MS/MS-Based Molecular Networking Approach for the Detection of Aplysiatoxin-Related Compounds in Environmental Marine Cyanobacteria

 , and
, and
Abstract
:1. Introduction
2. Results
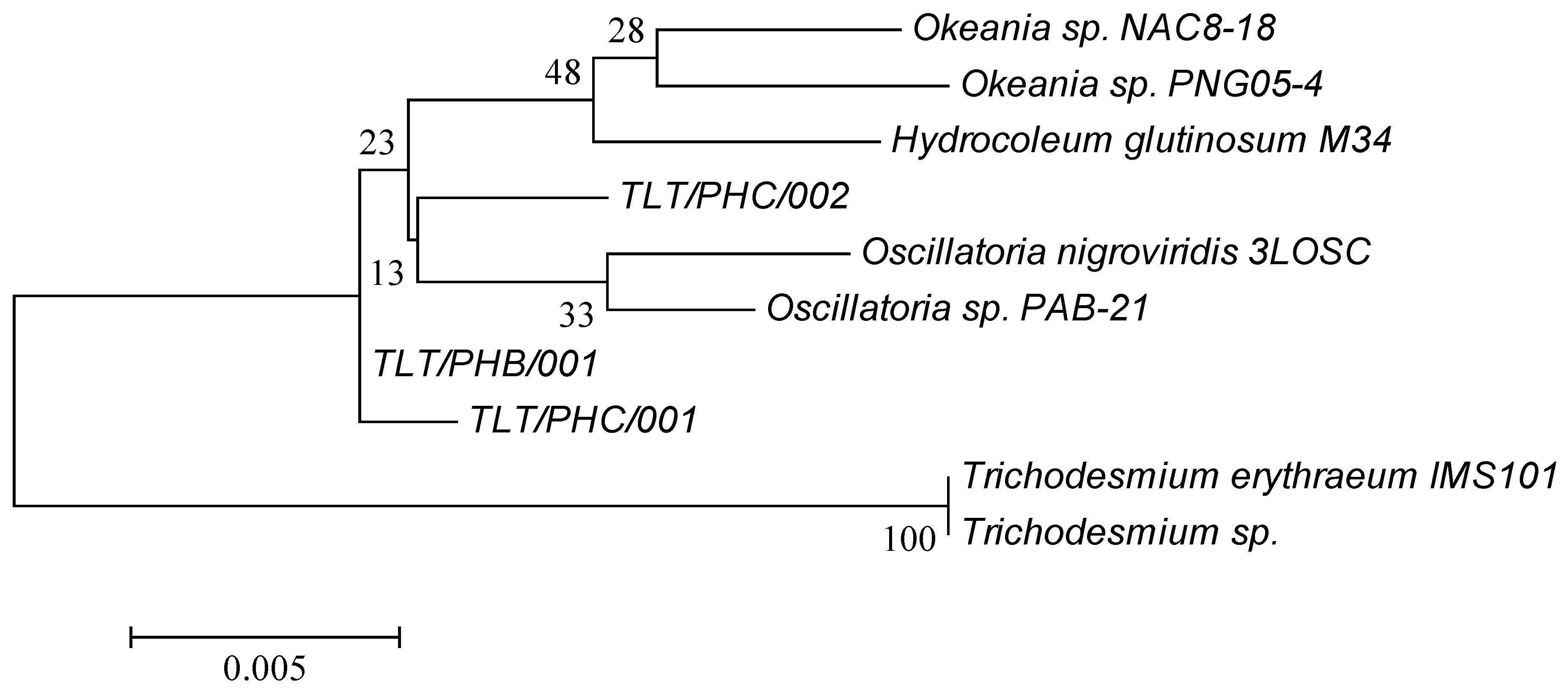
2.1. Phylogenetic Analysis of Environmental Marine Cyanobacterial Samples
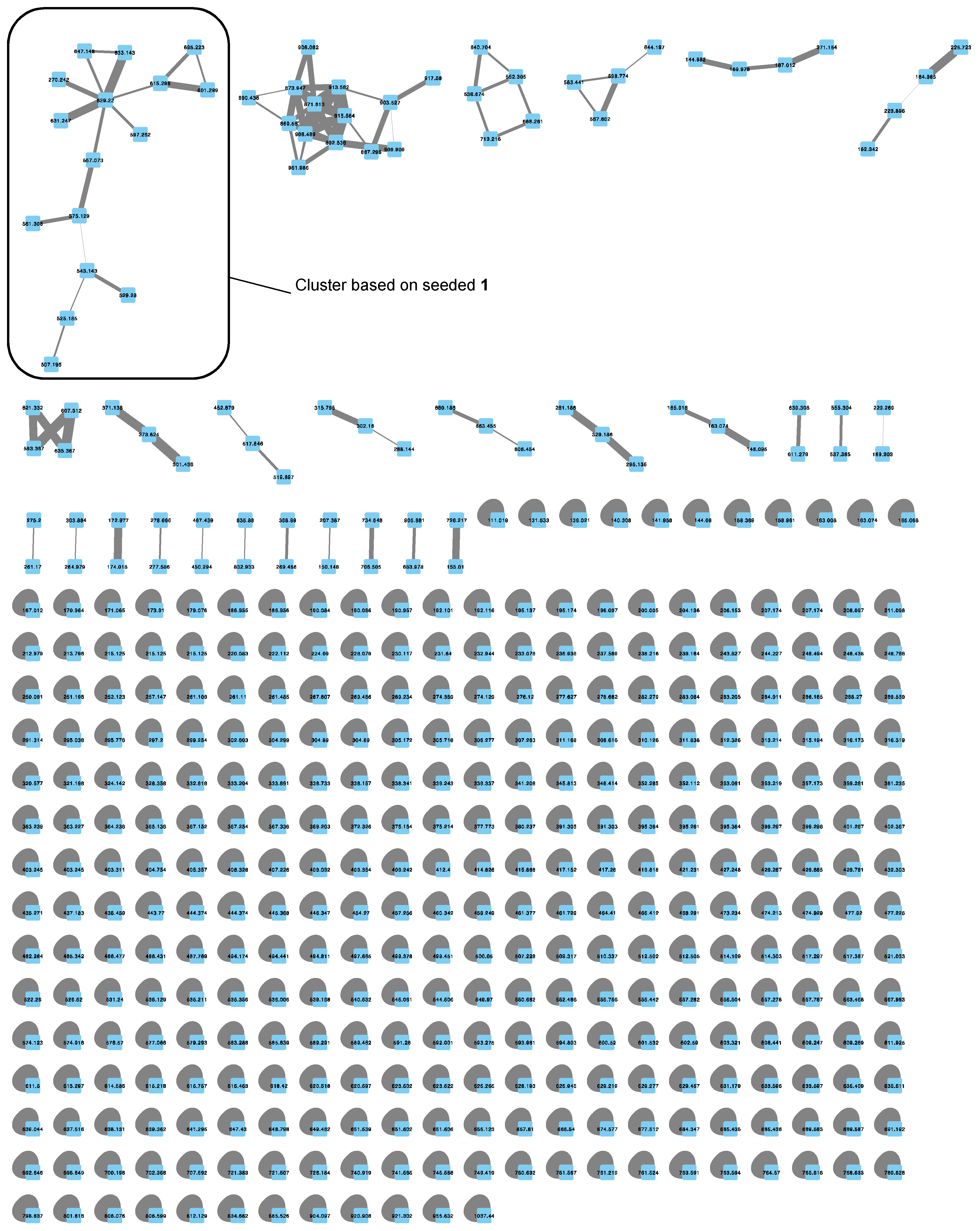
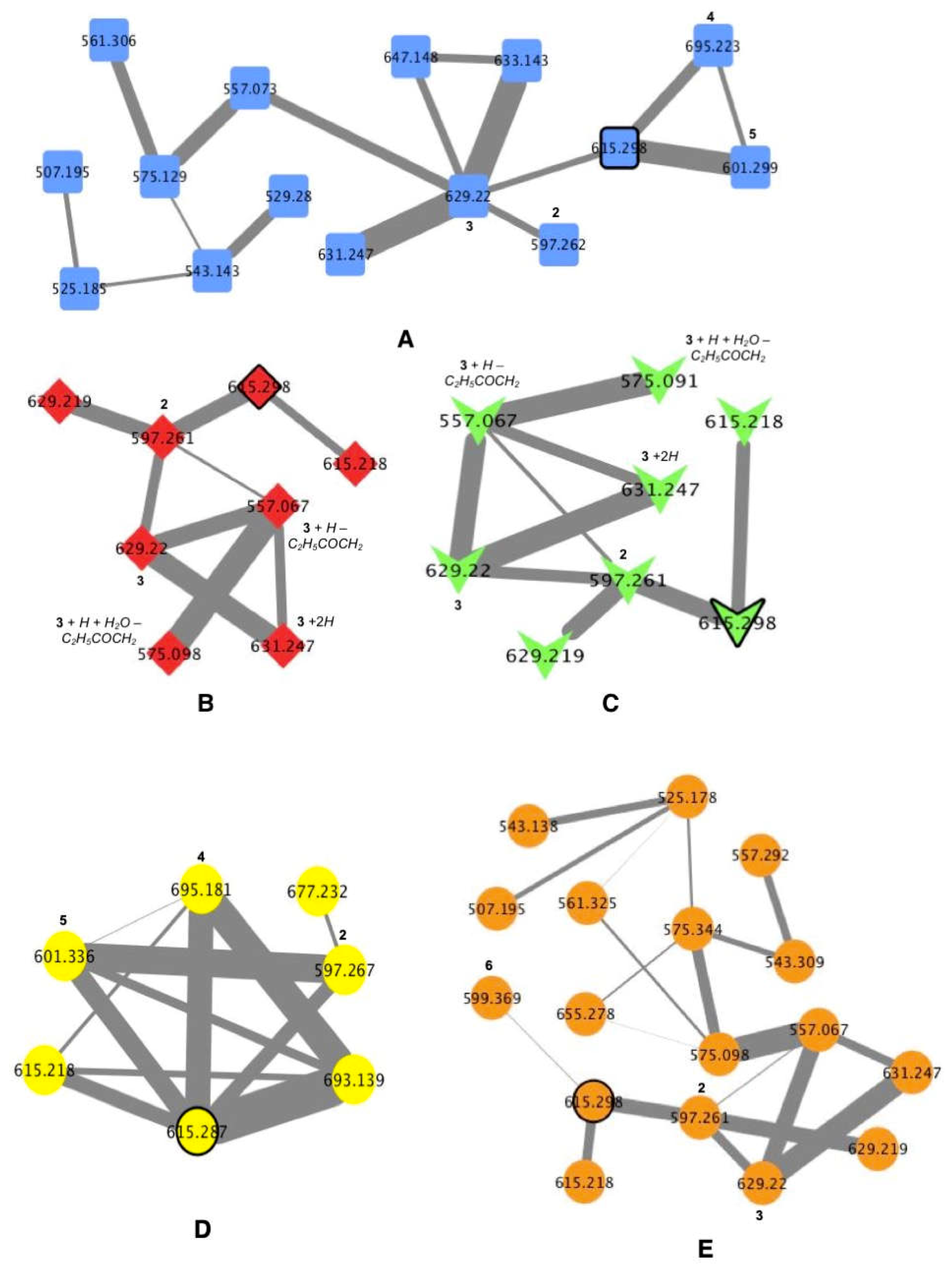
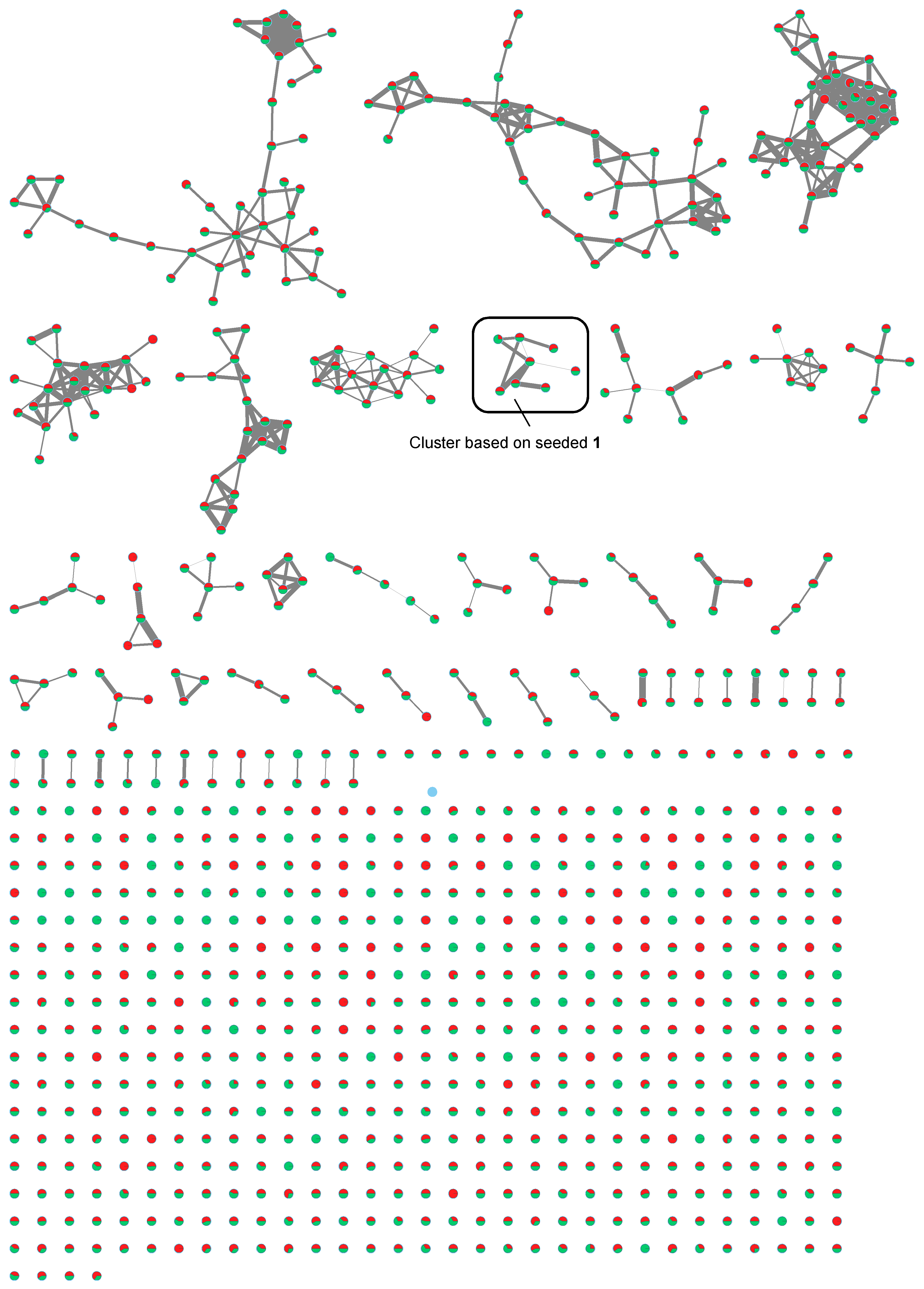
2.2. MS/MS-Based Molecular Networks of Marine Cyanobacterial Extracts
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Marine Cyanobacterial Collections
4.3. DNA Isolation and PCR Amplification
4.4. Phylogenetic Analysis
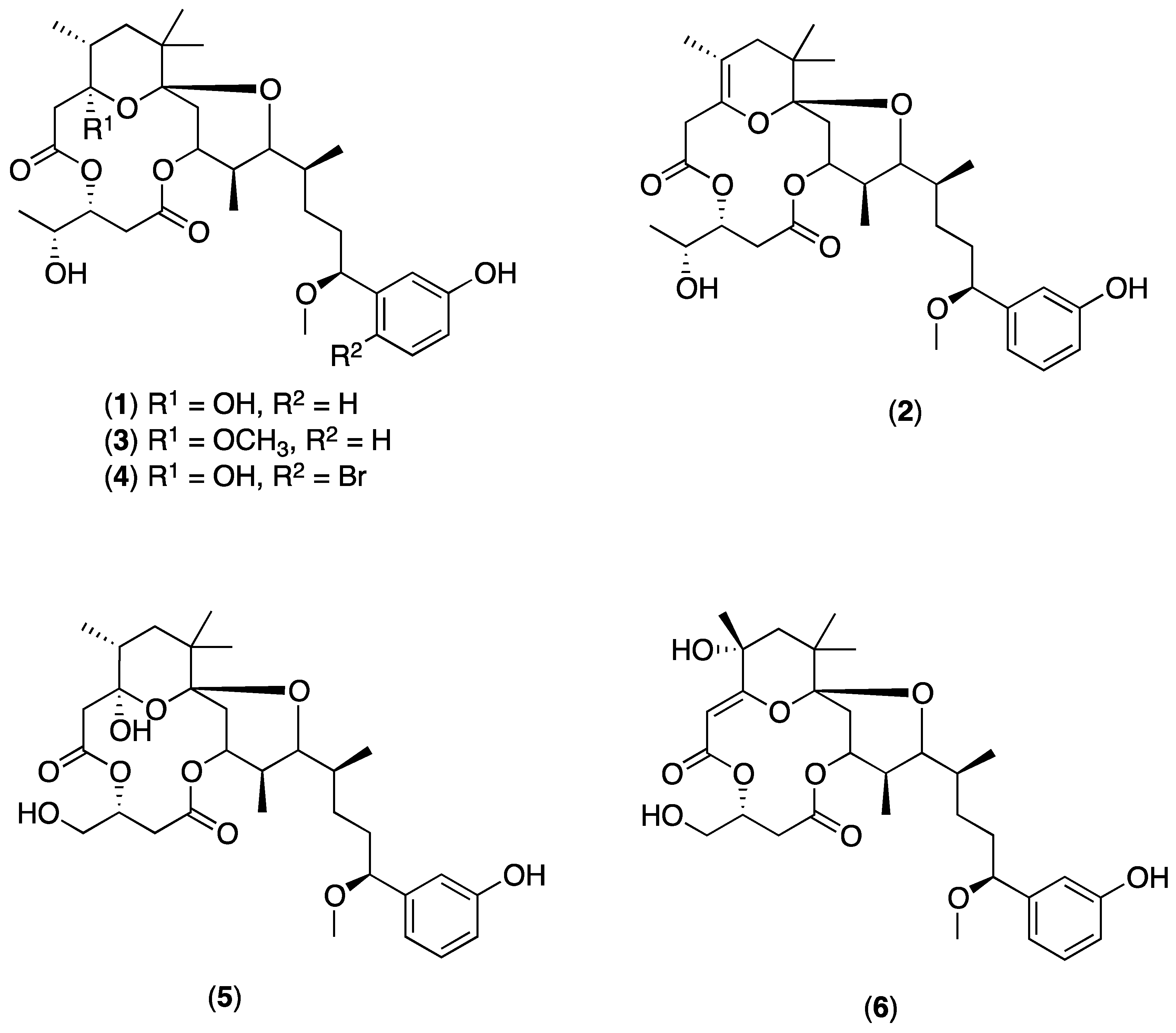
4.5. Extraction of Crude Extracts and Isolation of Debromoaplysiatoxin (1)
4.6. UPLC-HRMS/MS Analysis
4.7. HPLC-MS/MS Analysis
4.8. Molecular Networking Creation
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tan, L.T. Bioactive natural products from marine cyanobacteria for drug discovery. Phytochemistry 2007, 7, 954–979. [Google Scholar] [CrossRef]
- Abed, R.M.; Dobretsov, S.; Sudesh, K. Applications of cyanobacteria in biotechnology. J. Appl. Microbiol. 2009, 1, 1–12. [Google Scholar] [CrossRef]
- Ducat, D.C.; Way, J.C.; Silver, P.A. Engineering cyanobacteria to generate high-value products. Trends Biotechnol. 2011, 2, 95–103. [Google Scholar] [CrossRef]
