The Distribution of Asterosaponins, Polyhydroxysteroids and Related Glycosides in Different Body Components of the Far Eastern Starfish Lethasterias fusca

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
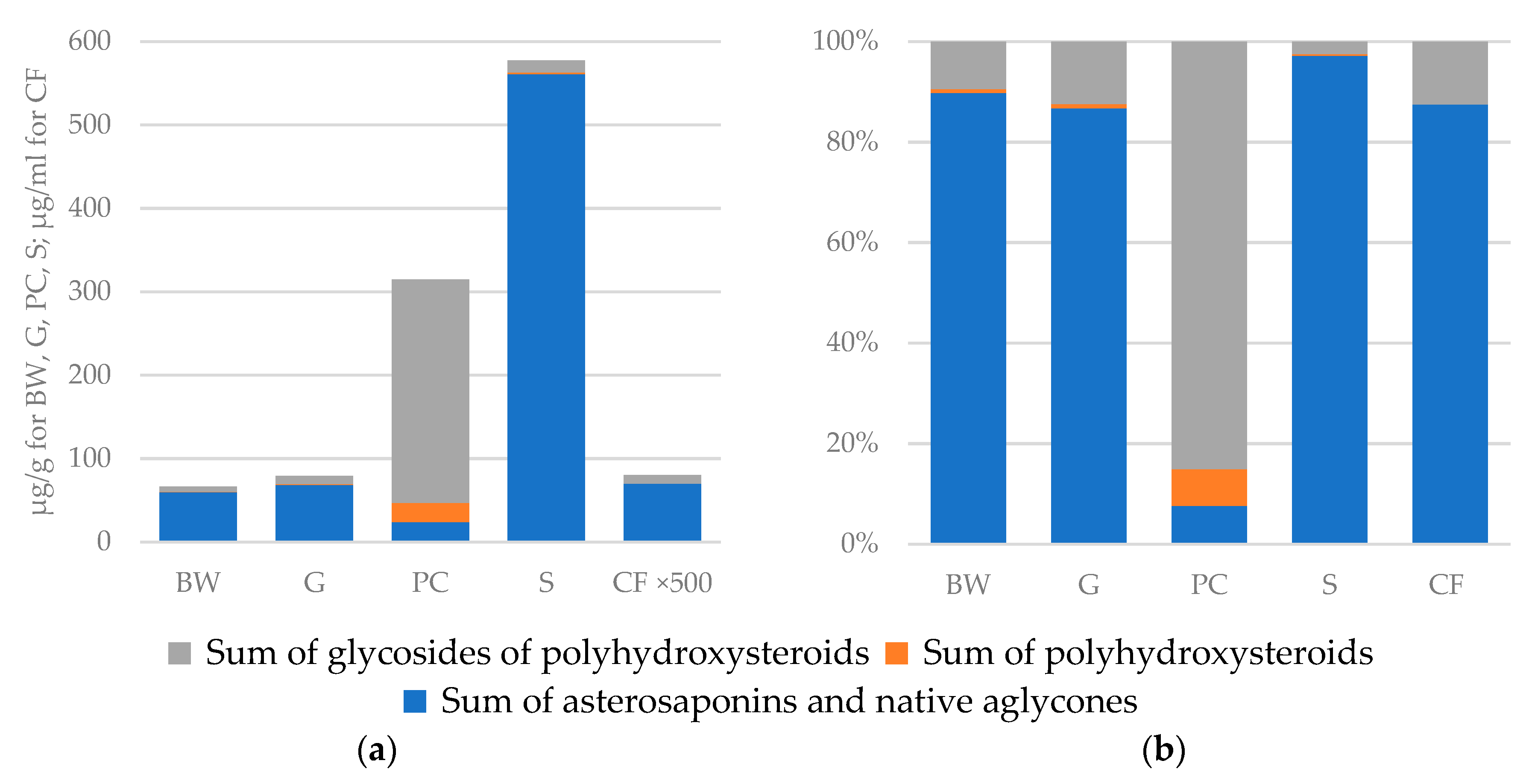
2.1. Distribution of Polar Steroids in Body Components of the Starfish L. fusca
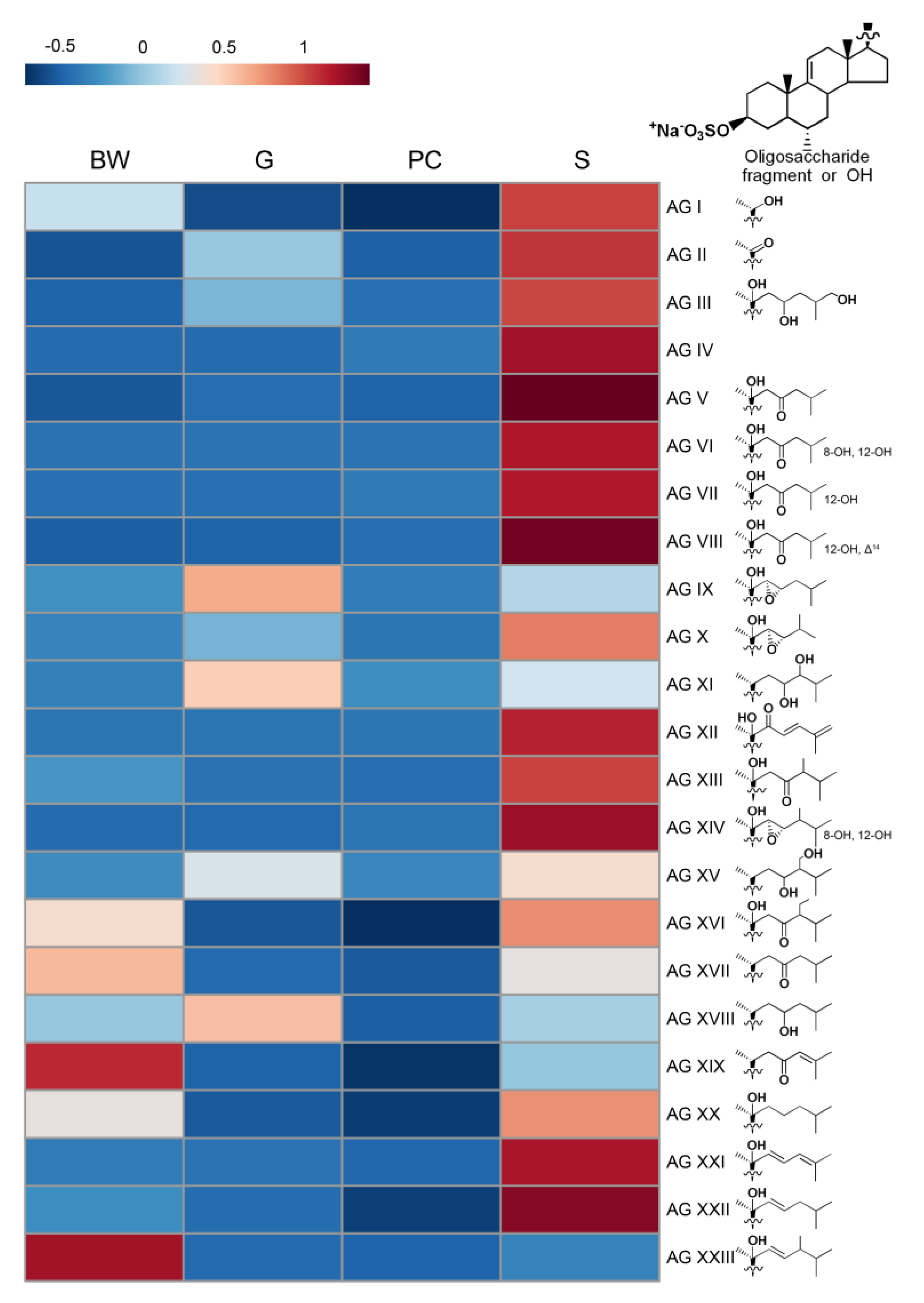
2.2. Distribution of Asterosaponins
2.3. Distribution of Polyhydroxysteroids and Glycosides of Polyhydroxysteroids
3. Materials and Methods
3.1. Chemicals
3.2. Animal Material
3.3. Sample Preparation and Solid-Phase Extraction (SPE)
3.4. LC/MS Analysis
3.5. Semi-Quantitative Analysis of Detected Polar Steroid Compounds
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Minale, L.; Riccio, R.; Zollo, F. Steroidal Oligoglycosides and Polyhydroxysteroids from Echinoderms. Fortschr. Chem. Org. Naturst. 1993, 62, 75–308. [Google Scholar]
- Stonik, V.A. Marine polar steroids. Russ. Chem. Rev. 2001, 70, 673–715. [Google Scholar] [CrossRef]
- Stonik, V.A.; Ivanchina, N.V.; Kicha, A.A. New polar steroids from starfish. Nat. Prod. Commun. 2008, 3, 1587–1610. [Google Scholar] [CrossRef]
- Ivanchina, N.V.; Kicha, A.A.; Stonik, V.A. Steroid glycosides from marine organisms. Steroids 2011, 76, 425–454. [Google Scholar] [CrossRef]
- Dong, G.; Xu, T.H.; Yang, B.; Lin, X.P.; Zhou, X.F.; Yang, X.W.; Liu, Y.H. Chemical Constituents and Bioactivities of Starfish. Chem. Biodivers. 2011, 8, 740–791. [Google Scholar] [CrossRef]
- Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Stonik, V.A. Recent studies of polar steroids from starfish: Structures, biological activities and biosynthesis. In Advances in Natural Products Discovery; Gomes, R., Rocha-Santos, T., Duarte, A., Eds.; Nova Sci.: New York, NY, USA, 2017; pp. 191–224. [Google Scholar]
- Yasumoto, T.; Tanaka, M.; Hashimoto, Y. Distribution of Saponin in Echinoderms. Bull. Jpn. Soc. Sci. Fish 1966, 32, 673–676. [Google Scholar] [CrossRef]
- Mackie, A.M.; Singh, H.T.; Owen, J.M. Studies on the distribution, biosynthesis and function of steroidal saponins in echinoderms. Comp. Biochem. Physiol. Part B Comp. Biochem. 1977, 56, 9–14. [Google Scholar] [CrossRef]
