
Impact of Astrocytic Coverage of Synapses on the Short-Term Memory of a Computational Neuron-Astrocyte Network


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
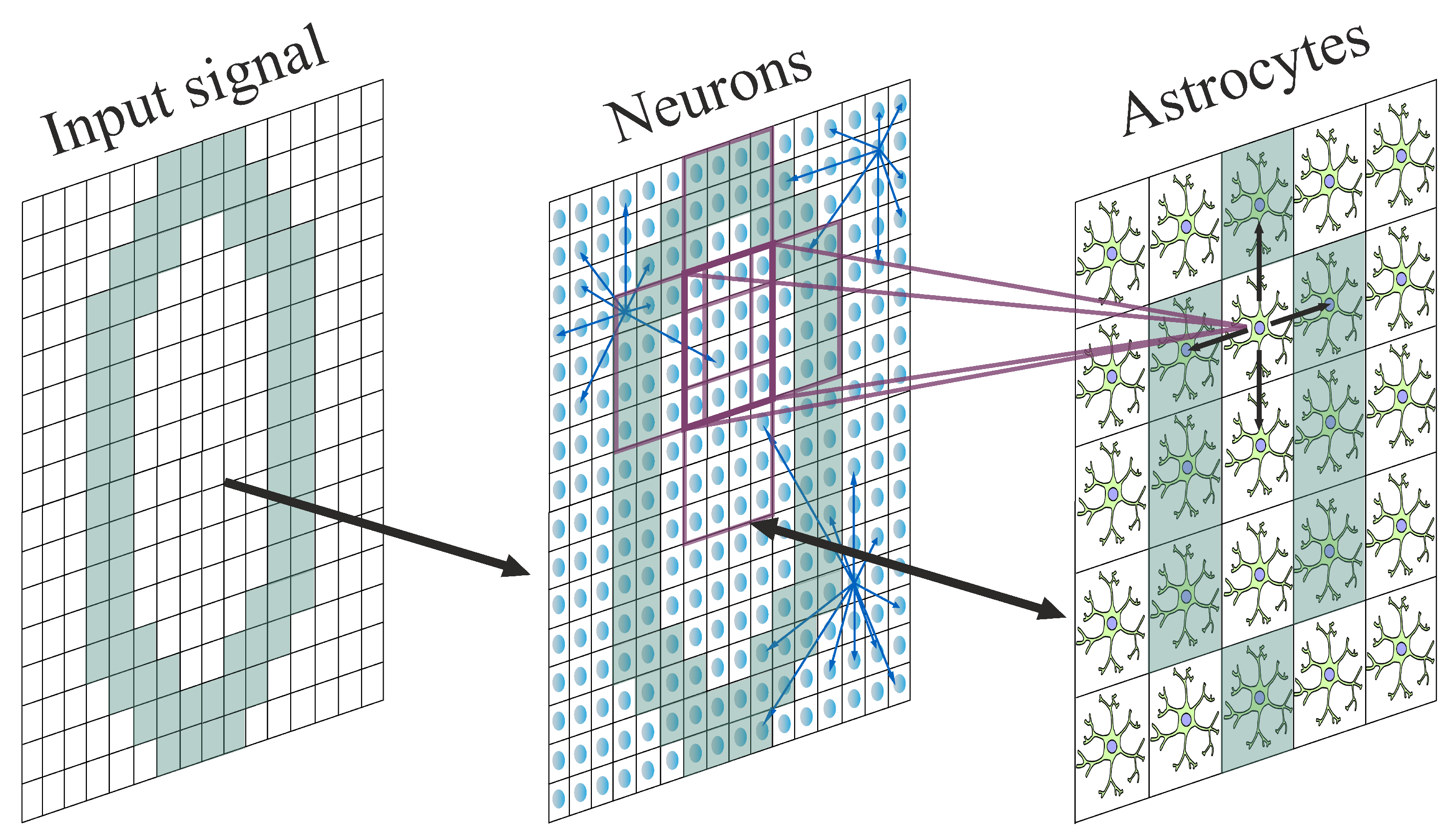
