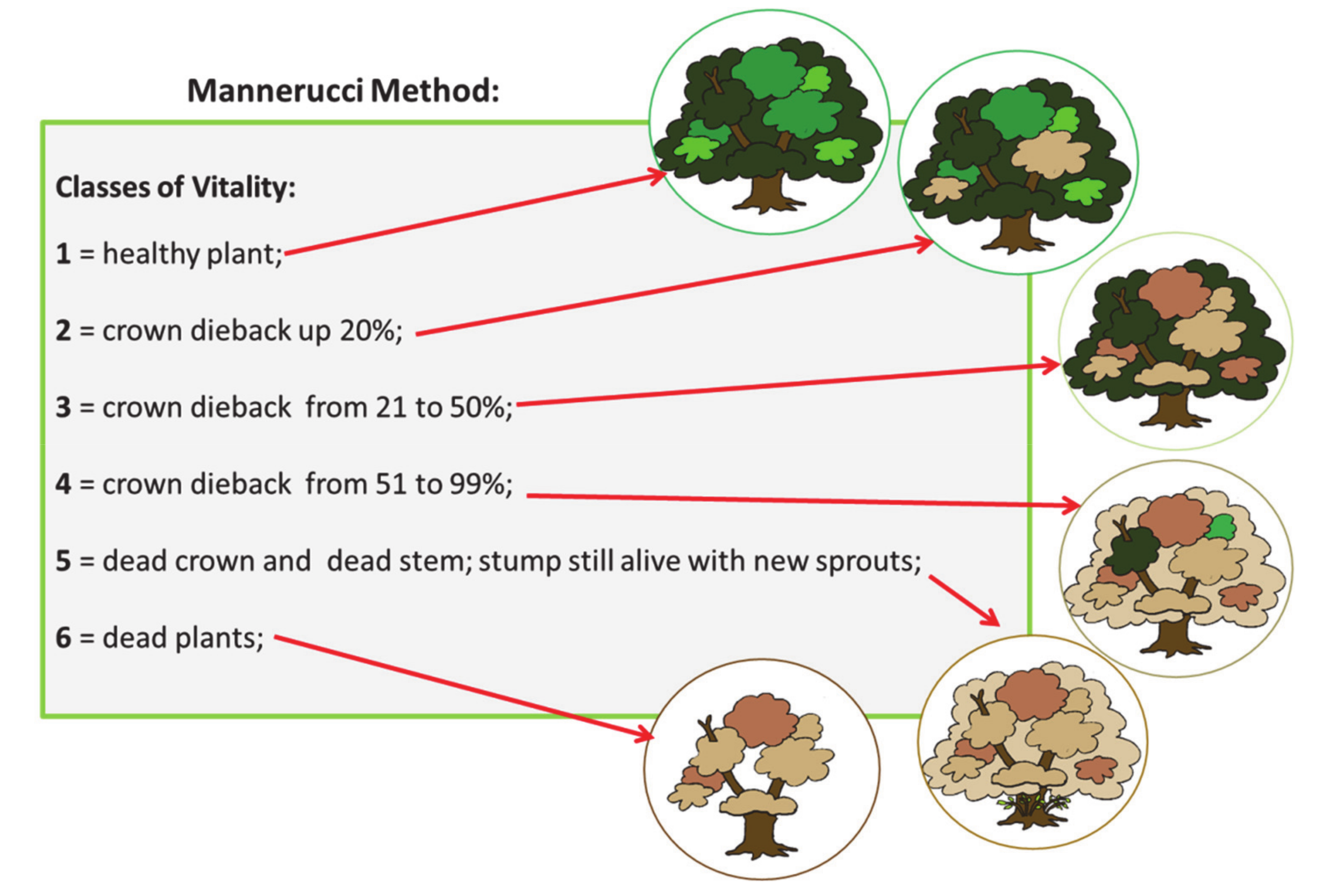
3.3.1. The Mannerucci Method
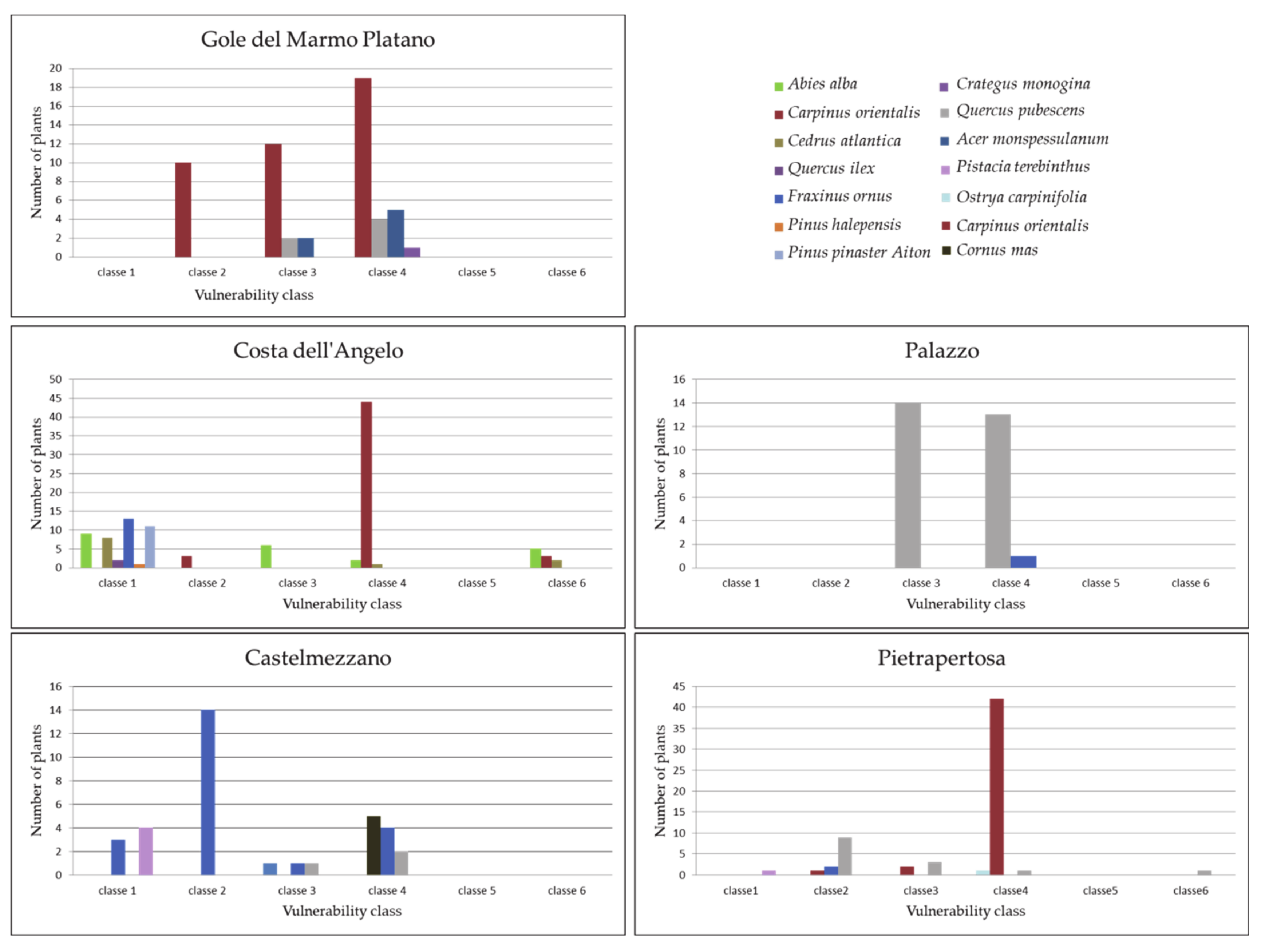
3.3.2. Floristic-Vegetational Sampling and Plant Functional Type Classification
4. Results and Discussion
4.1. Analysis of Meteorological Data
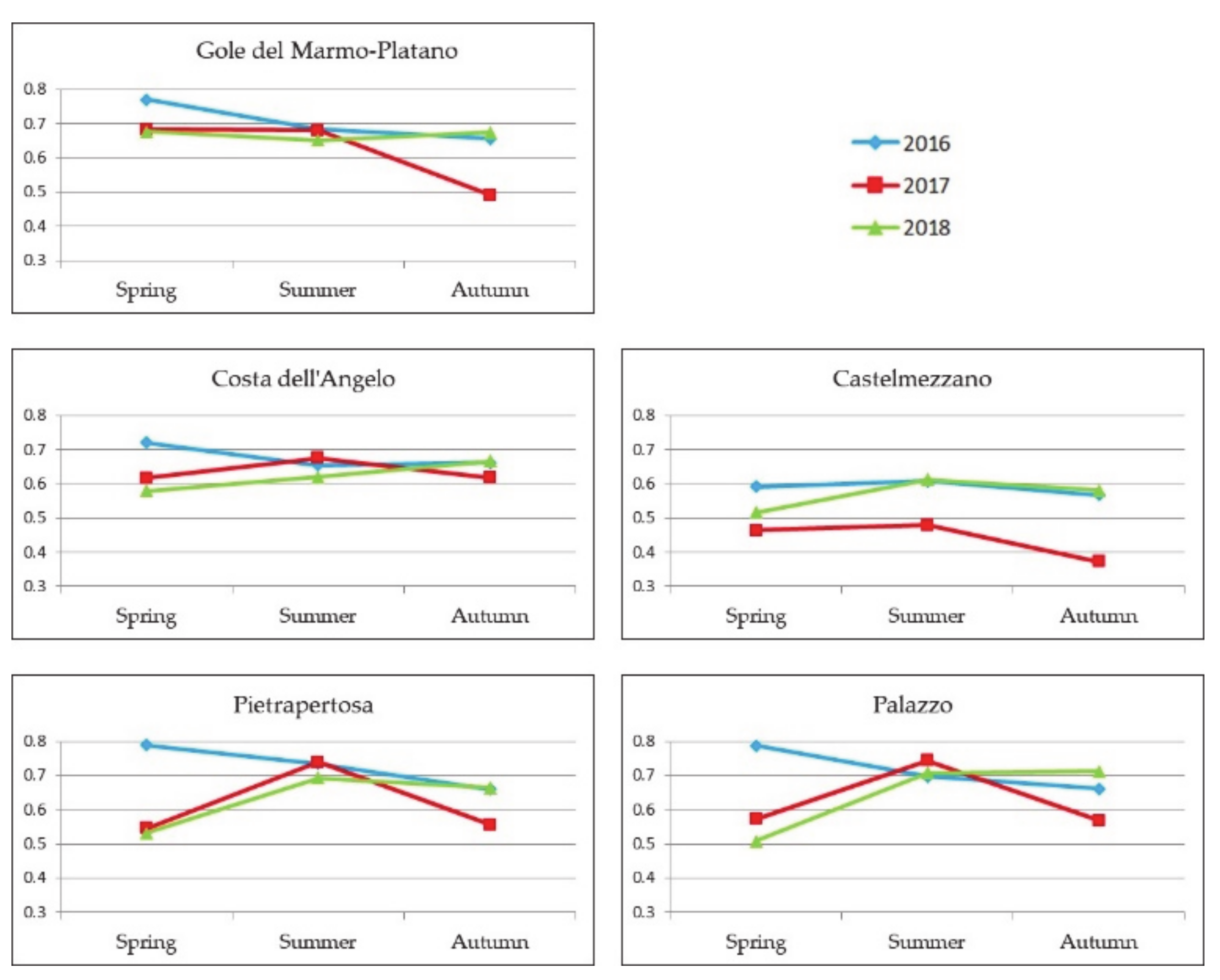
4.2. Analysis of Sentinel-2 NDVI
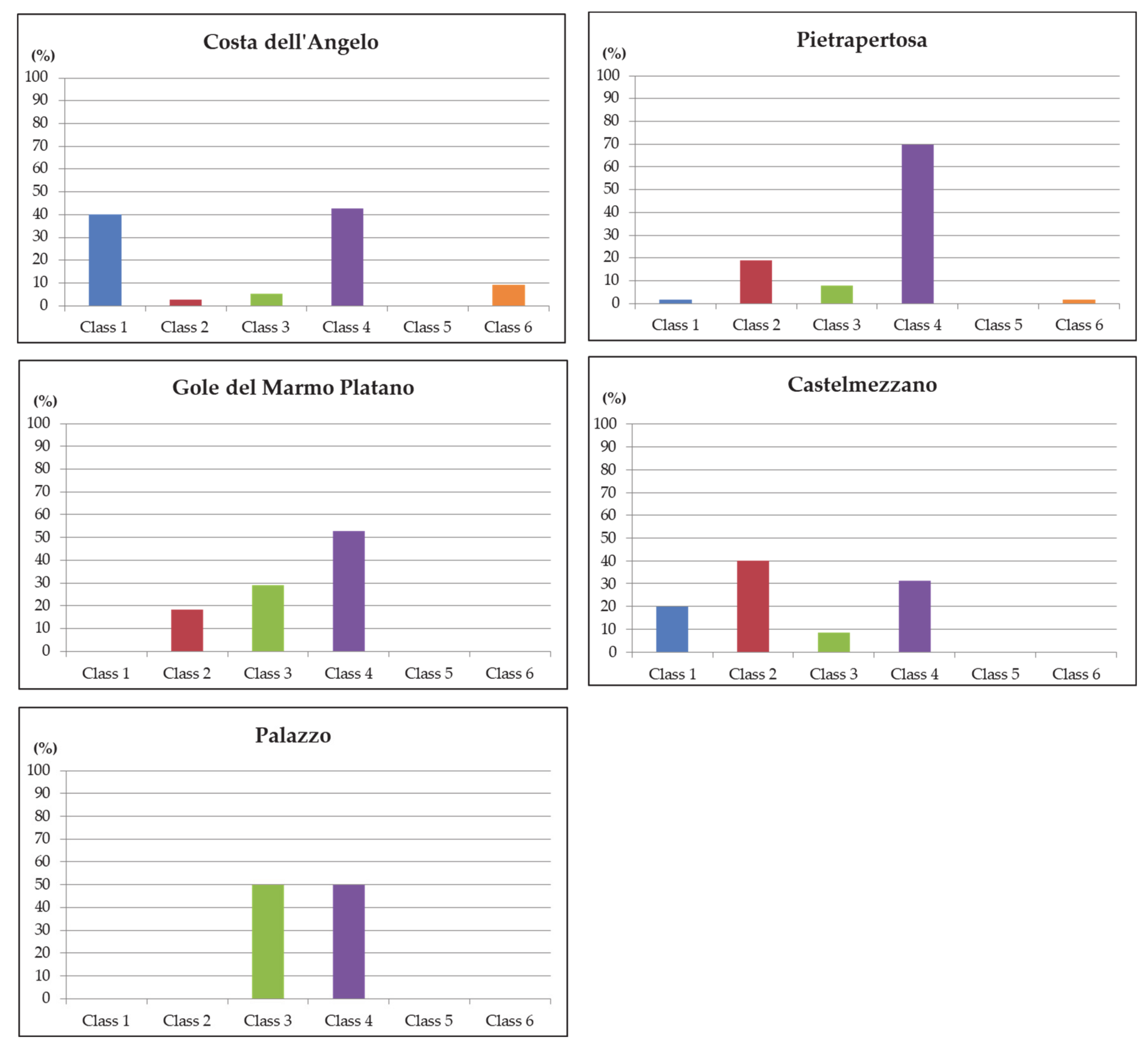
4.3. Analysis of In Situ Data
4.3.1. Gole del Marmo-Platano Site
4.3.2. Costa dell’Angelo Site
4.3.3. Palazzo Site
4.3.4. Castelmezzano Site
4.3.5. Pietrapertosa Site
4.4. Comparative Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lindner, M.; Fitzgerald, J.B.; Zimmermann, N.E.; Reyer, C.; Delzon, S.; van der Maaten, E.; Schelhaas, M.-J.; Lasch, P.; Eggers, J.; van der Maaten-Theunissen, M.; et al. Climate change and European forests: What do we know, what are the uncertainties, and what are the implications for forest management? J. Environ. Manag. 2014, 146, 69–83. [Google Scholar] [CrossRef] [Green Version]
- Millar, C.I.; Stephenson, N.L. Temperate forest health in an era of emerging megadisturbance. Science 2015, 349, 823–826. [Google Scholar] [CrossRef]
- The Intergovernmental Panel on Climate Change (IPCC). TAR Climate Change 2001: Impacts, Adaptation, and Vulnerability; Cambridge University Press: New York, NY, USA, 2001. [Google Scholar]
- Coumou, D.; Rahmstorf, S. A decade of weather extremes. Nat. Clim. Chang. 2012, 2, 491–496. [Google Scholar] [CrossRef]
- Easterling, D.R.; Meehl, G.A.; Parmesan, C.; Changnon, S.A.; Karl, T.R.; Mearns, L.O. Climate extremes: Observations, modeling, and impacts. Science 2000, 289, 2068–2074. [Google Scholar] [CrossRef] [Green Version]
- Ummenhofer, C.C.; Meehl, G.A. Extreme weather and climate events with ecological relevance: A review. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef]
- Field, C.B. Climate Change 2014—Impacts, Adaptation and Vulnerability: Regional Aspects; Cambridge University Press: New York, NY, USA, 2014; ISBN 978-1-107-05816-3. [Google Scholar]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Mariotti, A.; Pan, Y.; Zeng, N.; Alessandri, A. Long-term climate change in the Mediterranean region in the midst of decadal variability. Clim. Dyn. 2015, 44, 1437–1456. [Google Scholar] [CrossRef]
