
Morphological Alterations and Increased S100B Expression in Epidermal Langerhans Cells Detected in Skin from Patients with Progressive Vitiligo


Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Skin Specimens
2.2. Human Fluorescent Immunohistochemistry Staining
2.3. Human Pre-Embedding Immuno-Electron Microscopy
2.4. Quantitative and Morphometric Analyses
2.5. Statistical Analysis
3. Results
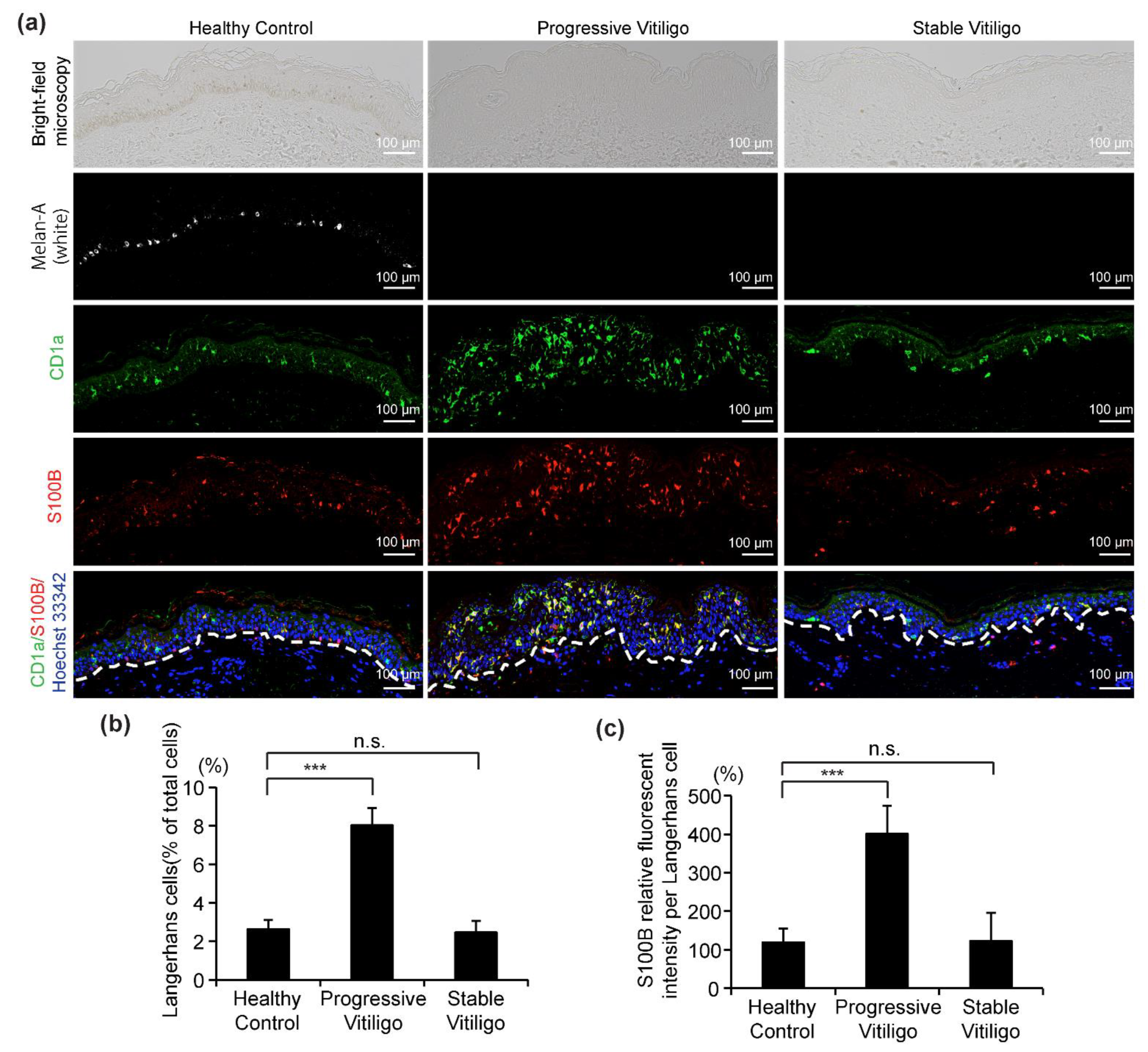
3.1. Epidermal LCs Are Increased in Lesional Skin from Progressive Vitiligo Relative to Normal Skin
3.2. Increased Number of Dendrites with Increased Length Detected in LCs of Lesional Skin from Progressive Vitiligo
3.3. Decreased BGs and Shorter BG Rods with Elevated Langerin Expression Detected in LCs of Progressive Vitiligo
3.4. Increased Expression Level of S100B in Epidermal LCs from Progressive Vitiligo
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nestle, F.O.; Di Meglio, P.; Qin, J.Z.; Nickoloff, B.J. Skin immune sentinels in health and disease. Nat. Rev. Immunol. 2009, 9, 679–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taieb, A.; Picardo, M. Clinical practice. Vitiligo. N. Engl. J. Med. 2009, 360, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Ezzedine, K.; Eleftheriadou, V.; Whitton, M.; van Geel, N. Vitiligo. Lancet 2015, 386, 74–84. [Google Scholar] [CrossRef]
- Nordlund, J.J. Vitiligo: A review of some facts lesser known about depigmentation. Indian J. Dermatol. 2011, 56, 180–189. [Google Scholar] [CrossRef]
- Katz, E.L.; Harris, J.E. Translational Research in Vitiligo. Front. Immunol. 2021, 12, 624517. [Google Scholar] [CrossRef]
- Hatchome, N.; Aiba, S.; Kato, T.; Torinuki, W.; Tagami, H. Possible functional impairment of Langerhans’ cells in vitiliginous skin. Reduced ability to elicit dinitrochlorobenzene contact sensitivity reaction and decreased stimulatory effect in the allogeneic mixed skin cell lymphocyte culture reaction. Arch. Dermatol. 1987, 123, 51–54. [Google Scholar] [CrossRef]
- Uehara, M.; Miyauchi, H.; Tanaka, S. Diminished contact sensitivity response in vitiliginous skin. Arch. Dermatol. 1984, 120, 195–198. [Google Scholar] [CrossRef]
- Breit, R. The Langerhans cell: The master key to contact dermatitis: A hypothesis. Arch. Dermatol. Res. 1982, 272, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Silberberg, I.; Baer, R.L.; Rosenthal, S.A. The role of Langerhans cells in allergic contact hypersensitivity. A review of findings in man and guinea pigs. J. Investig. Dermatol. 1976, 66, 210–217. [Google Scholar] [CrossRef] [Green Version]