2. Models and Methods
2.1. Neuronal Network
2.2. Action Potential-Induced Elevation of Glutamate and
2.3. Astrocytic Network
2.4. Variation of Astrocytic Coverage
2.5. Variation of Spatial Frequencies
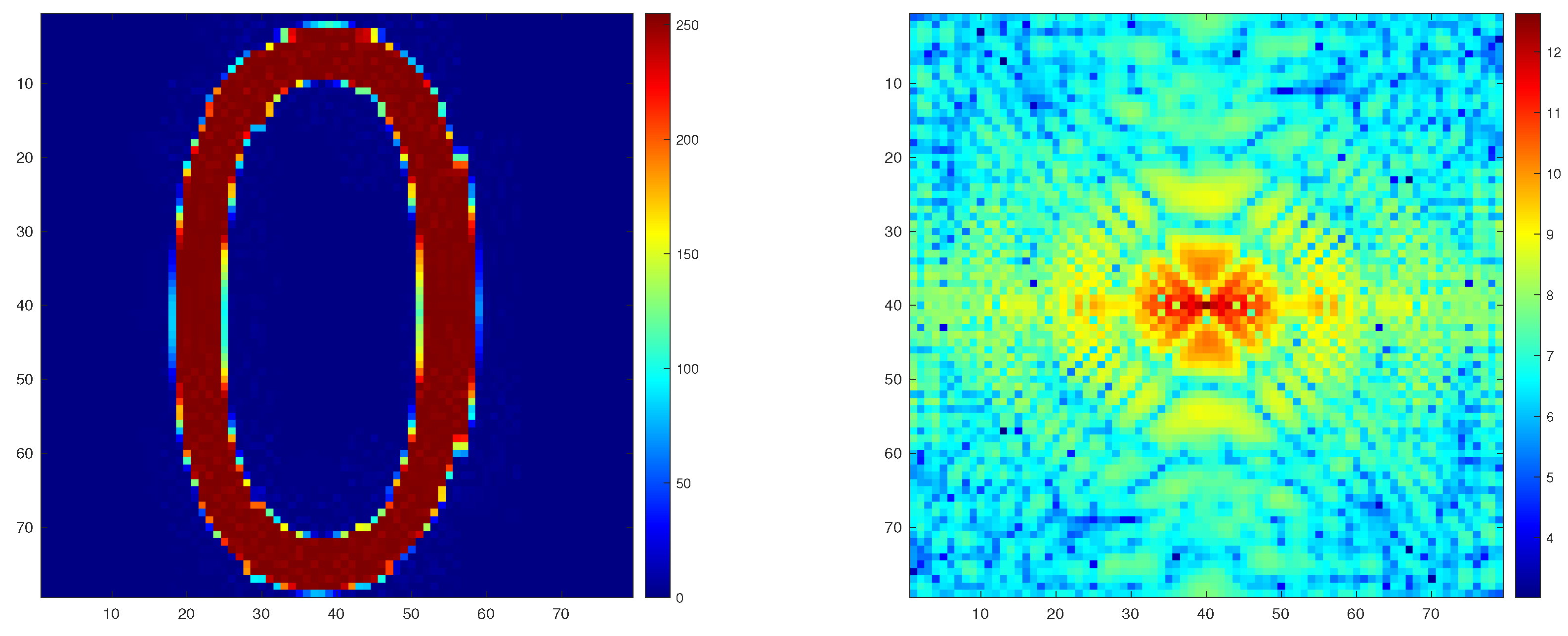
2.6. Simulation Protocols
2.7. Performance Measure
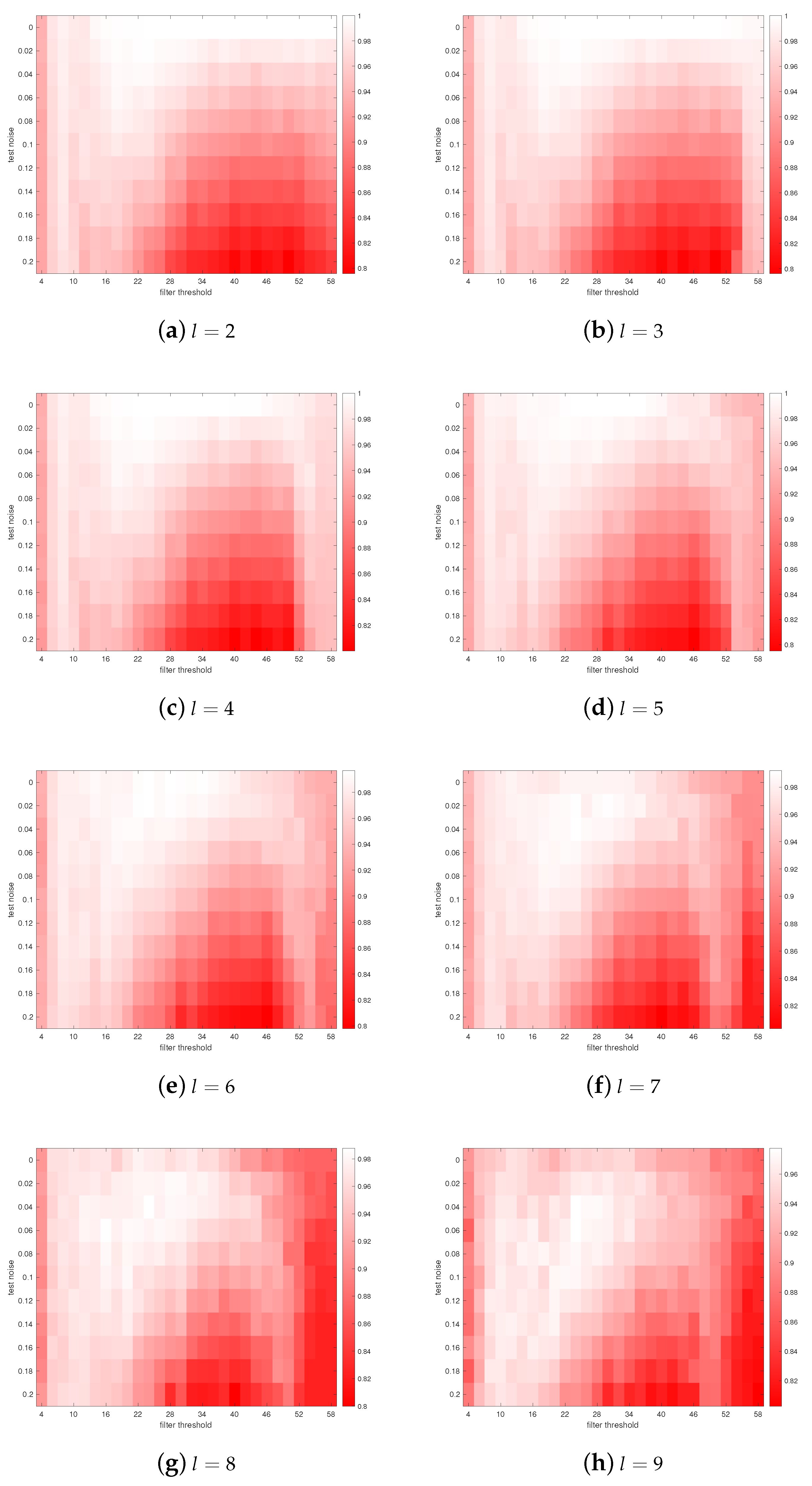
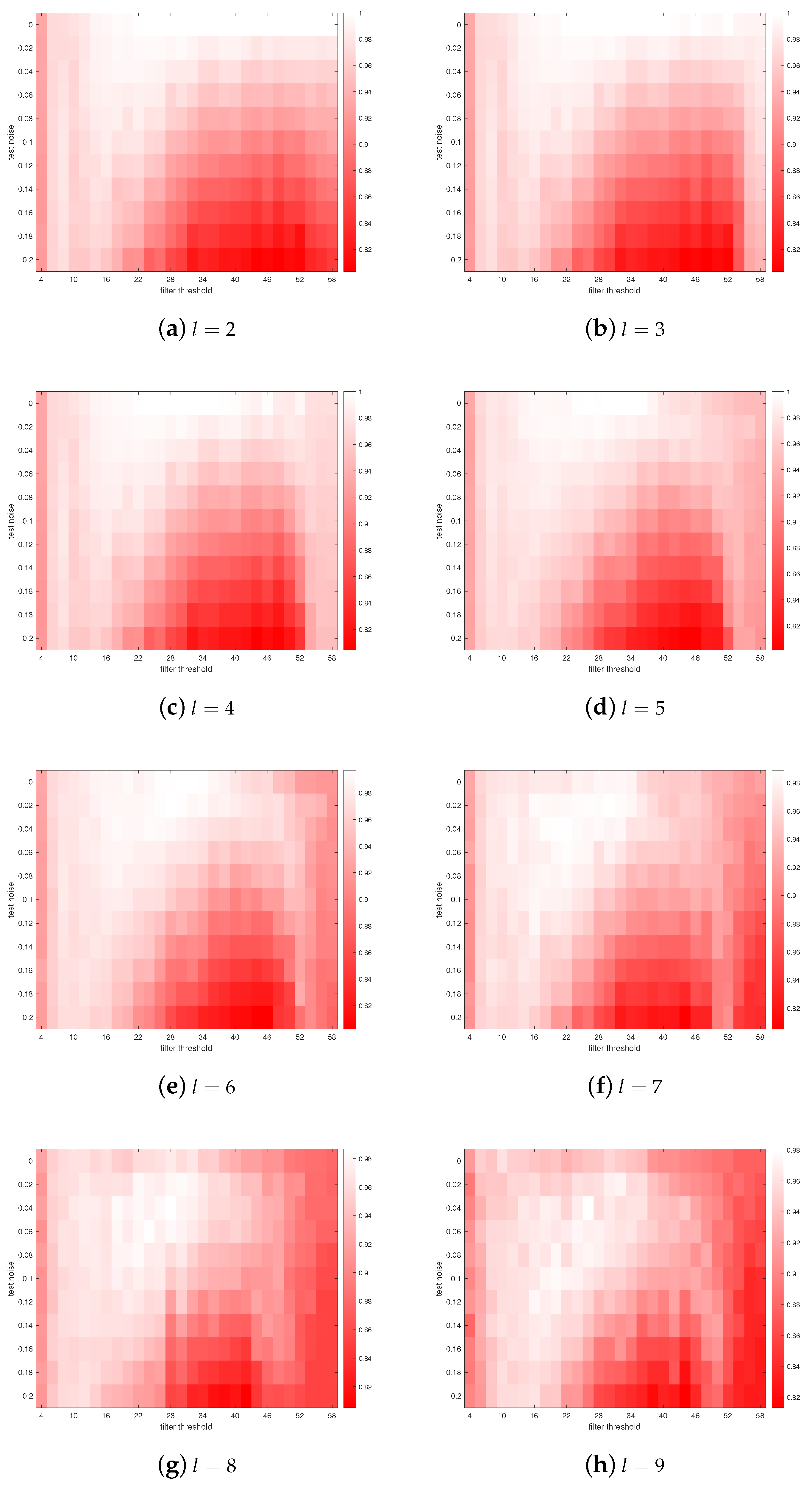
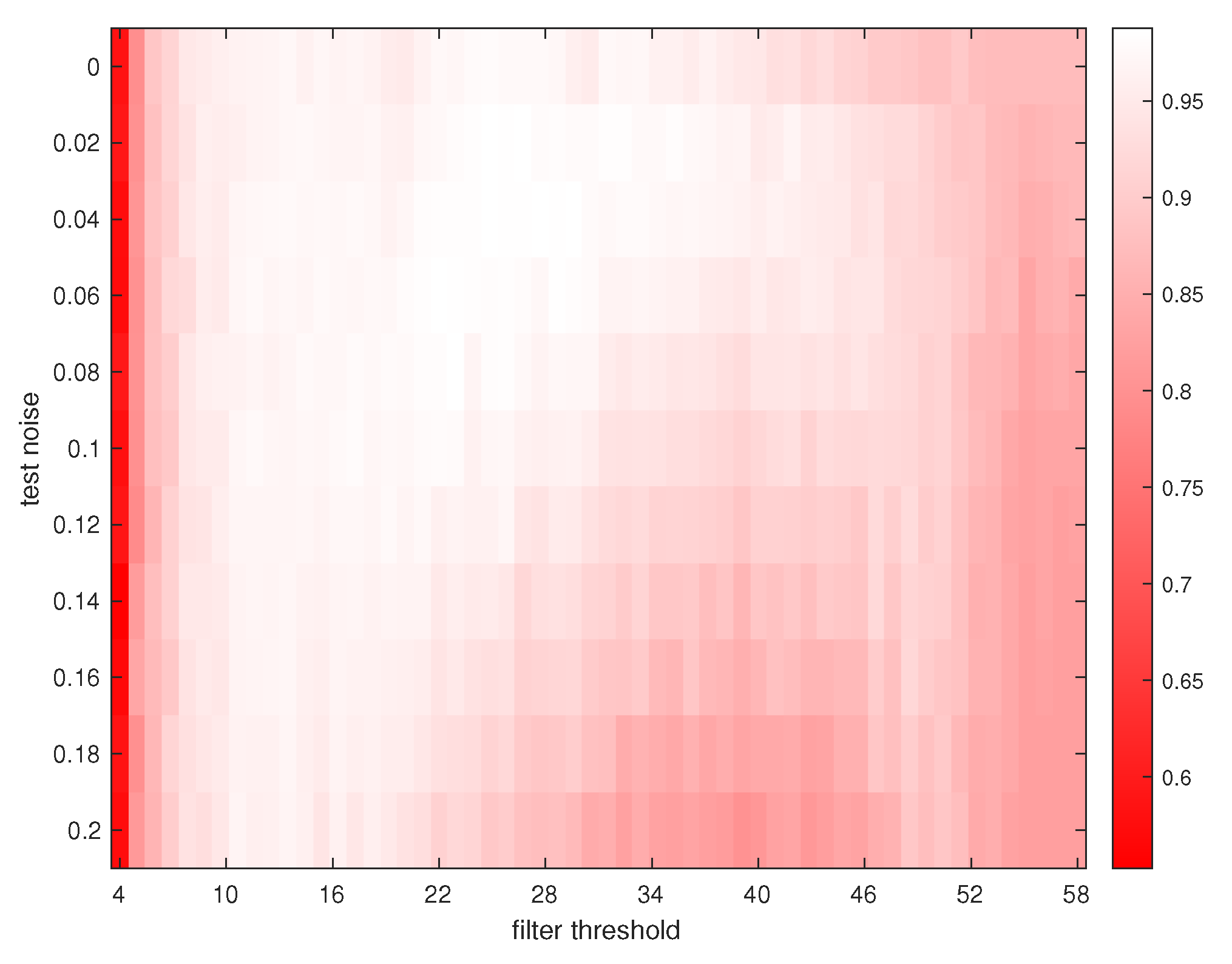
