Simultaneous Exposure of Cultured Human Lymphoblastic Cells to Simulated Microgravity and Radiation Increases Chromosome Aberrations

 ,
, 
 and
and
Abstract
:1. Introduction
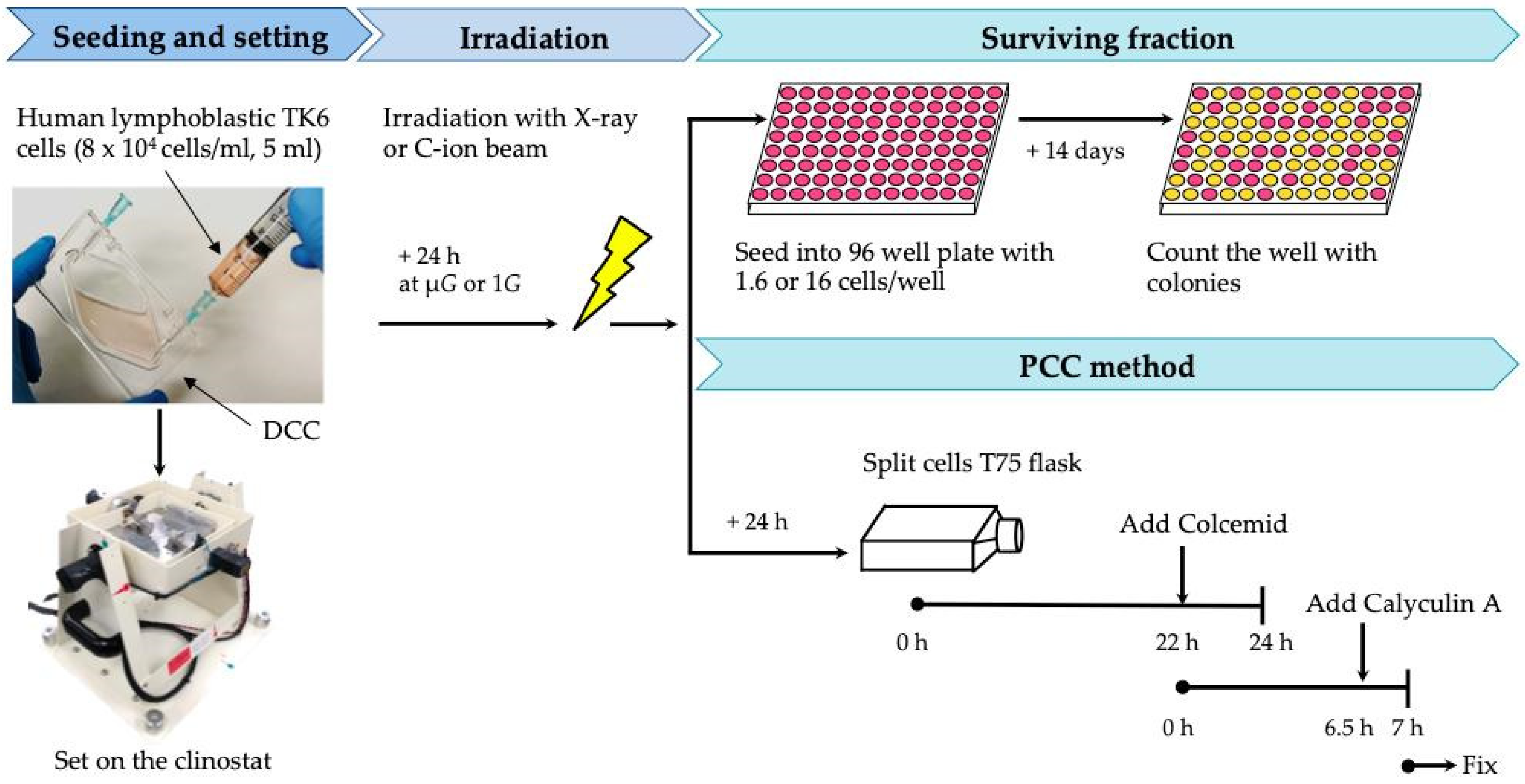
2. Materials and Methods
2.1. Cell Culture
2.2. Simultaneous Exposure of Cells to Radiation and Simulated μG
2.3. Cell Survival Fraction Analysis
2.4. Premature Chromosome Condensation (PCC)
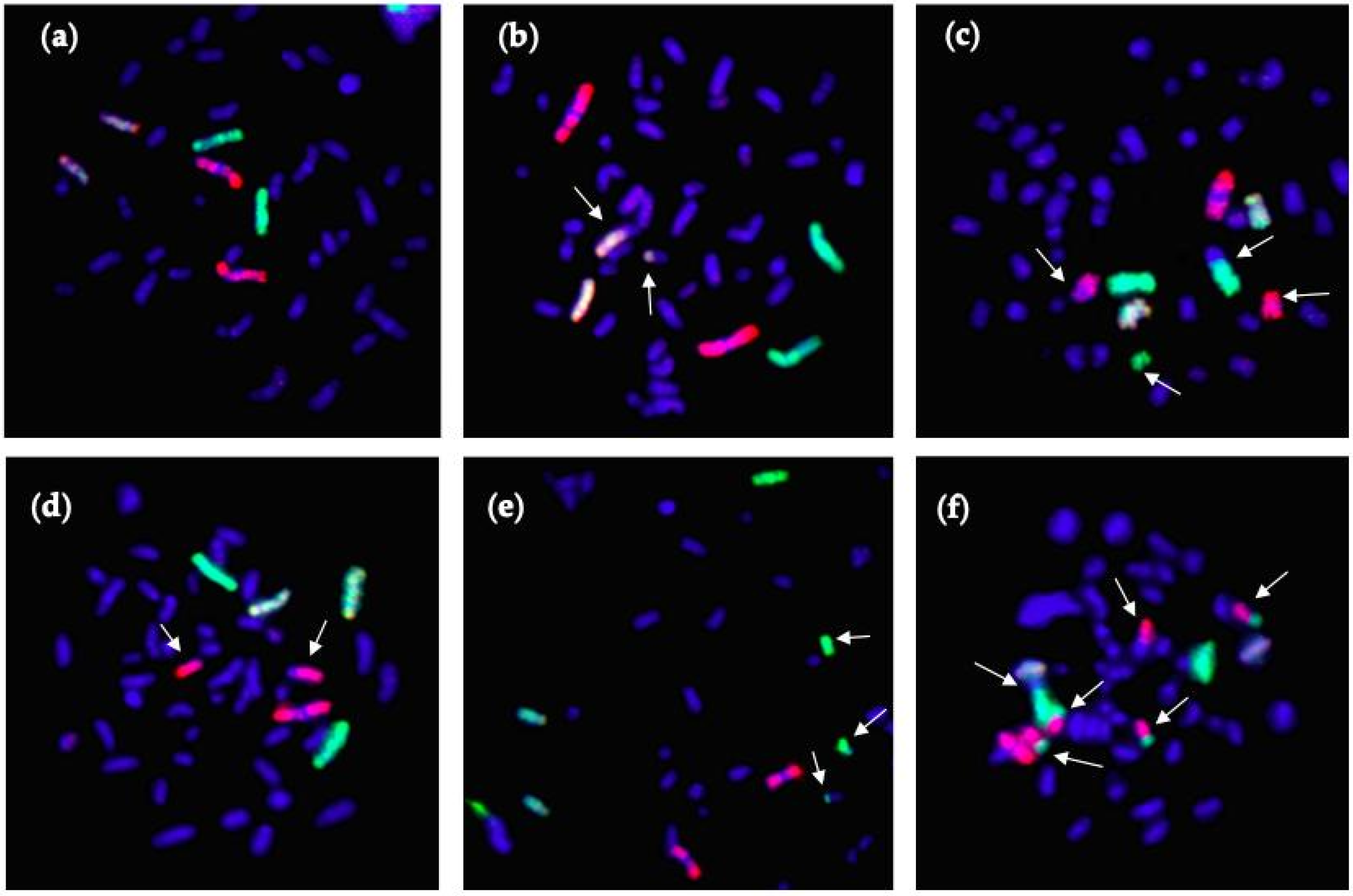
2.5. Fluorescence In Situ Hybridization (FISH)
2.6. Statistical Analysis
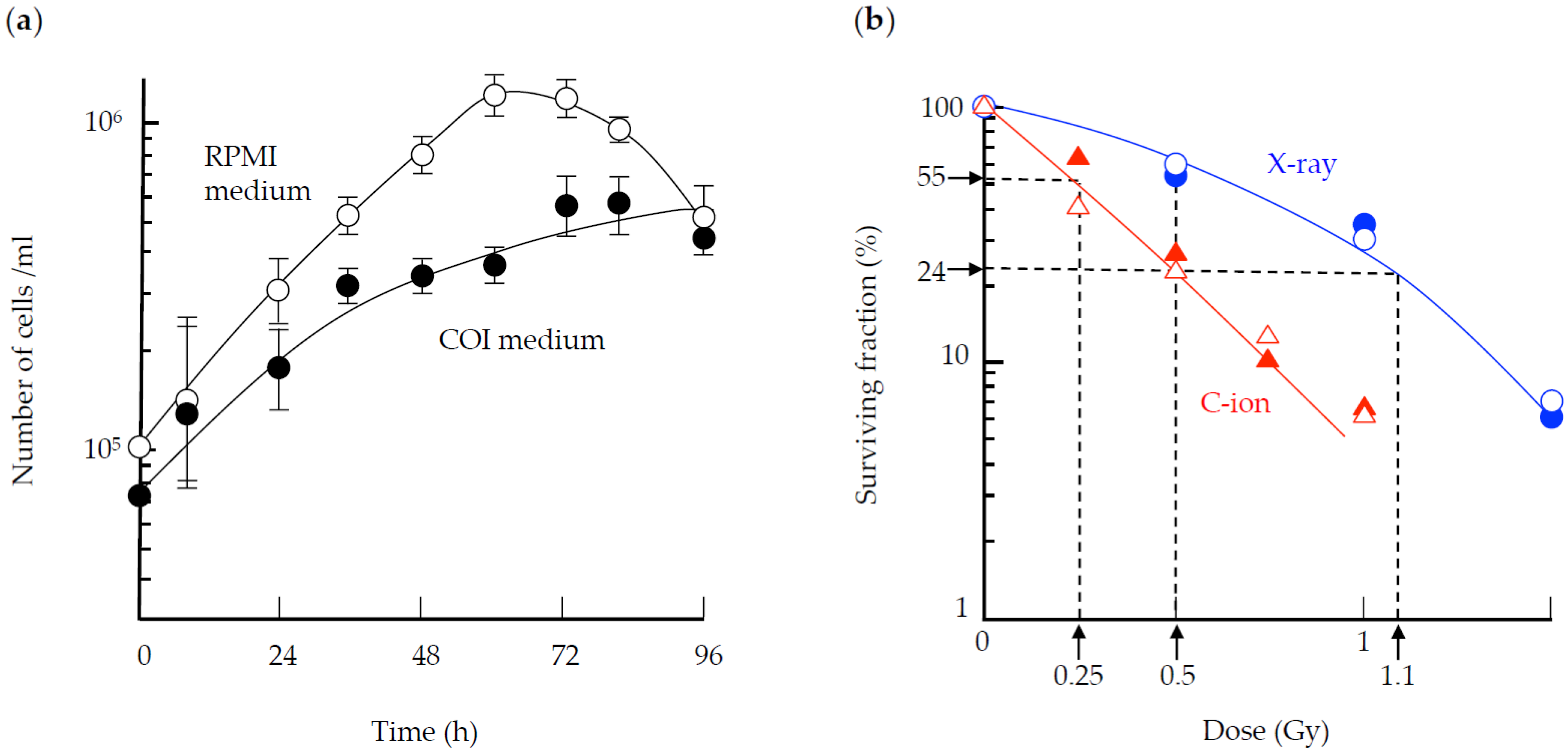
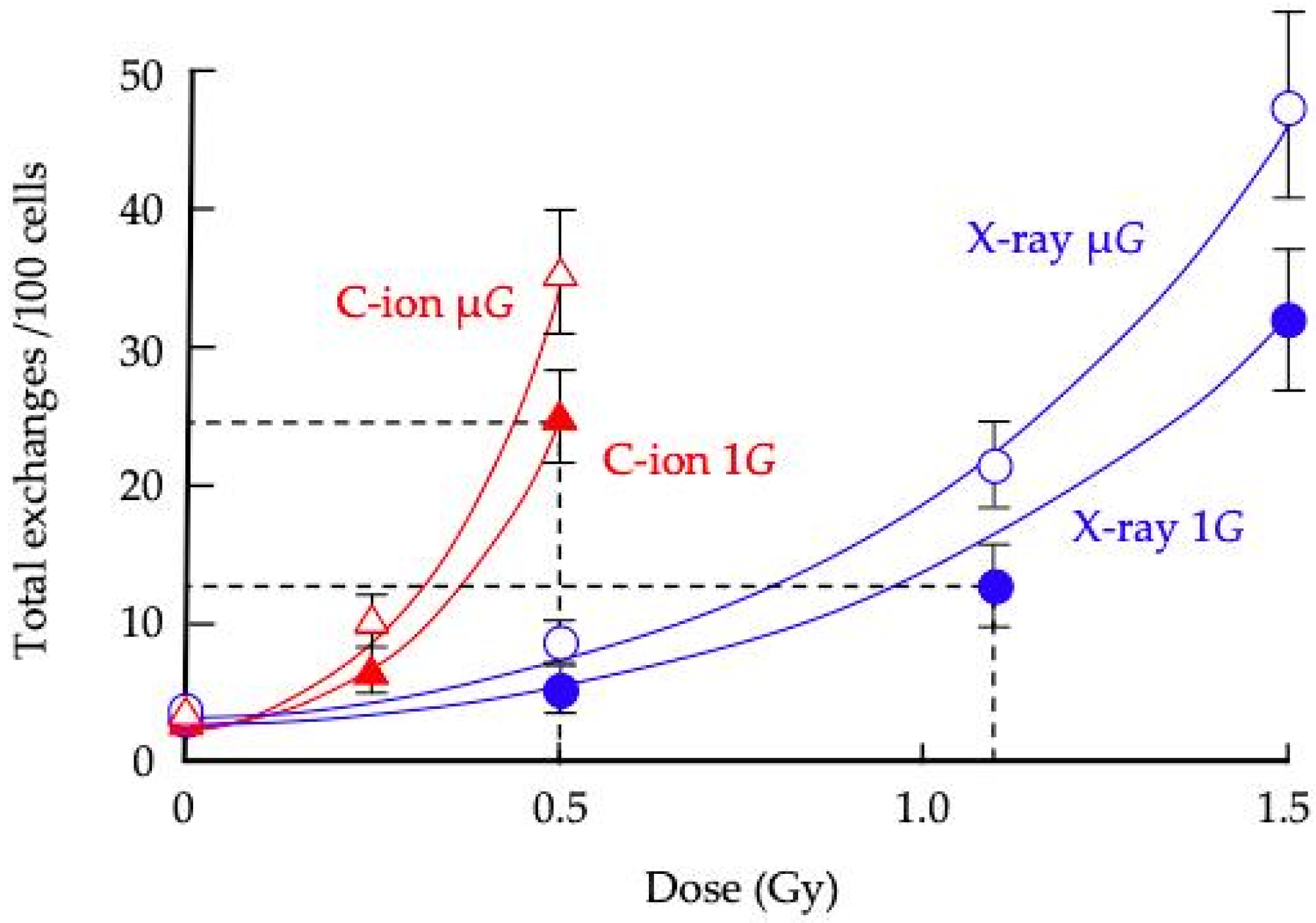
3. Results
3.1. Cell Survival
3.2. CA
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hassler, D.M.; Zeitlin, C.; Wimmer-Schweingruber, R.F.; Ehresmann, B.; Rafkin, S.; Eigenbrode, J.L.; Brinza, D.E.; Weigle, G.; Böttcher, S.; Böhm, E.; et al. Mars’ surface radiation environment measured with the Mars Science Laboratory’s Curiosity rover. Science 2014, 343, 1244797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, S.; Nagamatsu, A.; Nenoi, M.; Fujimori, A.; Kakinuma, S.; Katsube, T.; Wang, B.; Tsuruoka, C.; Shirai, T.; Nakamura, A.J.; et al. Space radiation biology for “Living in Space”. BioMed Res. Int. 2020, 2020, 4703286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, E.M.; Powell, D.R.; Li, Z.; Bell, J.S.K.; Barwick, B.G.; Feng, H.; McCrary, M.R.; Dwivedi, B.; Kowalski, J.; Dynan, W.S.; et al. Galactic cosmic radiation induces persistent epigenome alterations relevant to human lung cancer. Sci. Rep. 2018, 8, 6709. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Jella, K.K.; Jaafar, L.; Li, S.; Park, S.; Story, M.D.; Wang, H.; Wang, Y.; Dynan, W.S. Exposure to galactic cosmic radiation compromises DNA repair and increases the potential for oncogenic chromosomal rearrangement in bronchial epithelial cells. Sci. Rep. 2018, 8, 11038. [Google Scholar] [CrossRef] [PubMed]
- Tungjai, M.; Whorton, E.B.; Rithidech, K.N. Persistence of apoptosis and inflammatory responses in the heart and bone marrow of mice following whole-body exposure to ²⁸Silicon (²⁸Si) ions. Radiat. Environ. Biophys. 2013, 52, 339–350. [Google Scholar] [CrossRef]
- Yan, X.; Sasi, S.P.; Gee, H.; Lee, J.; Yang, Y.; Mehrzad, R.; Onufrak, J.; Song, J.; Enderling, H.; Agarwal, A.; et al. Cardiovascular risks associated with low dose ionizing particle radiation. PLoS ONE 2014, 9, e110269. [Google Scholar] [CrossRef] [PubMed]