- Nordlund, J.J. The melanocyte and the epidermal melanin unit: An expanded concept. Dermatol. Clin. 2007, 25, 271–281. [Google Scholar] [CrossRef]
- Itoi, S.; Tanemura, A.; Kotobuki, Y.; Wataya-Kaneda, M.; Tsuruta, D.; Ishii, M.; Katayama, I. Coexistence of Langerhans cells activation and immune cells infiltration in progressive nonsegmental vitiligo. J. Dermatol. Sci. 2014, 73, 83–85. [Google Scholar] [CrossRef]
- Prignano, F.; Ricceri, F.; Bianchi, B.; Guasti, D.; Bonciolini, V.; Lotti, T.; Pimpinelli, N. Dendritic cells: Ultrastructural and immunophenotypical changes upon nb-UVB in vitiligo skin. Arch. Dermatol. Res. 2011, 303, 231–238. [Google Scholar] [CrossRef]
- Shoeib, M.; Farag, A.G.A.; Eltorgman, M.; Abdou, A.G. Evaluation of the Langerhans cells role in vitiligo and its relationship to NB-UVB. J. Cosmet. Dermatol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Cocchia, D.; Michetti, F.; Donato, R. Immunochemical and Immuno-Cytochemical Localization of S-100 Antigen in Normal Human-Skin. Nature 1981, 294, 85–87. [Google Scholar] [CrossRef]
- Hofmann, M.A.; Drury, S.; Fu, C.; Qu, W.; Taguchi, A.; Lu, Y.; Avila, C.; Kambham, N.; Bierhaus, A.; Nawroth, P.; et al. RAGE mediates a novel proinflammatory axis: A central cell surface receptor for S100/calgranulin polypeptides. Cell 1999, 97, 889–901. [Google Scholar] [CrossRef] [Green Version]
- Michetti, F.; Corvino, V.; Geloso, M.C.; Lattanzi, W.; Bernardini, C.; Serpero, L.; Gazzolo, D. The S100B protein in biological fluids: More than a lifelong biomarker of brain distress. J. Neurochem. 2012, 120, 644–659. [Google Scholar] [CrossRef] [PubMed]
- Paradisi, A.; Guidi, B.; Diociaiuti, A.; Forni, F.; Scribano, D.; Sisto, T.; Amerio, P.L.; Capizzi, R. Increased S100B protein serum levels in psoriasis. J. Dermatol. Sci. 2007, 48, 148–150. [Google Scholar] [CrossRef] [PubMed]
- Birlea, S.A. S100B: Correlation with Active Vitiligo Depigmentation. J. Investig. Dermatol. 2017, 137, 1408–1410. [Google Scholar] [CrossRef] [Green Version]
- Cheong, K.A.; Noh, M.; Kim, C.H.; Lee, A.Y. S100B as a potential biomarker for the detection of cytotoxicity of melanocytes. Exp. Dermatol. 2014, 23, 165–171. [Google Scholar] [CrossRef]
- Badran, A.Y.; Gomaa, A.S.; El-Mahdy, R.I.; El Zohne, R.A.; Kamal, D.T.; Abou-Taleb, D.A.E. Serum level of S100B in vitiligo patients: Is it a marker of disease activity? Australas J. Dermatol. 2021, 62, e67–e72. [Google Scholar] [CrossRef]