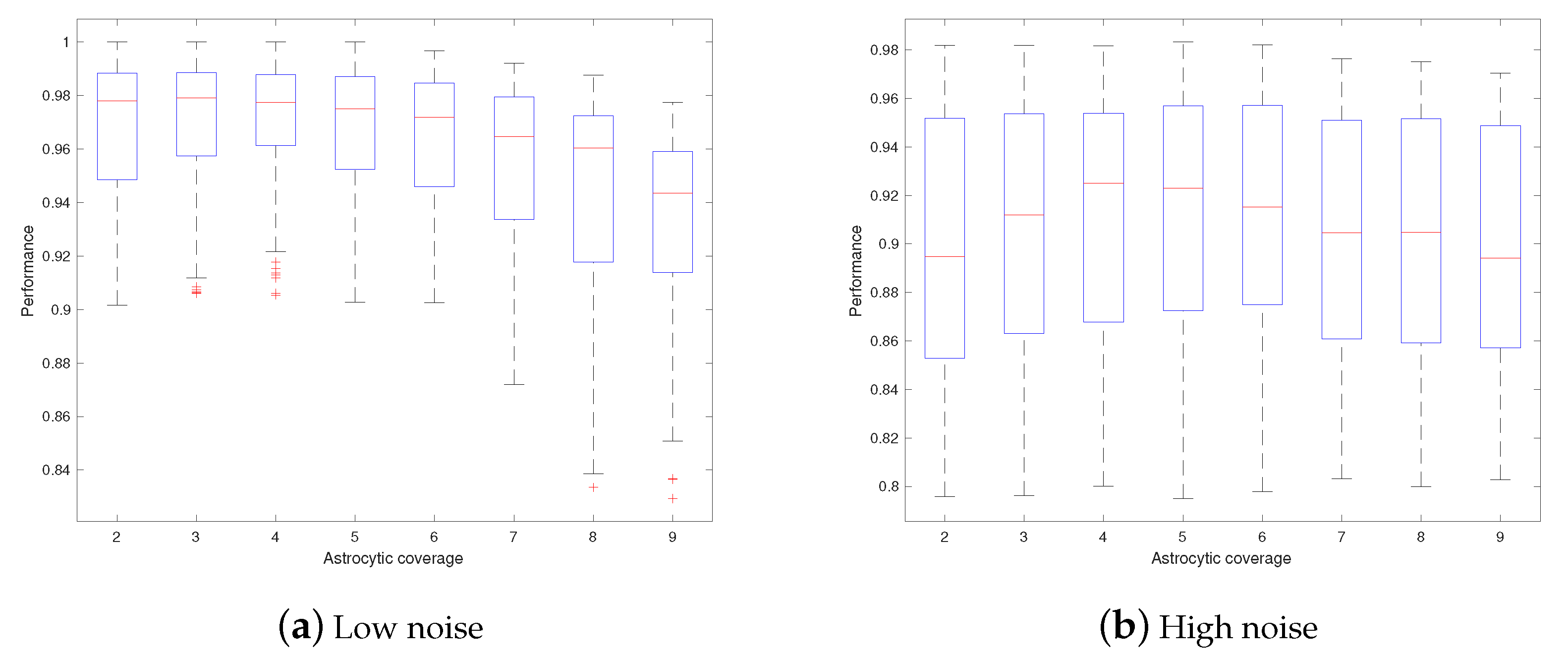
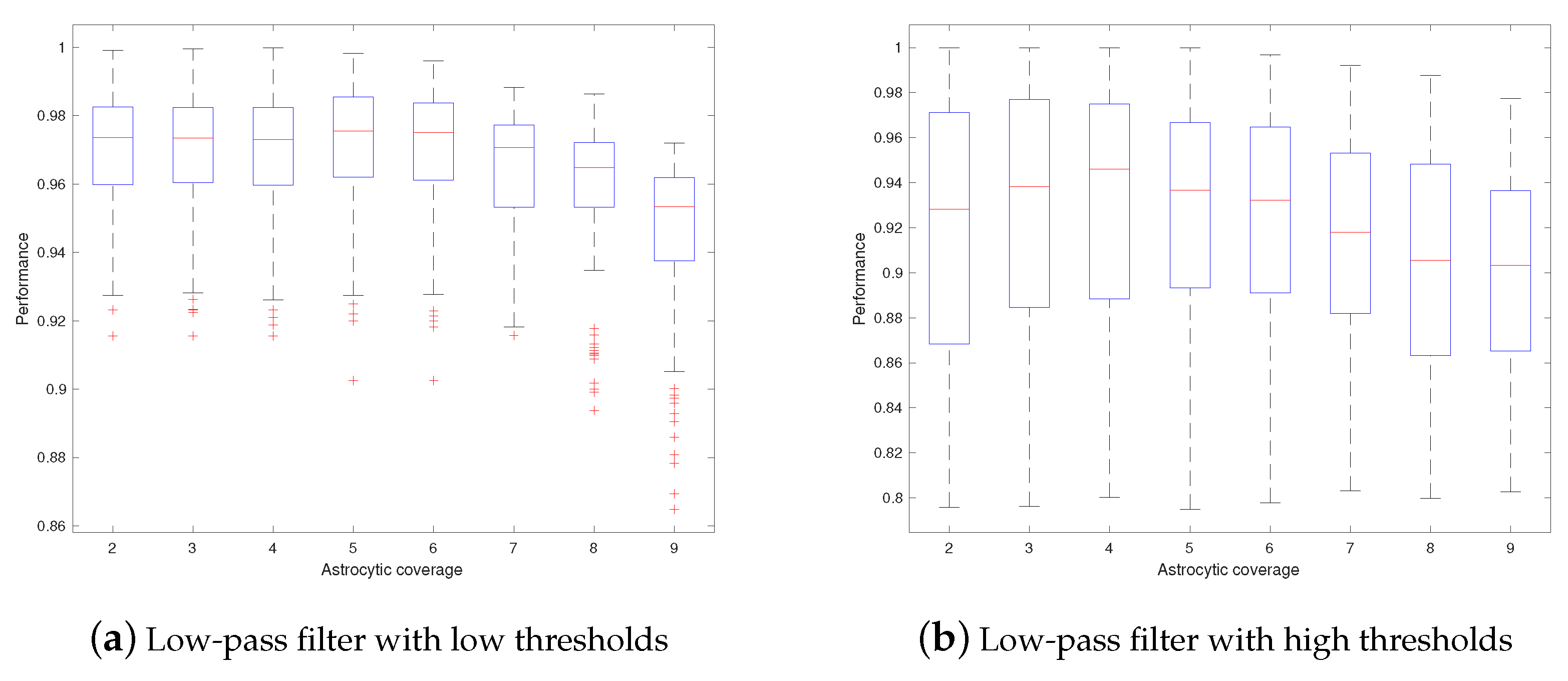
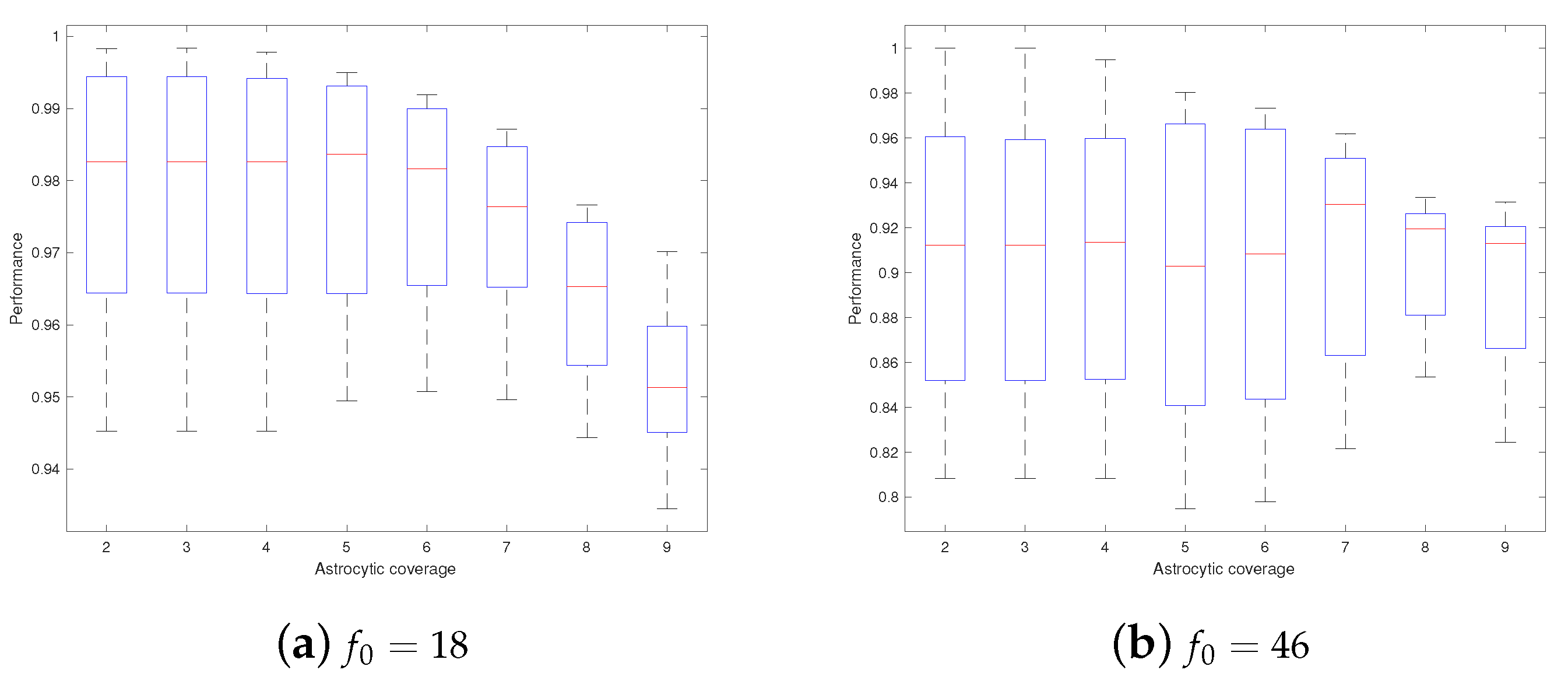
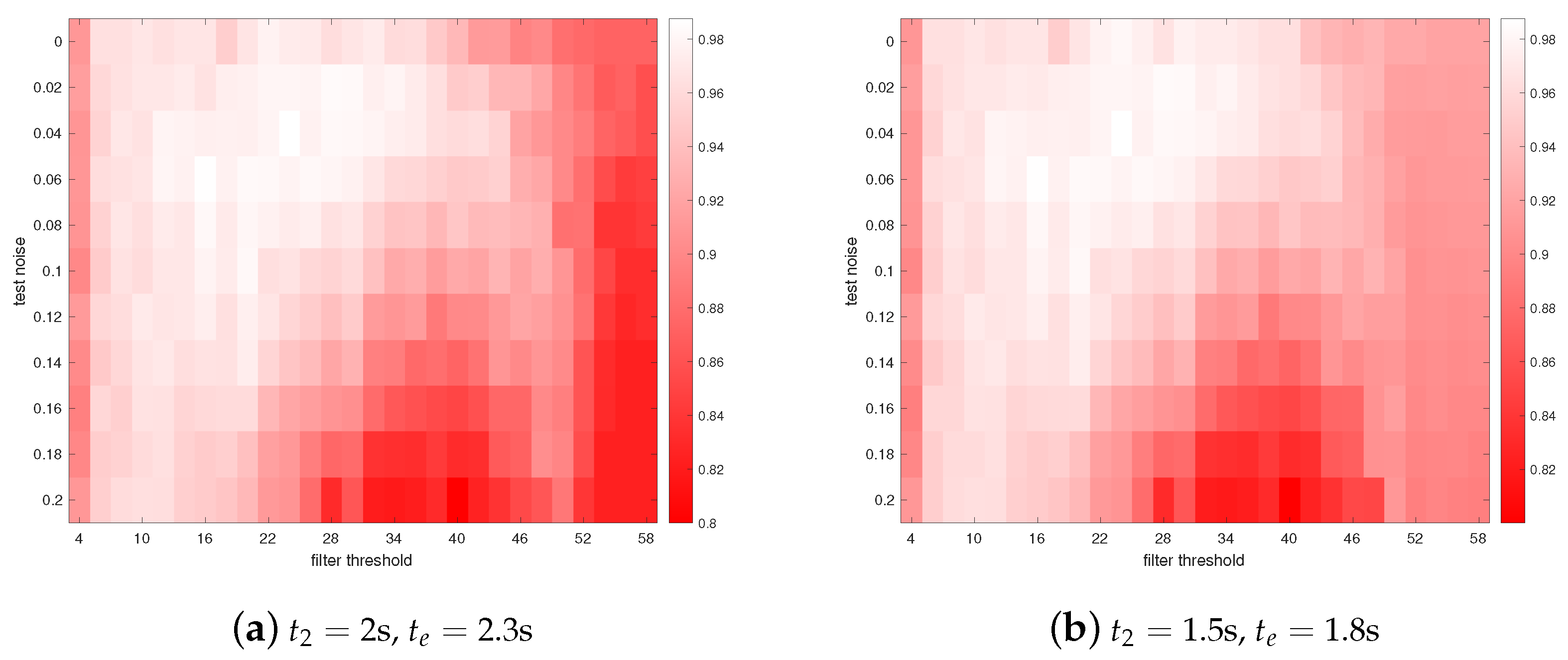
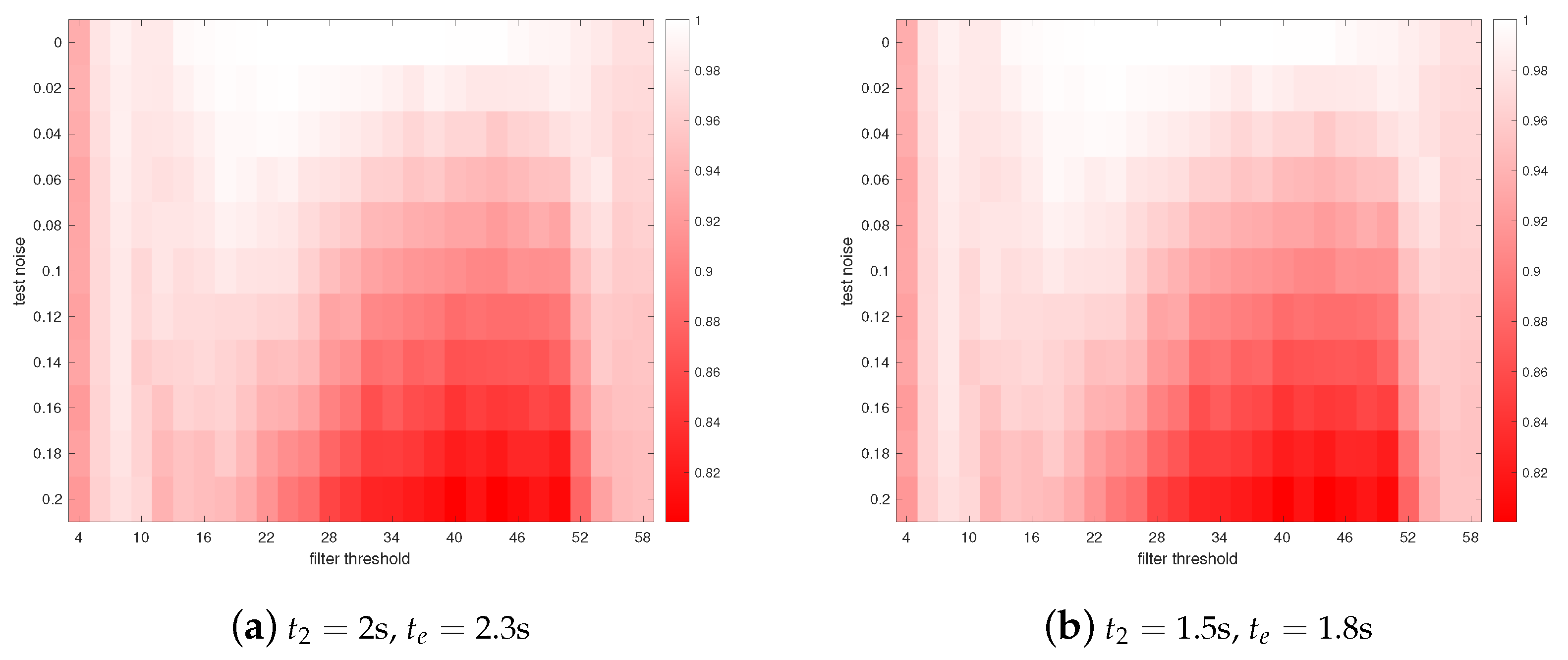
