Protective Effects of Fucoxanthin on Ultraviolet B-Induced Corneal Denervation and Inflammatory Pain in a Rat Model

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
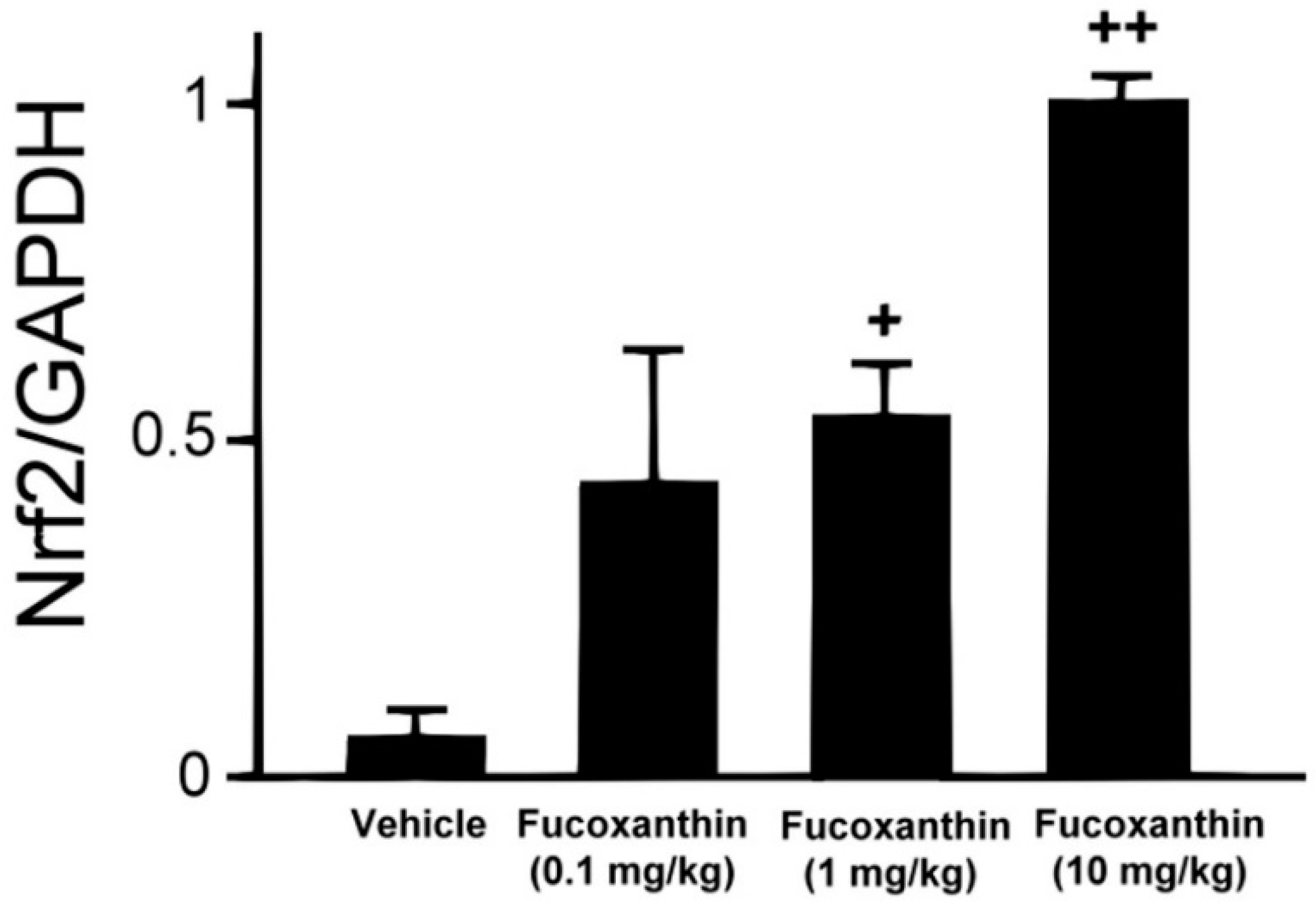
2.1. Effect of Fucoxanthin on Nrf2 Expression
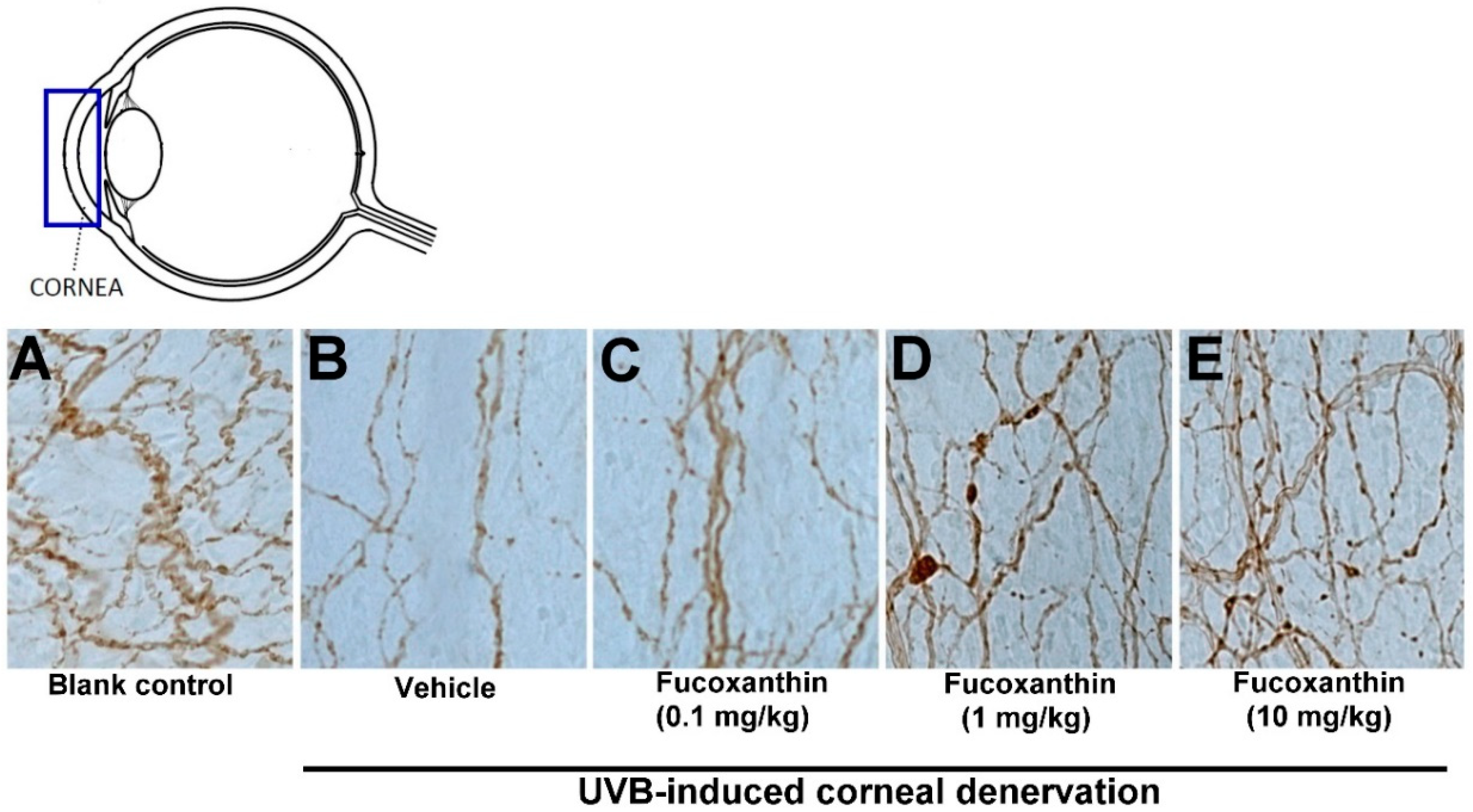
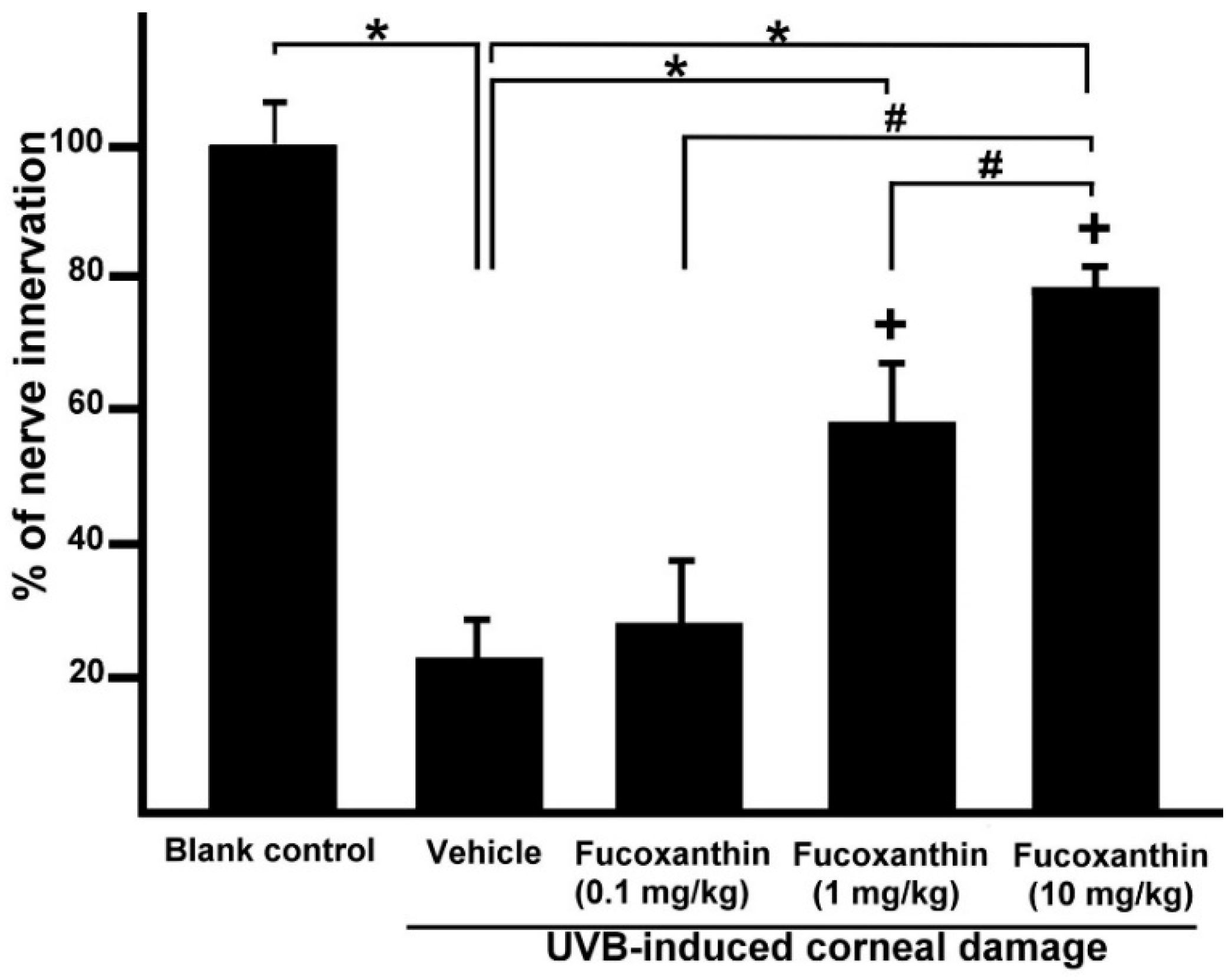
2.2. Effect of Fucoxanthin on UVB-Induced Corneal Denervation
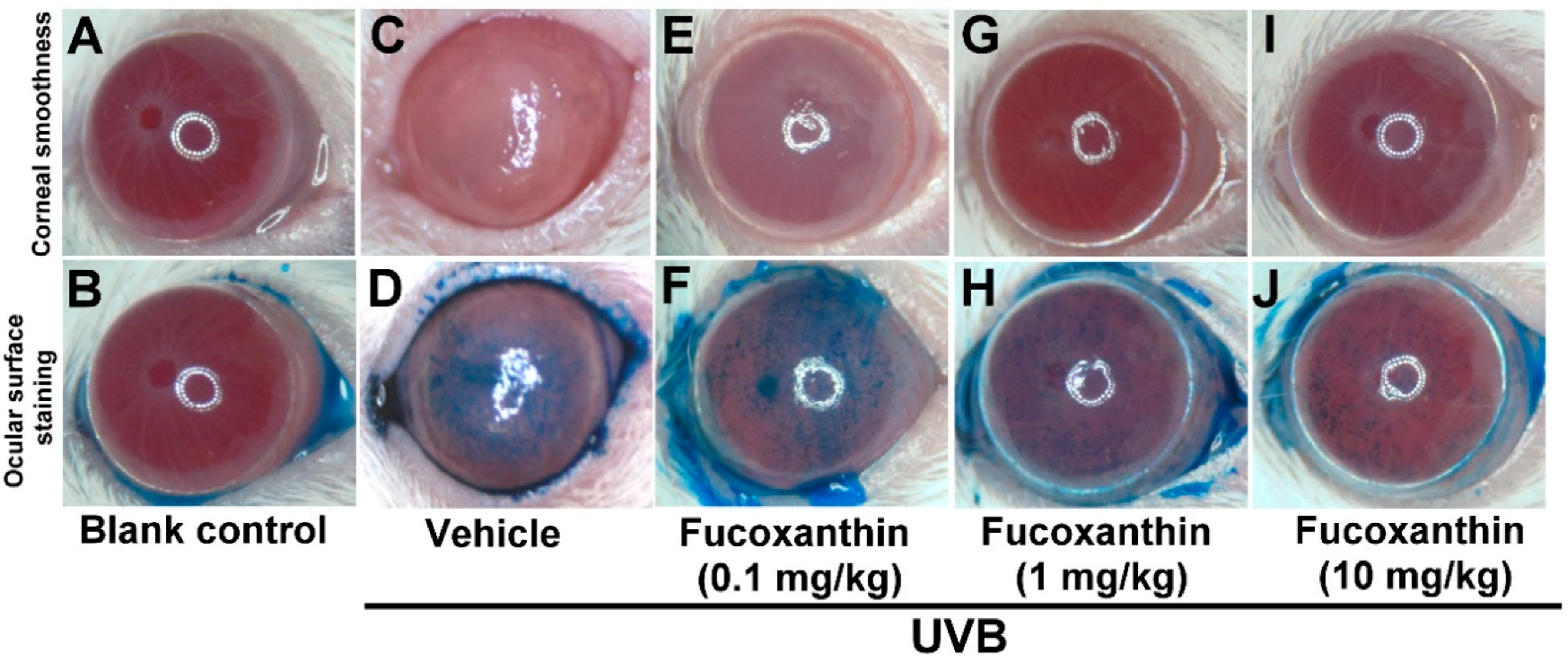
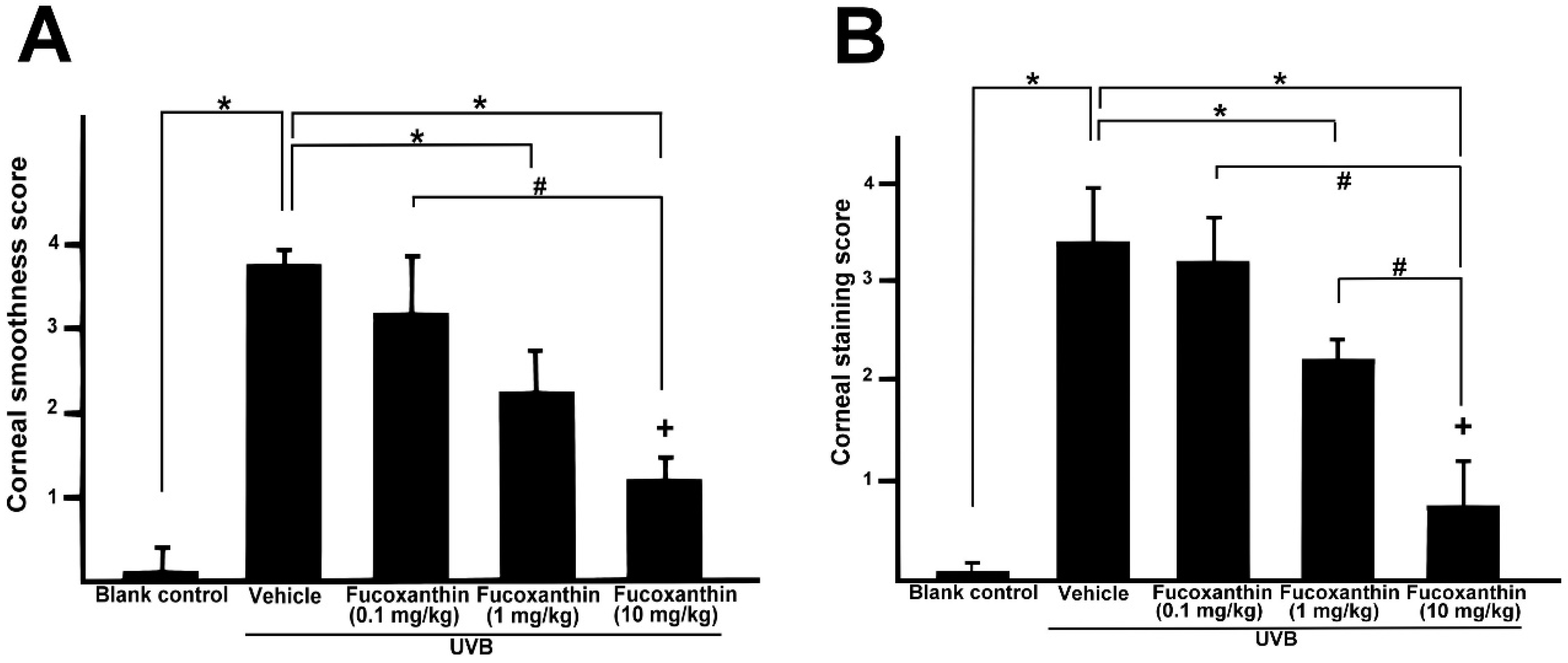
2.3. Effect of Fucoxanthin on UVB-Induced Corneal Epithelial Disorganization
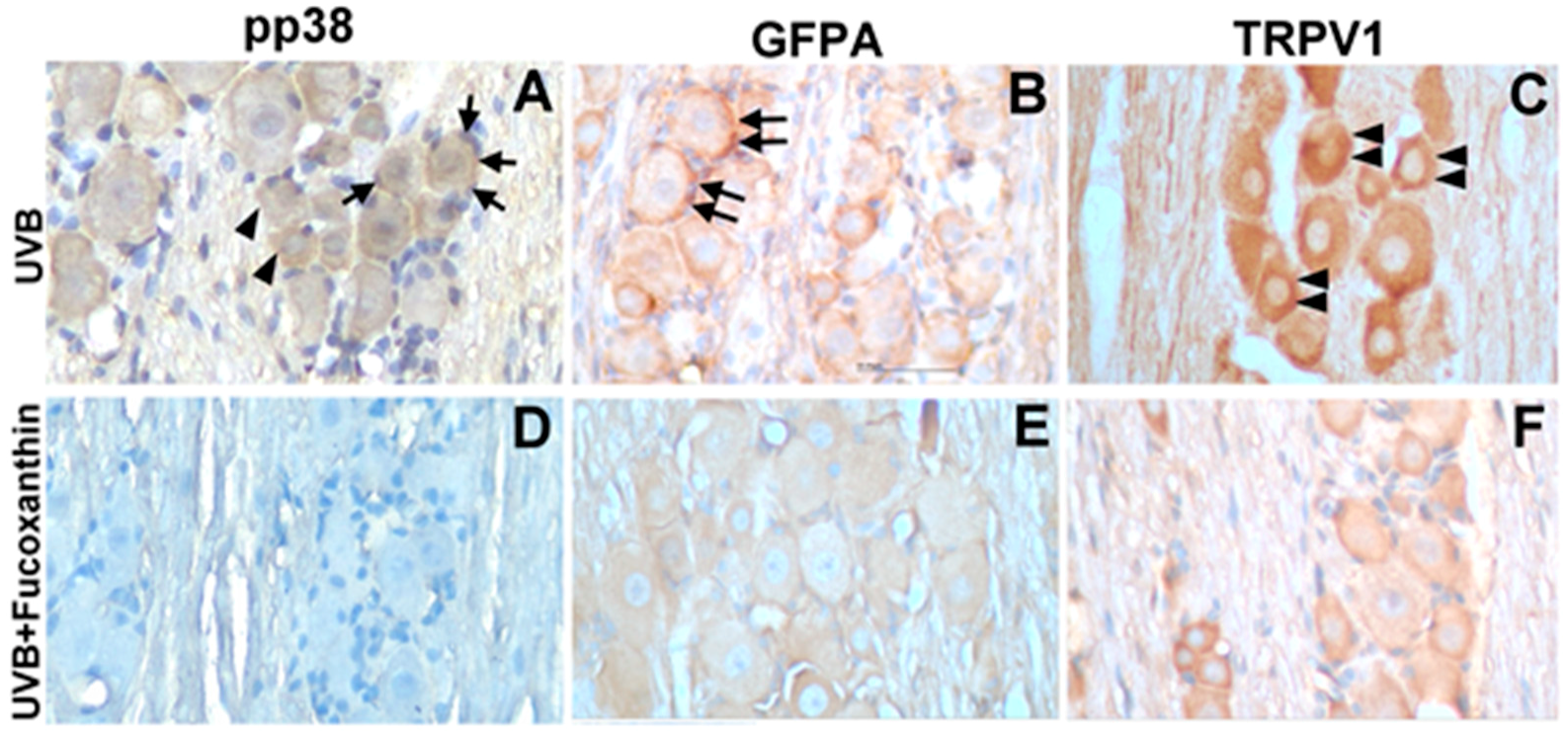
2.4. Effect of Fucoxanthin on UVB-Induced pp38, GFAP and TRPV1 Expression in the Trigeminal Ganglia
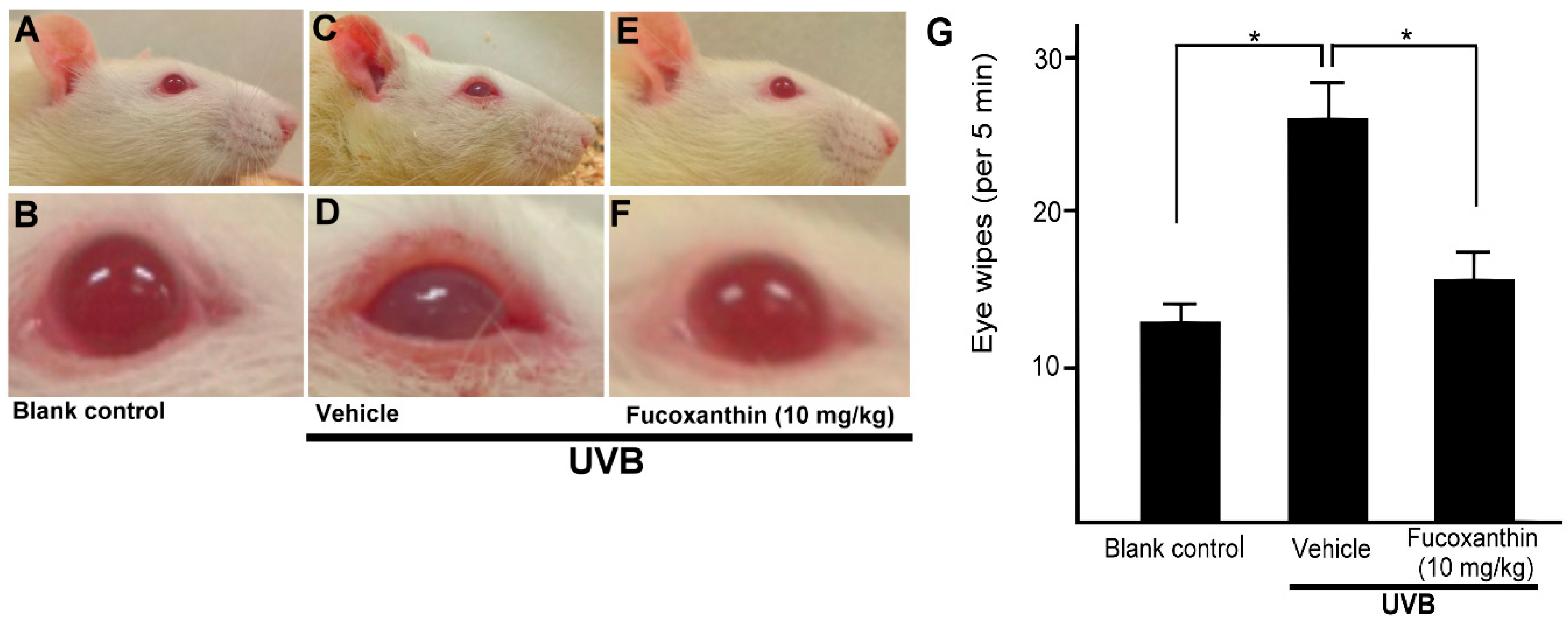
2.5. Effect of Fucoxanthin on UVB-Induced Corneal Injury-Evoked Eye Wipe Behavior
3. Discussions
4. Materials and Methods
4.1. Experimental Animals
4.2. UVB-Induced Photokeratitis and Fucoxanthin Treatment
4.3. Tissue Fractionation and Western Blot
4.4. Immunohistochemistry for Nerve Innervations in Corneal Tissues
4.5. Determination of Corneal Injuries
4.6. Immunohistochemistry of pp38, TRPV1 and GFAP in Trigeminal Ganglia
4.7. Eye Wipe Behavior Test
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Willmann, G. Ultraviolet Keratitis: From the Pathophysiological Basis to Prevention and Clinical Management. High Alt. Med. Biol. 2015, 16, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Podskochy, A.; Fagerholm, P. Cellular response and reactive hyaluronan production in UV-exposed rabbit corneas. Cornea 1998, 17, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Schein, O.D. Phototoxicity and the cornea. J. Natl. Med. Assoc. 1992, 84, 579–583. [Google Scholar] [PubMed]