- Shabaka, F.H.; Rashed, L.A.; Said, M.; Ibrahim, L. Sensitivity of serum S100B protein as a disease activity marker in Egyptian patients with vitiligo (case-control study). Arch. Physiol. Biochem. 2020, 1–8. [Google Scholar] [CrossRef]
- Yamamoto, A.; Yang, L.; Kuroda, Y.; Guo, J.; Teng, L.; Tsuruta, D.; Katayama, I. Local Epidermal Endocrine Estrogen Protects Human Melanocytes against Oxidative Stress, a Novel Insight into Vitiligo Pathology. Int. J. Mol. Sci. 2020, 22, 269. [Google Scholar] [CrossRef] [PubMed]
- Hayashi-Nishino, M.; Fujita, N.; Noda, T.; Yamaguchi, A.; Yoshimori, T.; Yamamoto, A. A subdomain of the endoplasmic reticulum forms a cradle for autophagosome formation. Nat. Cell Biol. 2009, 11, 1433–1437. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Matsunaga, K.; Suzuki, K.; Ito, A.; Tanemura, A.; Abe, Y.; Suzuki, T.; Yoshikawa, M.; Sumikawa, Y.; Yagami, A.; Masui, Y.; et al. Rhododendrol-induced leukoderma update I: Clinical findings and treatment. J. Dermatol. 2021. [Google Scholar] [CrossRef]
- Inoue, S.; Katayama, I.; Suzuki, T.; Tanemura, A.; Ito, S.; Abe, Y.; Sumikawa, Y.; Yoshikawa, M.; Suzuki, K.; Yagami, A.; et al. Rhododendrol-induced leukoderma update II: Pathophysiology, mechanisms, risk evaluation, and possible mechanism-based treatments in comparison with vitiligo. J. Dermatol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, T.; Hauser, C.; Budtz-Jorgensen, E. Langerhans cells: Structure, function and role in oral pathological conditions. J. Oral. Pathol. Med. 1993, 22, 193–202. [Google Scholar] [CrossRef]
- Speeckaert, R.; Voet, S.; Hoste, E.; van Geel, N. S100B Is a Potential Disease Activity Marker in Nonsegmental Vitiligo. J. Investig. Dermatol. 2017, 137, 1445–1453. [Google Scholar] [CrossRef] [Green Version]
- Seite, S.; Zucchi, H.; Moyal, D.; Tison, S.; Compan, D.; Christiaens, F.; Gueniche, A.; Fourtanier, A. Alterations in human epidermal Langerhans cells by ultraviolet radiation: Quantitative and morphological study. Br. J. Dermatol. 2003, 148, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Westerhof, W.; d’Ischia, M. Vitiligo puzzle: The pieces fall in place. Pigment Cell Res. 2007, 20, 345–359. [Google Scholar] [CrossRef] [PubMed]
- Langerhans, P. die Nerven der menschlichen Haut. Virchows Arch. 1868, 44, 325–327. [Google Scholar] [CrossRef] [Green Version]
- Merad, M.; Ginhoux, F.; Collin, M. Origin, homeostasis and function of Langerhans cells and other langerin-expressing dendritic cells. Nat. Rev. Immunol. 2008, 8, 935–947. [Google Scholar] [CrossRef] [PubMed]