- Rabin, B.M.; Poulose, S.M.; Carrihill-Knoll, K.L.; Ramirez, F.; Bielinski, D.F.; Heroux, N.; Shukitt-Hale, B. Acute effects of exposure to (56)Fe and (16)O particles on learning and memory. Radiat. Res. 2015, 184, 143–150. [Google Scholar] [CrossRef]
- Krukowski, K.; Jones, T.; Campbell-Beachler, M.; Nelson, G.; Rosi, S. Peripheral T cells as a biomarker for oxygen-ion-radiation-induced social impairments. Radiat. Res. 2018, 190, 186–193. [Google Scholar] [CrossRef]
- Pecaut, M.J.; Dutta-Roy, R.; Smith, A.L.; Jones, T.A.; Nelson, G.A.; Gridley, D.S. Acute effects of iron-particle radiation on immunity. Part I: Population distributions. Radiat. Res. 2006, 165, 68–77. [Google Scholar] [CrossRef]
- Gridley, D.S.; Obenaus, A.; Bateman, T.A.; Pecaut, M.J. Long-term changes in rat hematopoietic and other physiological systems after high-energy iron ion irradiation. Int. J. Radiat. Biol. 2008, 84, 549–559. [Google Scholar] [CrossRef]
- Akiyama, T.; Horie, K.; Hinoi, E.; Hiraiwa, M.; Kato, A.; Maekawa, Y.; Takahashi, A.; Furukawa, S. How does spaceflight affect the acquired immune system? NPJ Microgravity 2020, 6, 14. [Google Scholar] [CrossRef] [PubMed]
- Dietz, C.; Infanger, M.; Romswinkel, A.; Strube, F.; Kraus, A. Apoptosis induction and alteration of cell adherence in human lung cancer cells under simulated microgravity. Int. J. Mol. Sci. 2019, 20, 3601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merzlikina, N.V.; Buravkova, L.B.; Romanov, Y.A. The primary effects of clinorotation on cultured human mesenchymal stem cells. J. Gravit. Physiol. 2004, 11, 193–194. [Google Scholar]
- Nishikawa, M.; Ohgushi, H.; Tamai, N.; Osuga, K.; Uemura, M.; Yoshikawa, H.; Myoui, A. The effect of simulated microgravity by three-dimensional clinostat on bone tissue engineering. Cell Transplant. 2005, 14, 829–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camberos, V.; Baio, J.; Bailey, L.; Hasaniya, N.; Lopez, L.V.; Kearns-Jonker, M. Effects of spaceflight and simulated microgravity on YAP1 expression in cardiovascular progenitors: Implications for cell-based repair. Int. J. Mol. Sci. 2019, 20, 2742. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Villanueva, M.; Wong, M.; Lu, T.; Zhang, Y.; Wu, H. Interplay of space radiation and microgravity in DNA damage and DNA damage response. NPJ Microgravity 2017, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, H.; Souda, H.; Puspitasari, A.; Held, K.D.; Hidema, J.; Nikawa, T.; Yoshida, Y.; Kanai, T.; Takahashi, A. Development and performance evaluation of a three-dimensional clinostat synchronized heavy-ion irradiation system. Life Sci. Space Res. 2017, 12, 51–60. [Google Scholar] [CrossRef]
- Ikeda, H.; Souda, H.; Puspitasari, A.; Held, K.D.; Hidema, J.; Nikawa, T.; Yoshida, Y.; Kanai, T.; Takahashi, A. A new system for three-dimensional clinostat synchronized X-irradiation with a high-speed shutter for space radiation research. Biol. Sci. Space 2016, 30, 8–16. [Google Scholar] [CrossRef]
- Ikeda, H.; Muratani, M.; Hidema, J.; Hada, M.; Fujiwara, K.; Souda, H.; Yoshida, Y.; Takahashi, A. Expression profile of cell cycle-related genes in human fibroblasts exposed simultaneously to radiation and simulated microgravity. Int. J. Mol. Sci. 2019, 20, 4791. [Google Scholar] [CrossRef] [Green Version]
- Hada, M.; Ikeda, H.; Rhone, J.R.; Beitman, A.J.; Plante, I.; Souda, H.; Yoshida, Y.; Held, K.D.; Fujiwara, K.; Saganti, P.B.; et al. Increased chromosome aberrations in cells exposed simultaneously to simulated microgravity and radiation. Int. J. Mol. Sci. 2018, 20, 43. [Google Scholar] [CrossRef] [Green Version]
- Liber, H.L.; Thilly, W.G. Mutation assay at the thymidine kinase locus in diploid human lymphoblasts. Mutat. Res. 1982, 94, 467–485. [Google Scholar] [CrossRef]
- Xia, F.; Wang, X.; Wang, Y.H.; Tsang, N.M.; Yandell, D.W.; Kelsey, K.T.; Liber, H.L. Altered p53 status correlates with differences in sensitivity to radiation-induced mutation and apoptosis in two closely related human lymphoblast lines. Cancer Res. 1995, 55, 12–15. [Google Scholar] [PubMed]
- Kronenberg, A. Mutation induction in human lymphoid cells by energetic heavy ions. Adv. Space Res. 1994, 14, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Grosovsky, A.; Bethel, H.; Parks, K.; Ritter, L.; Gauny, S.; Wiese, C.; Kronenberg, A. Genomic instability in human lymphoid cells exposed to 1 GeV/amu Fe ions. Phys. Med. 2001, 17, 238–240. [Google Scholar]
- Wiese, C.; Gauny, S.; Liu, W.C.; Cherbonnel-Lasserre, C.L.; Kronenberg, A. Different mechanisms of radiation-induced loss of heterozygosity in two human lymphoid cell lines from a single donor. Cancer Res. 2001, 61, 1129–1137. [Google Scholar]