- Mureşan, S.; Filip, A.; Mureşan, A.; Şimon, V.; Moldovan, R.; Gal, A.F.; Miclăuş, V. Histological findings in the Wistar rat cornea following UVB irradiation. Rom. J. Morphol. Embryol. 2013, 54, 247–252. [Google Scholar] [PubMed]
- Pauloin, T.; Dutot, M.; Joly, F.; Warnet, J.M.; Rat, P. High molecular weight hyaluronan decreases UVB-induced apoptosis and inflammation in human epithelial corneal cells. Mol. Vis. 2009, 15, 577–583. [Google Scholar] [PubMed]
- Visconti, R.; Grieco, D. New insights on oxidative stress in cancer. Curr. Opin. Drug Discov. Dev. 2009, 12, 240–245. [Google Scholar]
- Yamanaka, O.; Liu, C.Y.; Kao, W.W. Fibrosis in the anterior segments of the eye. Endocr. Metab. Immune Disord. Drug Targets 2010, 10, 331–335. [Google Scholar] [CrossRef] [PubMed]
- De Castro, F.; Silos-Santiago, I.; De Armentia, M.L.; Barbacid, M.; Belmonte, C. Corneal innervation and sensitivity to noxious stimuli in trkA knockout mice. Eur. J. Neurosci. 1998, 10, 146–152. [Google Scholar] [CrossRef] [PubMed]
- De Leeuw, A.M.; Chan, K.Y. Corneal nerve regeneration. Correlation between morphology and restoration of sensitivity. Investig. Ophthalmol. Vis. Sci. 1989, 30, 1980–1990. [Google Scholar]
- Garcia-Hirschfeld, J.; Lopez-Briones, L.G.; Belmonte, C. Neurotrophic influences on corneal epithelial cells. Exp. Eye Res. 1994, 59, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Choi, J.S.; Joo, C.K. Effects of nicergoline on corneal epithelial wound healing in rat eyes. Investig. Ophthalmol. Vis. Sci. 2009, 50, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, C.; Aracil, A.; Acosta, M.C.; Luna, C.; Gallar, J. Nerves and sensations from the eye surface. Ocul. Surf. 2004, 2, 248–253. [Google Scholar] [CrossRef]
