
Computational Model of Noradrenaline Modulation of Astrocyte Responses to Synaptic Activity


Abstract
:1. Introduction
2. The Model
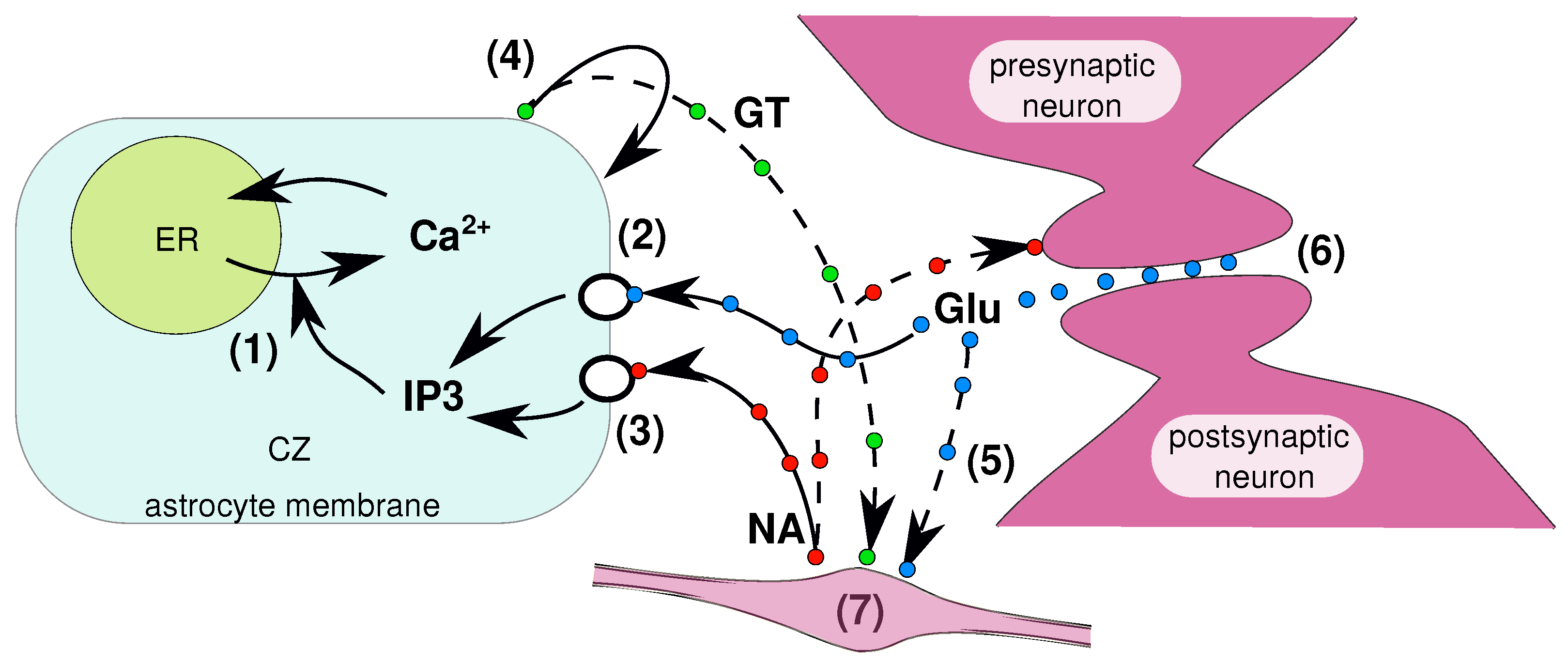
2.1. Pathways to Be Modeled
2.2. Topological Issues and AVF Approach

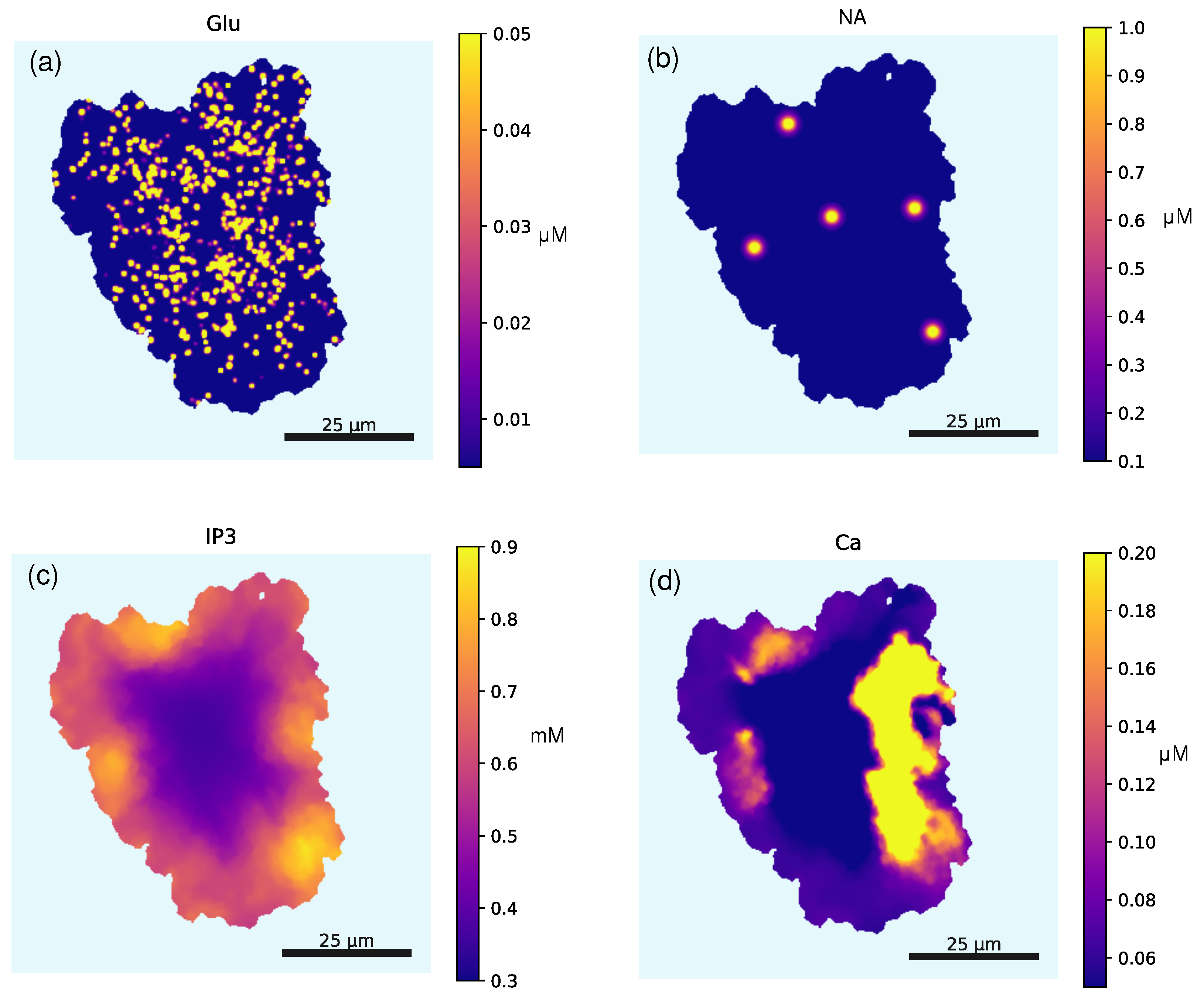
2.3. Randomly Seeded Synapses
- 1.
- We set the synaptic density to . This parameter reflects the maximum synapse coverage density in relative units.
- 2.
- 3.
- The probability of spawning a synapse at each point of the spatial template is given as
- 4.
- Finally, we use the defined probability to randomly pick the pixels where the synapses are located; then, these sites are enlarged to a 22 shape to match the effective area of our synapse representation.