- Voogt, P.A.; van Rheenen, J.W.A. Carbohydrate content and composition of asterosaponins from different organs of the sea star Asterias rubens: Relation to their haemolytic activity and implications for their biosynthesis. Comp. Biochem. Physiol. Part B Comp. Biochem. 1982, 72, 683–688. [Google Scholar] [CrossRef]
- Garneau, F.X.; Harvey, C.; Simard, J.L.; Apsimon, J.W.; Burnell, D.J.; Himmelman, J.H. The distribution of asterosaponins in various body components of the starfish Leptasterias polaris. Comp. Biochem. Physiol. Part B Comp. Biochem. 1989, 92, 411–416. [Google Scholar] [CrossRef]
- Demeyer, M.; De Winter, J.; Caulier, G.; Eeckhaut, I.; Flammang, P.; Gerbaux, P. Molecular diversity and body distribution of saponins in the sea star Asterias rubens by mass spectrometry. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2014, 168, 1–11. [Google Scholar] [CrossRef]
- Demeyer, M.; Wisztorski, M.; Decroo, C.; De Winter, J.; Caulier, G.; Hennebert, E.; Eeckhaut, I.; Fournier, I.; Flammang, P.; Gerbaux, P. Inter- and intra-organ spatial distributions of sea star saponins by MALDI imaging. Anal. Bioanal. Chem. 2015, 407, 8813–8824. [Google Scholar] [CrossRef]
- Kicha, A.A.; Ivanchina, N.V.; Gorshkova, I.A.; Ponomarenko, L.P.; Likhatskaya, G.N.; Stonik, V.A. The distribution of free sterols, polyhydroxysteroids and steroid glycosides in various body components of the starfish Patiria (=Asterina) pectinifera. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2001, 128, 43–52. [Google Scholar] [CrossRef]
- Kicha, A.A.; Ivanchina, N.V.; Stonik, V.A. Seasonal variations in the levels of polyhydroxysteroids and related glycosides in the digestive tissues of the starfish Patiria (Asterina) pectinifera. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2003, 136, 897–903. [Google Scholar] [CrossRef]
- Kicha, A.A.; Ivanchina, N.V.; Stonik, V.A. Seasonal variations in polyhydroxysteroids and related glycosides from digestive tissues of the starfish Patiria (=Asterina) pectinifera. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2004, 139, 581–585. [Google Scholar] [CrossRef]
- Ivanchina, N.V.; Malyarenko, T.V.; Kicha, A.A.; Kalinovskii, A.I.; Dmitrenok, P.S. Polar steroidal compounds from the Far-Eastern starfish Lethasterias fusca. Russ. Chem. Bull. 2008, 57, 204–208. [Google Scholar] [CrossRef]
- Ivanchina, N.V.; Kalinovsky, A.I.; Kicha, A.A.; Malyarenko, T.V.; Dmitrenok, P.S.; Ermakova, S.P.; Stonik, V.A. Two new asterosaponins from the Far Eastern starfish Lethasterias fusca. Nat. Prod. Commun. 2012, 7, 853–858. [Google Scholar] [CrossRef]
