Eighteen New Aeruginosamide Variants Produced by the Baltic Cyanobacterium Limnoraphis CCNP1324

 ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. Extraction and Isolation
3.2. LC-MS/MS Analysis
3.3. NMR Analysis
3.4. Cytotoxicity Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Burja, A.M.; Banaigs, B.; Abou-Mansour, E.; Burgess, J.G.; Wright, P.C. Marine cyanobacteria—A prolific source of natural products. Tetrahedron 2001, 57, 9347–9377. [Google Scholar] [CrossRef]
- Chlipala, G.E.; Mo, S.; Orjala, J. Chemodiversity in freshwater and terrestrial cyanobacteria—A source for drug discovery. Curr. Cancer Drug Targets 2011, 12, 1654–1673. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, E.; Gugger, M.; Sivonen, K.; Fewer, D.P. Natural product biosynthetic diversity and comparative genomics of the cyanobacteria. Trends Microbiol. 2015, 23, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.A.A.; Akhter, N.; Auckloo, B.N.; Khan, I.; Lu, Y.; Wang, K.; Wu, B.; Guo, Y. Structural diversity, biological properties and applications of natural products from cyanobacteria: A review. Mar. Drugs 2017, 15, 354. [Google Scholar] [CrossRef] [Green Version]
- Seddek, N.H.; Fawzy, M.A.; El-Said, W.A.; Ahmed, M.M.R. Evaluation of antimicrobial, antioxidant and cytotoxic activities and characterization of bioactive substances from freshwater blue-green algae. Glob. NEST J. 2019, 21, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Sivonen, K.; Leikoski, N.; Fewer, D.P.; Jokela, J. Cyanobactins—Ribosomal cyclic peptides produced by cyanobacteria. Appl. Microbiol. Biotechnol. 2010, 86, 1213–1225. [Google Scholar] [CrossRef] [Green Version]
- Martins, J.; Vasconcelos, V. Cyanobactins from cyanobacteria: Current genetic and chemical state of knowledge. Mar. Drugs 2015, 13, 6910–6946. [Google Scholar] [CrossRef] [Green Version]
- Donia, M.S.; Ravel, J.; Schmidt, E.W. A global assembly line for cyanobactins. Nat. Chem. Biol. 2008, 4, 341–343. [Google Scholar] [CrossRef]