- Davis, J.B.; Gray, J.; Gunthorpe, M.J.; Hatcher, J.P.; Davey, P.T.; Overend, P.; Harries, M.H.; Latcham, J.; Clapham, C.; Atkinson, K.; et al. Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature 2000, 405, 183–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.X.; Zhuang, Z.Y.; Woolf, C.J.; Ji, R.R. p38 mitogen-activated protein kinase is activated after a spinal nerve ligation in spinal cord microglia and dorsal root ganglion neurons and contributes to the generation of neuropathic pain. J. Neurosci. 2003, 23, 4017–4022. [Google Scholar] [CrossRef] [PubMed]
- Csáti, A.; Edvinsson, L.; Vécsei, L.; Toldi, J.; Fülöp, F.; Tajti, J.; Warfvinge, K. Kynurenic acid modulates experimentally induced inflammation in the trigeminal ganglion. J. Headache Pain 2015, 16, 99. [Google Scholar] [CrossRef] [PubMed]
- Komiya, H.; Shimizu, K.; Ishii, K.; Kudo, H.; Okamura, T.; Kanno, K.; Shinoda, M.; Ogiso, B.; Iwata, K. Connexin 43 expression in satellite glial cells contributes to ectopic tooth-pulp pain. J. Oral Sci. 2018, 60, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Hanani, M. Satellite glial cells in sensory ganglia: From form to function. Brain Res. Rev. 2005, 48, 457–476. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhao, L.; Gu, W.; Liu, Q.; Gao, Z.; Zhu, X.; Wu, Z.; He, H.; Huang, F.; Fan, W. Activation of satellite glial cells in trigeminal ganglion following dental injury and inflammation. J. Mol. Histol. 2018, 49, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Sherratt, P.J.; Pickett, C.B. Regulatory mechanisms controlling gene expression mediated by the antioxidant response element. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 233–260. [Google Scholar] [CrossRef] [PubMed]