- Yudhistiara, B.; Zwicker, F.; Weber, K.J.; Huber, P.E.; Ruehle, A.; Brons, S.; Haering, P.; Debus, J.; Hauswald, S.H. The influence of a magnetic field on photon beam radiotherapy in a normal human TK6 lymphoblastoid cell line. Radiat. Oncol. 2019, 14, 11. [Google Scholar] [CrossRef]
- Harada-Sukeno, A.; Kohno, S.; Nakao, R.; Hirasaka, K.; Higashibata, A.; Yamazaki, T.; Ishioka, N.; Suzuki, H.; Shimazu, T.; Maita, A.; et al. Myo Lab: A JAXA cell biology experiment in “Kibo (JEM)” of the international space station. Biol. Sci. Space 2009, 23, 189–193. [Google Scholar] [CrossRef] [Green Version]
- Tanigawa, N.; Yano, S.; Higashibata, A.; Tsuchiya, Y.; Tanii, H.; Ando, N.; Kubota, K.; Nagase, M. Development and validation of a closed chamber for cell culture experiments in space. Biol. Sci. Space 2014, 28, 1–5. [Google Scholar] [CrossRef]
- Ohno, T.; Kanai, T.; Yamada, S.; Yusa, K.; Tashiro, M.; Shimada, H.; Torikai, K.; Yoshida, Y.; Kitada, Y.; Katoh, H.; et al. Carbon ion radio-therapy at the Gunma University Heavy Ion Medical Center: New facility set-up. Cancers 2011, 3, 4046–4060. [Google Scholar] [CrossRef]
- Takahashi, A.; Ikeda, H.; Souda, H.; Puspitasari, A.; Held, K.D.; Hidema, J.; Nikawa, T.; Yoshida, Y.; Kanai, T. Performance evaluation of new devices to synchronize heavy-ion or X-ray irradiation and simulated microgravity for space radiation research—Improvement of a control stage. In Proceedings of the 2017 International Symposium on Space Technology and Science, Matsuyama, Japan, 3–9 June 2017. [Google Scholar]
- Hada, M.; Huff, J.L.; Patel, Z.S.; Kawata, T.; Pluth, J.M.; George, K.A.; Cucinotta, F.A. AT cells are not radiosensitive for simple chromosomal exchanges at low dose. Mutat. Res. 2011, 716, 76–83. [Google Scholar] [CrossRef]
- George, K.A.; Hada, M.; Jackson, L.J.; Elliott, T.; Kawata, T.; Pluth, J.M.; Cucinotta, F.A. Dose response of γ-rays and iron nuclei induction of chromosomal aberrations in normal and repair deficient cell lines. Radiat. Res. 2009, 171, 752–763. [Google Scholar] [CrossRef] [PubMed]