- Alcalay, J.; Goldberg, L.H.; Wolf, J.E., Jr.; Kripke, M.L. Variations in the number and morphology of Langerhans’ cells in the epidermal component of squamous cell carcinomas. Arch. Dermatol. 1989, 125, 917–920. [Google Scholar] [CrossRef] [PubMed]
- Bergfelt, L.; Larko, O.; Lindberg, M. Density and morphology of Langerhans cells in basal cell carcinomas of the face and trunk. Br. J. Dermatol. 1992, 127, 575–579. [Google Scholar] [CrossRef]
- Bergfelt, L. Langerhans cells, immunomodulation and skin lesions. A quantitative, morphological and clinical study. Acta Derm. Venereol. Suppl. 1993, 180, 1–37. [Google Scholar]
- Bergfelt, L.; Emilson, A.; Lindberg, M.; Scheynius, A. Quantitative and 3-dimensional analysis of Langerhans cells in basal cell carcinoma. A comparative study using light microscopy and confocal laser scanning microscopy. Br. J. Dermatol. 1994, 130, 273–280. [Google Scholar] [CrossRef]
- Romani, N.; Clausen, B.E.; Stoitzner, P. Langerhans cells and more: Langerin-expressing dendritic cell subsets in the skin. Immunol. Rev. 2010, 234, 120–141. [Google Scholar] [CrossRef] [Green Version]
- Arrighi, J.F.; Soulas, C.; Hauser, C.; Saeland, S.; Chapuis, B.; Zubler, R.H.; Kindler, V. TNF-alpha induces the generation of Langerin/(CD207)+ immature Langerhans-type dendritic cells from both CD14-CD1a and CD14+CD1a- precursors derived from CD34+ cord blood cells. Eur. J. Immunol. 2003, 33, 2053–2063. [Google Scholar] [CrossRef] [PubMed]
- Takahara, K.; Omatsu, Y.; Yashima, Y.; Maeda, Y.; Tanaka, S.; Iyoda, T.; Clausen, B.E.; Matsubara, K.; Letterio, J.; Steinman, R.M.; et al. Identification and expression of mouse Langerin (CD207) in dendritic cells. Int. Immunol. 2002, 14, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Kanitakis, J.; Zambruno, G.; Viac, J.; Panzini, H.; Thivolet, J. Expression of Neural-Tissue Markers (S-100 Protein and Leu-7 Antigen) by Sweat Gland Tumors of the Skin—An Immunohistochemical Study. J. Am. Acad. Dermatol. 1987, 17, 187–191. [Google Scholar] [CrossRef]
- Gonzalez, L.L.; Garrie, K.; Turner, M.D. Role of S100 proteins in health and disease. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118677. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient No. | Sex | Age (y) | Duration (y) | Type | Lesion Site | Activity |
|---|---|---|---|---|---|---|
| 1 | F | 49 | 2 | Non-segmental vitiligo | Back of hand | Stable |
| 2 | M | 79 | 7 | Non-segmental vitiligo | Forehead | Progressive |
| 3 | F | 69 | 1 | RDIL * | Forehead | Progressive |
| 4 | F | 55 | 10 | Non-segmental vitiligo | Abdomen | Progressive |