- Popov, R.S.; Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Dmitrenok, P.S. Structural characterization of polar steroid compounds of the Far Eastern starfish Lethasterias fusca by nanoflow liquid chromatography coupled to quadrupole time-of-flight tandem mass spectrometry. J. Am. Soc. Mass Spectrom. 2019, 30, 743–764. [Google Scholar] [CrossRef]
- Harvey, C.; Garneau, F.-X.; Himmelman, J. Chemodetection of the predatory sea-star Leptasterias polaris by the whelk Buccinum undatum. Mar. Ecol. Prog. Ser. 1987, 40, 79–86. [Google Scholar] [CrossRef]
- Van Dyck, S.; Flammang, P.; Meriaux, C.; Bonnel, D.; Salzet, M.; Fournier, I.; Wisztorski, M. Localization of secondary metabolites in marine invertebrates: Contribution of MALDI MSI for the study of saponins in cuvierian tubules of H. forskali. PLoS ONE 2010, 5, e13923. [Google Scholar] [CrossRef]
- Popov, R.S.; Ivanchina, N.V.; Silchenko, A.S.; Avilov, S.A.; Kalinin, V.I.; Dolmatov, I.Y.; Stonik, V.A.; Dmitrenok, P.S. Metabolite profiling of triterpene glycosides of the Far Eastern sea cucumber Eupentacta fraudatrix and their distribution in various body components using LC-ESI QTOF-MS. Mar. Drugs 2017, 15, 302. [Google Scholar] [CrossRef]
- Van Dyck, S.; Gerbaux, P.; Flammang, P. Qualitative and quantitative saponin contents in five sea cucumbers from the Indian Ocean. Mar. Drugs 2010, 8, 173–189. [Google Scholar] [CrossRef]
- Goad, L.J. Sterol biosynthesis and metabolism in marine invertebrates. Pure Appl. Chem. 1981, 53, 837–852. [Google Scholar] [CrossRef]
- Goad, L.J. The sterols of marine invertebrates: Composition, biosynthesis, and metabolites. In Marine Natural Products. Chemical and Biological Perspectives; Scheuer, P.J., Ed.; Academic Press: New York, NY, USA, 1978; pp. 76–173. [Google Scholar]
- Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Kalinovsky, A.I.; Dmitrenok, P.S.; Stonik, V.A. Biosynthesis of polar steroids from the Far Eastern starfish Patiria (=Asterina) pectinifera. Cholesterol and cholesterol sulfate are converted into polyhydroxylated sterols and monoglycoside asterosaponin P1 in feeding experiments. Steroids 2013, 78, 1183–1191. [Google Scholar] [CrossRef]
