Aging Promotes Mitochondria-Mediated Apoptosis in Rat Hearts

 , , , , , ,
, , , , , ,  ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments and Ethical Approval
2.2. Tissue Preparation
2.3. Hematoxylin and Eosin Staining
2.4. Western Immunoblotting
2.5. Mitochondrial Permeability Transition Pore (mPTP) Opening Sensitivity
2.6. Immunohistochemistry
2.7. Terminal Deoxynucleotidyl Transferase-Mediated dUTP Nick-End Labeling (TUNEL Assay)
2.8. Statistical Analysis
3. Results
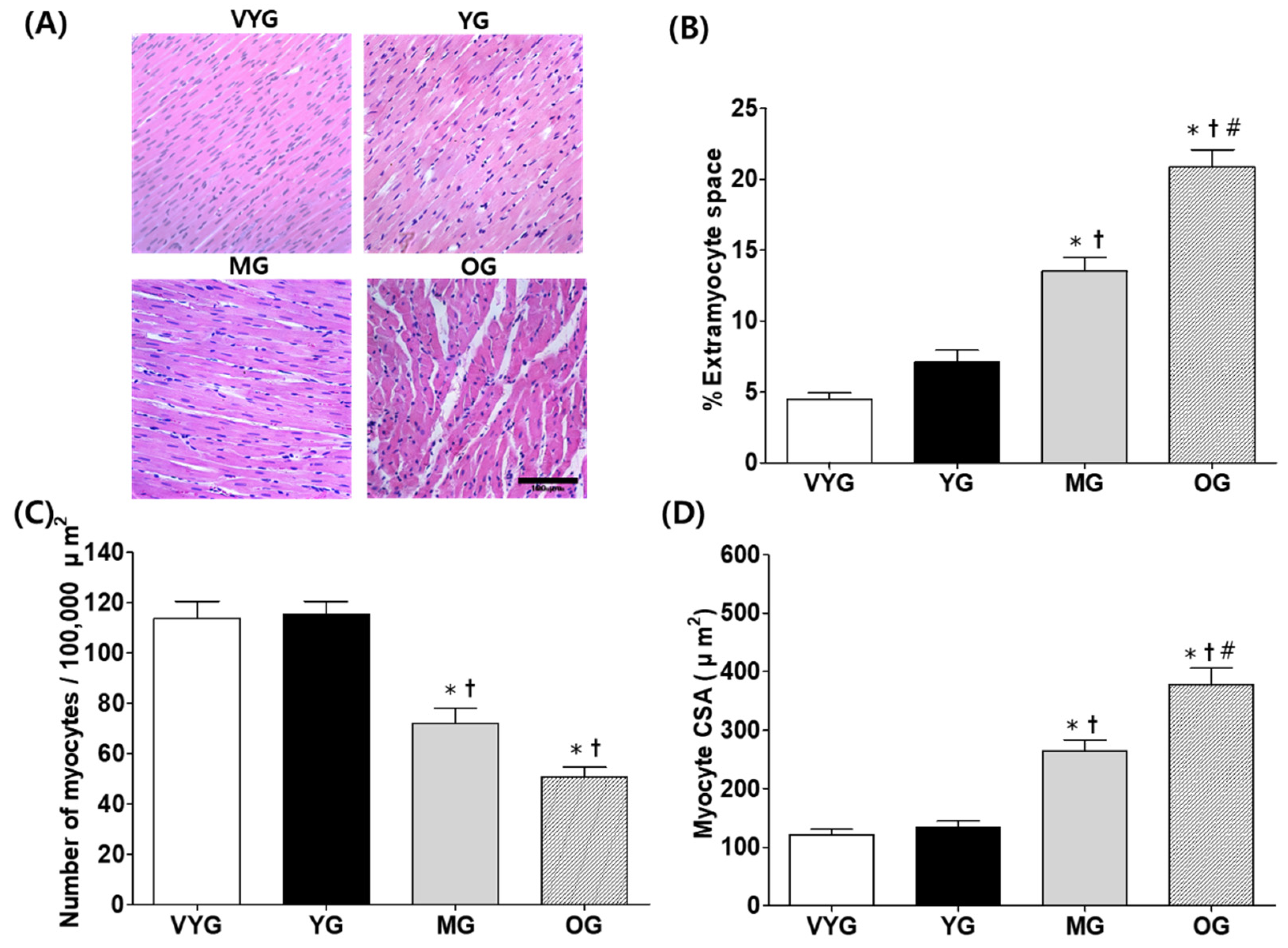
3.1. Effects of Aging on Cardiac Muscle Morphology
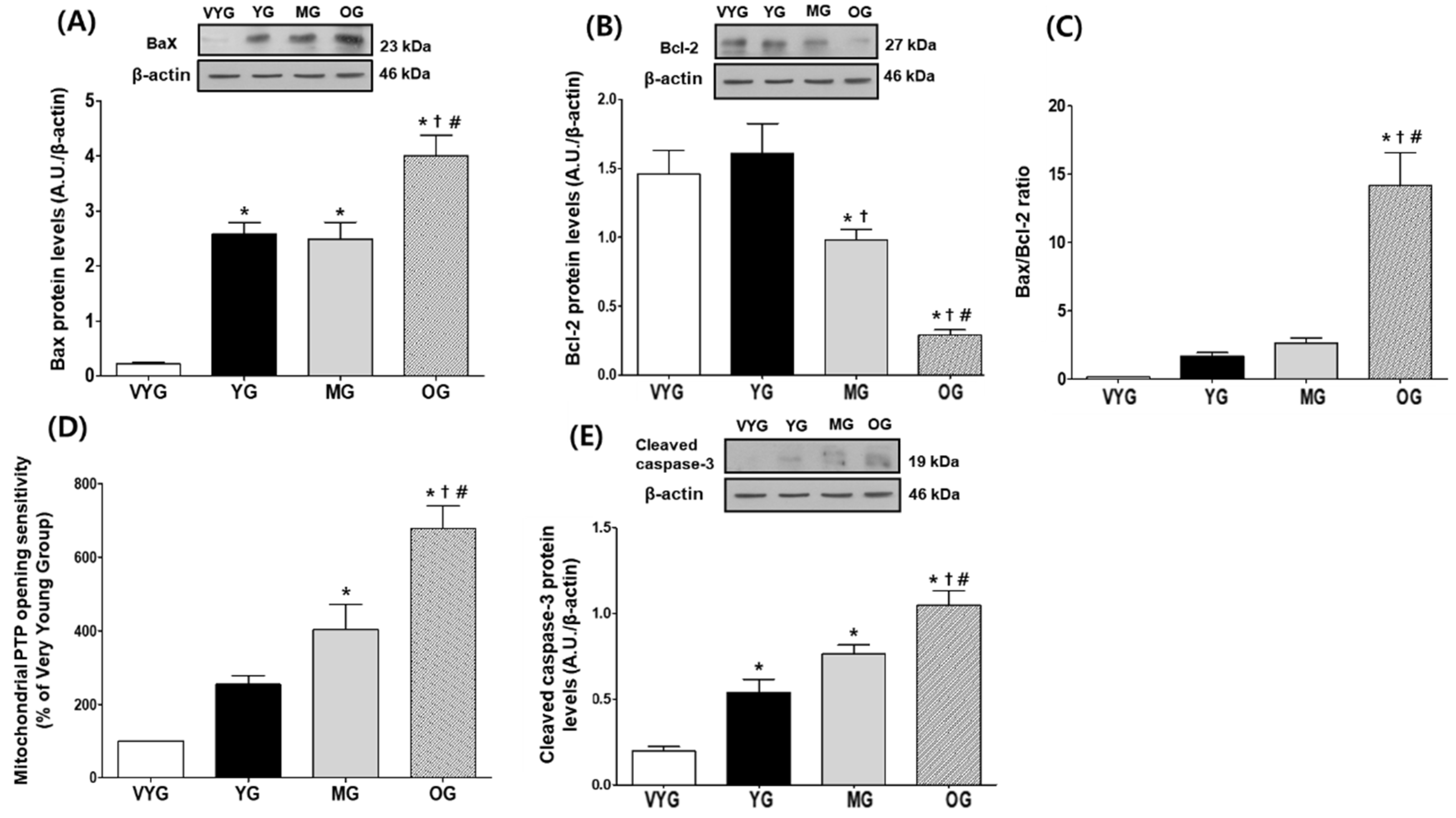
3.2. Effects of Aging on Mitochondria-Mediated Apoptotic Signaling in Cardiac Muscles
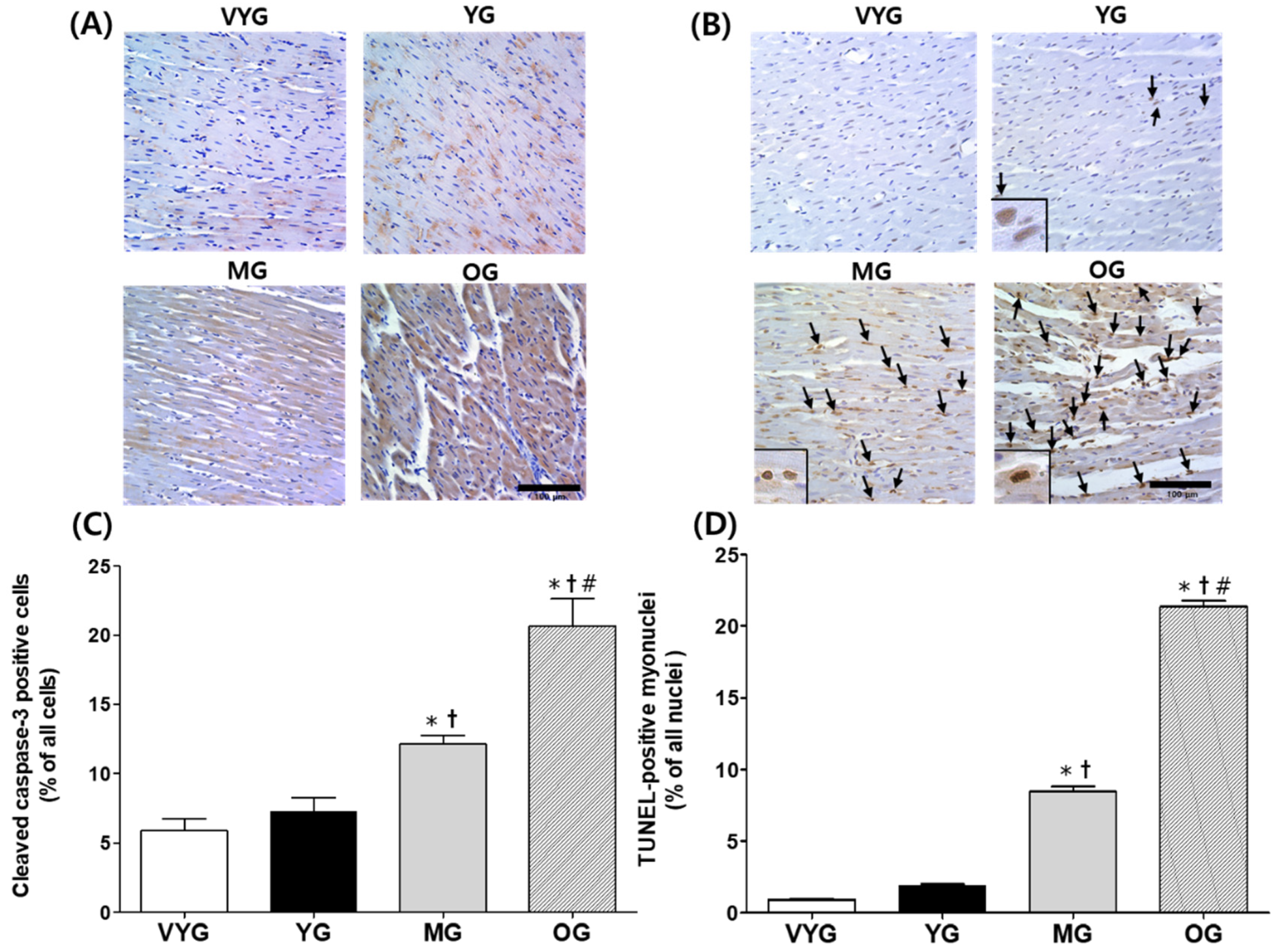
3.3. Effects of Aging on Cleaved Caspase-3-Positive Cells and TUNEL-Positive Myonuclei in Cardiac Muscles
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, Y.; Herman, B. Apoptosis and successful aging. Mech. Ageing Dev. 2002, 123, 563–565. [Google Scholar] [CrossRef]
- Nilsson, M.I.; Tarnopolsky, M.A. Mitochondria and aging-the role of exercise as a countermeasure. Biology 2019, 8, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higashi, Y.; Sukhanov, S.; Anwar, A.; Shai, S.Y.; Delafontaine, P. Aging, atherosclerosis, and IGF-1. J. Gerontol. A Biol. Sci. Med. Sci. 2012, 67, 626–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venditti, P.; Di Meo, S. The role of reactive oxygen species in the life cycle of the mitochondrion. Int. J. Mol. Sci. 2020, 21, 2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- No, M.H.; Heo, J.W.; Yoo, S.Z.; Jo, H.S.; Park, D.H.; Kang, J.H.; Seo, D.Y.; Han, J.; Kwak, H.B. Effects of aging on mitochondrial hydrogen peroxide emission and calcium retention capacity in rat heart. J. Exerc. Rehabil. 