- Pettit, G.R.; Kamano, Y.; Herald, C.L.; Tuinman, A.A.; Boettner, F.E.; Kizu, H.; Schmidt, J.M.; Baczynskyj, L.; Tomer, K.B.; Bontems, R.J. The isolation and structure of a remarkable marine animal antineoplastic constituent: dolastatin 10. J. Am. Chem. Soc. 1987, 22, 6883–6885. [Google Scholar] [CrossRef]
- Ju, W.; Zhang, M.; Wilson, K.M.; Petrus, M.N.; Bamford, R.N.; Zhang, X.; Guha, R.; Ferrer, M.; Thomas, C.J.; Waldmann, T.A. Augmented efficacy of Brentuximab Vedotin combined with Ruxolitinib and/or Navitoclax in a murine model of human Hodgkin’s lymphoma. Proc. Natl. Acad. Sci. USA 2016, 6, 1624–1629. [Google Scholar] [CrossRef]
- Pearson, L.A.; Dittmann, E.; Mazmouz, R.; Ongley, S.E.; D’Agostino, P.M.; Neilan, B.A. The genetics, biosynthesis and regulation of toxic specialized metabolites of cyanobacteria. Harmful Algae 2016, 54, 98–111. [Google Scholar] [CrossRef]
- Zanchett, G.; Oliveira-Filho, E.C. Cyanobacteria and cyanotoxins: from impacts on aquatic ecosystems and human health to anticarcinogenic effects. Toxins 2013, 10, 1896–1917. [Google Scholar] [CrossRef]
- Nagarajan, M.; Maruthanayagam, V.; Sundararaman, M. A review of pharmacological and toxicological potentials of marine cyanobacterial metabolites. J. Appl. Toxicol. 2012, 3, 153–185. [Google Scholar] [CrossRef]
- Yanagita, R.C.; Kamachi, H.; Tanaka, K.; Murakami, A.; Nakagawa, Y.; Tokuda, H.; Nagai, H.; Irie, K. Role of the phenolic hydroxyl group in the biological activities of simplified analogue of aplysiatoxin with antiproliferative activity. Bioorg. Med. Chem. Lett. 2010, 20, 6064–6066. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, Y.; Kikumori, M.; Yanagita, R.; Murakami, A.; Tokuda, H.; Nagai, H.; Irie, K. Synthesis and biological evaluation of the 12,12-dimethyl derivative of aplog-1, an anti-proliferative analog of tumor-promoting aplysiatoxin. Biosci. Biotechnol. Biochem. 2011, 75, 1167–1173. [Google Scholar] [CrossRef]
- Kamachi, H.; Tanaka, K.; Yanagita, R.C.; Murakami, A.; Murakami, K.; Tokuda, H.; Suzuki, N.; Nakagawa, Y.; Irie, K. Structure-activity studies on the side chain of a simplified analog of aplysiatoxin (aplog-1) with anti-proliferative activity. Bioorg. Med. Chem. 2013, 10, 2695–2702. [Google Scholar] [CrossRef]
- Kikumori, M.; Yanagita, R.C.; Tokuda, H.; Suzuki, N.; Nagai, H.; Suenaga, K.; Irie, K. Structure-activity studies on the spiroketal moiety of a simplified analogue of debromoaplysiatoxin with antiproliferative activity. J. Med. Chem. 2012, 11, 5614–5626. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Yanagita, R.C.; Hamada, N.; Murakami, A.; Takahashi, H.; Saito, N.; Nagai, H.; Irie, K. A simple analogue of tumor-promoting aplysiatoxin is an antineoplastic agent rather than a tumor promoter: Development of a synthetically accessible protein kinase C activator with bryostatin-like activity. J. Am. Chem. Soc. 2009, 22, 7573–7579. [Google Scholar] [CrossRef]
- Gupta, D.K.; Kaur, P.; Leong, S.T.; Tan, L.T.; Prinsep, M.R.; Chu, J.J. Anti-Chikungunya viral activities of aplysiatoxin-related compounds from the marine cyanobacterium Trichodesmium erythraeum. Mar. Drugs 2014, 1, 115–127. [Google Scholar] [CrossRef]