- Osburn, W.O.; Yates, M.S.; Dolan, P.D.; Chen, S.; Liby, K.T.; Sporn, M.B.; Taguchi, K.; Yamamoto, M.; Kensler, T.W. Genetic or pharmacologic amplification of nrf2 signaling inhibits acute inflammatory liver injury in mice. Toxicol. Sci. 2008, 104, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Kikuchi, M.; Kubodera, A.; Kawakami, Y. Proton-donative antioxidant activity of fucoxanthin with 1,1-diphenyl-2-picrylhydrazyl (DPPH). Biochem. Mol. Biol. Int. 1997, 42, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Asai, A.; Sugawara, T.; Ono, H.; Nagao, A. Biotransformation of fucoxanthinol into amarouciaxanthin A in mice and HepG2 cells: Formation and cytotoxicity of fucoxanthin metabolites. Drug Metab. Dispos. 2004, 32, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Gammone, M.A.; Riccioni, G.; D’Orazio, N. Marine Carotenoids against Oxidative Stress: Effects on Human Health. Mar. Drugs 2015, 13, 6226–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rengarajan, T.; Rajendran, P.; Nandakumar, N.; Balasubramanian, M.P.; Nishigaki, I. Cancer preventive efficacy of marine carotenoid fucoxanthin: Cell cycle arrest and apoptosis. Nutrients 2013, 5, 4978–4989. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.P.; Hou, Y.H. First evidence for the anti-inflammatory activity of fucoxanthin in high-fat-diet-induced obesity in mice and the antioxidant functions in PC12 cells. Inflammation 2014, 37, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Woo, M.N.; Jeon, S.M.; Shin, Y.C.; Lee, M.K.; Kang, M.A.; Choi, M.S. Anti-obese property of fucoxanthin is partly mediated by altering lipid-regulating enzymes and uncoupling proteins of visceral adipose tissue in mice. Mol. Nutr. Food Res. 2009, 53, 1603–1611. [Google Scholar] [CrossRef] [PubMed]