2.4. Noradrenaline Pathways
2.5. Calcium Dynamics
3. Simulation Results
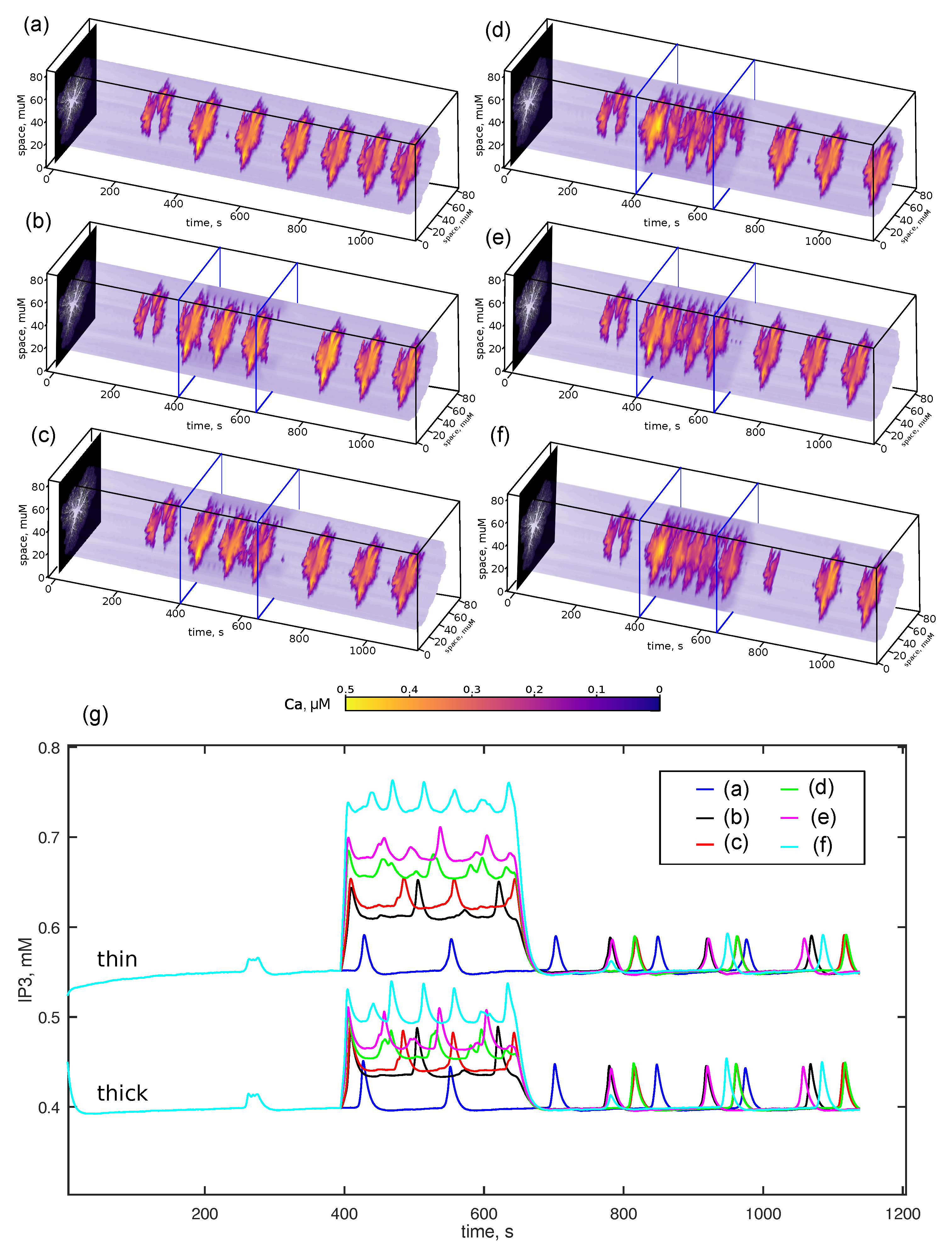
3.1. Dynamics Overview
3.2. Short Noradrenaline Surges Raise Gain of Astrocyte Responses to Glutamate
- (b)—, , corresponding to the additive effect of and Glu on IP production;
- (c)—, corresponding to cooperative interaction, increasing sensitivity to Glu;;
- (d)–(f)—additive NA effect combined with volume NA increases, simulated by setting NA to , , and m, respectively.