- Han, B.-N.; Liang, T.-T.; Keen, L.J.; Fan, T.-T.; Zhang, X.-D.; Xu, L.; Zhao, Q.; Wang, S.-P.; Lin, H.-W. Two marine cyanobacterial aplysiatoxin polyketides, neo-debromoaplysiatoxin A and B, with K+ channel inhibition activity. Org. Lett. 2018, 3, 578–581. [Google Scholar] [CrossRef]
- Richard, K.; Williams, D.; de Silva, E.; Brockman, M.; Brumme, Z.; Andersen, R.; Tietjen, I. Identification of novel Hiv-1 latency-reversing agents from a library of marine natural products. Viruses 2018, 10, 348. [Google Scholar] [CrossRef]
- Kaushik, R.; Balasubramanian, R. Methods and approaches used for detection of cyanotoxins in environmental samples: a review. Crit. Rev. Environ. Sci. Technol. 2013, 13, 1349–1383. [Google Scholar] [CrossRef]
- Nagai, H.; Kan, Y.; Fujita, T.; Sakamoto, B.; Hokama, Y. Manauealide C and anhydrodebromoaplysiatoxin, toxic constituents of the Hawaiian red alga, Gracilaria coronopifolia. Biosci. Biotechnol. Biochem. 1998, 5, 1011–1013. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef]
- Nguyen, D.D.; Wu, C.-H.; Moree, W.J.; Lamsa, A.; Medema, M.H.; Zhao, X.; Gavilan, R.G.; Aparicio, M.; Atencio, L.; Jackson, C.; et al. MS/MS networking guided analysis of molecule and gene cluster families. Proc. Nat. Acad. Sci. U.S.A. 2013, 28, E2611–E2620. [Google Scholar] [CrossRef]
- Gerwick, W.H. The face of a molecule. J. Nat. Prod. 2017, 9, 2583–2588. [Google Scholar] [CrossRef]