- Polade, S.D.; Gershunov, A.; Cayan, D.R.; Dettinger, M.D.; Pierce, D.W. Precipitation in a warming world: Assessing projected hydro-climate changes in California and other Mediterranean climate regions. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Bastida, F.; López-Mondéjar, R.; Baldrian, P.; Andrés-Abellán, M.; Jehmlich, N.; Torres, I.F.; García, C.; López-Serrano, F.R. When drought meets forest management: Effects on the soil microbial community of a Holm oak forest ecosystem. Sci. Total Environ. 2019, 662, 276–286. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Lopez-Moreno, J.-I.; Beguería, S.; Lorenzo-Lacruz, J.; Sanchez-Lorenzo, A.; García-Ruiz, J.M.; Azorin-Molina, C.; Morán-Tejeda, E.; Revuelto, J.; Trigo, R.; et al. Evidence of increasing drought severity caused by temperature rise in southern Europe. Environ. Res. Lett. 2014, 9, 044001. [Google Scholar] [CrossRef]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2013, 3, 52–58. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.T.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Shao, M.; Jia, X.; Wei, X. Relationship of Climatic and Forest Factors to Drought—And Heat-Induced Tree Mortality. PLoS ONE 2017, 12, e0169770. [Google Scholar] [CrossRef] [PubMed]
- Bréda, N.; Huc, R.; Granier, A.; Dreyer, E. Temperate forest trees and stands under severe drought: A review of ecophysiological responses, adaptation processes and long-term consequences. Ann. For. Sci. 2006, 63, 625–644. [Google Scholar] [CrossRef] [Green Version]
- Camarero, J.J.; Manzanedo, R.D.; Sanchez-Salguero, R.; Navarro-Cerrillo, R.M. Growth response to climate and drought change along an aridity gradient in the southernmost Pinus nigra relict forests. Ann. For. Sci. 2013, 70, 769–780. [Google Scholar] [CrossRef]
- Gentilesca, T.; Camele, I.; Colangelo, M.; Lauteri, M.; Lapolla, A.; Ripullone, F. Il declino dei soprassuoli di querce nel sud Italia: Il caso di studio del bosco di Gorgoglione. In Proceedings of the Atti del II Congresso Internazionale di Selvicoltura, Progettare il Futuro per il Settore Forestale, Florence, Italy, 26–29 November 2014; Volume 1, pp. 123–129. [Google Scholar]