- Hada, M.; Chappell, L.J.; Wang, M.; George, K.A.; Cucinotta, F.A. Induction of chromosomal aberrations at fluences of less than one HZE particle per cell nucleus. Radiat. Res. 2014, 182, 368–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hada, M.; Meador, J.A.; Cucinotta, F.A.; Gonda, S.R.; Wu, H. Chromosome aberrations induced by dual exposure of protons and iron ions. Radiat. Environ. Biophys. 2007, 46, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Luomahaara, S.; Lindholm, C.; Mustonen, R.; Salomaa, S. Distribution of radiation-induced exchange aberrations in human chromosomes 1, 2 and 4. Int. J. Radiat. Biol. 1999, 75, 1551–1556. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.N.; Poggensee, M.; Straume, T. The persistence of chromosome translocations in a radiation worker accidentally exposed to tritium. Cytogenet. Cell Genet. 1992, 60, 255–256. [Google Scholar] [CrossRef] [PubMed]
- George, K.A.; Hada, M.; Chappell, L.; Cucinotta, F.A. Biological effectiveness of accelerated particle for the induction of chromosome damage: Track structure effects. Radiat. Res. 2013, 180, 25–33. [Google Scholar] [CrossRef]
- George, K.; Hada, M.; Cucinotta, F.A. Biological effectiveness of accelerated protons for chromosome exchanges. Front. Oncol. 2015, 5, 226–232. [Google Scholar] [CrossRef] [Green Version]
- Plante, I.; Ponomarev, A.; Patel, Z.; Slaba, T.; Hada, M. RITCARD: Radiation induced tracks, chromosome aberrations, repair, and damage. Radiat. Res. 2019, 192, 282–298. [Google Scholar] [CrossRef]
- Plante, I.; Slaba, T.; Patel, Z.; Hada, M. A bi-exponential repair algorithm for radiation-induced double-strand breaks: Application to simulation of chromosome aberrations. Genes 2019, 10, 936. [Google Scholar] [CrossRef] [Green Version]
- Slaba, T.C.; Plante, I.; Ponomarev, A.; Patel, Z.S.; Hada, M. Determination of chromosome aberrations in human fibroblasts irradiated by mixed fields generated with shielding. Radiat. Res. 2020; in press. [Google Scholar]
- George, K.; Durante, M.; Wu, H.; Willingham, V.; Badhwar, G.; Cucinotta, F.A. Chromosome aberrations in the blood lymphocytes of astronauts after space flight. Radiat. Res. 2001, 156, 731–738. [Google Scholar] [CrossRef]
- George, K.; Willingham, V.; Cucinotta, F.A. Stability of chromosome aberrations in the blood lymphocytes of astronauts measured after space flight by FISH chromosome painting. Radiat. Res. 2005, 164, 474–480. [Google Scholar] [CrossRef]
- George, K.; Rhone, J.; Beitman, A.; Cucinotta, F.A. Cytogenetic damage in the blood lymphocytes of astronauts: Effects of repeat long-duration space missions. Mutat. Res. 2013, 756, 165–169. [Google Scholar] [CrossRef]