- Bertin, M.J.; Schwartz, S.L.; Lee, J.; Korobeynikov, A.; Dorrestein, P.C.; Gerwick, L.; Gerwick, W.H. Spongosine production by a Vibrio harveyi strain associated with the sponge Tectitethya crypta. J. Nat. Prod. 2015, 3, 493–499. [Google Scholar] [CrossRef]
- Teta, R.; Della Sala, G.; Glukhov, E.; Gerwick, L.; Gerwick, W.H.; Mangoni, A.; Costantino, V. Combined LC-MS/MS and molecular networking approach reveals new cyanotoxins from the 2014 cyanobacterial bloom in Green Lake, Seattle. Environ. Sci. Technol. 2015, 24, 14301–14310. [Google Scholar] [CrossRef]
- Briand, E.; Bormans, M.; Gugger, M.; Dorrestein, P.C.; Gerwick, W.H. Changes in secondary metabolic profiles of Microcystis aeruginosa strains in response to intraspecific interactions. Environ. Microbiol. 2015, 2, 384–400. [Google Scholar] [CrossRef]
- Benke, P.I.; Vinay Kumar, M.C.S.; Pan, D.; Swarup, S. A mass spectrometry-based unique fragment approach for the identification of microcystins. Analyst 2015, 4, 1198–1206. [Google Scholar] [CrossRef]
- Te, S.H.; Chen, E.Y.; Gin, K.Y.-H. Comparison of quantitative PCR and droplet digital PCR multiplex assays for two genera of bloom-forming cyanobacteria, Cylindrospermopsis and Microcystis. Appl. Environ. Microl. 2015, 15, 5203–5211. [Google Scholar] [CrossRef]
- Birungi, G.; Yau Li, S.F. Determination of cyanobacterial cyclic peptide hepatotoxins in drinking water using CE. Electrophoresis 2009, 15, 2737–2742. [Google Scholar] [CrossRef]