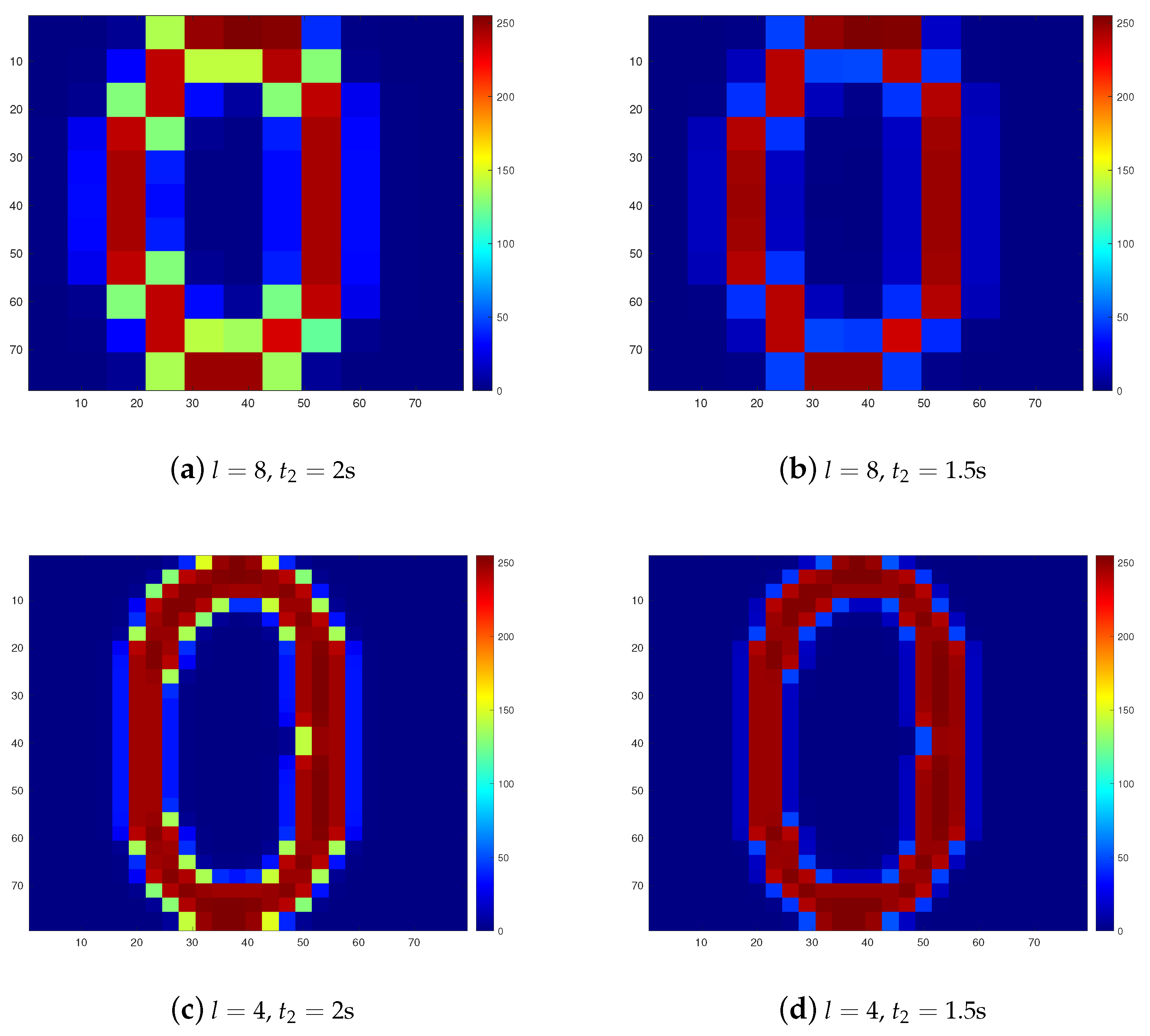
3. Results
4. Discussions
- Astrocytes may adjust their coverage areas in response to the change in spatial frequencies in order to optimize the short-term memory.