- Chambers, M.C.; MacLean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Storey, J.D.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Asterosaponins and Native Aglycones | Polyhydroxysteroids | Glycosides of Polyhydroxysteroids | Total | |
|---|---|---|---|---|
| Body walls (μg/g) | 59.7 | 0.5 | 6.3 | 66.5 |
| Gonads (μg/g) | 68.4 | 0.7 | 9.8 | 78.9 |
| Pyloric caeca (μg/g) | 23.8 | 23.1 | 267.5 | 314.4 |
| Stomach (μg/g) | 561.1 | 1.9 | 14.5 | 577.5 |
| Coelomic fluid (μg/mL) | 0.14 | - | 0.02 | 0.16 |
| N | Content in Different Organs (Mean ± SD, ng/g) | Proposed Structures | |||
|---|---|---|---|---|---|
| BW | G | PC | S | ||
| 3 | 27.5 ± 23.5 | 0.7 ± 0.9 | 0.8 ± 1 | 6.2 ± 1.5 | Hex–dHex–Hex–Xyl(-Qui)–Qui–AG I |
| 16 | 0.1 ± 0.2 | 0 ± 0 | 3.8 ± 3.5 | 87.7 ± 74.5 | dHex–dHex–Qui(-Qui)–Hex–AG IV |
| 17 | 50.1 ± 53.2 | 39.7 ± 24 | 79.3 ± 61.6 | 658.1 ± 232.1 | Hex–dHex–Hex–Xyl(-Qui)–Qui–AG II |
| 32 | 0.8 ± 0.8 | 3.5 ± 2.5 | 7.4 ± 3.4 | 818 ± 552.4 | dHex–dHex–Qui(-Qui)–DXU–AG VI |
| 33 | 45.4 ± 65.8 | 295.6 ± 526.5 | 20.3 ± 17.6 | 2749.1 ± 1759 | dHex–dHex–Qui(-Qui)–DXU–AG II |
| 34 | 93.2 ± 58.6 | 10.4 ± 6.5 | 13.8 ± 6.7 | 48.5 ± 32.5 | AG I |
| 40 | 23.9 ± 25.3 | 12.9 ± 18.7 | 18.3 ± 16.1 | 615.5 ± 394.5 | Hex–dHex–Hex–Xyl(-Qui)–Qui–AG VII |
| 41 | 0.9 ± 0.6 | 5.7 ± 9.6 | 5.8 ± 6.1 | 325.3 ± 204.6 | Hex–dHex–Hex–Xyl(-Qui)–Qui–AG VIII |
| 44 | 64.4 ± 70.9 | 8.9 ± 8.4 | 5.6 ± 4 | 350.2 ± 240.5 | Hex–dHex–Hex–Xyl(-Qui)–Hex–AG V |
| 51 | 86.1 ± 168.1 | 26.4 ± 22.3 | 45.4 ± 15.4 | 1059.8 ± 733.7 | Qui–Xyl–Qui–AG I |
| 60 | 80.7 ± 102.3 | 52 ± 39.6 | 67.3 ± 59.3 | 1232 ± 639.6 | dHex–Hex–Xyl(-Qui)–Hex–AG V |
| 65 | 0.5 ± 0.3 | 1.2 ± 2.1 | 3.9 ± 2.8 | 375.3 ± 370 | dHex–dHex–Qui(-Qui)–Hex–AG XII |
| 70 | 34.8 ± 15.4 | 39.4 ± 18.5 | 24 ± 11.3 | 2805.1 ± 929.5 | dHex–dHex–Glc(-Qui)–Hex–AG V |
| 74 | 4.8 ± 3.3 | 12 ± 6.4 | 11.7 ± 5.2 | 1468.7 ± 949.4 | dHex–dHex–Qui(-Qui)–DXU–AG VIII |
| 77 | 13 ± 4.9 | 30.6 ± 25.6 | 23.3 ± 8.2 | 2888.6 ± 1744.3 | dHex–dHex–Qui(-Qui)–DXU–AG VII |
| 78 | 2468.3 ± 2319.4 | 1881.1 ± 1119 | 953.2 ± 857.2 | 23,431.5 ± 7708.7 | Hex–dHex–Hex–Xyl(-Qui)–Qui–AG V |
| 88 | 77.5 ± 66.8 | 110.5 ± 52.2 | 33.4 ± 25.3 | 4674.9 ± 1961 | Hex–dHex–dHex–Glc(-Qui)–Qui–AG V |
| 90 | 0.2 ± 0.2 | 0 ± 0.1 | 2.6 ± 2.5 | 83.8 ± 67.6 | dHex–dHex–Glc(-Qui)–Qui–AG XIV |
| 93 | 104 ± 125.9 | 121.9 ± 83.6 | 71.4 ± 61 | 4978 ± 4835.3 | dHex–dHex–Qui(-Qui)–Hex–AG V |