- Graca, A.P.; Bondoso, J.; Gaspar, H.; Xavier, J.R.; Monteiro, M.C.; de la Cruz, M.; Oves-Costales, D.; Vicente, F.; Lage, O.M. Antimicrobial activity of heterotrophic bacterial communities from the marine sponge Erylus discophorus (Astrophorida, Geodiidae). PLoS ONE 2013, 11, e78992. [Google Scholar]
- Mincer, T.J.; Jensen, P.R.; Kauffman, C.A.; Fenical, W. Widespread and persistent populations of a major new marine actinomycete taxon in ccean sediments. Appl. Environ. Microbiol. 2002, 10, 5005–5011. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids. Res. 2004, 5, 1792–1797. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA 6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 12, 2725–2729. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster | Compound | Molecular Formula of Observed [M + Na]+ | Calculated m/z | Observed m/z | aΔ (ppm) |
|---|---|---|---|---|---|
| A | 1 | C32H48O10Na | 615.2987 | 615.2988 | +0.1 |
| 2 | C32H46O9Na | 597.3040 | 597.3047 | +1.1 | |
| 3 | C33H50O10Na | 629.3302 | 629.3308 | +0.9 | |
| 4 | C32H47O10BrNa | 695.2236 | 695.2227 | −0.4 | |
| 5 | C31H46O10Na | 601.2989 | 601.2982 | −1.2 | |
| B | 1 | C32H48O10Na | 615.2987 | 615.3005 | +2.9 |
| 2 | C32H46O9Na | 597.3040 | 597.3024 | −2.7 | |
| 3 | C33H50O10Na | 629.3302 | 629.3307 | +0.8 | |
| C | 1 | C32H48O10Na | 615.2987 | 615.2986 | −0.1 |
| 2 | C32H46O9Na | 597.3040 | 597.3058 | +3.0 | |
| 3 | C33H50O10Na | 629.3302 | 629.3290 | −1.9 | |
| E | 1 | C32H48O10Na | 615.2987 | 615.2985 | −0.3 |