- Different astrocytes may have different coverage areas in order to process different frequency components in order to optimize the short-term memory.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Latham, P.E.; Richmond, B.J.; Nelson, P.G.; Nirenberg, S. Intrinsic dynamics in neuronal networks. I. Theory. J. Neurophysiol. 2000, 83, 808–827. [Google Scholar] [CrossRef] [PubMed]
- Van Pelt, J.; Vajda, I.; Wolters, P.S.; Corner, M.A.; Ramakers, G.J. Dynamics and plasticity in developing neuronal networks in vitro. Prog. Brain Res. 2005, 147, 171–188. [Google Scholar]
- Roxin, A.; Montbrió, E. How effective delays shape oscillatory dynamics in neuronal networks. Phys. Nonlinear Phenom. 2011, 240, 323–345. [Google Scholar] [CrossRef]
- Zhang, H.; Ye, M.; Ye, R.; Cao, J. Synchronization stability of Riemann–Liouville fractional delay-coupled complex neural networks. Phys. Stat. Mech. Appl. 2018, 508, 155–165. [Google Scholar] [CrossRef]
- Michel, C.M.; Koenig, T. EEG microstates as a tool for studying the temporal dynamics of whole-brain neuronal networks: A review. Neuroimage 2018, 180, 577–593. [Google Scholar] [CrossRef]
- Zhang, H.; Cheng, J.; Zhang, H.; Zhang, W.; Cao, J. Quasi-uniform synchronization of Caputo type fractional neural networks with leakage and discrete delays. Chaos Solitons Fractals 2021, 152, 111432. [Google Scholar] [CrossRef]
- Zhang, H.; Cheng, Y.; Zhang, H.; Zhang, W.; Cao, J. Hybrid control design for Mittag-Leffler projective synchronization on FOQVNNs with multiple mixed delays and impulsive effects. Math. Comput. Simul. 2022, 197, 341–357. [Google Scholar] [CrossRef]
- Agulhon, C.; Petravicz, J.; McMullen, A.B.; Sweger, E.J.; Minton, S.K.; Taves, S.R.; Casper, K.B.; Fiacco, T.A.; McCarthy, K.D. What is the role of astrocyte calcium in neurophysiology? Neuron 2008, 59, 932–946. [Google Scholar] [CrossRef]
- Booth, H.D.; Hirst, W.D.; Wade-Martins, R. The role of astrocyte dysfunction in Parkinson’s disease pathogenesis. Trends Neurosci. 2017, 40, 358–370. [Google Scholar] [CrossRef]
- Upadhya, R.; Zingg, W.; Shetty, S.; Shetty, A.K. Astrocyte-derived extracellular vesicles: Neuroreparative properties and role in the pathogenesis of neurodegenerative disorders. J. Control. Release 2020, 323, 225–239. [Google Scholar] [CrossRef]
- Rose, C.F.; Verkhratsky, A.; Parpura, V. Astrocyte glutamine synthetase: Pivotal in health and disease. Biochem. Soc. Trans. 2013, 41, 1518–1524. [Google Scholar] [CrossRef] [PubMed]
- Brusilow, S.W.; Koehler, R.C.; Traystman, R.J.; Cooper, A.J. Astrocyte glutamine synthetase: Importance in hyperammonemic syndromes and potential target for therapy. Neurotherapeutics 2010, 7, 452–470. [Google Scholar] [CrossRef]
- Gordleeva, S.; Kanakov, O.; Ivanchenko, M.; Zaikin, A.; Franceschi, C. Brain aging and garbage cleaning. Semin. Immunopathol. 2020, 42, 647–665. [Google Scholar] [CrossRef] [PubMed]
- Haydon, P.G. GLIA: Listening and talking to the synapse. Nat. Rev. Neurosci. 2001, 2, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Parpura, V.; Haydon, P.G. Physiological astrocytic calcium levels stimulate glutamate release to modulate adjacent neurons. Proc. Natl. Acad. Sci. USA 2000, 97, 8629–8634. [Google Scholar] [CrossRef]
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Tripartite synapses: Glia, the unacknowledged partner. Trends Neurosci. 1999, 22, 208–215. [Google Scholar] [CrossRef]
- Perea, G.; Araque, A. GLIA modulates synaptic transmission. Brain Res. Rev. 2010, 63, 93–102. [Google Scholar] [CrossRef]
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Glutamate-dependent astrocyte modulation of synaptic transmission between cultured hippocampal neurons. Eur. J. Neurosci. 1998, 10, 2129–2142. [Google Scholar] [CrossRef]
- Halassa, M.M.; Fellin, T.; Haydon, P.G. The tripartite synapse: Roles for gliotransmission in health and disease. Trends Mol. Med. 2007, 13, 54–63. [Google Scholar] [CrossRef]
- Santello, M.; Calì, C.; Bezzi, P. Gliotransmission and the tripartite synapse. Synaptic Plast. 2012, 970, 307–331. [Google Scholar]
- Volterra, A.; Magistretti, P.J.; Haydon, P.G. The Tripartite Synapse: Glia in Synaptic Transmission; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Perea, G.; Navarrete, M.; Araque, A. Tripartite synapses: Astrocytes process and control synaptic information. Trends Neurosci. 2009, 32, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Panatier, A.; Robitaille, R. Astrocytic mGluR5 and the tripartite synapse. Neuroscience 2016, 323, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Fellin, T.; Pascual, O.; Gobbo, S.; Pozzan, T.; Haydon, P.G.; Carmignoto, G. Neuronal synchrony mediated by astrocytic glutamate through activation of extrasynaptic NMDA receptors. Neuron 2004, 43, 729–743. [Google Scholar] [CrossRef] [PubMed]
- Allegrini, P.; Fronzoni, L.; Pirino, D. The influence of the astrocyte field on neuronal dynamics and synchronization. J. Biol. Phys. 2009, 35, 413–423. [Google Scholar] [CrossRef]
- Wade, J.J.; McDaid, L.J.; Harkin, J.; Crunelli, V.; Kelso, J.S. Bidirectional coupling between astrocytes and neurons mediates learning and dynamic coordination in the brain: A multiple modeling approach. PLoS ONE 2011, 6, e29445. [Google Scholar]
- Chen, N.; Sugihara, H.; Sharma, J.; Perea, G.; Petravicz, J.; Le, C.; Sur, M. Nucleus basalis-enabled stimulus-specific plasticity in the visual cortex is mediated by astrocytes. Proc. Natl. Acad. Sci. USA 2012, 109, E2832–E2841. [Google Scholar] [CrossRef]
- Gordleeva, S.Y.; Lebedev, S.A.; Rumyantseva, M.A.; Kazantsev, V.B. Astrocyte as a detector of synchronous events of a neural network. JETP Lett. 2018, 107, 440–445. [Google Scholar] [CrossRef]
- Pankratova, E.V.; Kalyakulina, A.I.; Stasenko, S.V.; Gordleeva, S.Y.; Lazarevich, I.A.; Kazantsev, V.B. Neuronal synchronization enhanced by neuron–astrocyte interaction. Nonlinear Dyn. 2019, 97, 647–662. [Google Scholar] [CrossRef]
- Gordleeva, S.Y.; Ermolaeva, A.V.; Kastalskiy, I.A.; Kazantsev, V.B. Astrocyte as spatiotemporal integrating detector of neuronal activity. Front. Physiol. 2019, 10, 294. [Google Scholar] [CrossRef]