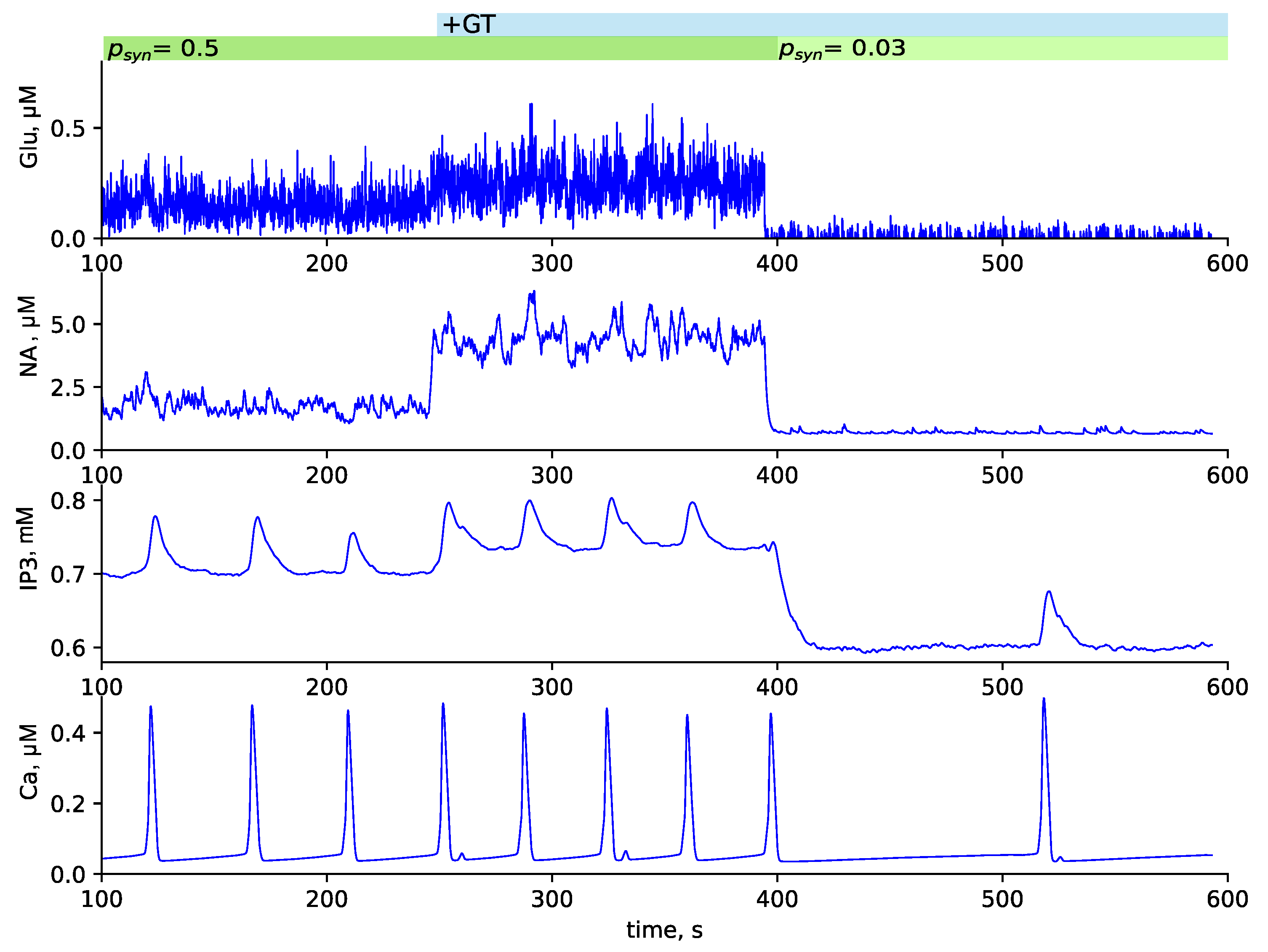
3.3. Testing GANE Hypothesis
4. Discussion
4.1. Noradrenaline and Glu: Synergy or Independent Effects?
4.2. Comparison between Spatially Confined and Volume NA Signal
4.3. GANE Mechanism Seems to Be Viable at Microscopic Scale
4.4. Additional Considerations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NVU | neurovascular unit |
| AVF | astrocyte volume fraction |
| SVR | surface–volume ratio |
| GANE | glutamate amplifies noradrenergic effects |
| ER | endoplasmic reticulum |
| LC | Locus Coeruleus |
| inositol trisphosphate | |
| Glu | glutamate |
| NA | noradrenaline |
| Gt | gliotransmitter |
| NMDA | N-methyl-D-aspartate |
| GPCR | G-protein-coupled receptors |
References
- Agnati, L.; Zoli, M.; Strömberg, I.; Fuxe, K. Intercellular communication in the brain: Wiring versus volume transmission. Neuroscience 1995, 69, 711–726. [Google Scholar] [CrossRef] [PubMed]
- Sykova, E.; Nicholson, C. Diffusion in brain extracellular space. Physiol. Rev. 2008, 88, 1277–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sykova, E. Extrasynaptic volume transmission and diffusion parameters of the extracellular space. Neuroscience 2004, 129, 861–876. [Google Scholar] [CrossRef] [PubMed]
- Semyanov, A.; Verkhratsky, A. Astrocytic processes: From tripartite synapses to the active milieu. Trends Neurosci. 2021, 44, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Araque, A.; Carmignoto, G.; Haydon, P.G. Dynamic signaling between astrocytes and neurons. Annu. Rev. Physiol. 2001, 63, 795–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volterra, A.; Meldolesi, J. Astrocytes, from brain glue to communication elements: The revolution continues. Nat. Rev. Neurosci. 2005, 6, 626–640. [Google Scholar] [CrossRef]
- Parpura, V.; Verkhratsky, A. Astrocytes revisited: Concise historic outlook on glutamate homeostasis and signaling. Croat. Med. J. 2012, 53, 518–528. [Google Scholar] [CrossRef] [Green Version]
- Volterra, A.; Magistretti, P.; Haydon, P. The Tripartite Synapse: Glia in Synaptic Transmission; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Rose, C.R.; Chatton, J.Y. Astrocyte sodium signaling and neuro-metabolic coupling in the brain. Neuroscience 2016, 323, 121–134. [Google Scholar] [CrossRef] [Green Version]
- Araque, A.; Carmignoto, G.; Haydon, P.G.; Oliet, S.H.; Robitaille, R.; Volterra, A. Gliotransmitters travel in time and space. Neuron 2014, 81, 728–739. [Google Scholar] [CrossRef] [Green Version]
- Muoio, V.; Persson, P.; Sendeski, M. The neurovascular unit—Concept review. Acta Physiol. 2014, 210, 790–798. [Google Scholar] [CrossRef]
- Iadecola, C. The neurovascular unit coming of age: A journey through neurovascular coupling in health and disease. Neuron 2017, 96, 17–42. [Google Scholar] [CrossRef] [Green Version]
- Schaeffer, S.; Iadecola, C. Revisiting the neurovascular unit. Nat. Neurosci. 2021, 24, 1198–1209. [Google Scholar] [CrossRef]
- Nadkarni, S.; Jung, P. Dressed neurons: Modeling neural-glial interactions. Phys. Biol. 2004, 1, 35–41. [Google Scholar] [CrossRef]