- Gu, W.; Dong, S.H.; Sarkar, S.; Nair, S.K.; Schmidt, E.W. The biochemistry and structural biology of cyanobactin pathways: Enabling combinatorial biosynthesis. Methods Enzymol. 2018, 604, 113–163. [Google Scholar] [CrossRef]
- Arnison, P.G.; Bibb, M.J.; Bierbaum, G.; Bowers, A.A.; Bugni, T.S.; Bulaj, G.; Camarero, J.A.; Campopiano, D.J.; Challis, G.L.; Clardy, J.; et al. Ribosomally synthesized and post-translationally modified peptide natural products: Overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 2013, 30, 108–160. [Google Scholar] [CrossRef]
- McIntosh, J.A.; Donia, M.S.; Nair, S.K.; Schmidt, E.W. Enzymatic basis of ribosomal peptide prenylation in cyanobacteria. J. Am. Chem. Soc. 2011, 133, 13698–13705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, A.R.; Feng, Y.; Bowden, B.F.; Coll, J.C. Studies of Australian ascidians. 5. Virenamides A–C, new cytotoxic linear peptides from the colonial didemnid ascidian Diplosoma Virens. J. Org. Chem. 1996, 61, 4059–4061. [Google Scholar] [CrossRef] [PubMed]
- Wesson, K.J.; Hamann, M. Keenamide A, a bioactive cyclic peptide from the marine mollusc Pleurobranchus forskalii. J. Nat. Prod. 1996, 59, 631–659. [Google Scholar] [CrossRef]
- Rudi, A.; Aknin, M.; Gaydou, E.M.; Kashman, Y. Four new cytotoxic cyclic hexa- and heptapeptides from the marine ascidian Didemnum molle. Tetrahedron 1998, 54, 13203–13210. [Google Scholar] [CrossRef]
- Ishida, K.; Nakagawa, H.; Murakami, M. Microcyclamide, a cytotoxic cyclic hexapeptide from the cyanobacterium Microcystis aeruginosa. J. Nat. Prod. 2000, 63, 1315–1317. [Google Scholar] [CrossRef] [PubMed]
- Ireland, C.M.; Durso, A.R., Jr.; Newman, R.A.; Hacker, M.P. Antineoplastic cyclic peptides from the marine tunicate Lissoclinum patella. J. Org. Chem. 1982, 47, 1807–1811. [Google Scholar] [CrossRef]