- Beppu, F.; Hosokawa, M.; Yim, M.J.; Shinoda, T.; Miyashita, K. Down-regulation of hepatic stearoyl-CoA desaturase-1 expression by fucoxanthin via leptin signaling in diabetic/obese KK-A(y) mice. Lipids 2013, 48, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, T.; Matsubara, K.; Akagi, R.; Mori, M.; Hirata, T. Antiangiogenic activity of brown algae fucoxanthin and its deacetylated product, fucoxanthinol. J. Agric. Food Chem. 2006, 54, 9805–9810. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.J.; Lee, C.J.; Lin, T.B.; Liu, H.J.; Huang, S.Y.; Chen, J.Z.; Tseng, K.W. Inhibition of Ultraviolet B-Induced Expression of the Proinflammatory Cytokines TNF-α and VEGF in the Cornea by Fucoxanthin Treatment in a Rat Model. Mar. Drugs 2016, 14, 13. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Luna, A.; Ávila-Román, J.; González-Rodríguez, M.L.; Cózar, M.J.; Rabasco, A.M.; Motilva, V.; Talero, E. Fucoxanthin-containing cream prevents epidermal hyperplasia and UVB-induced skin erythema in mice. Mar. Drugs 2018, 16, 378. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, I.; Cao, M.; Su, Z.Y.; Wu, R.; Guo, Y.; Fang, M.; Kong, A.N. Fucoxanthin elicits epigenetic modifications, Nrf2 activation and blocking transformation in mouse skin JB6 P+ Cells. AAPS J. 2018, 20, 32. [Google Scholar] [CrossRef] [PubMed]
- Muller, L.J.; Marfurt, C.F.; Kruse, F.; Tervo, T.M. Corneal nerves: Structure, contents and function. Exp. Eye Res. 2003, 76, 521–542. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Z.H.; Lin, Z.H.; Shi, X.Z. Eyes in pituitary disorders. J. Biol. Regul. Homeost. Agents 2018, 32, 97–99. [Google Scholar] [PubMed]