- Alpen, E.L.; Powers-Risius, P. The relative biological effect of high-Z, high-LET charged particles for spermatogonial killing. Radiat. Res. 1981, 88, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J.L.; Jordan, R.; Evans, H.H.; Lenarczyk, M.; Liber, H.L. Baseline levels of chromosome instability in the human lymphoblastoid cell TK6. Mutagenesis 2004, 6, 477–482. [Google Scholar] [CrossRef]
- Manti, L.; Durante, M.; Cirrone, G.A.P.; Grossi, G.; Lattuada, M.; Pugliese, M.; Sabini, M.G.; Scampoli, P.; Valastro, L.; Gialanella, G. Modeled microgravity does not modify the yield of chromosome aberrations induced by high-energy protons in human lymphocytes. Int. J. Radiat. Biol. 2005, 81, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Mosesso, P.; Schuber, M.; Seibt, D.; Schmitz, C.; Fiore, M.; Schinoppi, A.; Penna, S.; Palitti, F. X-ray-induced chromosome aberrations in human lymphocytes in vitro are potentiated under simulated microgravity conditions (Clinostat). Phys. Med. 2001, 17, 264–266. [Google Scholar] [PubMed]
- Ohnishi, T.; Takahashi, A.; Nagamatsu, A.; Omori, K.; Suzuki, H.; Shimazu, T.; Ishioka, N. Detection of space radiation-induced double strand breaks as a track in cell nucleus. Biochem. Biophys. Res. Commun. 2009, 18, 485–488. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Suzuki, H.; Omori, K.; Seki, M.; Hashizume, T.; Shimazu, T.; Ishioka, N.; Ohnishi, T. The expression of p53-regulated genes in human cultured lymphoblastoid TSCE5 and WTK1 cell lines during spaceflight. Int. J. Radiat. Biol. 2010, 86, 669–681. [Google Scholar] [CrossRef]
- Yagatai, F.; Honma, M.; Takahashi, A.; Omori, K.; Suzuki, H.; Shimazu, T.; Seki, M.; Hashizume, T.; Ukai, A.; Sugasawa, K.; et al. Frozen human cells can record radiation damage accumulated during space flight: Mutation induction and radioadaptation. Radiat. Environ. Biophys. 2011, 50, 125–134. [Google Scholar] [CrossRef]
- Yagatai, F.; Honma, M.; Omori, K.; Suzuki, J.; Shimazu, T.; Takahashi, A.; Ohnishi, T.; Dohmae, N.; Ishioka, N. Preliminary results of space experiment: Implications for the effects of space radiation and microgravity on survival and mutation induction in human cells. Adv. Space Res. 2012, 49, 476–486. [Google Scholar]