- Degnan, B.M.; Hawkins, C.J.; Lavin, M.F.; McCaffrey, E.J.; Parry, D.L.; van den Brenk, A.L.; Watterst, D.J. New cyclic peptides with cytotoxic activity from the ascidian Lissoclinum patella. J. Med. Chem. 1989, 32, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Linington, R.G.; Gonzàles, J.; Ureña, L.-D.; Romero, L.I.; Ortega-Barria, E.; Gerwick, W.H. Venturamides A and B: Antimalarial constituents of the Panamanian marine cyanobacterium Oscillatoria sp. J. Nat. Prod. 2007, 70, 397–401. [Google Scholar] [CrossRef]
- Todorova, A.K.; Jüttner, F.; Linden, A.; Plüss, T.; von Philipsborn, W. Nostocyclamide: A new macrocyclic, thiazole-containing allelochemical from Nostoc sp. 31 (cyanobacteria). J. Org. Chem. 1995, 60, 7891–7895. [Google Scholar] [CrossRef]
- Jüttner, F.; Todorova, A.K.; Walch, N.; von Philipsborn, W. Nostocyclamide M: A cyanobacterial cyclic peptide with allelopathic activity from Nostoc 31. Phytochemistry 2001, 57, 613–619. [Google Scholar] [CrossRef]
- Ireland, C.; Scheuer, P.J. Ulicyclamide and ulithiacyclamide, two new small peptides from a marine Tunicate. J. Am. Chem. Soc. 1980, 102, 5688–5691. [Google Scholar] [CrossRef]
- Long, P.F.; Dunlap, W.C.; Battershill, C.N.; Jaspars, M. Shotgun cloning and heterologous expression of the patellamide gene cluster as a strategy to achieving sustained metabolite production. Chem. Biol. Chem. 2005, 6, 1760–1765. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, E.W.; Nelson, J.T.; Rasko, D.A.; Sudek, S.; Eisen, J.A.; Haygood, M.G.; Ravel, J. Patellamide A and C biosynthesis by a microcin-like pathway in Prochloron didemni, the cyanobacterial symbiont of Lissoclinum patella. Proc. Natl. Acad. Sci. USA 2005, 102, 7315–7320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leikoski, N.; Fewer, D.P.; Jokela, J.; Wahlsten, M.; Rouhiainen, L.; Sivonen, K. Highly diverse cyanobactins in strains of the genus Anabaena. Appl. Environ. Microbiol. 2010, 76, 701–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donia, M.S.; Schmidt, E.W. Linking chemistry and genetics in the growing cyanobactin natural products family. Chem. Biol. 2011, 18, 508–519. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, J.A.; Lin, Z.; Tianero, M.D.; Schmidt, E.W. Aestuaramides, a natural library of cyanobactin cyclic peptides resulting from isoprene-derived Claisen rearrangements. ACS Chem. Biol. 2013, 8, 877–883. [Google Scholar] [CrossRef] [Green Version]
