
Loss of Group II Metabotropic Glutamate Receptor Signaling Exacerbates Hypertension in Spontaneously Hypertensive Rats



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Dorsal Hindbrain mGluR2/3 Treatment
2.3. Measurement of BP and HR
2.4. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (PCR)
2.5. Echocardiographic and Renal Ultrasonographic Measurements
2.6. Measurement of Catecholamine Concentration
2.7. Implantation of Telemetry Device for Electrocardiography Recording
2.8. HRV Analysis
2.9. Assessment of Baroreflex Sensitivity
2.10. Statistical Analysis
3. Results
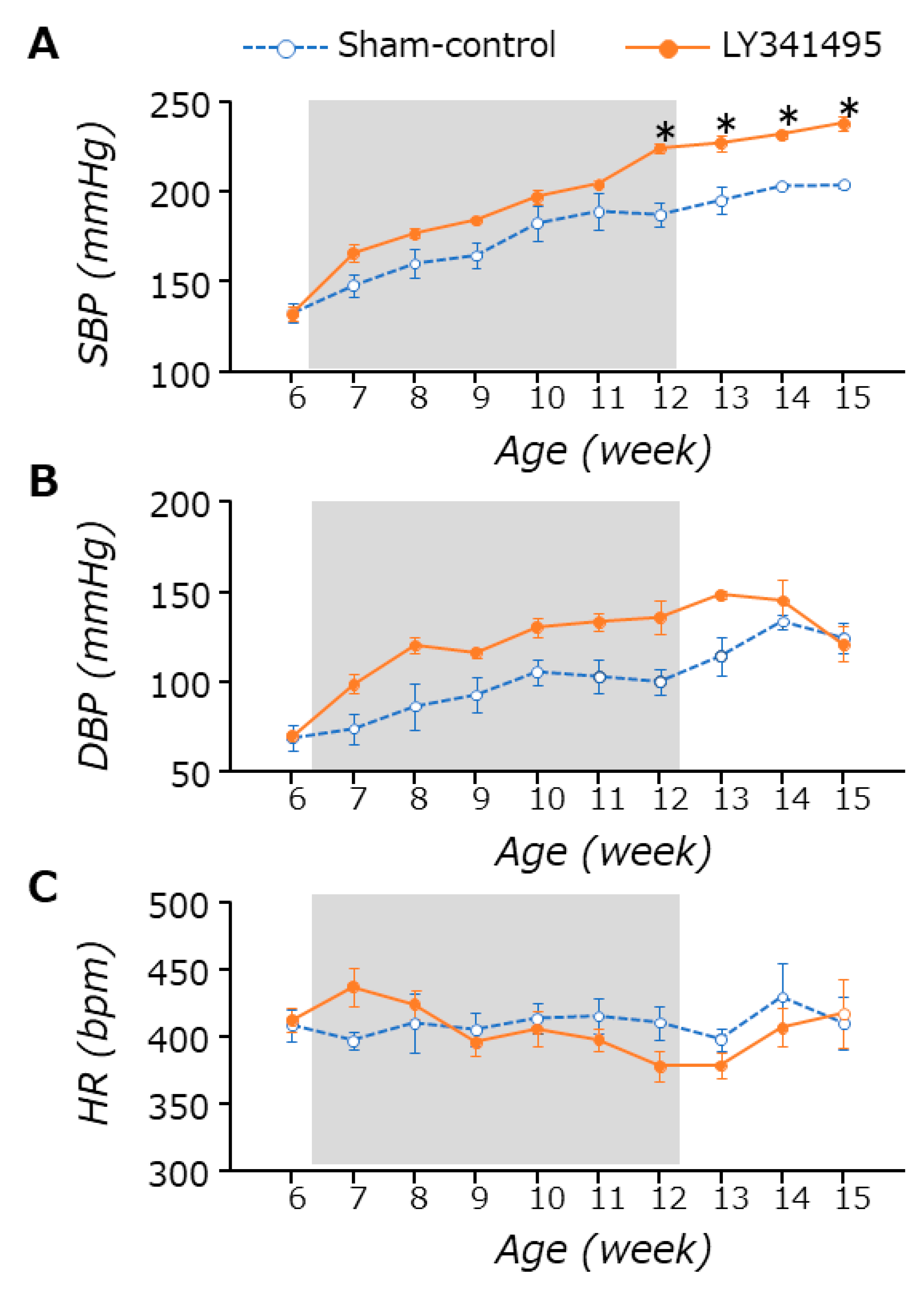
3.1. BP and HR Changes
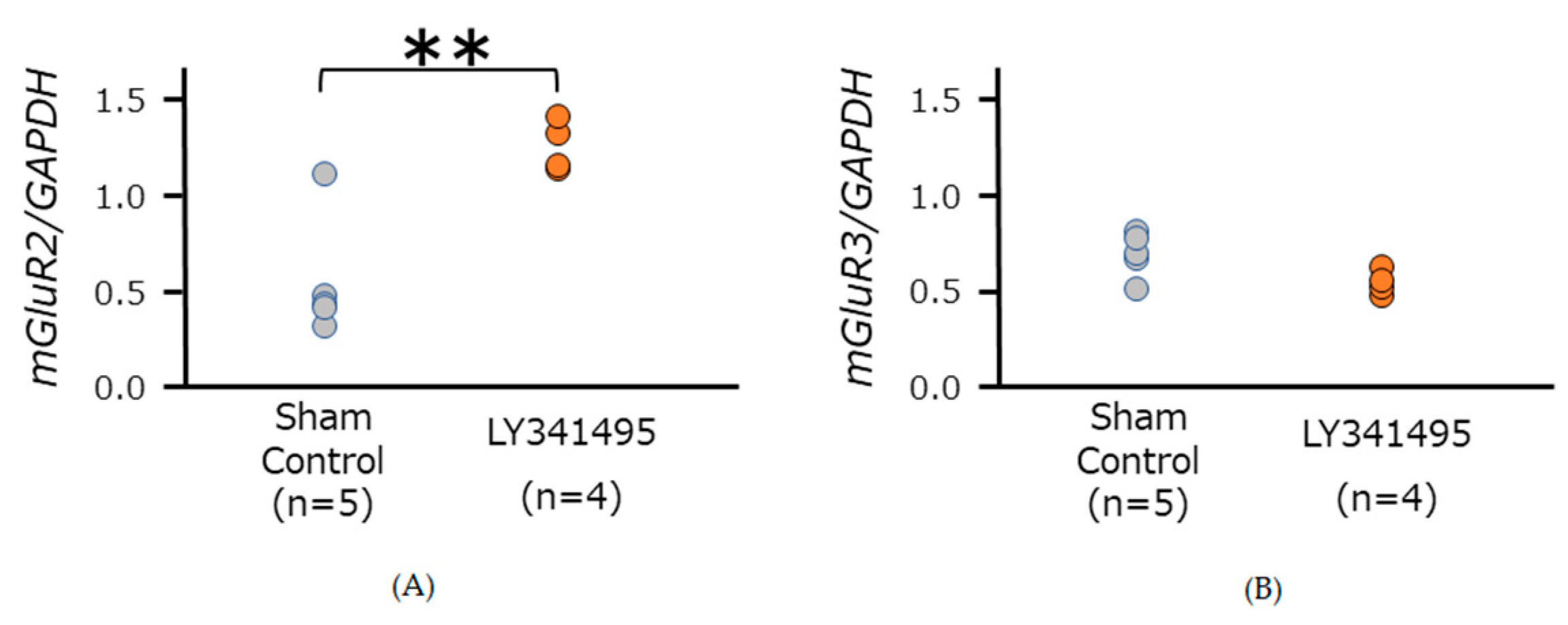
3.2. mGluR2/3 Expression in Medulla Oblongata in SHR
3.3. Echocardiography and Renal Ultrasonography
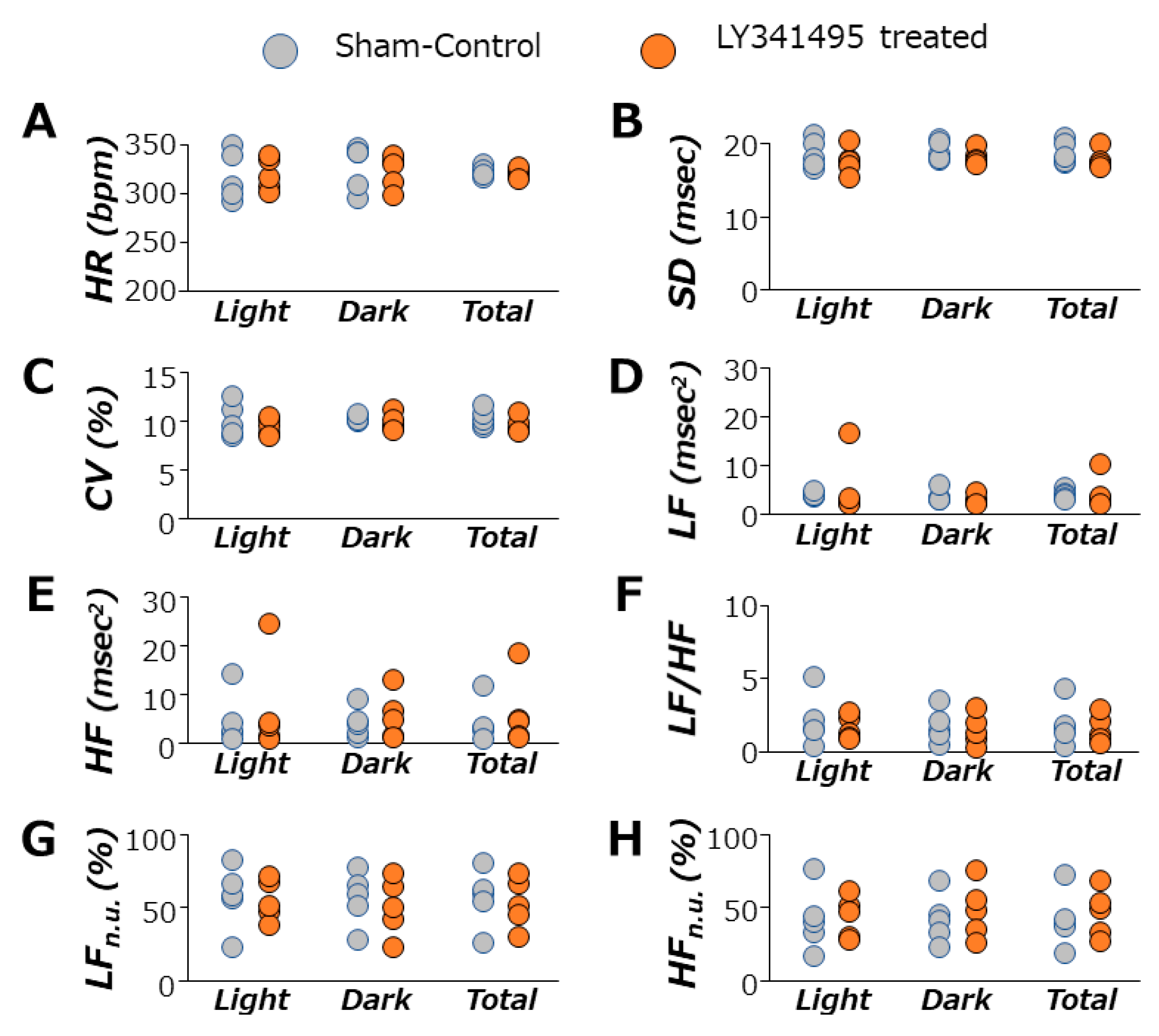
3.4. Autonomic Nervous System Function during mGluR2/3 Antagonist Treatment in SHRs
3.5. Catecholamine Blood Test in SHRs Treated with LY341495
3.6. Effects of Dorsal Hindbrain Treatment with mGluR2/3 Antagonist on Baroreflex Function
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- World Health Organization. A Global Brief on Hypertension: Silent Killer, Global Public Health Crisis; World Health Organization: Geneva, Switzerland, 2013. WHO Reference Number: WHO/DCO/WHD/2013.2. Available online: https://apps.who.int/iris/rest/bitstreams/195800/retrieve (accessed on 18 July 2021).