- Ripullone, F.; Borghetti, M.; Raddi, S.; Vicinelli, E.; Baraldi, R.; Guerrieri, M.R.; Nolè, A.; Magnani, F. Physiological and structural changes in response to altered precipitation regimes in a Mediterranean macchia ecosystem. Trees 2009, 23, 823. [Google Scholar] [CrossRef]
- Puletti, N.; Mattioli, W.; Bussotti, F.; Pollastrini, M. Monitoring the effects of extreme drought events on forest health by Sentinel-2 imagery. JARS 2019, 13, 020501. [Google Scholar] [CrossRef]
- Adams, H.D.; Zeppel, M.J.B.; Anderegg, W.R.L.; Hartmann, H.; Landhäusser, S.M.; Tissue, D.T.; Huxman, T.E.; Hudson, P.J.; Franz, T.E.; Allen, C.D.; et al. A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat. Ecol. Evol. 2017, 1, 1285–1291. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Anderegg, W.R.L.; Vicente-Serrano, S.M. Impacts of droughts on the growth resilience of Northern Hemisphere forests. Glob. Ecol. Biogeogr. 2017, 26, 166–176. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Navarro-Cerrillo, R.; Camarero, J.; Fernández-Cancio, Á. Selective drought-induced decline of pine species in southeastern Spain. Clim. Chang. 2012, 113, 767–785. [Google Scholar] [CrossRef]
- Colangelo, M.; Camarero, J.J.; Borghetti, M.; Gentilesca, T.; Oliva, J.; Redondo, M.-A.; Ripullone, F. Drought and Phytophthora Are Associated With the Decline of Oak Species in Southern Italy. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Arzac, A.; García-Cervigón, A.I.; Vicente-Serrano, S.M.; Loidi, J.; Olano, J.M. Phenological shifts in climatic response of secondary growth allow Juniperus sabina L. to cope with altitudinal and temporal climate variability. Agric. For. Meteorol. 2016, 217, 35–45. [Google Scholar] [CrossRef]
- Colangelo, M.; Camarero, J.J.; Borghetti, M.; Gazol, A.; Gentilesca, T.; Ripullone, F. Size Matters a Lot: Drought-Affected Italian Oaks Are Smaller and Show Lower Growth Prior to Tree Death. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rita, A.; Camarero, J.J.; Nolè, A.; Borghetti, M.; Brunetti, M.; Pergola, N.; Serio, C.; Vicente-Serrano, S.M.; Tramutoli, V.; Ripullone, F. The impact of drought spells on forests depends on site conditions: The case of 2017 summer heat wave in southern Europe. Glob. Chang. Biol. 2020, 26, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Gentilesca, T.; Camarero, J.J.; Colangelo, M.; Nolè, A.; Ripullone, F. Drought-induced oak decline in the western Mediterranean region: An overview on current evidences, mechanisms and management options to improve forest resilience. iForest Biogeosci. For. 2017, 10, 796. [Google Scholar] [CrossRef] [Green Version]
- Azzari, G.; Grassini, P.; Edreira, J.I.R.; Conley, S.; Mourtzinis, S.; Lobell, D.B. Satellite mapping of tillage practices in the North Central US region from 2005 to 2016. Remote Sens. Environ. 2019, 221, 417–429. [Google Scholar] [CrossRef]
- Sharma, S.; Dhakal, K.; Wagle, P.; Kilic, A. Retrospective tillage differentiation using the Landsat-5 TM archive with discriminant analysis. Agrosyst. Geosci. Environ. 2020, 3, e20000. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.; Wu, B.; Yan, N.; Davdai, B.; Nasanbat, E. Suitability Assessment of Satellite-Derived Drought Indices for Mongolian Grassland. Remote Sens. 2017, 9, 650. [Google Scholar] [CrossRef] [Green Version]
- Moffet, C.A.; Taylor, J.B.; Booth, D.T. Very Large Scale Aerial (VLSA) Imagery for assessing postfire bitterbrush recovery. In Proceedings of the-Shrublands under fire: Disturbance and Recovery in a Changing World, Cedar City, UT, USA, 6–8 June 2006; Kitchen, S.G., Pendleton, R.L., Monaco, T.A., Vernon, J., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2008; Volume 52, pp. 161–168. [Google Scholar]
- Byer, S.; Jin, Y. Detecting Drought-Induced Tree Mortality in Sierra Nevada Forests with Time Series of Satellite Data. Remote Sens. 2017, 9, 929. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Ochsner, T.E.; Twidwell, D.; Carlson, J.D.; Krueger, E.S.; Engle, D.M.; Fuhlendorf, S.D. Nondestructive estimation of standing crop and fuel moisture content in tallgrass prairie. Rangel. Ecol. Manag. 2018, 71, 356–362. [Google Scholar] [CrossRef]
- Zhao, H.; Yang, C.; Guo, W.; Zhang, L.; Zhang, D. Automatic Estimation of Crop Disease Severity Levels Based on Vegetation Index Normalization. Remote Sens. 2020, 12, 1930. [Google Scholar] [CrossRef]
- Dotzler, S.; Hill, J.; Buddenbaum, H.; Stoffels, J. The Potential of EnMAP and Sentinel-2 Data for Detecting Drought Stress Phenomena in Deciduous Forest Communities. Remote Sens. 2015, 7, 14227–14258. [Google Scholar] [CrossRef] [Green Version]