- Ishida, K.; Matsuda, H.; Murakami, M.; Yamaguchi, K. Kawaguchipeptin B, an antibacterial cyclic undecapeptide from the cyanobacterium Microcystis aeruginosa. J. Nat. Prod. 1997, 60, 724–726. [Google Scholar] [CrossRef]
- Lawton, L.A.; Morris, L.A.; Jaspars, M. A bioactive modified peptide, aeruginosamide, isolated from the cyanobacterium Microcystis aeruginosa. J. Org. Chem. 1999, 64, 5329–5332. [Google Scholar] [CrossRef]
- Gesner-Apter, S.; Carmeli, S. Three novel metabolites from a bloom of the cyanobacterium Microcystis sp. Tetrahedron 2008, 64, 6628–6634. [Google Scholar] [CrossRef]
- Portmann, C.; Blom, J.F.; Kaiser, M.; Brun, R.; Jüttner, F.; Gademann, K. Isolation of aerucyclamides C and D and structure revision of microcyclamide 7806A: Heterocyclic ribosomal peptides from Microcystis aeruginosa PCC 7806 and their antiparasite evaluation. J. Nat. Prod. 2008, 71, 1891–1896. [Google Scholar] [CrossRef]
- Crnkovic, C.M.; Braesel, J.; Krunic, A.; Eustáquio, A.S.; Orjala, J. Scytodecamide from the cultured Scytonema sp. UIC 10036 expands the chemical and genetic diversity of cyanobactins. ChemBioChem 2020, 21, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.; Leikoski, N.; Wahlsten, M.; Azevedo, J.; Antunes, J.; Jokela, J.; Sivonen, K.; Vasconcelos, V.; Fewer, D.P.; Leão, P.N. Sphaerocyclamide, a prenylated cyanobactin from the cyanobacterium Sphaerospermopsis sp. LEGE 00249. Sci. Rep. 2018, 8, 14537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leikoski, N.; Fewer, D.P.; Sivonen, K. Widespread occurrence and lateral transfer of the cyanobactin biosynthesis gene cluster in cyanobacteria. Appl. Environ. Microbiol. 2009, 75, 853–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leikoski, N.; Liu, L.; Jokela, J.; Wahlsten, M.; Gugger, M.; Calteau, A.; Permi, P.; Kerfeld, C.A.; Sivonen, K.; Fewer, D.P. Genome mining expands the chemical diversity of the cyanobactin family to include highly modified linear peptides. Chem. Biol. 2013, 20, 1033–1043. [Google Scholar] [CrossRef] [PubMed]