- Whelton, P.K.; Carey, R.M.; Aronow, W.S.; Casey, D.E., Jr.; Collins, K.J.; Dennison, H.C.; De Palma, S.M.; Gidding, S.; Jamerson, K.A.; Jones, D.W.; et al. 2017 ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA Guideline for the Prevention, Detection, Evaluation, and Management of High Blood Pressure in Adults: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Hypertension 2018, 71, 1269–1324. [Google Scholar] [PubMed]
- Colombari, E.; Sato, M.A.; Cravo, S.L.; Bergamaschi, C.T.; Campos, R.R., Jr.; Lopes, O.U. Role of the medulla oblongata in hypertension. Hypertension 2001, 38 Pt 2, 549–554. [Google Scholar] [CrossRef] [Green Version]
- Ernsberger, P.; Meeley, M.P.; Mann, J.J.; Reis, D.J. Clonidine binds to imidazole binding sites as well as a2-adrenoceptors in the ventrolateral medulla. Eur. J. Pharmacol. 1987, 134, 1–13. [Google Scholar] [CrossRef]
- McCorry, L.K. Physiology of the autonomic nervous system. Am. J. Pharm. Educ. 2007, 71, 78. [Google Scholar] [CrossRef] [Green Version]
- Furlan, R.; Heusser, K.; Minonzio, M.; Shiffer, D.; Cairo, B.; Tank, J.; Jordan, J.; Diedrich, A.; Gauger, P.; Zamuner, A.R.; et al. Cardiac and vascular sympathetic baroreflex control during orthostatic pre-syncope. J. Clin. Med. 2019, 8, 1434. [Google Scholar] [CrossRef] [Green Version]
- Bristow, J.D.; Honour, A.J.; Pickering, G.W.; Sleight, P.; Smyth, H.S. Diminished baroreflex sensitivity in high blood pressure. Circulation 1969, 39, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Laterza, M.C.; de Matos, L.D.N.J.; Trombetta, I.C.; Braga, A.M.W.; Roveda, F.; Alves, M.J.N.N.; Krieger, E.M.; Negrão, C.E.; Rondon, M.U.P.B. Exercise training restores baroreflex sensitivity in never-treated hypertensive patients. Hypertension 2007, 49, 1298–1306. [Google Scholar] [CrossRef] [Green Version]
- Gordon, F.J.; Sved, A.F. Neurotransmitters in central cardiovascular regulation: Glutamate and GABA. Clin. Exp. Pharmacol. Physiol. 2002, 29, 522–524. [Google Scholar] [CrossRef]
- Mandel, D.A.; Schreihofer, A.M. Glutamatergic inputs to the CVLM independent of the NTS promote tonic inhibition of sympathetic vasomotor tone in rats. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1772–H1779. [Google Scholar] [CrossRef]
- Sekizawa, S.; Bonham, A.C. Group I metabotropic glutamate receptors on second-order baroreceptor neurons are tonically activated and induce a Na+-Ca2+ exchange current. J. Neurophysiol. 2006, 95, 882–892. [Google Scholar] [CrossRef]
- Martinez, D.; Rogers, R.C.; Hermann, G.E.; Hasser, E.M.; Kline, D.D. Astrocytic glutamate transporters reduce the neuronal and physiological influence of metabotropic glutamate receptors in nucleus tractus solitarii. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020, 318, R545–R564. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, S. Molecular diversity of glutamate receptors and implications for brain function. Science 1992, 258, 597–603. [Google Scholar] [CrossRef]
- Chen, C.Y.; Ling, E.H.; Horowitz, J.M.; Bonham, A.C. Synaptic transmission in nucleus tractus solitarius is depressed by Group II and III but not Group I presynaptic metabotropic glutamate receptors in rats. J. Physiol. 2002, 538 Pt 3, 773–786. [Google Scholar] [CrossRef]