- Stepp, M.A.; Tadvalkar, G.; Hakh, R.; Pal-Ghosh, S. Corneal epithelial cells function as surrogate Schwann cells for their sensory nerves. Glia 2017, 65, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Bishop, T.; Hewson, D.W.; Yip, P.K.; Fahey, M.S.; Dawbarn, D.; Young, A.R.; McMahon, S.B. Characterisation of ultraviolet-B-induced inflammation as a model of hyperalgesia in the rat. Pain 2007, 131, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Bishop, T.; Marchand, F.; Young, A.R.; Lewin, G.R.; McMahon, S.B. Ultraviolet-B-induced mechanical hyperalgesia: A role for peripheral sensitisation. Pain 2010, 50, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Davies, E.K.; Boyle, Y.; Chizh, B.A.; Lumb, B.M.; Murrell, J.C. Ultraviolet B-induced inflammation in the rat: A model of secondary hyperalgesia? Pain 2011, 152, 2844–2851. [Google Scholar] [CrossRef] [PubMed]
- Sacchetti, M.; Abicca, I.; Bruscolini, A.; Cavaliere, C.; Nebbioso, M.; Lambiase, A. Allergic conjunctivitis: Current concepts on pathogenesis and management. J. Biol. Regul. Homeost. Agents 2018, 32, 49–60. [Google Scholar] [PubMed]
- Moalem, G.; Tracey, D.J. Immune and inflammatory mechanisms in neuropathic pain. Brain Res. Rev. 2006, 51, 240–264. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, A.J.; Mickle, A.D.; Golden, J.P.; Mack, M.R.; Halabi, C.M.; de Kloet, A.D.; Samineni, V.K.; Kim, B.S.; Krause, E.G.; Gereau, R.W., 4th; et al. Macrophage angiotensin II type 2 receptor triggers neuropathic pain. Proc. Natl. Acad. Sci. USA 2018, 115, E8057–E8066. [Google Scholar] [CrossRef] [PubMed]
- Karai, L.; Brown, D.C.; Mannes, A.J.; Connelly, S.T.; Brown, J.; Gandal, M.; Wellisch, O.M.; Neubert, J.K.; Olah, Z.; Iadarola, M.J. Deletion of vanilloid receptor 1-expressing primary afferent neurons for pain control. J. Clin. Investig. 2004, 113, 1344–1352. [Google Scholar] [CrossRef] [PubMed]
- Julius, D. TRP channels and pain. Annu. Rev. Cell Dev. Biol. 2013, 29, 355–384. [Google Scholar] [CrossRef] [PubMed]