2018, 14, 920–926. [Google Scholar] [CrossRef] [PubMed]
- Kwak, H.B. Effects of aging and exercise training on apoptosis in the heart. J. Exerc. Rehabil. 2013, 9, 212–219. [Google Scholar] [CrossRef]
- Orogo, A.M.; Gustafsson, A.B. Cell death in the myocardium: My heart won’t go on. IUBMB Life 2013, 65, 651–656. [Google Scholar] [CrossRef]
- Kumar, D.; Jugdutt, B.I. Apoptosis and oxidants in the heart. J. Lab. Clin. Med. 2003, 142, 288–297. [Google Scholar] [CrossRef]
- Jose Corbalan, J.; Vatner, D.E.; Vatner, S.F. Myocardial apoptosis in heart disease: Does the emperor have clothes? Basic Res. Cardiol. 2016, 111, 31. [Google Scholar] [CrossRef]
- Martin-Fernandez, B.; Gredilla, R. Mitochondria and oxidative stress in heart aging. Age (Dordr) 2016, 38, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.Z.; No, M.H.; Heo, J.W.; Chang, E.; Park, D.H.; Kang, J.H.; Seo, D.Y.; Han, J.; Jung, S.J.; Hwangbo, K.; et al. Effects of a single bout of exercise on mitochondria-mediated apoptotic signaling in rat cardiac and skeletal muscles. J. Exerc. Rehabil. 2019, 15, 512–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollack, M.; Leeuwenburgh, C. Apoptosis and aging: Role of the mitochondria. J. Gerontol. A Biol. Sci. Med. Sci. 2001, 56, B475–B482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemura, G.; Kanoh, M.; Minatoguchi, S.; Fujiwara, H. Cardiomyocyte apoptosis in the failing heart--a critical review from definition and classification of cell death. Int. J. Cardiol. 2013, 167, 2373–2386. [Google Scholar] [CrossRef]