| 105 | 8.8 ± 10.7 | 11.6 ± 12 | 10.1 ± 5.5 | 789.8 ± 563.5 | dHex–dHex–Glc(-Qui)–Hex–AG XIII |
| 106 | 29.7 ± 24.6 | 21.4 ± 12.8 | 39.8 ± 31.9 | 566 ± 187.1 | dHex–dHex–Hex–Xyl(-Qui)–Qui–AG V |
| 111 | 304.4 ± 169.6 | 153.5 ± 38 | 111.4 ± 65.9 | 44,57.6 ± 3242.3 | dHex–Hex–Glc(-Qui)–Qui–AG V |
| 112 | 53.5 ± 51.3 | 16.2 ± 15.1 | 7.5 ± 5.7 | 655.2 ± 516.7 | Hex–dHex–Hex–Xyl(-Qui)–Hex–AG XVII |
| 118 | 831.6 ± 848.4 | 2816.5 ± 1624.1 | 2521.9 ± 2733.1 | 10,972.3 ± 4393.7 | Fuc–Gal–Xyl(-Qui)–Qui–AG V (thornasteroside A) |
| 122 | 2553.5 ± 1926.5 | 6991.5 ± 3200.9 | 3934.5 ± 2011.3 | 173,398.3 ± 148,397.4 | Fuc–Qui–Glc(-Qui)–Qui–AG V (luidiaquinoside) |
| 123 | 2846.4 ± 2424.5 | 162.9 ± 111.1 | 42.8 ± 46.9 | 333.7 ± 264.9 | Hex–dHex–Hex–Xyl(-Qui)–Qui–AG XX |
| 129 | 37.4 ± 27.4 | 68.1 ± 40.1 | 25.7 ± 12.6 | 2677.6 ± 1160.6 | Hex–dHex–dHex–Qui(-Qui)–Qui–AG V |
| 135 | 437.3 ± 207.6 | 2343.4 ± 2062 | 273.3 ± 196.3 | 64,570.3 ± 23,737.3 | dHex–dHex–Qui(-Qui)–DXU–AG V |
| 150 | 1249.7 ± 1349 | 1098.9 ± 860.2 | 224 ± 215.9 | 28,498.5 ± 24,555.7 | dHex–dHex–Qui(-Qui)–Hex–AG XXI |
| 152 | 399.9 ± 462.7 | 931 ± 1346 | 114 ± 78.6 | 6581 ± 1828.8 | dHex–dHex–Glc(-Qui)–Hex–AG XXII |
| 159 | 1114.2 ± 860 | 305.4 ± 187.7 | 66.7 ± 57.1 | 364.7 ± 155.4 | Fuc–Gal–Xyl(-Qui)–Qui–AG XVIII (lethasterioside B) |
| 174 | 1.7 ± 2 | 13.6 ± 10.1 | 91.8 ± 61.2 | 818.9 ± 580.7 | AG VIII |
| 175 | 141.9 ± 90.5 | 355 ± 174.4 | 225.1 ± 120.2 | 7623.4 ± 3936.2 | dHex–dHex–Qui(-Qui)–Qui–AG V |
| 178 | 4.7 ± 5.2 | 25.3 ± 21.7 | 219.6 ± 166.5 | 1023.6 ± 609.4 | AG VII |
| 180 | 51 ± 40.9 | 725.5 ± 1061.2 | 95 ± 52 | 4869.7 ± 3668.5 | dHex–dHex–Qui(-Qui)–(C6H6O3)–AG V |
| 182 | 3440.1 ± 3370.9 | 1962.9 ± 1574 | 614.6 ± 567.6 | 36,581.4 ± 30,632.4 | dHex–dHex–Qui(-Qui)–Hex–AG XXII |
| 186 | 7.8 ± 5.5 | 633.3 ± 1139.3 | 50 ± 35.4 | 3777.4 ± 3427.8 | dHex–dHex–Qui(-Qui)–(C7H9NO4)–AG V |
| 194 | 295 ± 239.4 | 9.3 ± 5.8 | 4.7 ± 4.2 | 30.3 ± 38.5 | Hex–dHex–Hex–Xyl(-Qui)–Qui–AG XXIII |
| 195 | 1935.9 ± 1321.3 | 228.1 ± 163.7 | 14.6 ± 5.5 | 746.4 ± 802.9 | dHex–dHex–Qui(-Qui)–DXU–AG XIX |
| 199 | 4.8 ± 6.4 | 61.6 ± 76 | 68.6 ± 88.9 | 7221.9 ± 7112.6 | dHex–dHex–Qui(-Qui)–(C7H9NO4)–AG V |
| 205 | 1096.5 ± 566.1 | 10,113.8 ± 13,871.4 | 7737.1 ± 4085.7 | 4,5330.5 ± 21,263.7 | 3-O-sulfothornasterol A (AG V) |
| 207 | 477.9 ± 255.2 | 154.4 ± 155.8 | 320.3 ± 99 | 664.4 ± 284.8 | 3-O-sulfo-24,25-dihydromarthasterone (AG XVII) |