- Kaji, K.; Shinoda, M.; Honda, K.; Unno, S.; Shimizu, N.; Iwata, K. Connexin 43 contributes to ectopic orofacial pain following inferior alveolar nerve injury. Mol. Pain 2016, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Retamal, M.A.; Riquelme, M.A.; Stehberg, J.; Alcayaga, J. Connexin43 hemichannels in satellite glial cells, can they influence sensory neuron activity? Front. Mol. Neurosci. 2017, 10, 374. [Google Scholar] [CrossRef] [PubMed]
- Murray, H.; Locker, D.; Mock, D.; Tenenbaum, H.C. Pain and the quality of life in patients referred to a craniofacial pain unit. J. Orofac. Pain 1996, 10, 316–323. [Google Scholar] [PubMed]
- Hodges, P.W.; Tucker, K. Moving differently in pain: A new theory to explain the adaptation to pain. Pain 2011, 152, S90–S98. [Google Scholar] [CrossRef] [PubMed]
- Piao, Z.G.; Cho, I.H.; Park, C.K.; Hong, J.P.; Choi, S.Y.; Lee, S.J.; Lee, S.; Park, K.; Kim, J.S.; Oh, S.B. Activation of glia and microglial p38 MAPK in medullary dorsal horn contributes to tactile hypersensitivity following trigeminal sensory nerve injury. Pain 2006, 121, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.F.; Zhang, S.; Chiang, C.Y.; Hu, J.W.; Dostrovsky, J.O.; Sessle, B.J. Involvement of glia in central sensitization in trigeminal subnucleus caudalis (medullary dorsal horn). Brain Behav. Immun. 2007, 21, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Lu, J.; Tay, S.S.; Jiang, H.; He, B.P. Induced NG2 expressing microglia in the facial motor nucleus after facial nerve axotomy. Neuroscience 2010, 166, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.; Kwon, M.J.; Nam, T.J. Nrf2 and NF-κB signaling pathways contribute to porphyra-334-mediated inhibition of UVA-induced inflammation in skin fibroblasts. Mar. Drugs 2015, 13, 4721–4732. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.H.; Wang, T.C.; Lee, Y.C.; Shen, P.T.; Chang, J.Y.; Yeh, T.K.; Huang, C.H.; Chang, H.H.; Cheng, S.Y.; Lin, C.Y.; et al. Novel Nrf2/ARE activator, trans-coniferylaldehyde, induces a HO-1-mediated defense mechanism through a dual p38α/MAPKAPK-2 and PK-N3 signaling pathway. Chem. Res. Toxicol. 2015, 28, 1681–1692. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.Q.; Zhang, M.; Matis, K.I.; Kim, C.; Rosenblatt, M.I. Vascular endothelial growth factor mediates corneal nerve repair. Investig. Ophthalmol. Vis. Sci. 2008, 49, 3870–3878. [Google Scholar] [CrossRef] [PubMed]
- Farazifard, R.; Safarpour, F.; Sheibani, V.; Javan, M. Eye-wiping test: A sensitive animal model for acute trigeminal pain studies. Brain Res. Protoc. 2005, 16, 44–49. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.-J.; Lee, C.-J.; Lin, T.-B.; Peng, H.-Y.; Liu, H.-J.; Chen, Y.-S.; Tseng, K.-W. Protective Effects of Fucoxanthin on Ultraviolet B-Induced Corneal Denervation and Inflammatory Pain in a Rat Model. Mar. Drugs 2019, 17, 152. https://0-doi-org.brum.beds.ac.uk/10.3390/md17030152
Chen S-J, Lee C-J, Lin T-B, Peng H-Y, Liu H-J, Chen Y-S, Tseng K-W. Protective Effects of Fucoxanthin on Ultraviolet B-Induced Corneal Denervation and Inflammatory Pain in a Rat Model. Marine Drugs. 2019; 17(3):152. https://0-doi-org.brum.beds.ac.uk/10.3390/md17030152
Chicago/Turabian StyleChen, Shiu-Jau, Ching-Ju Lee, Tzer-Bin Lin, Hsien-Yu Peng, Hsiang-Jui Liu, Yu-Shan Chen, and Kuang-Wen Tseng. 2019. "Protective Effects of Fucoxanthin on Ultraviolet B-Induced Corneal Denervation and Inflammatory Pain in a Rat Model" Marine Drugs 17, no. 3: 152. https://0-doi-org.brum.beds.ac.uk/10.3390/md17030152