- Dang, B.; Yang, Y.; Zhang, E.; Li, W.; Mi, X.; Meng, Y.; Yan, S.; Wang, Z.; Wei, W.; Shao, C.; et al. Simulated microgravity increases heavy ion radiation-induced apoptosis in human B lymphoblasts. Life Sci. 2014, 97, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Mognato, M.; Celotti, L. Modeled microgravity affects cell survival and HPRT mutant frequency, but not the expression of DNA repair genes in human lymphocytes irradiated with ionising radiation. Mutat. Res. 2005, 578, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Ritter, S.; Durante, M. Heavy-ion induced chromosomal aberrations: A review. Mutat. Res. 2010, 701, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Friedland, W.; Schmitt, E.; Kundrát, P.; Dingfelder, M.; Baiocco, G.; Barbieri, S.; Ottolenghi, A. Comprehensive track-structure based evaluation of DNA damage by light ions from radiotherapy-relevant energies down to stopping. Sci. Rep. 2017, 7, 45161. [Google Scholar] [CrossRef]
- Hada, M.; Georgakilas, A.G. Formation of clustered DNA damage after high-LET irradiation: A review. J. Radiat. Res. 2008, 49, 203–210. [Google Scholar] [CrossRef]
- Kumari, R.; Singh, K.P.; Dumond, J.W., Jr. Simulated microgravity decreases DNA repair capacity and induces DNA damage in human lymphocytes. J. Cell. Biochem. 2009, 107, 723–731. [Google Scholar] [CrossRef]
- Wei, L.; Liu, C.; Kang, L.; Liu, Y.; Shi, S.; Wu, Q.; Li, Y. Experimental study on effect of simulated microgravity on structural chromosome instability of human peripheral blood lymphocytes. PLoS ONE 2014, 9, e100595. [Google Scholar] [CrossRef] [Green Version]
- Cucinotta, F.A.; Cacaoa, E.; Kim, M.H.Y.; Saganti, P.B. Benchmarking risk predictions and uncertainties in the NSCR model of GCR cancer risks with revised low LET risk coefficients. Life Sci. Space Res. 2020, 27, 64–73. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
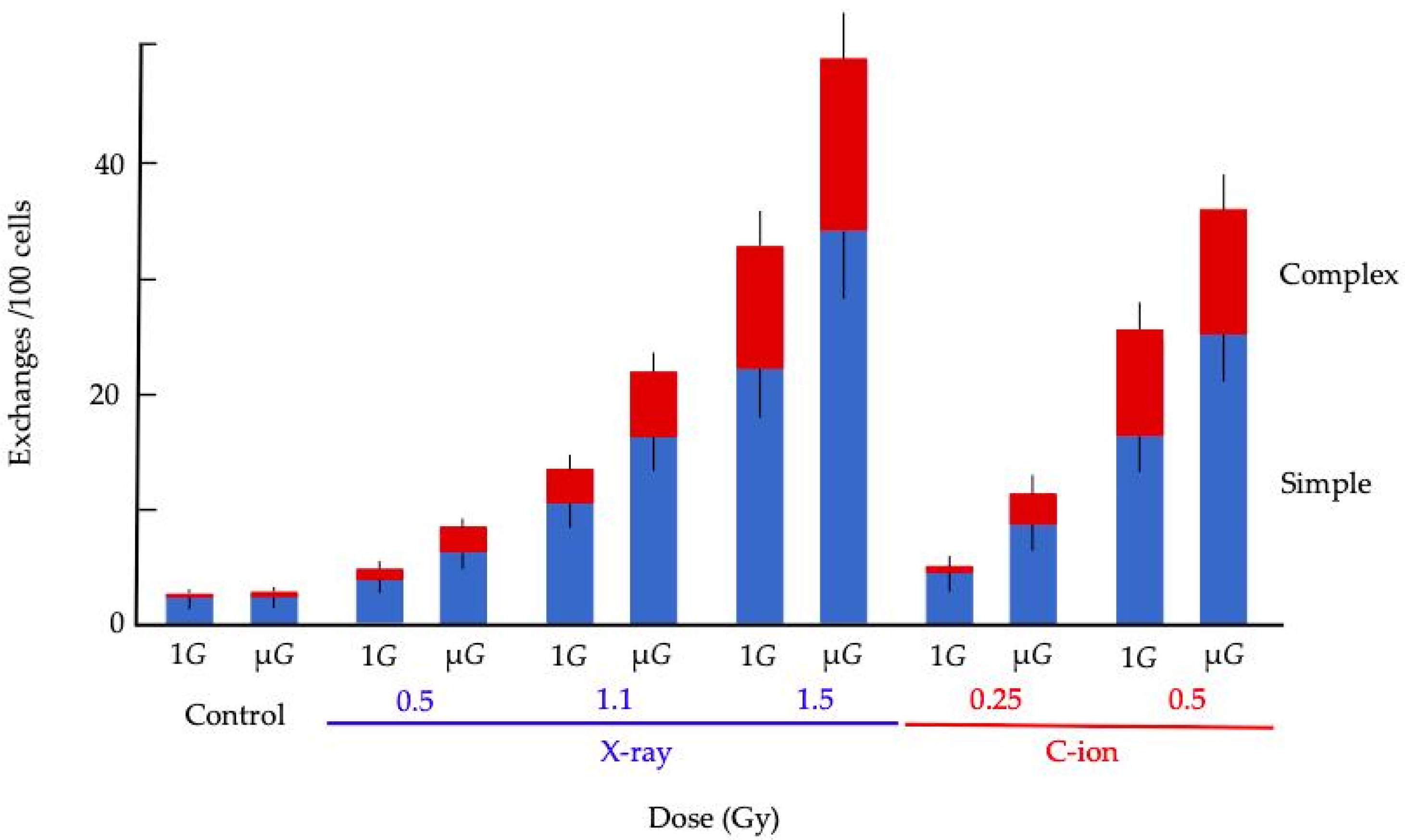{kind=link}
| Radiation | 1G or μG | Total Spreads Scored | No. of Aberrant Spreads | Simple Exchanges * | Complex Exchanges * | Total Exchanges * |
|---|---|---|---|---|---|---|
| Control (0 Gy) | 1G | 1049 | 29 | 2.13 ± 0.71 | 0.24 ± 0.24 | 2.36 ± 0.75 |
| μG | 1027 | 27 | 2.65 ± 0.88 | 0.48 ± 0.34 | 3.14 ± 0.87 | |
| (p-value) | (0.50) | (0.62) | (0.54) | |||
| X-ray | 1G | 1034 | 34 | 4.07 ± 0.99 | 0.96 ± 0.48 | 5.03 ± 1.10 |
| (0.5 Gy) | μG | 1039 | 39 | 6.44 ± 1.24 | 2.15 ± 0.72 | 8.59 ± 1.43 |