- Bajocco, S.; De Angelis, A.; Salvati, L. A satellite-based green index as a proxy for vegetation cover quality in a Mediterranean region. Ecol. Indic. 2012, 23, 578–587. [Google Scholar] [CrossRef]
- D’Emilio, M.; Coluzzi, R.; Macchiato, M.; Imbrenda, V.; Ragosta, M.; Sabia, S.; Simoniello, T. Satellite data and soil magnetic susceptibility measurements for heavy metals monitoring: Findings from Agri Valley (Southern Italy). Environ. Earth Sci. 2018, 77, 63. [Google Scholar] [CrossRef]
- Greco, S.; Infusino, M.; De Donato, C.; Coluzzi, R.; Imbrenda, V.; Lanfredi, M.; Simoniello, T.; Scalercio, S. Late Spring Frost in Mediterranean Beech Forests: Extended Crown Dieback and Short-Term Effects on Moth Communities. Forests 2018, 9, 388. [Google Scholar] [CrossRef] [Green Version]
- Simoniello, T.; Coluzzi, R.; Imbrenda, V.; Lanfredi, M. Land cover changes and forest landscape evolution (1985–2009) in a typical Mediterranean agroforestry system (high Agri Valley). Nat. Hazards Earth Syst. Sci. 2015, 15, 1201–1214. [Google Scholar] [CrossRef] [Green Version]
- Simoniello, T.; Lanfredi, M.; Coppola, R.; Imbrenda, V.; Macchiato, M. Correlation of Vegetation and Air Temperature Seasonal Profiles—Spatial Arrangement and Temporal Variability. In Phenology Climate Change; IntechOpen: Rijeka, Croatia, 2012. [Google Scholar] [CrossRef]
- Imbrenda, V.; D’Emilio, M.; Lanfredi, M.; Simoniello, T.; Ragosta, M.; Macchiato, M. Integrated Indicators for the Estimation of Vulnerability to Land Degradation. In Soil Processes and Current Trends in Quality Assessment; IntechOpen: Rijeka, Croatia, 2013. [Google Scholar] [CrossRef] [Green Version]
- Bonfiglio, A.; Cuomo, V.; Lanfredi, M.; Macchiato, M. Interfacing NOAA/ANHRR NDVI and soil truth maps for monitoring vegetation phenology at a local scale in a heterogeneous landscape of Southern Italy. Int. J. Remote Sens. 2002, 23, 4181–4195. [Google Scholar] [CrossRef]
- Mancino, G.; Nolè, A.; Ripullone, F.; Ferrara, A. Landsat TM imagery and NDVI differencing to detect vegetation change: Assessing natural forest expansion in Basilicata, southern Italy. iForest Biogeosci. For. 2014, 7, 75. [Google Scholar] [CrossRef] [Green Version]