| 5 | F | 66 | 0.4 | RDIL | Back of hand | Progressive |
| 6 | F | 63 | 4 | Non-segmental vitiligo | Lumbar | Stable |
| 7 | F | 74 | 0.6 | RDIL | Forehead | Progressive |
| 8 | F | 63 | 1 | RDIL | Abdomen | Progressive |
| 9 | F | 38 | 0.5 | RDIL | Neck | Progressive |
| 10 | F | 59 | 0.8 | RDIL | Back of hand | Progressive |
| 11 | M | 80 | 0.5 | Non-segmental vitiligo | Forehead | Progressive |
| 12 | F | 70 | 6 | Non-segmental vitiligo | Chin | Stable |
| 13 | F | 43 | 3 | Non-segmental vitiligo | Neck | Stable |
| 14 | F | 66 | 8 | Non-segmental vitiligo | Chest | Stable |
| 15 | F | 59 | 3 | Non-segmental vitiligo | Arm | Stable |
| 16 | F | 60 | 4 | Non-segmental vitiligo | Leg | Progressive |
| 17 | F | 64 | 5 | Non-segmental vitiligo | Shoulder | Stable |
| 18 | M | 15 | 1 | Non-segmental vitiligo | Face | Progressive |
| 19 | F | 84 | 1 | RDIL | Neck | Progressive |
| 20 | M | 35 | 5 | Non-segmental vitiligo | Chest | Progressive |
| 21 | F | 42 | 0.3 | RDIL | Face | Progressive |
| 22 | F | 48 | 0.5 | RDIL | Neck | Progressive |
| 23 | M | 48 | 4 | Non-segmental vitiligo | Face | Stable |
| 24 | M | 58 | 3 | Non-segmental vitiligo | Wrist | Stable |
| 25 | M | 36 | 5 | Non-segmental vitiligo | Abdomen | Progressive |
| 26 | F | 35 | 0.5 | RDIL | Back of hand | Progressive |
| 27 | F | 71 | 1 | RDIL | Neck | Progressive |
| 28 | F | 67 | 0.3 | RDIL | Forehead | Progressive |
| 29 | F | 38 | 3 | Non-segmental vitiligo | Forehead | Progressive |
| 30 | F | 66 | 1 | RDIL | Abdomen | Progressive |
| 31 | F | 77 | 3 | Non-segmental vitiligo | Back of hand | Stable |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, F.; Yang, L.; Teng, L.; Zhang, H.; Katayama, I. Morphological Alterations and Increased S100B Expression in Epidermal Langerhans Cells Detected in Skin from Patients with Progressive Vitiligo. Life 2021, 11, 579. https://0-doi-org.brum.beds.ac.uk/10.3390/life11060579
Yang F, Yang L, Teng L, Zhang H, Katayama I. Morphological Alterations and Increased S100B Expression in Epidermal Langerhans Cells Detected in Skin from Patients with Progressive Vitiligo. Life. 2021; 11(6):579. https://0-doi-org.brum.beds.ac.uk/10.3390/life11060579
Chicago/Turabian StyleYang, Fei, Lingli Yang, Lanting Teng, Huimin Zhang, and Ichiro Katayama. 2021. "Morphological Alterations and Increased S100B Expression in Epidermal Langerhans Cells Detected in Skin from Patients with Progressive Vitiligo" Life 11, no. 6: 579. https://0-doi-org.brum.beds.ac.uk/10.3390/life11060579