| (p-value) | (0.022) | (0.27) | (0.059) | |||
| X-ray | 1G | 670 | 50 | 10.36 ± 1.96 | 2.96 ± 1.05 | 13.31 ± 2.22 |
| (1.1 Gy) | μG | 523 | 50 | 16.11 ± 2.76 | 5.69 ± 1.64 | 21.80 ± 3.21 |
| (p-value) | (0.70) | (0.17) | (0.021) | |||
| X-ray | 1G | 330 | 50 | 23.28 ± 4.18 | 10.51 ± 2.81 | 33.04 ± 4.98 |
| (1.5 Gy) | μG | 253 | 50 | 34.28 ± 5.79 | 14.69 ± 3.79 | 48.97 ± 6.93 |
| (p-value) | (0.50) | (0.44) | (0.041) | |||
| C-ion | 1G | 1034 | 34 | 5.51 ± 1.15 | 1.20 ± 0.54 | 6.71 ± 1.27 |
| (0.25 Gy) | μG | 817 | 49 | 8.49 ± 1.60 | 1.82 ± 0.74 | 10.31 ± 1.77 |
| (p-value) | (0.83) | (0.55) | (0.091) | |||
| C-ion | 1G | 435 | 49 | 16.52 ± 3.07 | 9.11 ± 2.28 | 25.63 ± 3.82 |
| (0.5 Gy) | μG | 344 | 45 | 23.77 ± 4.14 | 10.81 ± 2.79 | 36.02 ± 5.09 |
| (p-value) | (0.074) | (0.71) | (0.079) |
| Radiation Type | Coefficient | Standard Error of Coefficient | p-Value | Odds Ratio | 95% CI for OR * | ||
|---|---|---|---|---|---|---|---|
| Lower | Upper | ||||||
| X-ray | Dose | 1.906 | 0.136 | 2.12 × 10−44 | 6.727 | 5.150 | 8.789 |
| Gravity | 0.491 | 0.132 | 2.04 × 10−4 | 1.635 | 1.261 | 2.118 | |
| Constant | −4.826 | 0.171 | 3.30 × 10−174 | ||||
| C-ion | Dose | 5.218 | 0.447 | 1.85 × 10−31 | 184.559 | 76.821 | 443.396 |
| Gravity | 0.397 | 0.156 | 1.1 × 10−2 | 1.487 | 1.096 | 2.018 | |
| Constant | −4.797 | 0.193 | 4.75 × 10−136 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamanouchi, S.; Rhone, J.; Mao, J.-H.; Fujiwara, K.; Saganti, P.B.; Takahashi, A.; Hada, M. Simultaneous Exposure of Cultured Human Lymphoblastic Cells to Simulated Microgravity and Radiation Increases Chromosome Aberrations. Life 2020, 10, 187. https://0-doi-org.brum.beds.ac.uk/10.3390/life10090187
Yamanouchi S, Rhone J, Mao J-H, Fujiwara K, Saganti PB, Takahashi A, Hada M. Simultaneous Exposure of Cultured Human Lymphoblastic Cells to Simulated Microgravity and Radiation Increases Chromosome Aberrations. Life. 2020; 10(9):187. https://0-doi-org.brum.beds.ac.uk/10.3390/life10090187
Chicago/Turabian StyleYamanouchi, Sakuya, Jordan Rhone, Jian-Hua Mao, Keigi Fujiwara, Premkumar B. Saganti, Akihisa Takahashi, and Megumi Hada. 2020. "Simultaneous Exposure of Cultured Human Lymphoblastic Cells to Simulated Microgravity and Radiation Increases Chromosome Aberrations" Life 10, no. 9: 187. https://0-doi-org.brum.beds.ac.uk/10.3390/life10090187