- Donia, M.S.; Schmidt, E.W. Comprehensive natural products II chemistry and biology. In Cyanobactins—Ubiquitous Cyanobacterial Ribosomal Peptide Metabolites; Elsevier: Oxford, UK, 2010; pp. 539–558. [Google Scholar]
- Nagatsu, A.; Kajitani, H.; Sakakibara, J. Muscoride A: A new oxazole peptide alkaloid from freshwater cyanobacterium Nostoc muscorum. Tetrahedron Lett. 1995, 36, 4097–4100. [Google Scholar] [CrossRef]
- Mattila, A.; Andsten, R.M.; Jumppanen, M.; Assante, M.; Jokela, J.; Wahlsten, M.; Mikula, K.M.; Sigindere, C.; Kwak, D.H.; Gugger, M.; et al. Biosynthesis of the bis-prenylated alkaloids muscoride A and B. ACS Chem. Biol. 2019, 14, 2683–2690. [Google Scholar] [CrossRef]
- Ziemert, N.; Ishida, K.; Liaimer, A.; Hertweck, C.; Dittmann, E. Ribosomal synthesis of tricyclic depsipeptides in bloom-forming cyanobacteria. Angew. Chem. Int. Ed. Engl. 2008, 47, 7756–7759. [Google Scholar] [CrossRef]
- Botta, B.; Vitali, A.; Menendez, P.; Misiti, D.; Monache, G. Prenylated flavonoids: Pharmacology and biotechnology. Curr. Med. Chem. 2005, 12, 717–739. [Google Scholar] [CrossRef]
- Wang, M.; Casey, P.J. Protein prenylation: Unique fats make their mark on biology. Nat. Rev. Mol. Cell Biol. 2016, 17, 110–122. [Google Scholar] [CrossRef]
- Wong, C.P.; Awakawa, T.; Nakashima, Y.; Mori, T.; Zhu, Q.; Liu, X.; Abe, I. Two distinct substrate binding modes for the normal and reverse prenylation of hapalindoles by the prenyltransferase AmbP3. Angew. Chem. Int. Ed. Engl. 2018, 57, 560–563. [Google Scholar] [CrossRef]
- Wätjen, W.; Weber, N.; Lou, Y.J.; Wang, Z.Q.; Chovolou, Y.; Kampkötter, A.; Kahl, R.; Proksch, P. Prenylation enhances cytotoxicity of apigenin and liquiritigenin in rat H4IIE hepatoma and C6 glioma cells. Food Chem. Toxicol. 2007, 45, 119–124. [Google Scholar] [CrossRef] [PubMed]