- Makovkin, S.Y.; Shkerin, I.V.; Gordleeva, S.Y.; Ivanchenko, M.V. Astrocyte-induced intermittent synchronization of neurons in a minimal network. Chaos Solitons Fractals 2020, 138, 109951. [Google Scholar] [CrossRef]
- Makovkin, S.; Kozinov, E.; Ivanchenko, M.; Gordleeva, S. Controlling synchronization of gamma oscillations by astrocytic modulation in a model hippocampal neural network. Sci. Rep. 2022, 12, 6970. [Google Scholar] [CrossRef] [PubMed]
- Gordleeva, S.; Tsybina, Y.A.; Krivonosov, M.I.; Tyukin, I.Y.; Kazantsev, V.B.; Zaikin, A.A.; Gorban, A.N. Situation-based memory in spiking neuron-astrocyte network. arXiv 2022, arXiv:2202.07218. [Google Scholar]
- Tsybina, Y.; Gordleeva, S.; Krivonosov, M.; Kastalskiy, I.; Zaikin, A.; Gorban, A. Modelling working memory in neuron-astrocyte network. In Proceedings of the 2021 International Joint Conference on Neural Networks (IJCNN) 2021, Shenzhen, China, 18–22 July 2021; pp. 1–6. [Google Scholar]
- Gordleeva, S.Y.; Tsybina, Y.A.; Krivonosov, M.I.; Ivanchenko, M.V.; Zaikin, A.A.; Kazantsev, V.B.; Gorban, A.N. Modelling working memory in spiking neuron network accompanied by astrocytes. Front. Cell. Neurosci. 2021, 15, 86. [Google Scholar] [CrossRef] [PubMed]
- Tsybina, Y.; Kastalskiy, I.; Krivonosov, M.; Zaikin, A.; Kazantsev, V.; Gorban, A.; Gordleeva, S. Astrocytes mediate analogous memory in a multi-layer neuron-astrocytic network. Neural Comput. Appl. 2022, 34, 9147–9160. [Google Scholar] [CrossRef]
- Tsybina, Y.; Krivonosov, M.; Gordleeva, S.; Zaikin, A.; Gorban, A. Short-term memory in neuron-astrocyte network. In Proceedings of the 2020 4th Scientific School on Dynamics of Complex Networks and Their Application in Intellectual Robotics (DCNAIR), Innopolis, Russia, 9–11 September 2020; pp. 245–247. [Google Scholar]
- Kastalskiy, I.; Tsybina, Y.; Kazantsev, V.; Gordleeva, S. Astrocytes’ signals guided storage and retrieval of patterns by an SNN. In Proceedings of the 2021 Third International Conference Neurotechnologies and Neurointerfaces (CNN), Kaliningrad, Russia, 13–15 September 2021; pp. 34–37. [Google Scholar]
- Abrego, L.; Gordleeva, S.; Kanakov, O.; Krivonosov, M.; Zaikin, A. Estimating integrated information in bidirectional neuron-astrocyte communication. Phys. Rev. E 2021, 103, 022410. [Google Scholar] [CrossRef] [PubMed]
- Kanakov, O.; Gordleeva, S.; Ermolaeva, A.; Jalan, S.; Zaikin, A. Astrocyte-induced positive integrated information in neuron-astrocyte ensembles. Phys. Rev. E 2019, 99, 012418. [Google Scholar] [CrossRef]
- Abrego Rangel, L.A. Information Processing in Biological Complex Systems: A View to Bacterial and Neural Complexity. Ph.D. Thesis, UCL (University College London), London, UK, 2021. [Google Scholar]
- Kanakov, O.; Gordleeva, S.; Zaikin, A. Integrated Information in the Spiking–Bursting Stochastic Model. Entropy 2020, 22, 1334. [Google Scholar] [CrossRef]
- Suzuki, A.; Stern, S.A.; Bozdagi, O.; Huntley, G.W.; Walker, R.H.; Magistretti, P.J.; Alberini, C.M. Astrocyte-neuron lactate transport is required for long-term memory formation. Cell 2011, 144, 810–823. [Google Scholar] [CrossRef]
- Habbas, S.; Santello, M.; Becker, D.; Stubbe, H.; Zappia, G.; Liaudet, N.; Klaus, F.R.; Kollias, G.; Fontana, A.; Pryce, C.R.; et al. Neuroinflammatory TNFα impairs memory via astrocyte signaling. Cell 2015, 163, 1730–1741. [Google Scholar] [CrossRef]
- Adamsky, A.; Kol, A.; Kreisel, T.; Doron, A.; Ozeri-Engelhard, N.; Melcer, T.; Refaeli, R.; Horn, H.; Regev, L.; Groysman, M.; et al. Astrocytic activation generates de novo neuronal potentiation and memory enhancement. Cell 2018, 174, 59–71. [Google Scholar] [CrossRef]
- Kozachkov, L.; Michmizos, K.P. Sequence Learning in Associative Neuronal-Astrocytic Networks. In Proceedings of the International Conference on Brain Informatics, Padua, Italy, 19 September 2020; Springer: Cham, Switzerland, 2020; pp. 349–360. [Google Scholar]
- De Luca, S.N.; Soch, A.; Sominsky, L.; Nguyen, T.X.; Bosakhar, A.; Spencer, S.J. Glial remodeling enhances short-term memory performance in Wistar rats. J. Neuroinflamm. 2020, 17, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cowan, N. Working memory underpins cognitive development, learning, and education. Educ. Psychol. Rev. 2014, 26, 197–223. [Google Scholar] [CrossRef] [PubMed]
- Baddeley, A. Working memory: The interface between memory and cognition. J. Cogn. Neurosci. 1992, 4, 281–288. [Google Scholar] [CrossRef]
- Nairne, J.S. Short-term/working memory. In Memory 1 January 1996; Academic Press: Cambridge, MA, USA, 1996; pp. 349–360. [Google Scholar]
- Lima, A.; Sardinha, V.M.; Oliveira, A.F.; Reis, M.; Mota, C.; Silva, M.A.; Marques, F.; Cerqueira, J.J.; Pinto, L.; Sousa, N.; et al. Astrocyte pathology in the prefrontal cortex impairs the cognitive function of rats. Mol. Psychiatry 2014, 19, 834–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robin, L.M.; da Cruz, J.F.; Langlais, V.C.; Martin-Fernez, M.; Metna-Laurent, M.; Busquets-Garcia, A.; Bellocchio, L.; Soria-Gomez, E.; Papouin, T.; Varilh, M.; et al. Astroglial CB1 receptors determine synaptic D-serine availability to enable recognition memory. Neuron 2018, 98, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Santello, M.; Toni, N.; Volterra, A. Astrocyte function from information processing to cognition and cognitive impairment. Nat. Neurosci. 2019, 22, 154–166. [Google Scholar] [CrossRef]
- Akther, S.; Hirase, H. Assessment of astrocytes as a mediator of memory and learning in rodents. Glia 2021, 70, 1484–1505. [Google Scholar] [CrossRef]
- Chung, W.S.; Allen, N.J.; Eroglu, C. Astrocytes control synapse formation, function, and elimination. Cold Spring Harb. Perspect. Biol. 2015, 7, a020370. [Google Scholar] [CrossRef]
- Heller, J.P.; Rusakov, D.A. Morphological plasticity of astroglia: Understanding synaptic microenvironment. Glia 2015, 63, 2133–2151. [Google Scholar] [CrossRef]