- Nadkarni, S.; Jung, P. Modeling synaptic transmission of the tripartite synapse. Phys. Biol. 2007, 4, 1. [Google Scholar] [CrossRef]
- Farr, H.; David, T. Models of neurovascular coupling via potassium and EET signalling. J. Theor. Biol. 2011, 286, 13–23. [Google Scholar] [CrossRef]
- Dormanns, K.; Brown, R.; David, T. The role of nitric oxide in neurovascular coupling. J. Theor. Biol. 2016, 394, 1–17. [Google Scholar] [CrossRef]
- Kenny, A.; Plank, M.J.; David, T. The role of astrocytic calcium and TRPV4 channels in neurovascular coupling. J. Comput. Neurosci. 2018, 44, 97–114. [Google Scholar] [CrossRef]
- Mathias, E.J.; Kenny, A.; Plank, M.J.; David, T. Integrated models of neurovascular coupling and BOLD signals: Responses for varying neural activations. NeuroImage 2018, 174, 69–86. [Google Scholar] [CrossRef]
- Gordleeva, S.Y.; Tsybina, Y.A.; Krivonosov, M.I.; Ivanchenko, M.V.; Zaikin, A.A.; Kazantsev, V.B.; Gorban, A.N. Modeling working memory in a spiking neuron network accompanied by astrocytes. Front. Cell. Neurosci. 2021, 15, 631485. [Google Scholar] [CrossRef]
- Tsybina, Y.; Kastalskiy, I.; Krivonosov, M.; Zaikin, A.; Kazantsev, V.; Gorban, A.N.; Gordleeva, S. Astrocytes mediate analogous memory in a multi-layer neuron–astrocyte network. Neural Comput. Appl. 2022, 34, 9147–9160. [Google Scholar] [CrossRef]
- Ayata, C.; Lauritzen, M. Spreading depression, spreading depolarizations, and the cerebral vasculature. Physiol. Rev. 2015, 95, 953–993. [Google Scholar] [CrossRef] [Green Version]
- Dahlem, M.A.; Graf, R.; Strong, A.J.; Dreier, J.P.; Dahlem, Y.A.; Sieber, M.; Hanke, W.; Podoll, K.; Schöll, E. Two-dimensional wave patterns of spreading depolarization: Retracting, re-entrant, and stationary waves. Phys. D Nonlinear Phenom. 2010, 239, 889–903. [Google Scholar] [CrossRef] [Green Version]
- Postnov, D.; Postnov, D.; Schimansky-Geier, L. Self-terminating wave patterns and self-organized pacemakers in a phenomenological model of spreading depression. Brain Res. 2012, 1434, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Verisokin, A.Y.; Verveyko, D.; Postnov, D. Turing-like structures in a functional model of cortical spreading depression. Phys. Rev. E 2017, 96, 062409. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; O’Donnell, J.; Xu, Q.; Kang, N.; Goldman, N.; Nedergaard, M. Changes in the composition of brain interstitial ions control the sleep-wake cycle. Science 2016, 352, 550–555. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, J.; Ding, F.; Nedergaard, M. Distinct functional states of astrocytes during sleep and wakefulness: Is norepinephrine the master regulator? Curr. Sleep Med. Rep. 2015, 1, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ingiosi, A.M.; Frank, M.G. Goodnight, astrocyte: Waking up to astroglial mechanisms in sleep. FEBS J. 2022. [Google Scholar] [CrossRef]
- Haydon, P.G. Astrocytes and the modulation of sleep. Curr. Opin. Neurobiol. 2017, 44, 28–33. [Google Scholar] [CrossRef]
- Savtchenko, L.P.; Bard, L.; Jensen, T.P.; Reynolds, J.P.; Kraev, I.; Medvedev, N.; Stewart, M.G.; Henneberger, C.; Rusakov, D.A. Disentangling astroglial physiology with a realistic cell model in silico. Nat. Commun. 2018, 9, 3554. [Google Scholar] [CrossRef]