- López-Ogalla, J.; García-Palomero, E.; Sánchez-Quesada, J.; Rubio, L.; Delgado, E.; García, P.; Medina, M.; Castro, A.; Muñoz, P. Bioactive prenylated phenyl derivatives derived from marine natural products: Novel scaffolds for the design of BACE inhibitors. Med. Chem. Commun. 2014, 5, 474. [Google Scholar] [CrossRef]
- Fu, X.; Do, T.; Schmitz, F.J.; Andrusevich, V.; Engel, M.H. New cyclic peptides from the ascidian Lissoclinum patella. J. Nat. Prod. 1998, 61, 1547–1551. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-J.; Li, Y.-Y.; Liu, X.-Y.; Lu, X.-L.; Cao, X.; Jiao, B.-H. Marine antibody-drug conjugates: Design strategies and research progress. Mar. Drugs 2017, 15, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luesch, H.; Moore, R.E.; Paul, V.J.; Mooberry, S.L.; Corbett, T.H. Isolation of dolastatin 10 from the marine cyanobacterium Symploca species VP642 and total stereochemistry and biological evaluation of its analogue symplostatin 1. J. Nat. Prod. 2001, 64, 907–910. [Google Scholar] [CrossRef]
- Senter, P.D.; Sievers, E.L. The discovery and development of brentuximab vedotin for use in relapsed hodgkin lymphoma and systemic anaplastic large cell lymphoma. Nat. Biotechnol. 2012, 30, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Felczykowska, A.; Pawlik, A.; Mazur-Marzec, H.; Toruńska-Sitarz, A.; Narajczyk, M.; Richert, M.; Węgrzyn, G.; Herman-Antosiewicz, A. Selective inhibition of cancer cells’ proliferation by compounds included in extracts from Baltic Sea cyanobacteria. Toxicon 2015, 108, 1–10. [Google Scholar] [CrossRef]
- Szubert, K.; Wiglusz, M.; Mazur-Marzec, H. Bioactive metabolites produced by Spirulina subsalsa from the Baltic Sea. Oceanologia 2018, 60, 245–255. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aeruginosamide AEG | m/z | Retention Time [min] | Relative Peak Area of Extracted Ion | AEG Chemical Structure | Reference |
|---|---|---|---|---|---|
| AEG-A | 561 | - | - | (Pre)2+Ile+Val+Pyr+TzlCOOMe | [28] |
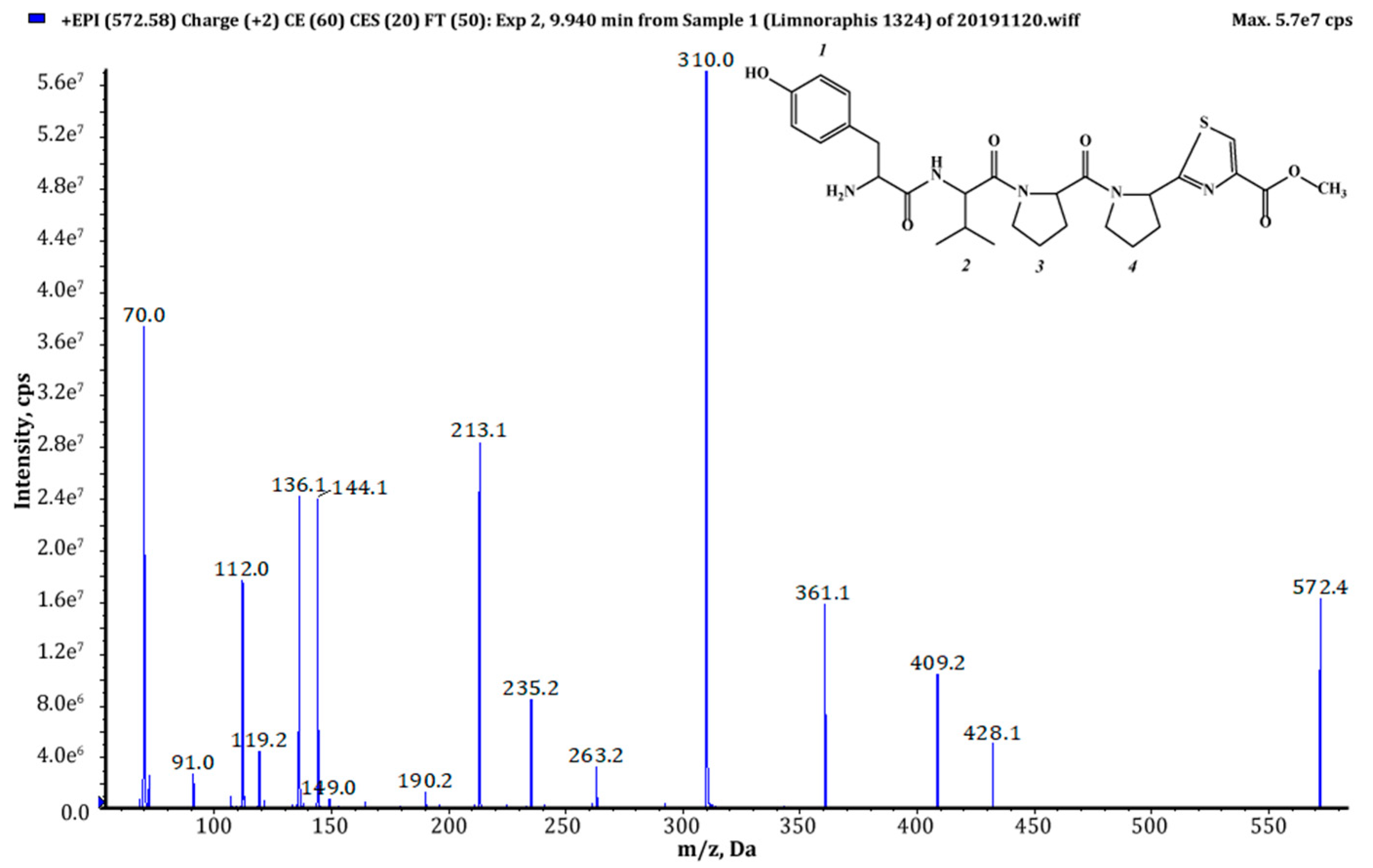
| 571 (1),(2) | 572 | 9.94 | 1.39 × 109 | Tyr+Val+Pro+Pyr+TzlCOOMe | This study |