- Kwak, H.B.; Song, W.; Lawler, J.M. Exercise training attenuates age-induced elevation in Bax/Bcl-2 ratio, apoptosis, and remodeling in the rat heart. FASEB J. 2006, 20, 791–793. [Google Scholar] [CrossRef]
- Song, W.; Kwak, H.B.; Lawler, J.M. Exercise training attenuates age-induced changes in apoptotic signaling in rat skeletal muscle. Antioxid. Redox Signal. 2006, 8, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Azhar, G.; Gao, W.; Zhang, X.; Wei, J.Y. Bcl-2 and Bax expression in adult rat hearts after coronary occlusion: Age-associated differences. Am. J. Physiol. 1998, 275, R315–R322. [Google Scholar] [CrossRef] [PubMed]
- Nitahara, J.A.; Cheng, W.; Liu, Y.; Li, B.; Leri, A.; Li, P.; Mogul, D.; Gambert, S.R.; Kajstura, J.; Anversa, P. Intracellular calcium, DNase activity and myocyte apoptosis in aging Fischer 344 rats. J. Mol. Cell. Cardiol. 1998, 30, 519–535. [Google Scholar] [CrossRef]
- Horn, M.A.; Graham, H.K.; Richards, M.A.; Clarke, J.D.; Greensmith, D.J.; Briston, S.J.; Hall, M.C.; Dibb, K.M.; Trafford, A.W. Age-related divergent remodeling of the cardiac extracellular matrix in heart failure: Collagen accumulation in the young and loss in the aged. J. Mol. Cell. Cardiol. 2012, 53, 82–90. [Google Scholar] [CrossRef]