- Matyash, V.; Kettenmann, H. Heterogeneity in astrocyte morphology and physiology. Brain Res. Rev. 2010, 63, 2–10. [Google Scholar] [CrossRef]
- Khakh, B.S.; Sofroniew, M.V. Diversity of astrocyte functions and phenotypes in neural circuits. Nat. Neurosci. 2015, 18, 942–952. [Google Scholar] [CrossRef]
- Verisokin, A.; Verveyko, D.; Postnov, D.; Brazhe, A. Modeling of Astrocyte Networks: Toward Realistic Topology and Dynamics. Front. Cell. Neurosci. 2021, 15, 50. [Google Scholar] [CrossRef]
- Verveyko, D.V.; Verisokin, A.Y.; Postnov, D.E.; Brazhe, A.R. Connectivity promotes repeatable activation patterns in the model of astrocytic networks. Eur. Phys. J. Plus 2021, 136, 732. [Google Scholar] [CrossRef]
- Paukert, M.; Agarwal, A.; Cha, J.; Doze, V.A.; Kang, J.U.; Bergles, D.E. Norepinephrine controls astroglial responsiveness to local circuit activity. Neuron 2014, 82, 1263–1270. [Google Scholar] [CrossRef] [Green Version]
- Ingiosi, A.M.; Frank, M.G. Noradrenergic Signaling in Astrocytes Influences Mammalian Sleep Homeostasis. Clocks Sleep 2022, 4, 332–345. [Google Scholar] [CrossRef]
- Mander, B.A.; Winer, J.R.; Jagust, W.J.; Walker, M.P. Sleep: A novel mechanistic pathway, biomarker, and treatment target in the pathology of Alzheimer’s disease? Trends Neurosci. 2016, 39, 552–566. [Google Scholar] [CrossRef] [Green Version]
- Semyachkina-Glushkovskaya, O.; Postnov, D.; Penzel, T.; Kurths, J. Sleep as a novel biomarker and a promising therapeutic target for cerebral small vessel disease: A review focusing on Alzheimer’s disease and the blood-brain barrier. Int. J. Mol. Sci. 2020, 21, 6293. [Google Scholar] [CrossRef]
- Mather, M.; Clewett, D.; Sakaki, M.; Harley, C.W. Norepinephrine ignites local hotspots of neuronal excitation: How arousal amplifies selectivity in perception and memory. Behav. Brain Sci. 2016, 39. [Google Scholar] [CrossRef] [Green Version]
- Kimelberg, H.K.; Norenberg, M.D. Astrocytes. Sci. Am. 1989, 260, 66–77. [Google Scholar] [CrossRef]
- Keener, J.; Sneyd, J. Mathematical Physiology 1: Cellular Physiology; Springer: New York, NY, USA, 2009; Volume 2. [Google Scholar]
- Houart, G.; Dupont, G.; Goldbeter, A. Bursting, chaos and birhythmicity originating from self-modulation of the inositol 1, 4, 5-trisphosphate signal in a model for intracellular Ca2+ oscillations. Bull. Math. Biol. 1999, 61, 507–530. [Google Scholar] [CrossRef]
- Bushong, E.A.; Martone, M.E.; Ellisman, M.H. Maturation of astrocyte morphology and the establishment of astrocyte domains during postnatal hippocampal development. Int. J. Dev. Neurosci. 2004, 22, 73–86. [Google Scholar] [CrossRef]
- Martone, M.E.; Zhang, S.; Gupta, A.; Qian, X.; He, H.; Price, D.L.; Wong, M.; Santini, S.; Ellisman, M.H. The cell-centered database. Neuroinformatics 2003, 1, 379–395. [Google Scholar] [CrossRef] [PubMed]
- Héja, L.; Szabó, Z.; Péter, M.; Kardos, J. Spontaneous Ca2+ Fluctuations Arise in Thin Astrocytic Processes With Real 3D Geometry. Front. Cell. Neurosci. 2021, 15, 617989. [Google Scholar] [CrossRef] [PubMed]