- Sekizawa, S.; Bechtold, A.G.; Tham, R.C.; Bonham, A.C. A novel postsynaptic group II metabotropic glutamate receptor role in modulating baroreceptor signal transmission. J. Neurosci. 2009, 29, 11807–11816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, V.R.; Machado, B.H. Antagonism of glutamatergic metabotropic receptors in the NTS of awake rats does not affect the gain of the baroreflex. Auton. Neurosci. 2003, 103, 65–71. [Google Scholar] [CrossRef]
- Matsumura, K.; Tsuchihashi, T.; Kagiyama, S.; Abe, I.; Fujishima, M. Subtypes of metabotropic glutamate receptors in the nucleus of the solitary tract of rats. Brain Res. 1999, 842, 461–468. [Google Scholar] [CrossRef]
- Viard, E.; Sapru, H.N. Cardiovascular responses to activation of metabotropic glutamate receptors in the nTS of the rat. Brain Res. 2002, 952, 308–321. [Google Scholar] [CrossRef]
- Pinto, Y.M.; Paul, M.; Ganten, D. Lessons from rat models of hypertension: From Goldblatt to genetic engineering. Cardiovasc. Res. 1998, 39, 77–88. [Google Scholar] [CrossRef]
- Anishchenko, A.M.; Aliev, O.I.; Sidekhmenova, A.V.; Shamanaev, A.Y.; Plotnikov, M.B. Dynamics of blood pressure elevation and endothelial dysfunction in SHR rats during the development of arterial hypertension. Bull. Exp. Biol. Med. 2015, 159, 591–593. [Google Scholar] [CrossRef]
- Okamoto, K.; Aoki, K. Development of a strain of spontaneously hypertensive rats. Jpn. Circ. J. 1963, 27, 282–293. [Google Scholar] [CrossRef]
- Hsu, J.C.N.; Sekizawa, S.; Tochinai, R.; Kuwahara, M. Chronic stimulation of group II metabotropic glutamate receptors in the medulla oblongata attenuates hypertension development in spontaneously hypertensive rats. PLoS ONE 2021, 16, e0251495. [Google Scholar] [CrossRef]
- Simms, A.E.; Paton, J.F.R.; Pickering, A.E. Disinhibition of the cardiac limb of the arterial baroreflex in rat: A role for metabotropic glutamate receptors in the nucleus tractus solitarii. J. Physiol. 2006, 575 Pt 3, 727–738. [Google Scholar] [CrossRef]
- Kuwahara, M.; Sugano, S.; Yayou, K.; Tsubone, H.; Kobayashi, H. Evaluation of a new tail-cuff method for blood pressure measurements in rats with special reference to the effects of ambient temperature. Exp. Anim. 1991, 40, 331–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tochinai, R.; Komatsu, K.; Murakami, J.; Nagata, Y.; Ando, M.; Hata, C.; Suzuki, T.; Kado, S.; Kobayashi, T.; Kuwahara, M. Histopathological and functional changes in a single-dose model of combretastatin A4 disodium phosphate-induced myocardial damage in rats. J. Toxicol. Pathol. 2018, 31, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Petersen, L.J.; Petersen, J.R.; Talleruphuus, U.; Ladefoged, S.D.; Mehlsen, J.; Jensen, H.A. The pulsatility index and the resistive index in renal arteries. Associations with long-term progression in chronic renal failure. Nephrol. Dial. Transplant. 1997, 12, 1376–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, M.; Kuwahara, M.; Tsubone, H.; Sugano, S. Diurnal variation of autonomic nervous activity in the rat: Investigation by power spectral analysis of heart rate variability. J. Electrocardiol. 1999, 32, 167–171. [Google Scholar] [CrossRef]
- Kuwahara, M.; Yayou, K.; Ishii, K.; Hashimoto, S.; Tsubone, H.; Sugano, S. Power spectral analysis of heart rate variability as a new method for assessing autonomic activity in the rat. J. Electrocardiol. 1994, 27, 333–337. [Google Scholar] [CrossRef]