| Content in Different Organs (Mean ± SD, ng/g) | Proposed Structures | ||||
|---|---|---|---|---|---|
| BW | G | PC | S | ||
| 1 | 1.1 ± 0.7 | 4.3 ± 7.3 | 369.8 ± 244.8 | 9.4 ± 7.6 | 24-O-pentosyl-5α-cholestane-3β,6,8,15α,16β,24-hexaol |
| 2 | 1.5 ± 1.5 | 3.5 ± 5.2 | 421.5 ± 320.8 | 11.2 ± 7.2 | 24-O-hexosyl-5α-cholestane-3β,6,7,8,15α,16β,24-heptaol |
| 4 | 0.1 ± 0.1 | 0.2 ± 0.5 | 123.8 ± 53.7 | 1.1 ± 0.4 | 5α-cholestane-3β,4β,6α,7α,8,15α,16β,26-octaol |
| 11 | 0.4 ± 0.2 | 1 ± 1.7 | 325.9 ± 257.2 | 8.2 ± 6.4 | 5α-cholestane-3β,6α,7α,8,15α,16β,26-heptaol |
| 15 | 2.2 ± 1.4 | 7.8 ± 11.3 | 1016.8 ± 582 | 15.3 ± 9.8 | 5α-cholestane-3β,4β,6α,7α,8,15β,16β,26-octaol |
| 18 | 22.9 ± 18 | 70.6 ± 108.7 | 3276.3 ± 2782.6 | 167.5 ± 142.1 | pycnopodioside A |
| 21 | 17.8 ± 8.5 | 41.1 ± 51.9 | 2153.6 ± 1149.2 | 80.3 ± 53.5 | desulfated minutoside A |
| 23 | 26.3 ± 26.9 | 82.4 ± 148 | 1579.6 ± 1499.7 | 60.4 ± 54.8 | 3-O-pentosyl-5α-cholestane-3β,6,7,8,15,16β,26-heptaol 26-O-sulfate |
| 53 | 10.6 ± 19.3 | 8.3 ± 8.3 | 164.8 ± 109 | 27.4 ± 42.9 | 24-O-sulfohexosyl-5α-cholestane-3β,6,8,15,16β,24-hexaol |
| 58 | 64.8 ± 36.7 | 53.9 ± 62.7 | 2845.4 ± 1755.4 | 125.4 ± 81.2 | - |
| 59 | 49.8 ± 52.5 | 38.8 ± 40 | 2129.6 ± 1784.9 | 101.8 ± 62.5 | 3-O-pentosyl-5α-cholestane-3β,6,8,15,26-pentaol 26-O-sulfate |
| 63 | 12.2 ± 12.5 | 16.9 ± 28.8 | 455.5 ± 388.1 | 22.8 ± 14 | 5α-cholestane-3β,4,6,7,8,15α,16β,26-octaol 6-O-sulfate |
| 71 | 10.7 ± 6.9 | 4.5 ± 1.7 | 509.3 ± 373 | 19.8 ± 15.5 | 26-O-sulfohexosyl-27-nor-5α-ergost-22-ene-3β,6,8,15,16β,26-hexaol |
| 81 | 177.8 ± 126.7 | 337.9 ± 578.6 | 7196.8 ± 3778.9 | 309.9 ± 243.4 | 24-O-pentosyl-5α-cholestane-3β,6,8,15,24-pentaol 3-O-sulfate |
| 83 | 18.2 ± 18.4 | 8.9 ± 2.9 | 837.4 ± 721 | 35.5 ± 28 | 28-O-sulfohexosyl-5α-ergostane-3β,6,8,15,16β,28-hexaol |
| 91 | 11.1 ± 10 | 5.4 ± 2.8 | 654.6 ± 581 | 20.4 ± 11.8 | 26-O-sulfohexosyl-27-nor-5α-ergost-22-ene-3β,6,8,15,16β,26-hexaol |
| 92 | 35.2 ± 23.6 | 24.6 ± 32.1 | 924.1 ± 715.2 | 26.3 ± 9.1 | 3-O-pentosyl-24-O-sulfopentosyl-5α-cholestane-3β,6,8,15,24-pentaol |
| 94 | 16.3 ± 14.1 | 19.1 ± 22.9 | 551.8 ± 333.7 | 23.8 ± 12.6 | 3-O-pentosyl-26-O-sulfohexosyl-5α-ergost-22-ene-3β,6,8,15,26-pentaol |
| 99 | 10.4 ± 6.9 | 16.7 ± 21.8 | 582.9 ± 330 | 26.8 ± 14.5 | 28-O-sulfohexosyl-5α-ergost-20(22)-ene-3β,6,8,15,16β,28-hexaol |