- Genoud, C.; Knott, G.W.; Sakata, K.; Lu, B.; Welker, E. Altered synapse formation in the adult somatosensory cortex of brain-derived neurotrophic factor heterozygote mice. J. Neurosci. 2004, 24, 2394–2400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santuy, A.; Tomás-Roca, L.; Rodríguez, J.R.; González-Soriano, J.; Zhu, F.; Qiu, Z.; Grant, S.; DeFelipe, J.; Merchan-Perez, A. Estimation of the number of synapses in the hippocampus and brain-wide by volume electron microscopy and genetic labeling. Sci. Rep. 2020, 10, 14014. [Google Scholar] [CrossRef]
- Verisokin, A.; Kirsanov, A.; Verveyko, D.; Postnov, D.; Brazhe, A. Poppy-seeding synapses within astrocytic domains: The role of synaptic spatial patterns in the astrocyte-neuron communication. In Proceedings of the Computational Biophysics and Nanobiophotonics; Postnov, D.E., Khlebtsov, B.N., Eds.; International Society for Optics and Photonics, SPIE: Bellingham, WA, USA, 2022; Volume 12194, p. 121940A. [Google Scholar] [CrossRef]
- Moreno-Bote, R. Poisson-like spiking in circuits with probabilistic synapses. PLoS Comput. Biol. 2014, 10, e1003522. [Google Scholar] [CrossRef] [Green Version]
- Burkitt, A.N. A review of the integrate-and-fire neuron model: II. Inhomogeneous synaptic input and network properties. Biol. Cybern. 2006, 95, 97–112. [Google Scholar] [CrossRef]
- Brazhe, A.R.; Postnov, D.E.; Sosnovtseva, O. Astrocyte calcium signaling: Interplay between structural and dynamical patterns. Chaos 2018, 28, 106320. [Google Scholar] [CrossRef]
- Tass, P.A. Phase Resetting in Medicine and Biology: Stochastic Modelling and Data Analysis; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Gray, S.R.; Ye, L.; Ye, J.Y.; Paukert, M. Noradrenergic terminal short-term potentiation enables modality-selective integration of sensory input and vigilance state. Sci. Adv. 2021, 7, eabk1378. [Google Scholar] [CrossRef]
- Ryzhov, S.; Goldstein, A.E.; Biaggioni, I.; Feoktistov, I. Cross-talk between G(s)- and G(q)-coupled pathways in regulation of interleukin-4 by A(2B) adenosine receptors in human mast cells. Mol. Pharmacol. 2006, 70, 727–735. [Google Scholar] [CrossRef]
- Pikovsky, A.S.; Kurths, J. Coherence resonance in a noise-driven excitable system. Phys. Rev. Lett. 1997, 78, 775. [Google Scholar] [CrossRef] [Green Version]
- Perc, M. Spatial coherence resonance in excitable media. Phys. Rev. E 2005, 72, 016207. [Google Scholar] [CrossRef] [Green Version]
- Fuxe, K.; Agnati, L.F.; Marcoli, M.; Borroto-Escuela, D.O. Volume Transmission in Central Dopamine and Noradrenaline Neurons and Its Astroglial Targets. Neurochem. Res. 2015, 40, 2600–2614. [Google Scholar] [CrossRef]
- Feng, J.; Zhang, C.; Lischinsky, J.E.; Jing, M.; Zhou, J.; Wang, H.; Zhang, Y.; Dong, A.; Wu, Z.; Wu, H.; et al. A Genetically Encoded Fluorescent Sensor for Rapid and Specific In Vivo Detection of Norepinephrine. Neuron 2019, 102, 745–761.e8. [Google Scholar] [CrossRef]
- Wahis, J.; Holt, M.G. Astrocytes, Noradrenaline, α1-Adrenoreceptors, and Neuromodulation: Evidence and Unanswered Questions. Front. Cell. Neurosci. 2021, 15, 645691. [Google Scholar] [CrossRef]