- Goto, K.; Goto, T.; Nmor, J.C.; Minematsu, K.; Gotoh, K. Evaluating salinity damage to crops through satellite data analysis: Application to typhoon affected areas of southern Japan. Nat. Hazards 2015, 75, 2815–2828. [Google Scholar] [CrossRef] [Green Version]
- Rivero, R.G.; Grunwald, S.; Binford, M.W.; Osborne, T.Z. Integrating spectral indices into prediction models of soil phosphorus in a subtropical wetland. Remote Sens. Environ. 2009, 113, 2389–2402. [Google Scholar] [CrossRef]
- Pause, M.; Schweitzer, C.; Rosenthal, M.; Keuck, V.; Bumberger, J.; Dietrich, P.; Heurich, M.; Jung, A.; Lausch, A. In Situ/Remote Sensing Integration to Assess Forest Health—A Review. Remote Sens. 2016, 8, 471. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, M.J.; Jessen, B.; Barry, M.J.; Figueroa, M.; McIntosh, J.; Murray, T.; Schmid, J.; Muller-Karger, F.E. Automated High-Resolution Time Series Mapping of Mangrove Forests Damaged by Hurricane Irma in Southwest Florida. Remote Sens. 2020, 12, 1740. [Google Scholar] [CrossRef]
- Meiforth, J.J.; Buddenbaum, H.; Hill, J.; Shepherd, J.D.; Dymond, J.R. Stress Detection in New Zealand Kauri Canopies with WorldView-2 Satellite and LiDAR Data. Remote Sens. 2020, 12, 1906. [Google Scholar] [CrossRef]
- Coluzzi, R.; Imbrenda, V.; Lanfredi, M.; Simoniello, T. A first assessment of the Sentinel-2 Level 1-C cloud mask product to support informed surface analyses. Remote Sens. Environ. 2018, 217, 426–443. [Google Scholar] [CrossRef]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; Oxford University Press: London, UK, 1934. [Google Scholar]
- Mannerucci, F.; Sicoli, G. Oak decline in Apulia southern Italy: An ecological indicator of landscape evolution? In Patterns and Processes in Forest Landscapes. Consequences of Human Management; Lafortezza, R., Sanesi, G., Eds.; Locorotondo: Bari, Italy, 2006. [Google Scholar]
- Coluzzi, R.; D’Emilio, M.; Imbrenda, V.; Giorgio, G.A.; Lanfredi, M.; Macchiato, M.; Ragosta, M.; Simoniello, T.; Telesca, V. Investigating climate variability and long-term vegetation activity across heterogeneous Basilicata agroecosystems. Geomat. Nat. Hazards Risk 2019, 10, 168–180. [Google Scholar] [CrossRef] [Green Version]