- Almeida, J.; Oliveira, L.A.; Benini, R.; Crestani, C.C. Differential roles of hippocampal nNOS and iNOS in the control of baroreflex function in conscious rats. Brain Res. 2019, 1710, 109–116. [Google Scholar] [CrossRef]
- Lai, C.C.; Yuan, Z.F.; Chu, L.Y.; Chuang, K.T.; Lin, H.H. Roles of cocaine- and amphetamine-regulated transcript peptide in the rostral ventrolateral medulla in cardiovascular regulation in rats. Brain Res. 2019, 1710, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Pak, C.H. Plasma Adrenaline and Noradrenaline Concentrations of the Spontaneously Hypertensive Rat. Jpn. Heart J. 1981, 22, 987–995. [Google Scholar] [CrossRef] [Green Version]
- Kuo, Y.J.; Keeton, T.K. Captopril increases norepinephrine spillover rate in conscious spontaneously hypertensive rats. J. Pharmacol. Exp. Ther. 1991, 258, 223–231. [Google Scholar]
- Berg, T.; Walaas, S.I.; Roberg, B.Å.; Huynh, T.T.; Jensen, J. Plasma norepinephrine in hypertensive rats reflects α2-adrenoceptor release control only when re-uptake is inhibited. Front. Neurol. 2012, 3, 160. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, K.; Averill, D.B.; Ferrario, C.M. Role of AT1 receptors in area postrema on baroreceptor reflex in spontaneously hypertensive rats. Brain Res. 1999, 850, 166–172. [Google Scholar] [CrossRef]
- Minami, N.; Mori, N.; Nagasaka, M.; Ito, O.; Kurosawa, H.; Kanazawa, M.; Kaku, K.; Lee, E.; Kohzuki, M. Mechanism behind augmentation in baroreflex sensitivity after acute exercise in spontaneously hypertensive rats. Hypertens Res. 2006, 29, 117–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertagnolli, M.; Campos, C.; Schenkel, P.C.; de Oliveira, V.L.L.; De Angelis, K.; Belló-Klein, A.; Rigatto, K.; Irigoyen, M.C. Baroreflex sensitivity improvement is associated with decreased oxidative stress in trained spontaneously hypertensive rat. J. Hypertens. 2006, 24, 2437–2443. [Google Scholar] [CrossRef]
- Gross, P.M.; Wall, K.M.; Pang, J.J.; Shaver, S.W.; Wainman, D.S. Microvascular specializations promoting rapid interstitial solute dispersion in nucleus tractus solitarius. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1990, 259, R1131–R1138. [Google Scholar] [CrossRef]
- Melo, M.R.; Gasparini, S.; Speretta, G.F.; Silva, E.F.; Pedrino, G.R.; Menani, J.V.; Zoccal, D.B.; Colombari, D.S.A.; Colombari, E. Importance of the commissural nucleus of the solitary tract in renovascular hypertension. Hypertens. Res. 2019, 42, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Gehrmann, J.; Hammer, P.E.; Maguire, C.T.; Wakimoto, H.; Triedman, J.K.; Berul, C.I. Phenotypic screening for heart rate variability in the mouse. Am. J. Physiol. Heart Circ. Physiol. 2000, 279, H733–H740. [Google Scholar] [CrossRef]
- Aicher, S.A.; Saravay, R.H.; Cravo, S.L.; Jeske, I.; Morrison, S.F.; Reis, D.J.; Milner, T.A. Monosynaptic projections from the nucleus tractus solitarii to C1 adrenergic neurons in the rostral ventrolateral medulla: Comparison with input from the caudal ventrolateral medulla. J. Comp. Neurol. 1996, 373, 62–75. [Google Scholar] [CrossRef]
- Koshiya, N.; Guyenet, P.G. NTS neurons with carotid chemoreceptor inputs arborize in the rostral ventrolateral medulla. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1996, 270, R1273–R1278. [Google Scholar] [CrossRef]