| 2 | C32H46O9Na | 597.3040 | 597.3027 | −2.1 | |
| 3 | C33H50O10Na | 629.3302 | 629.3308 | +0.9 | |
| 6 | C31H44O10Na | 599.2832 | 599.2843 | +1.8 |
| Compound | Molecular Formula | Molecular Mass | Calculated m/z [M + H]+ | Calculated m/z [M + Na]+ |
|---|---|---|---|---|
| Aplysiatoxin | C32H47O10Br | 671.621 | 672.622 | 694.604 |
| Debromoaplysiatoxin | C32H48O10 | 592.725 | 593.727 | 615.709 |
| Oscillatoxin A | C31H46O10 | 578.698 | 579.700 | 601.682 |
| Anhydro-19-bromoaplysiatoxin | C32H44O9Br2 | 732.502 | 733.502 | 755.484 |
| Anhydro-19,21-dibromoaplysiatoxin | C32H43O9Br3 | 811.398 | 812.398 | 834.380 |
| 17-Bromooscillatoxin A | C31H45O10Br | 657.594 | 658.595 | 680.577 |
| 17,19-Dibromooscillatoxin A | C31H44O10Br2 | 736.491 | 737.491 | 759.473 |
| 19-Bromoaplysiatoxin | C32H46O10Br2 | 750.517 | 751.518 | 773.500 |
| Oscillatoxin B1 | C32H46O10 | 590.709 | 591.711 | 613.692 |
| Oscillatoxin B2 | C32H46O10 | 590.709 | 591.711 | 613.692 |
| 31-Noroscillatoxin B | C31H44O10 | 576.683 | 577.684 | 599.666 |
| Oscillatoxin D | C31H42O8 | 542.668 | 543.670 | 565.651 |
| 3-Methoxyaplysiatoxin | C33H49O10Br | 685.641 | 686.648 | 709.639 |
| 3-Methoxydebromoaplysiatoxin | C32H46O10 | 606.745 | 607.753 | 629.735 |
| Nhatrangin A | C21H32O8 | 412.474 | 413.482 | 435.465 |
| Nhatrangin B | C21H31O8Br | 491.370 | 492.378 | 514.360 |
| Anhydrodebromoaplysiatoxin | C32H46O9 | 574.702 | 575.711 | 597.693 |
| 30-Methyloscillatoxin D | C32H44O8 | 556.695 | 557.696 | 579.678 |