- Jugdutt, B.I.; Jelani, A.; Palaniyappan, A.; Idikio, H.; Uweira, R.E.; Menon, V.; Jugdutt, C.E. Aging-related early changes in markers of ventricular and matrix remodeling after reperfused ST-segment elevation myocardial infarction in the canine model: Effect of early therapy with an angiotensin II type 1 receptor blocker. Circulation 2010, 122, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Bujak, M.; Kweon, H.J.; Chatila, K.; Li, N.; Taffet, G.; Frangogiannis, N.G. Aging-related defects are associated with adverse cardiac remodeling in a mouse model of reperfused myocardial infarction. J. Am. Coll. Cardiol. 2008, 51, 1384–1392. [Google Scholar] [CrossRef] [Green Version]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef]
- Piek, A.; de Boer, R.A.; Sillje, H.H. The fibrosis-cell death axis in heart failure. Heart Fail. Rev. 2016, 21, 199–211. [Google Scholar] [CrossRef] [Green Version]
- Dai, D.F.; Chen, T.; Johnson, S.C.; Szeto, H.; Rabinovitch, P.S. Cardiac aging: From molecular mechanisms to significance in human health and disease. Antioxid. Redox Signal. 2012, 16, 1492–1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, S.; Mu, T.; Wang, G.; Jiang, X. Mitochondria-mediated apoptosis in mammals. Protein Cell 2014, 5, 737–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonsson, B. Mitochondria and the Bcl-2 family proteins in apoptosis signaling pathways. Mol. Cell. Biochem. 2004, 256–257, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Smith, M.A.; Schnellmann, R.G. Calpains, mitochondria, and apoptosis. Cardiovasc. Res. 2012, 96, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Fannin, J.; Rice, K.M.; Thulluri, S.; Arvapalli, R.K.; Wehner, P.; Blough, E.R. The