| AEG-B | 575 | - | - | Pre+Phe+Phe+Pyr+TzlCOOMe | [34] |
| 595 (2) | 596 | 5.12 | T | 187+Val+Pro+Pyr+TzlCOOMe | This study |
| 603 (2) | 604 | 9.74 | T | Phe+Phe+Pro+Pyr+TzlCOOMe | This study |
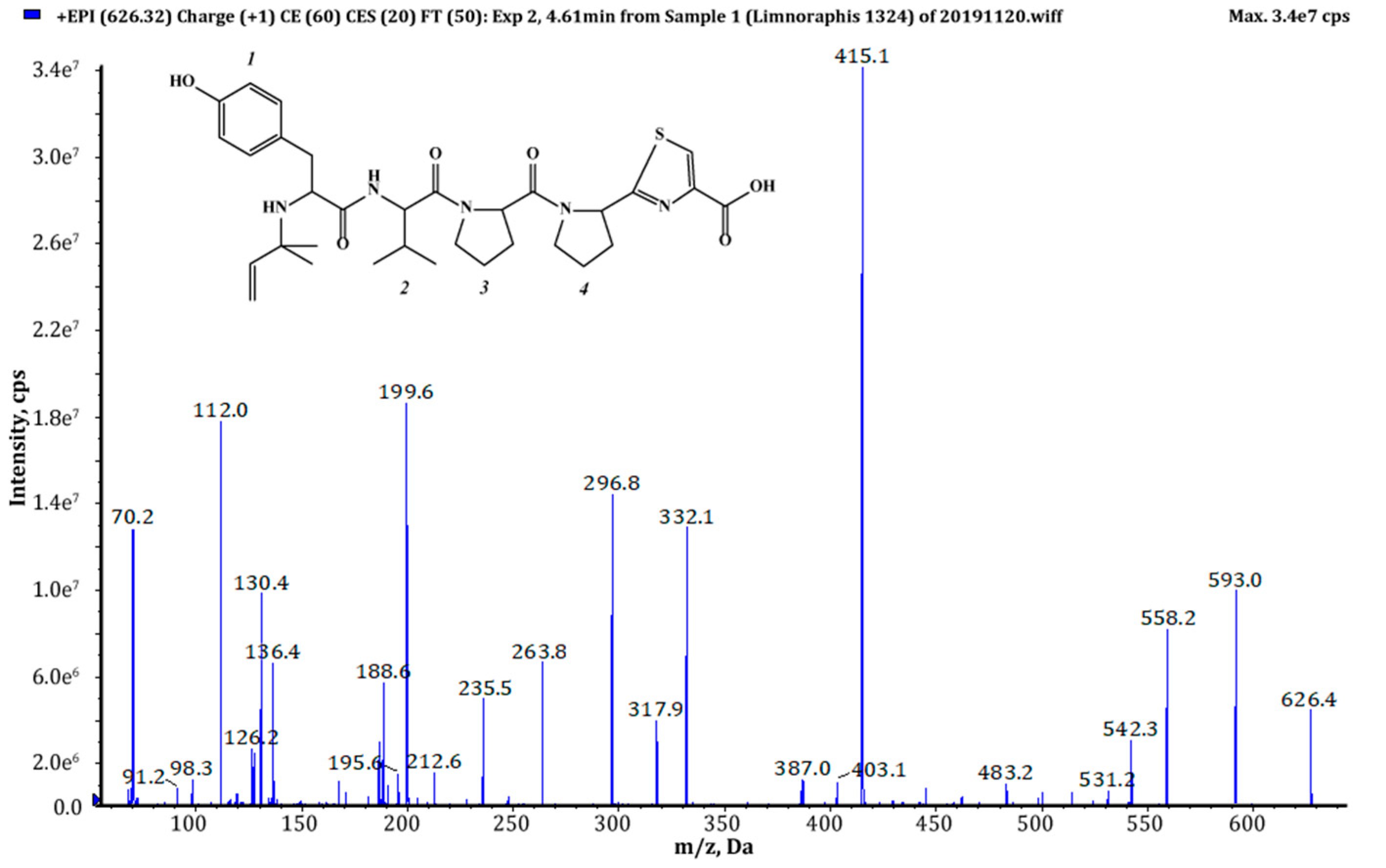
| 625 (1),(2) | 626 | 4.61 | 6.82 × 108 | Pre+Tyr+Val+Pro+Pyr+TzlCOOH | This study |
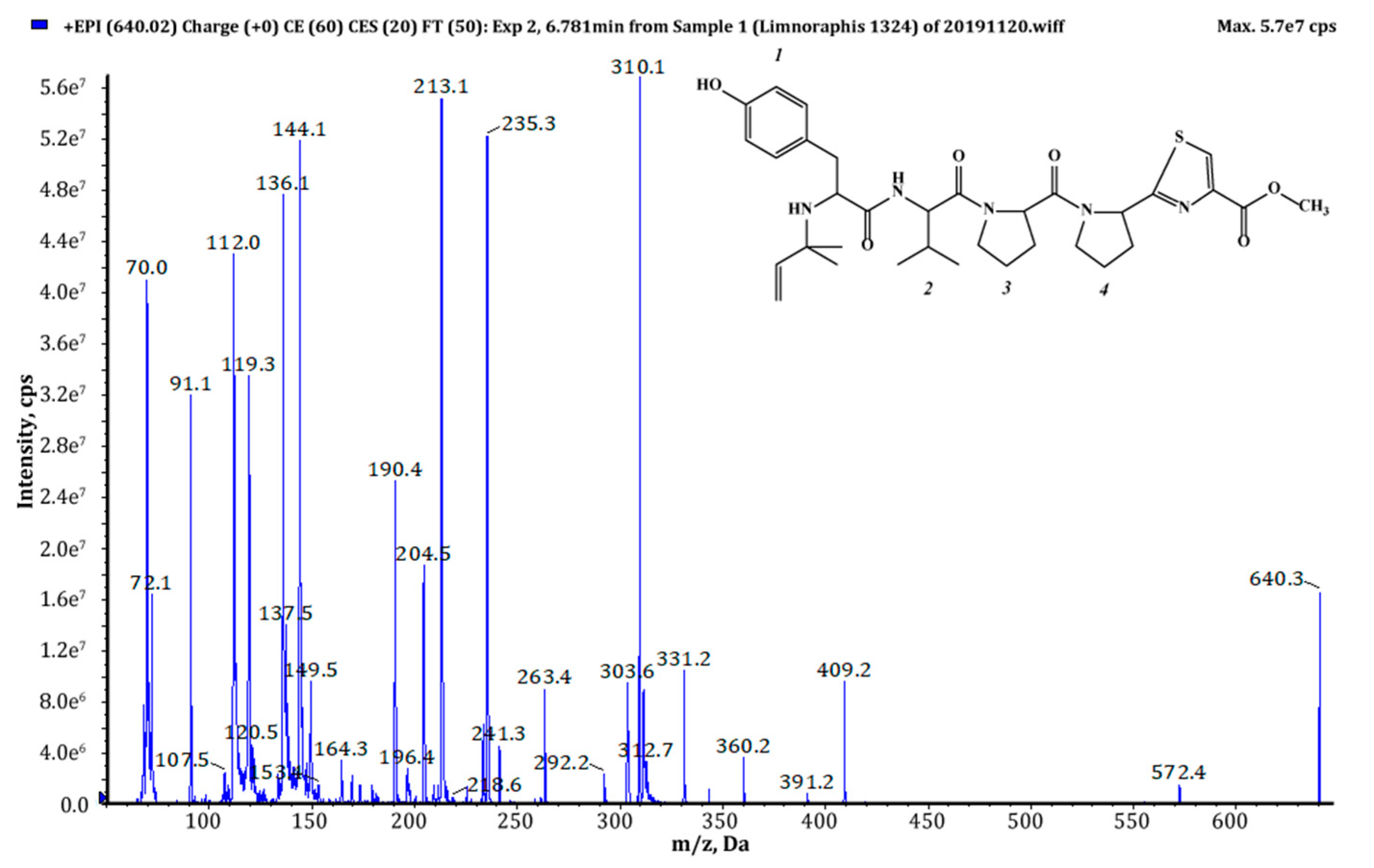
| 639 (1),(2) | 640 | 6.78 | 1.23 × 109 | Pre+Tyr+Val+Pro+Pyr+TzlCOOMe | This study |
| 657 (1),(2) | 658 | 2.27 | 5.8 × 108 | Pre+Phe+Phe+Pro+Pyr+TzlCOOH | This study |
| 667 (2) | 668 | 8.22 | T | Pre+MeHTyr+Val+Pro+Pyr+TzlCOOMe | This study |
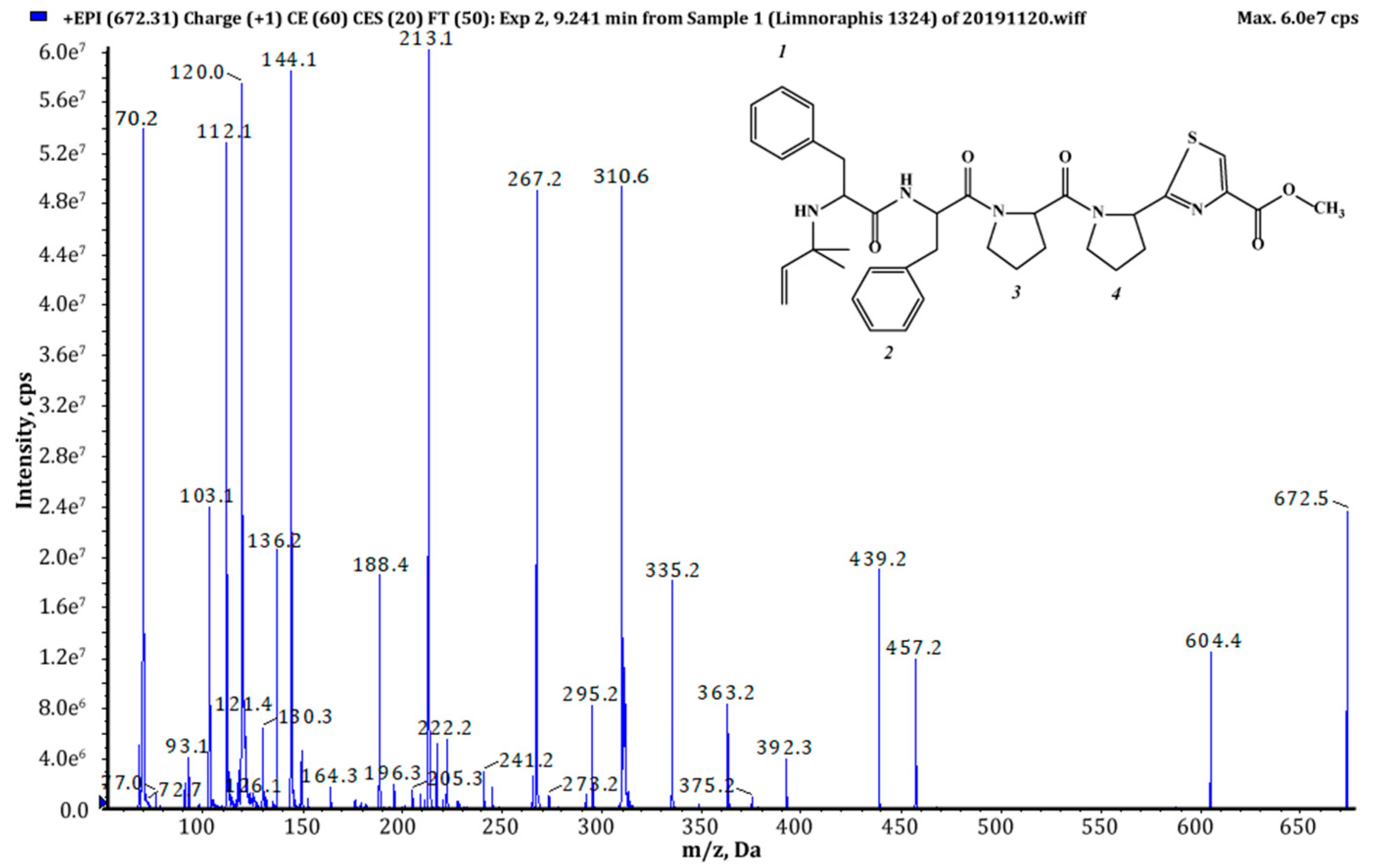
| 671 (1),(2) | 672 | 9.24 | 4.45 × 108 | Pre+Phe+Phe+Pro+Pyr+TzlCOOMe | This study |
| AEG-C | 674 | - | - | Pre+Phe+Phe+Pro+Val+TzlCOOMe | [34] |
| 681a (2) | 682a | 7.16 | T | Pre+205+Val+Pro+Pyr+TzlCOOMe | This study |
| 681b (2) | 682b | 11.78 | T | 225+Phe+Pro+Pyr+TzlCOOMe | This study |
| 683 (2) | 684 | 12.34 | T | Pre+207+Val+Pro+Pyr+TzlCOOMe | This study |
| 685 (2) | 686 | 12.62 | T | Pre+Phe+Hph/MePhe+Pro+Pyr+TzlCOOMe | This study |
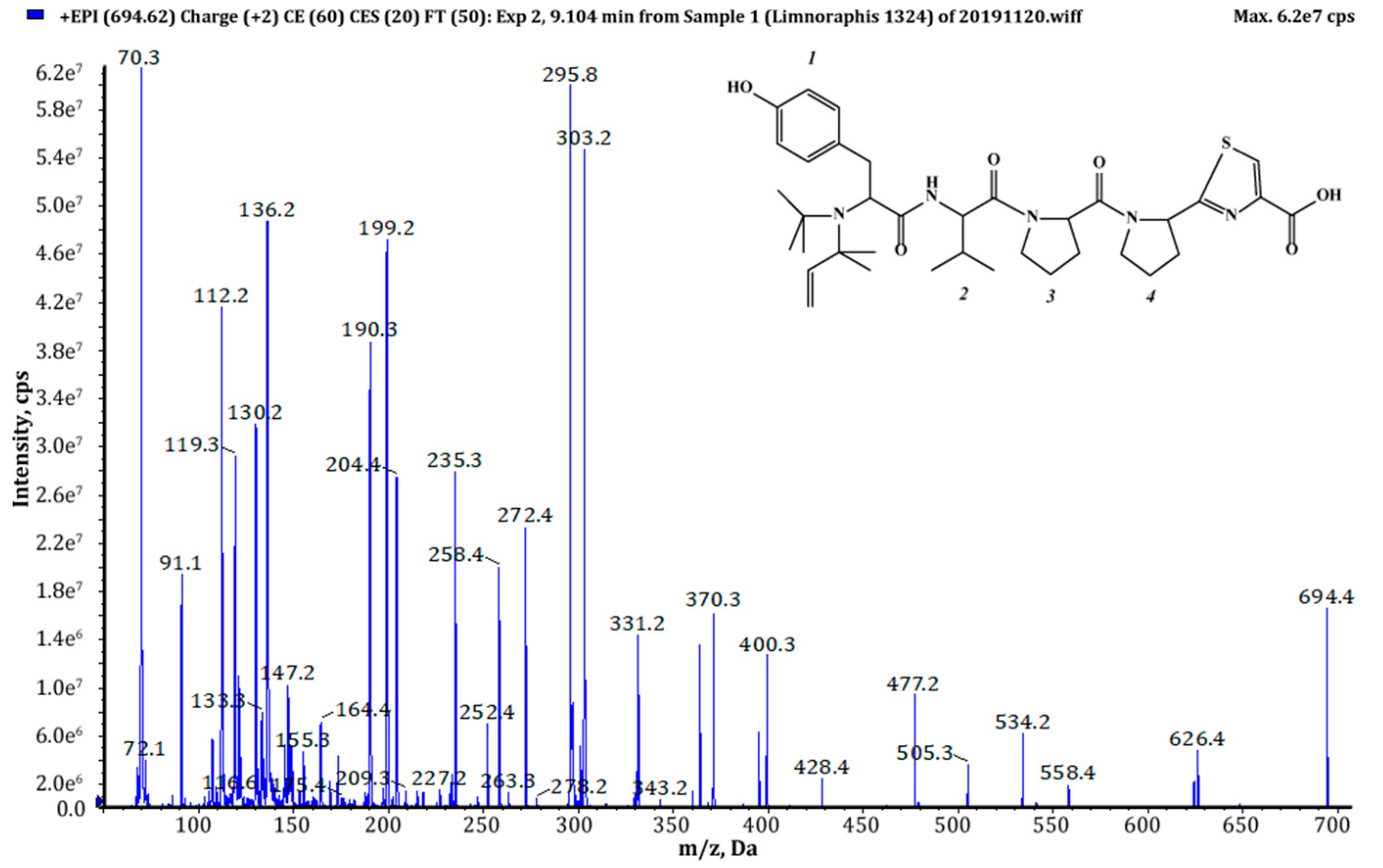
| 693 (1),(2) | 694 | 9.10 | 2.85 × 109 | (Pre)2+Tyr+Val+Pro+Pyr+TzlCOOH | This study |
| 705 (2) | 706 | 8.73 | T | (Pre)2+Hph/MePhe+Val+Pro+Pyr+TzlCOOMe | This study |
| 707 (1),(2) | 708 | 9.76 | 8.69 × 109 | (Pre)2+Tyr+Val+Pro+Pyr+TzlCOOMe | This study |
| 721 (1),(2) | 722 | 10.12 | 1.89 × 108 | (Pre)2+Tyr+Ile/Leu+Pro+Pyr+TzlCOOMe | This study |
| 735 (2) | 736 | 11.26 | T | Pre+225+Phe+Pro+Pyr+TzlCOOH | This study |
| 749 (2) | 750 | 11.50 | T | Pre+225+Phe+Pro+Pyr+TzlCOOMe | This study |
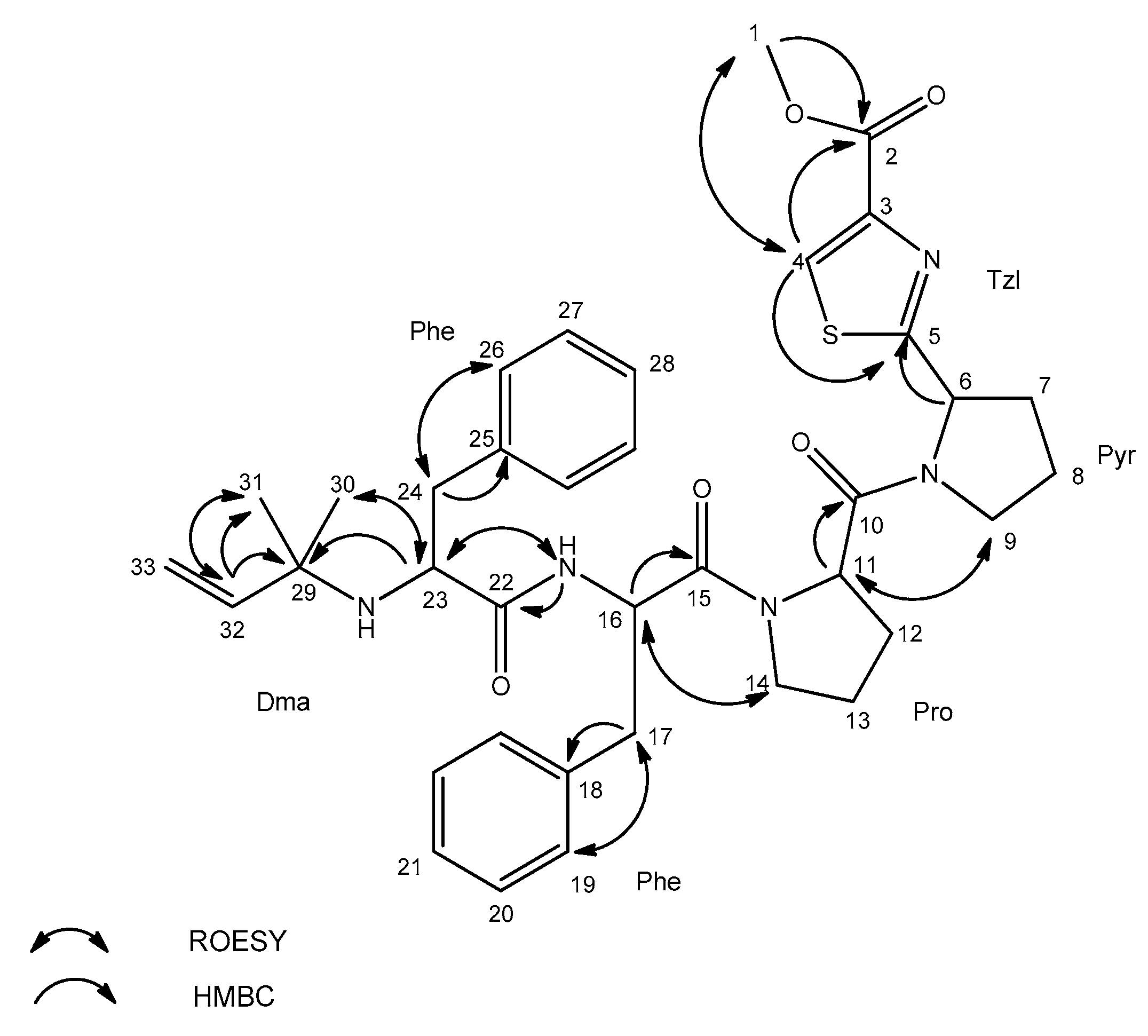
| Residue | Position | äC, Type | äH (J in Hz) | ROESY a | HMBC b |
|---|---|---|---|---|---|
| Tzl-COOMe | 1 | 51.1, CH3 | 3.81, s | 4 | 2 |
| 2 | 160.3, C | ||||
| 3 | 144.5, C | ||||
| 4 | 128.0, CH | 8.43, s | 1 | 2, 5 | |
| 5 | 173.4, C | ||||
| Pyr | 6 | 57.5, CH | 5.32, dd (8.2, 2.4) | 5 | |
| 7 | 30.5, CH2 | 2.05, m | |||