| Manauealide A | C32H47O10Cl | 627.168 | 628.171 | 650.153 |
| Manauealide B | C32H47O10Br | 671.621 | 672.622 | 694.604 |
| Manauealide C | C34H50O11 | 634.762 | 635.763 | 657.745 |
| Anhydrooscillatoxin A | C31H44O9 | 560.683 | 561.685 | 583.667 |
| Anhydroaplysiatoxin | C32H45O9Br | 653.606 | 654.607 | 676.589 |
| Neo-debromoaplysiatoxin A | C32H46O10 | 590.703 | 591.712 | 613.693 |
| Neo-debromoaplysiatoxin B | C27H38O6 | 458.588 | 459.596 | 481.578 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, C.Y.G.; Pang, L.M.; Liang, Z.-X.; Goh, K.K.K.; Glukhov, E.; Gerwick, W.H.; Tan, L.T. MS/MS-Based Molecular Networking Approach for the Detection of Aplysiatoxin-Related Compounds in Environmental Marine Cyanobacteria. Mar. Drugs 2018, 16, 505. https://0-doi-org.brum.beds.ac.uk/10.3390/md16120505
Ding CYG, Pang LM, Liang Z-X, Goh KKK, Glukhov E, Gerwick WH, Tan LT. MS/MS-Based Molecular Networking Approach for the Detection of Aplysiatoxin-Related Compounds in Environmental Marine Cyanobacteria. Marine Drugs. 2018; 16(12):505. https://0-doi-org.brum.beds.ac.uk/10.3390/md16120505
Chicago/Turabian StyleDing, Chi Ying Gary, Li Mei Pang, Zhao-Xun Liang, Kau Kiat Kelvin Goh, Evgenia Glukhov, William H. Gerwick, and Lik Tong Tan. 2018. "MS/MS-Based Molecular Networking Approach for the Detection of Aplysiatoxin-Related Compounds in Environmental Marine Cyanobacteria" Marine Drugs 16, no. 12: 505. https://0-doi-org.brum.beds.ac.uk/10.3390/md16120505