- Covelo, A.; Araque, A. Neuronal activity determines distinct gliotransmitter release from a single astrocyte. Elife 2018, 7, e32237. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Set 1 | Set 2 | Set 3 | Set 4 | Description |
|---|---|---|---|---|---|
| , Ms | 1 | 0; 1 | 0; 1 | 1 | NA influx in some points |
| , s | 50 | 0; 50 | 0; 50 | 50 | Glu influence on NA dynamics |
| , M | 0 | 0; 1 | 0 | 0 | Synergetic dependence on NA of the Glu-dependent IP3 production |
| , s | 0 | 0 | 0 | 0; 5 | Gt influence on NA dynamics |
| , M | 0 | 0 | 0 | 0.5 | NA influence on the probability of Glu release |
| , M s | 0.1 | 0; 0.1 | 0.1 | 0.1 | rate of IP3 production through NA |
| , M | 0 | 0–0.01 | 0 | 0 | NA ambient |
| , Hz | 0.5 | 0.5 | 0.5 | 0.0(3); 0.5 | Glu release probability from synapses (number of events per unit time) |
| Parameter | Value | Description |
|---|---|---|
| 0.185 | ER/cytosolic volume ratio | |
| 6.0 s | Maximum Ca channel flux | |
| 0.13 M | Dissociation constant for IP | |
| 0.082 M | Ca activation constant | |
| 0.11 s | Ca leak constant | |
| 2.2 M/s | Maximum Ca uptake | |
| 1.0 s | Rate constant of Ca extrusion | |
| 0.01 M/s | Rate of Ca leak across plasma membrane | |
| 0.2 M/s | Maximal rate of activation-dependent Ca influx | |
| 1.0 M | Half-saturation constant for agonist-dependent Ca entry | |
| 0.1 M | Activation constant for Ca-pump | |
| 0.25 M/s | Maximal rate of IP production | |
| 1.1 M | Dissociation constant for Ca stimulation of IP production | |
| 0.78 M/s | Dissociation constant for Glu stimulation of IP production | |
| 1.049 M | Dissociation constant for Ca inhibition | |
| 0.8 Ms | Dissociation constant of NA stimulation of IP3 | |
| 0.1 Ms | Dissociation constant of Ca stimulation of Gt | |
| 0.1 M/s | Rate of IP production through Glu | |
| 7.143 s | Rate constant for loss of IP | |
| 0.16 M | Steady-state IP | |
| 0 M | Steady-state Gt | |
| 0.943 M | Receptor dissociation constant for IP | |
| 0.14 Ms | Ca inhibition constant | |
| 1.0 Ms | Diffusion coefficient for Ca | |
| IP3 | 10.0 Ms | Diffusion coefficient for IP |
| 0.4 Ms | Diffusion coefficient for NA | |
| 0.1 Ms | Diffusion coefficient for Glu | |
| 0.1 Ms | Diffusion coefficient for Gt | |
| 0.1 Ms | Gt production rate through Ca | |
| 15 s | Gt loss rate constant | |
| 1.8 s | NA loss rate constant |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verisokin, A.; Verveyko, D.; Kirsanov, A.; Brazhe, A.; Postnov, D. Computational Model of Noradrenaline Modulation of Astrocyte Responses to Synaptic Activity. Mathematics 2023, 11, 628. https://0-doi-org.brum.beds.ac.uk/10.3390/math11030628
Verisokin A, Verveyko D, Kirsanov A, Brazhe A, Postnov D. Computational Model of Noradrenaline Modulation of Astrocyte Responses to Synaptic Activity. Mathematics. 2023; 11(3):628. https://0-doi-org.brum.beds.ac.uk/10.3390/math11030628
Chicago/Turabian StyleVerisokin, Andrey, Darya Verveyko, Artem Kirsanov, Alexey Brazhe, and Dmitry Postnov. 2023. "Computational Model of Noradrenaline Modulation of Astrocyte Responses to Synaptic Activity" Mathematics 11, no. 3: 628. https://0-doi-org.brum.beds.ac.uk/10.3390/math11030628