| 2.27, m | |||||
| 8 | 23.3, CH2 | 1.87, m | |||
| 1.96, m | |||||
| 9 | 45.9, CH2 | 3.70, m | 11 | ||
| Pro | 10 | 169.7, C | |||
| 11 | 56.7, CH | 4.68, brs | 9 | 10 | |
| 12 | 27.1, CH2 | 1.81, m | |||
| 2.20, m | |||||
| 13 | 23.7, CH2 | 2.04, m | |||
| 14 | 46.0, CH2 | 3.50, m | 16 | ||
| 3.66, m | |||||
| Phe | 15 | 168.2, C | |||
| 16 | 50.0, CH | 4.77, brs | 14 | 15, 18 | |
| 17 | 36.1, CH2 | 2.76, dd (14.1, 8.8) | 19 | 15, 16, 18 | |
| 2.97, dd (14.1, 4.3) | |||||
| 18 | 136.3, C | ||||
| 19 | 128.6, CH | 7.11, d (7.1) | 17 | ||
| 20 | 127.1, CH | 7.23, m | |||
| 21 | 125.3, CH | 7.18, m | |||
| NH(1) | 8.18, d (8.9) | 23 | 22 | ||
| Phe | 22 | 173.3, C | |||
| 23 | 57.1, CH | 3.03, dd (9.0, 4.2) | 30, NH(1) | 29 | |
| 24 | 39.6, CH2 | 2.35, dd (13.3, 9.0) | 26 | 22, 23, 25 | |
| 2.67, dd (13.3, 4.1) | |||||
| 25 | 137.4, C | ||||
| 26 | 128.6, CH | 7.11, d (7.1) | 24 | ||
| 27 | 127.1, CH | 7.23, m | |||
| 28 | 125.4, CH | 7.18, m | |||
| NH(2) | |||||
| Dma c | 29 | 53.2, C | |||
| 30 | 24.7, CH3 | 0.79, s | 23 | ||
| 31 | 26.6, CH3 | 0.76, s | 32 | ||
| 32 | 144.9, CH | 5.26, dd (17.5, 10.7) | 31 | 29, 31 | |
| 33 | 110.9, CH2 | 4.72, m |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cegłowska, M.; Szubert, K.; Wieczerzak, E.; Kosakowska, A.; Mazur-Marzec, H. Eighteen New Aeruginosamide Variants Produced by the Baltic Cyanobacterium Limnoraphis CCNP1324. Mar. Drugs 2020, 18, 446. https://0-doi-org.brum.beds.ac.uk/10.3390/md18090446
Cegłowska M, Szubert K, Wieczerzak E, Kosakowska A, Mazur-Marzec H. Eighteen New Aeruginosamide Variants Produced by the Baltic Cyanobacterium Limnoraphis CCNP1324. Marine Drugs. 2020; 18(9):446. https://0-doi-org.brum.beds.ac.uk/10.3390/md18090446
Chicago/Turabian StyleCegłowska, Marta, Karolia Szubert, Ewa Wieczerzak, Alicja Kosakowska, and Hanna Mazur-Marzec. 2020. "Eighteen New Aeruginosamide Variants Produced by the Baltic Cyanobacterium Limnoraphis CCNP1324" Marine Drugs 18, no. 9: 446. https://0-doi-org.brum.beds.ac.uk/10.3390/md18090446