| 108 | 31.1 ± 18.9 | 53.7 ± 81.4 | 1191.2 ± 499.9 | 58.5 ± 26.7 | 3-O-pentosyl-24-O-methylsulfopentosyl-5α-cholestane-3β,6,8,15,24-pentaol |
| 109 | 255.3 ± 194.8 | 213.7 ± 276.4 | 9046.4 ± 7355.2 | 476.4 ± 512.3 | 24-O-pentosyl-5α-cholest-22-ene-3β,6,8,15,24-pentaol 3-O-sulfate |
| 113 | 68.3 ± 82.6 | 26.2 ± 17.1 | 2005.6 ± 1910.1 | 173.9 ± 243 | 26-O-sulfohexosyl-5α-ergost-22-ene-3β,6,8,15,16β,26-hexaol |
| 115 | 33.9 ± 14.4 | 48.1 ± 68.6 | 1687.4 ± 1086.5 | 94.9 ± 64.4 | 24-O-sulfohexosyl-5α-cholest-20(22)-ene-3β,6,8,15,24-pentaol |
| 117 | 21 ± 12.8 | 12.8 ± 7.8 | 994.9 ± 636 | 109 ± 89.2 | 5α-cholestane-3β,6,8,15α,16β,26-hexaol 6-O-sulfate |
| 120 | 66.3 ± 74.4 | 24.5 ± 6.8 | 2870.3 ± 2423.8 | 122.3 ± 97.6 | 28-O-sulfohexosyl-5α-ergostane-3β,6,8,15,16β,28-hexaol |
| 125 | 29.5 ± 13 | 30.4 ± 45.5 | 1280.1 ± 177.9 | 269.9 ± 315.4 | - |
| 126 | 63.6 ± 61.2 | 142 ± 260.4 | 2519.8 ± 1747.5 | 105.1 ± 107.1 | 28-O-sulfohexosyl-5α-ergostane-3β,6,8,15,28-pentaol |
| 131 | 108.1 ± 73.9 | 124.3 ± 152.8 | 4377.2 ± 4373.3 | 194.2 ± 39.6 | 26-O-sulfohexosyl-27-nor-5α-ergost-20(22)-ene-3β,6,8,15,26-pentaol |
| 133 | 21.1 ± 18.2 | 4.2 ± 1.4 | 620.1 ± 555.7 | 62 ± 59.2 | 5α-cholestane-3β,6,8,15β,16β,26-hexaol 6-O-sulfate |
| 136 | 274.8 ± 165.5 | 549.3 ± 866.2 | 12,619.1 ± 6096.6 | 785.4 ± 489.5 | 5α-cholestane-3β,6,8,15,24-pentaol 24-O-sulfate |
| 138 | 82 ± 41.9 | 185.6 ± 252.1 | 3811 ± 1281.6 | 122.3 ± 84.6 | 26-O-sulfohexosyl-27-nor-5α-ergost-20(22)-ene-3β,6,8,15,26-pentaol |
| 146 | 10.8 ± 10.2 | 5.6 ± 4.1 | 584.6 ± 501.6 | 26.7 ± 25.3 | 5α-ergost-22-ene-3β,6,7,8,15α,16β,26-heptaol 6-O-sulfate |
| 147 | 25.1 ± 22.7 | 14.4 ± 8.8 | 1053.4 ± 813.6 | 60 ± 48.5 | 29-O-sulfohexosyl-5α-stigmast-20(22)-ene-3β,6,8,15,16β,29-hexaol |
| 148 | 14.9 ± 8.3 | 9.1 ± 6.5 | 777.4 ± 500.7 | 27.2 ± 19.5 | 28-O-sulfohexosyl-5α-ergost-22-ene-3β,6,8,15,16β,28-hexaol |
| 151 | 76.2 ± 95.2 | 41.2 ± 14 | 3204.2 ± 2986.5 | 206.6 ± 204.7 | 28-O-sulfopentosyl-5α-ergost-20(22)-ene-3β,6,8,15,16β,28-hexaol |
| 153 | 82.6 ± 58.9 | 40.5 ± 18.6 | 4829.9 ± 3939.8 | 342.7 ± 214.2 | 5α-ergost-22-ene-3β,6,8,15,16β,26-hexaol 6-O-sulfate |
| 154 | 248.2 ± 182.6 | 488.2 ± 798.4 | 9881.2 ± 6654.5 | 629.1 ± 380.1 | 28-O-sulfohexosyl-5α-ergost-22-ene-3β,6,8,15,28-pentaol |
| 163 | 70.5 ± 33.4 | 105.7 ± 164.8 | 3006.8 ± 503.1 | 172.5 ± 49.5 | 29-O-sulfohexosyl-5α-stigmastane-3β,6,8,15,16β,29-hexaol |