- Colombari, E.; Menani, J.V.; Talman, W.T. Commissural NTS contribute to pressor responses to glutamate injected into the medial NTS of awake rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1996, 270, R1220–R1225. [Google Scholar] [CrossRef]
- Blessing, W.W.; Yu, Y.H.; Nalivaiko, E. Medullary projections of rabbit carotid sinus nerve. Brain Res. 1999, 816, 405–410. [Google Scholar] [CrossRef]
- Ghali, M.G.Z. The brainstem network controlling blood pressure an important role for pressor sites in the caudal medulla and cervical spinal cord. J. Hypertens. 2017, 35, 1938–1947. [Google Scholar] [CrossRef]
- Chen, C.Y.; Bonham, A.C. Glutamate suppresses GABA release via presynaptic metabotropic glutamate receptors at baroreceptor neurones in rats. J. Physiol. 2005, 562 Pt 2, 535–551. [Google Scholar] [CrossRef]
- Slama, M.; Susic, D.; Varagic, J.; Ahn, J.; Frohlich, E.D. Echocardiographic measurement of cardiac output in rats. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H691–H697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, D.C.; Johnson, A.K. Development of autonomic control of heart rate in genetically hypertensive and normotensive rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1984, 246, R570–R577. [Google Scholar] [CrossRef]
- Rinaman, L. Hindbrain noradrenergic A2 neurons: Diverse roles in autonomic, endocrine, cognitive, and behavioral functions. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R222–R235. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters (Unit) | Wistar Kyoto Rats (n = 5) | SHRs (with LY341495) (n = 5) | p-Value (t-Test) |
|---|---|---|---|
| Cardiac | |||
| Heart Rate (BPM) | 329.5 ± 14.0 | 343.4 ± 15.2 | 0.520 |
| Stroke Volume (µL) | 213.4 ± 9.5 | 241.0 ± 20.9 | 0.264 |
| Ejection Fraction (%) | 69.1 ± 2.8 | 61.6 ± 3.6 | 0.138 |
| Fractional Shortening (%) | 40.3 ± 2.2 | 34.8 ± 2.7 | 0.147 |
| Cardiac Output (mL/min) | 70.3 ± 4.1 | 81.8 ± 5.1 | 0.117 |
| Renal | |||
| LRA PSV (mm/s) | 780.6 ± 86.3 | 440.8 ± 100.6 | 0.033 * |
| RRA PSV (mm/s) | 786.3 ± 65.5 | 544.2 ± 74.0 | 0.040 * |
| LRA LDV (mm/s) | 237.4 ± 34.0 | 207.9 ± 46.1 | 0.621 |
| RRA LDV (mm/s) | 231.7 ± 41.8 | 215.4 ± 25.6 | 0.748 |
| LRA PI | 1.304 ± 0.276 | 0.746 ± 0.042 | 0.081 |
| RRA PI | 1.377 ± 0.200 | 0.905 ± 0.071 | 0.057 |
| LRA RI | 0.673 ± 0.060 | 0.526 ± 0.023 | 0.052 |
| RRA RI | 0.701 ± 0.048 | 0.596 ± 0.021 | 0.080 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, J.C.-N.; Sekizawa, S.; Tochinai, R.; Kuwahara, M. Loss of Group II Metabotropic Glutamate Receptor Signaling Exacerbates Hypertension in Spontaneously Hypertensive Rats. Life 2021, 11, 720. https://0-doi-org.brum.beds.ac.uk/10.3390/life11070720
Hsu JC-N, Sekizawa S, Tochinai R, Kuwahara M. Loss of Group II Metabotropic Glutamate Receptor Signaling Exacerbates Hypertension in Spontaneously Hypertensive Rats. Life. 2021; 11(7):720. https://0-doi-org.brum.beds.ac.uk/10.3390/life11070720
Chicago/Turabian StyleHsu, Julia Chu-Ning, Shinichi Sekizawa, Ryota Tochinai, and Masayoshi Kuwahara. 2021. "Loss of Group II Metabotropic Glutamate Receptor Signaling Exacerbates Hypertension in Spontaneously Hypertensive Rats" Life 11, no. 7: 720. https://0-doi-org.brum.beds.ac.uk/10.3390/life11070720