- Rivas-Martinez, S.; Penas, A.; Diaz, T.E. Biogeographic map of Europe; University of Leon: Leon, Spain, 2004. [Google Scholar]
- Pesaresi, S.; Biondi, E.; Casavecchia, S. Bioclimates of Italy. J. Maps 2017, 13, 955–960. [Google Scholar] [CrossRef]
- Blasi, C.; Capotorti, G.; Copiz, R.; Guida, D.; Mollo, B.; Smiraglia, D.; Zavattero, L. Classification and mapping of the ecoregions of Italy. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2014, 148, 1255–1345. [Google Scholar] [CrossRef]
- Rouse, J.W.; Deering, D.W.; Haas, R.H.; Schell, J.A. Monitoring Vegetation Systems in the Great Plains with ERTS; NASA: Washington, DC, USA, 1974; Volume 1. [Google Scholar]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef] [Green Version]
- Simoniello, T.; Lanfredi, M.; Liberti, M.; Coppola, R.; Macchiato, M. Estimation of vegetation cover resilience from satellite time series. Hydrol. Earth Syst. Sci. 2008, 12, 1053–1064. [Google Scholar] [CrossRef] [Green Version]
- Lanfredi, M.; Coppola, R.; Simoniello, T.; Coluzzi, R.; D’Emilio, M.; Imbrenda, V.; Macchiato, M. Early Identification of Land Degradation Hotspots in Complex Bio-Geographic Regions. Remote Sens. 2015, 7, 8154–8179. [Google Scholar] [CrossRef] [Green Version]
- Quaranta, G.; Salvia, R.; Salvati, L.; Paola, V.D.; Coluzzi, R.; Imbrenda, V.; Simoniello, T. Long-term impacts of grazing management on land degradation in a rural community of Southern Italy: Depopulation matters. Land Degrad. Dev. 2020, 2020, 1–16. [Google Scholar] [CrossRef]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Zhou, L.; Chen, A. Effect of climate and CO2 changes on the greening of the Northern Hemisphere over the past two decades. Geophys. Res. Lett. 2006, 33. [Google Scholar] [CrossRef] [Green Version]
- Wardlow, B.D.; Tadesse, T.; Brown, J.F.; Callahan, K.; Swain, S.; Hunt, E. Vegetation Drought Response Index: An Integration of Satellite, Climate, and Biophysical Data; Drought Mitigation Center Faculty Publications: Boca Raton, FL, USA, 2012. [Google Scholar]
- QGIS Development Team QGIS Geographic Information System. Open Source Geospatial Foundation Project (2019). Available online: https://www.qgis.org/en/site/ (accessed on 28 August 2020).