- Rusakov, D.A. Disentangling calcium-driven astrocyte physiology. Nat. Rev. Neurosci. 2015, 16, 226–233. [Google Scholar] [CrossRef]
- Tanaka, M.; Shih, P.Y.; Gomi, H.; Yoshida, T.; Nakai, J.; Ando, R.; Furuichi, T.; Mikoshiba, K.; Semyanov, A.; Itohara, S. Astrocytic Ca2+ signals are required for the functional integrity of tripartite synapses. Mol. Brain 2013, 6, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.W.; Tang, X.; Arizono, M.; Bannai, H.; Shih, P.Y.; Dembitskaya, Y.; Kazantsev, V.; Tanaka, M.; Itohara, S.; Mikoshiba, K.; et al. Spatiotemporal calcium dynamics in single astrocytes and its modulation by neuronal activity. Cell Calcium 2014, 55, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Kustikova, V.; Krivonosov, M.; Pimashkin, A.; Denisov, P.; Zaikin, A.; Ivanchenko, M.; Meyerov, I.; Semyanov, A. CalciumCV: Computer vision software for calcium signaling in astrocytes. In Proceedings of the International Conference on Analysis of Images, Social Networks and Texts, Moscow, Russia, 5 July 2018; Springer: Cham, Switzerland, 2018; pp. 168–179. [Google Scholar]
- Matrosov, V.; Gordleeva, S.; Boldyreva, N.; Ben-Jacob, E.; Kazantsev, V.; Pittà, M.D. Emergence of regular and complex calcium oscillations by inositol 1, 4, 5-trisphosphate signaling in astrocytes. In Springer Series in Computational Neuroscience; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; pp. 151–176. [Google Scholar]
- Izhikevich, E.M. Simple model of spiking neurons. IEEE Trans. Neural Netw. 2003, 14, 1569–1572. [Google Scholar] [CrossRef]
- Kazantsev, V.B.; Asatryan, S.Y. Bistability induces episodic spike communication by inhibitory neurons in neuronal networks. Phys. Rev. E 2011, 84, 031913. [Google Scholar] [CrossRef]
- Kazantsev, V.; Gordleeva, S.; Stasenko, S.; Dityatev, A. A homeostatic model of neuronal fring governed by feedback signals from the extracellular matrix. PLoS ONE 2012, 7, e41646. [Google Scholar] [CrossRef]
- Gordleeva, S.Y.; Stasenko, S.V.; Semyanov, A.V.; Dityatev, A.E.; Kazantsev, V.B. Bi-directional astrocytic regulation of neuronal activity within a network. Front. Comput. Neurosci. 2012, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J. The inositol trisphosphate/calcium signaling pathway in health and disease. Physiol. Rev. 2016, 96, 1261–1296. [Google Scholar] [CrossRef]
- Berridge, M.J. Inositol trisphosphate and calcium signalling. Nature 1993, 361, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Nadkarni, S.; Jung, P. Modeling synaptic transmission of the tripartite synapse. Phys. Biol. 2007, 4, 1. [Google Scholar] [CrossRef]
- Oschmann, F.; Berry, H.; Obermayer, K.; Lenk, K. From in silico astrocyte cell models to neuron-astrocyte network models: A review. Brain Res. Bull. 2018, 136, 76–84. [Google Scholar] [CrossRef]
- Liu, L.; Gao, H.; Zaikin, A.; Chen, S. Unraveling Aβ-mediated multi-pathway calcium dynamics in astrocytes: Implications for Alzheimer’s Disease treatment from simulations. Front. Physiol. 2021, 12, 767892. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.W.; Gordleeva, S.; Tang, X.; Shih, P.Y.; Dembitskaya, Y.; Semyanov, A. Morphological profile determines the frequency of spontaneous calcium events in astrocytic processes. Glia 2019, 67, 246–262. [Google Scholar] [CrossRef] [PubMed]
- Ullah, G.; Jung, P.; Cornell-Bell, A.H. Anti-phase calcium oscillations in astrocytes via inositol (1, 4, 5)-trisphosphate regeneration. Cell Calcium 2006, 39, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Belanger, M.; Allaman, I.; Magistretti, P.J. Brain energy metabolism: Focus on astrocyte-neuron metabolic cooperation. Cell Metab. 2011, 14, 724–738. [Google Scholar] [CrossRef] [Green Version]
- Stobart, J.L.; Anderson, C.M. Multifunctional role of astrocytes as gatekeepers of neuronal energy supply. Front. Cell. Neurosci. 2013, 7, 38. [Google Scholar] [CrossRef]
- Kastanenka, K.V.; Moreno-Bote, R.; De Pittà, M.; Perea, G.; Eraso-Pichot, A.; Masgrau, R.; Poskanzer, K.E.; Galea, E. A roadmap to integrate astrocytes into Systems Neuroscience. Glia 2020, 68, 5–26. [Google Scholar] [CrossRef]
- Tavanaei, A.; Maida, A.S. Multi-layer unsupervised learning in a spiking convolutional neural network. In Proceedings of the 2017 International Joint Conference on Neural Networks (IJCNN), Anchorage, AK, USA, 14–19 May 2017; pp. 2023–2030. [Google Scholar]
- Lee, C.; Srinivasan, G.; Panda, P.; Roy, K. Deep spiking convolutional neural network trained with unsupervised spike-timing-dependent plasticity. IEEE Trans. Cogn. Dev. Syst. 2018, 11, 384–394. [Google Scholar]
- Tavanaei, A.; Ghodrati, M.; Kheradpisheh, S.R.; Masquelier, T.; Maida, A. Deep learning in spiking neural networks. Neural Netw. 2019, 111, 47–63. [Google Scholar] [CrossRef]
- Panatier, A.; Theodosis, D.T.; Mothet, J.P.; Touquet, B.; Pollegioni, L.; Poulain, D.A.; Oliet, S.H. Glia-derived D-serine controls NMDA receptor activity and synaptic memory. Cell 2006, 125, 775–784. [Google Scholar] [CrossRef]
- Pannasch, U.; Freche, D.; Dallérac, G.; Ghézali, G.; Escartin, C.; Ezan, P.; Cohen-Salmon, M.; Benchenane, K.; Abudara, V.; Dufour, A.; et al. Connexin 30 sets synaptic strength by controlling astroglial synapse invasion. Nat. Neurosci. 2014, 17, 549–558. [Google Scholar] [CrossRef]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Tsybina, Y.; Gordleeva, S.; Zaikin, A. Impact of Astrocytic Coverage of Synapses on the Short-Term Memory of a Computational Neuron-Astrocyte Network. Mathematics 2022, 10, 3275. https://0-doi-org.brum.beds.ac.uk/10.3390/math10183275
Li Z, Tsybina Y, Gordleeva S, Zaikin A. Impact of Astrocytic Coverage of Synapses on the Short-Term Memory of a Computational Neuron-Astrocyte Network. Mathematics. 2022; 10(18):3275. https://0-doi-org.brum.beds.ac.uk/10.3390/math10183275
Chicago/Turabian StyleLi, Zonglun, Yuliya Tsybina, Susanna Gordleeva, and Alexey Zaikin. 2022. "Impact of Astrocytic Coverage of Synapses on the Short-Term Memory of a Computational Neuron-Astrocyte Network" Mathematics 10, no. 18: 3275. https://0-doi-org.brum.beds.ac.uk/10.3390/math10183275