| 164 | 210.1 ± 127.7 | 256.7 ± 404.1 | 6606.4 ± 2715.5 | 565.2 ± 205 | 26-O-sulfohexosyl-5α-ergost-22-ene-3β,6,8,15,26-pentaol |
| 165 | 130.1 ± 93.8 | 86.5 ± 38.4 | 5780.8 ± 4339.3 | 315.8 ± 242.6 | 28-O-sulfohexosyl-5α-ergost-22-ene-3β,6,8,15,16β,28-hexaol |
| 171 | 216.3 ± 130.9 | 419.9 ± 643.2 | 7547.9 ± 3816.2 | 478.3 ± 231.9 | 28-O-sulfohexosyl-5α-ergost-22-ene-3β,6,8,15,28-pentaol |
| 172 | 18.1 ± 10 | 38.3 ± 54.7 | 1052 ± 570.4 | 123.6 ± 116.4 | 24-O-methylsulfopentosyl-5α-cholestane-3β,6,8,15,24-pentaol |
| 179 | 276.4 ± 232.1 | 536.3 ± 873.4 | 10,891.4 ± 6548.5 | 681.7 ± 327.8 | 26-O-sulfohexosyl-5α-ergost-22-ene-3β,6,8,15,26-pentaol |
| 181 | 53.5 ± 47.7 | 21.9 ± 4.9 | 2603.9 ± 2287.1 | 130.8 ± 110 | 29-O-sulfohexosyl-5α-stigmastane-3β,6,8,15,16β,29-hexaol |
| 183 | 25 ± 27.3 | 10.4 ± 4.5 | 1081.5 ± 1033.7 | 67.5 ± 59.6 | 29-O-sulfohexosyl-5α-stigmast-22-ene-3β,6,8,15,16β,29-hexaol |
| 189 | 24.3 ± 20.3 | 17.7 ± 11.1 | 1287.3 ± 919.2 | 65.8 ± 38.2 | 29-O-sulfohexosyl-5α-stigmast-22-ene-3β,6,8,15,16β,29-hexaol |
| 196 | 335.5 ± 360.2 | 827.8 ± 1416.6 | 14,640.2 ± 14,245.6 | 1044.1 ± 925.3 | 24-O-sulfopentosyl-5α-cholest-22-ene-3β,6,8,15,24-pentaol |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popov, R.S.; Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Grebnev, B.B.; Stonik, V.A.; Dmitrenok, P.S. The Distribution of Asterosaponins, Polyhydroxysteroids and Related Glycosides in Different Body Components of the Far Eastern Starfish Lethasterias fusca. Mar. Drugs 2019, 17, 523. https://0-doi-org.brum.beds.ac.uk/10.3390/md17090523
Popov RS, Ivanchina NV, Kicha AA, Malyarenko TV, Grebnev BB, Stonik VA, Dmitrenok PS. The Distribution of Asterosaponins, Polyhydroxysteroids and Related Glycosides in Different Body Components of the Far Eastern Starfish Lethasterias fusca. Marine Drugs. 2019; 17(9):523. https://0-doi-org.brum.beds.ac.uk/10.3390/md17090523
Chicago/Turabian StylePopov, Roman S., Natalia V. Ivanchina, Alla A. Kicha, Timofey V. Malyarenko, Boris B. Grebnev, Valentin A. Stonik, and Pavel S. Dmitrenok. 2019. "The Distribution of Asterosaponins, Polyhydroxysteroids and Related Glycosides in Different Body Components of the Far Eastern Starfish Lethasterias fusca" Marine Drugs 17, no. 9: 523. https://0-doi-org.brum.beds.ac.uk/10.3390/md17090523