- Costantini, G.; Bellotti, A.; Mancino, G.; Borghetti, M.; Ferrara, A. Forest map of Regione Basilicata—Atlas; INEA: Potenza, Italy, 2006; ISBN 88-8145-062-3. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie: Grundzüge der Vegetationskunde; Springer: Vienna, Austria; New York, NY, USA, 1964; ISBN 978-3-7091-4078-9. [Google Scholar]
- Duckworth, J.C.; Kent, M.; Ramsay, P.M. Plant functional types: An alternative to taxonomic plant community description in biogeography? Prog. Phys. Geogr. Earth Environ. 2000, 24, 515–542. [Google Scholar] [CrossRef]
- Godron, M.; Daget, P.; Emberger, L.; Long, G.; Le Floch, E.; Poissonet, J.; Sauvage, C.; Wacquant, J.P. Vade-mecum pour le Relevé Méthodique de la Végétation et du Milieu—CNRS Editions; Éditions du Centre National de la Recherche Scientifique: Paris, France, 1969. [Google Scholar]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; Wiley: New York, NY, USA, 1974; ISBN 978-0-471-62290-1. [Google Scholar]
- Cain, S.A. Life-Forms and Phytoclimate. Bot. Rev. 1950, 16, 1–32. [Google Scholar] [CrossRef]
- Fitchett, J.M.; Grab, S.W.; Thompson, D.I. Plant phenology and climate change: Progress in methodological approaches and application. Prog. Phys. Geogr. Earth Environ. 2015, 39, 460–482. [Google Scholar] [CrossRef]
- Parmesan, C.; Hanley, M.E. Plants and climate change: Complexities and surprises. Ann. Bot. 2015, 116, 849–864. [Google Scholar] [CrossRef]
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italy, 1982. [Google Scholar]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Lanfredi, M.; Coluzzi, R.; Imbrenda, V.; Macchiato, M.; Simoniello, T. Analyzing Space–Time Coherence in Precipitation Seasonality across Different European Climates. Remote Sens. 2020, 12, 171. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Area | Coordinates | Altitude (m, a.s.l.) | Aspect | Slope (%) |
|---|---|---|---|---|
| Gole del Marmo Platano | 40°37.039′ N | 500 | North/West | 30 |
| 15°27.600′ E | ||||
| Costa dell’Angelo | 40°34.211′ N | 770 | North/Northwest | 30 |
| 15°33.466′ E | ||||
| Palazzo | 40°30.948′ N | 790 | West | 15 |
| 16°8.878′ E | ||||
| Castelmezzano | 40°32.040′ N | 682 | South/Southeast | 50 |
| 16°3.284′ E | ||||
| Pietrapertosa | 40°32.015′ N | 625 | North/Northwest | 35 |
| 16°3.476′E |
| Sensing Date | Cloud Percentage (%) |
|---|---|
| 23 May 2016 | 0 |
| 22 July 2016 | 0 |
| 30 September 2016 | 2.5 |
| 8 April 2017 | 0 |
| 12 July 2017 | 0 |
| 5 October 2017 | 0 |
| 13 April 2018 | 0.12 |
| 2 July 2018 | 0 |
| 30 September 2018 | 0.47 |
| NDVI Autumn Values—NDVI Summer Values | |||
|---|---|---|---|
| 2016 | 2017 | 2018 | |
| Costa dell’Angelo | 0.01 | −0.06 | 0.05 |
| Gole del Marmo Platano | −0.03 | −0.19 | 0.02 |
| Pietrapertosa | −0.07 | −0.18 | −0.03 |
| Castelmezzano | −0.04 | −0.11 | −0.03 |
| Palazzo | −0.04 | −0.18 | 0.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coluzzi, R.; Fascetti, S.; Imbrenda, V.; Italiano, S.S.P.; Ripullone, F.; Lanfredi, M. Exploring the Use of Sentinel-2 Data to Monitor Heterogeneous Effects of Contextual Drought and Heatwaves on Mediterranean Forests. Land 2020, 9, 325. https://0-doi-org.brum.beds.ac.uk/10.3390/land9090325
Coluzzi R, Fascetti S, Imbrenda V, Italiano SSP, Ripullone F, Lanfredi M. Exploring the Use of Sentinel-2 Data to Monitor Heterogeneous Effects of Contextual Drought and Heatwaves on Mediterranean Forests. Land. 2020; 9(9):325. https://0-doi-org.brum.beds.ac.uk/10.3390/land9090325
Chicago/Turabian StyleColuzzi, Rosa, Simonetta Fascetti, Vito Imbrenda, Santain Settimio Pino Italiano, Francesco Ripullone, and Maria Lanfredi. 2020. "Exploring the Use of Sentinel-2 Data to Monitor Heterogeneous Effects of Contextual Drought and Heatwaves on Mediterranean Forests" Land 9, no. 9: 325. https://0-doi-org.brum.beds.ac.uk/10.3390/land9090325