effects of aging on indices of oxidative stress and apoptosis in the female fischer 344/Nnia X Brown Norway/BiNia rat heart. Open Cardiovasc. Med. J. 2013, 7, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Roh, J.; Rhee, J.; Chaudhari, V.; Rosenzweig, A. The role of exercise in cardiac aging: From physiology to molecular mechanisms. Circ. Res. 2016, 118, 279–295. [Google Scholar] [CrossRef]
- Lesnefsky, E.J.; Chen, Q.; Hoppel, C.L. Mitochondrial metabolism in aging heart. Circ. Res. 2016, 118, 1593–1611. [Google Scholar] [CrossRef] [Green Version]
- Klinge, C.M. Estrogenic control of mitochondrial function. Redox Biol. 2020, 31, 101435. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
No, M.-H.; Choi, Y.; Cho, J.; Heo, J.-W.; Cho, E.-J.; Park, D.-H.; Kang, J.-H.; Kim, C.-J.; Seo, D.Y.; Han, J.; et al. Aging Promotes Mitochondria-Mediated Apoptosis in Rat Hearts. Life 2020, 10, 178. https://0-doi-org.brum.beds.ac.uk/10.3390/life10090178
No M-H, Choi Y, Cho J, Heo J-W, Cho E-J, Park D-H, Kang J-H, Kim C-J, Seo DY, Han J, et al. Aging Promotes Mitochondria-Mediated Apoptosis in Rat Hearts. Life. 2020; 10(9):178. https://0-doi-org.brum.beds.ac.uk/10.3390/life10090178
Chicago/Turabian StyleNo, Mi-Hyun, Youngju Choi, Jinkyung Cho, Jun-Won Heo, Eun-Jeong Cho, Dong-Ho Park, Ju-Hee Kang, Chang-Ju Kim, Dae Yun Seo, Jin Han, and et al. 2020. "Aging Promotes Mitochondria-Mediated Apoptosis in Rat Hearts" Life 10, no. 9: 178. https://0-doi-org.brum.beds.ac.uk/10.3390/life10090178