
Expression and Biochemical Characterization of a Novel Marine Chitosanase from Streptomyces niveus Suitable for Preparation of Chitobiose


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
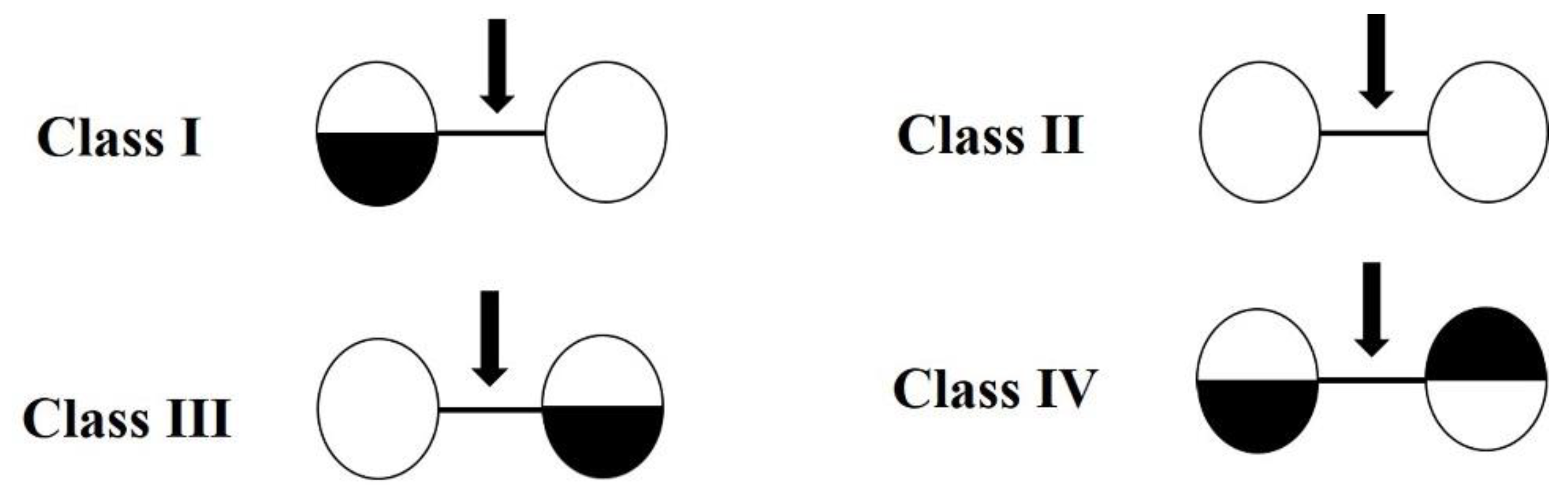
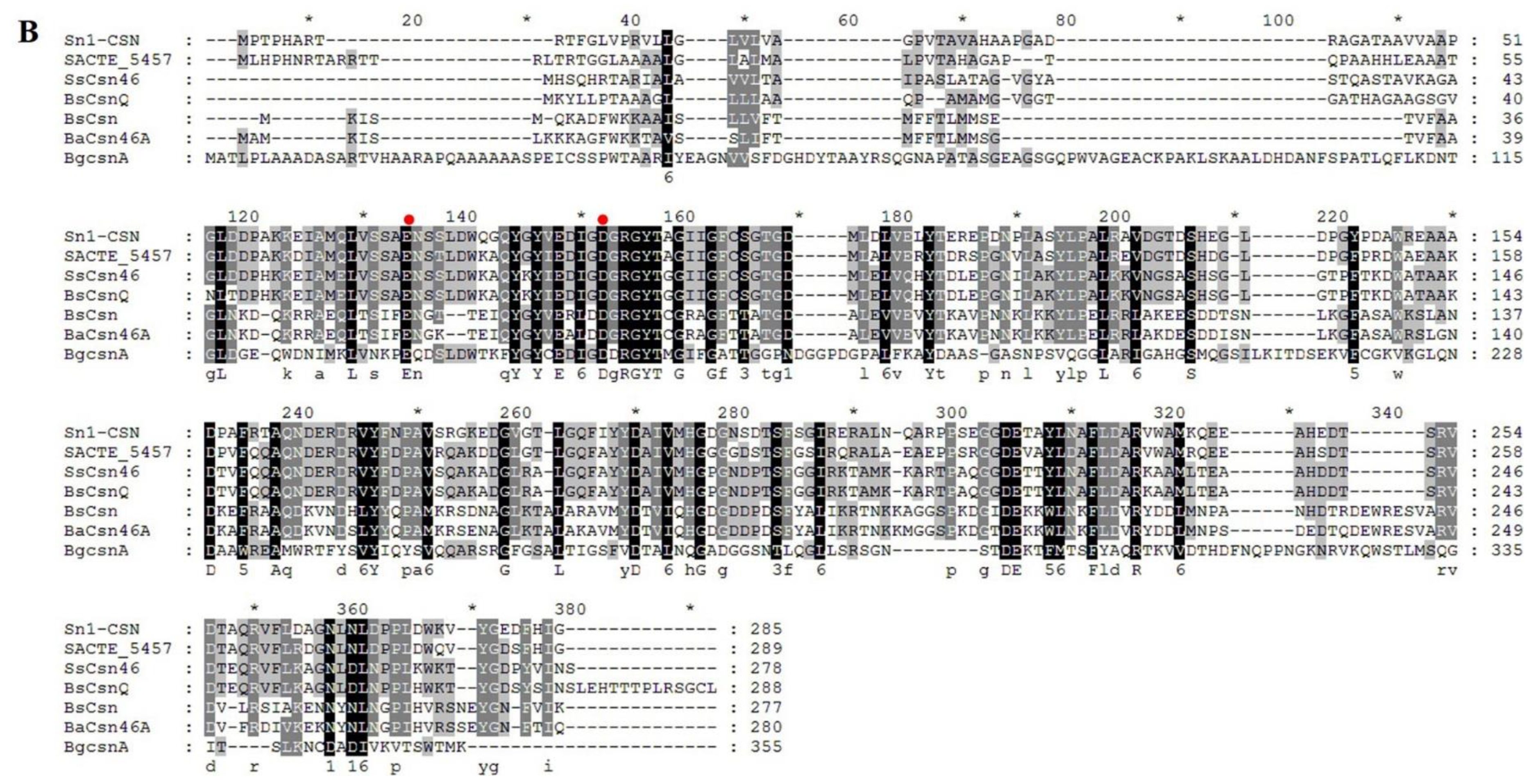
2.1. Sequence Alignment and Phylogenetic Analysis of Sn1-CSN with Characterized GH 46 Chitosanases


2.2. Overexpression and Purification of Sn1-CSN in E. coli
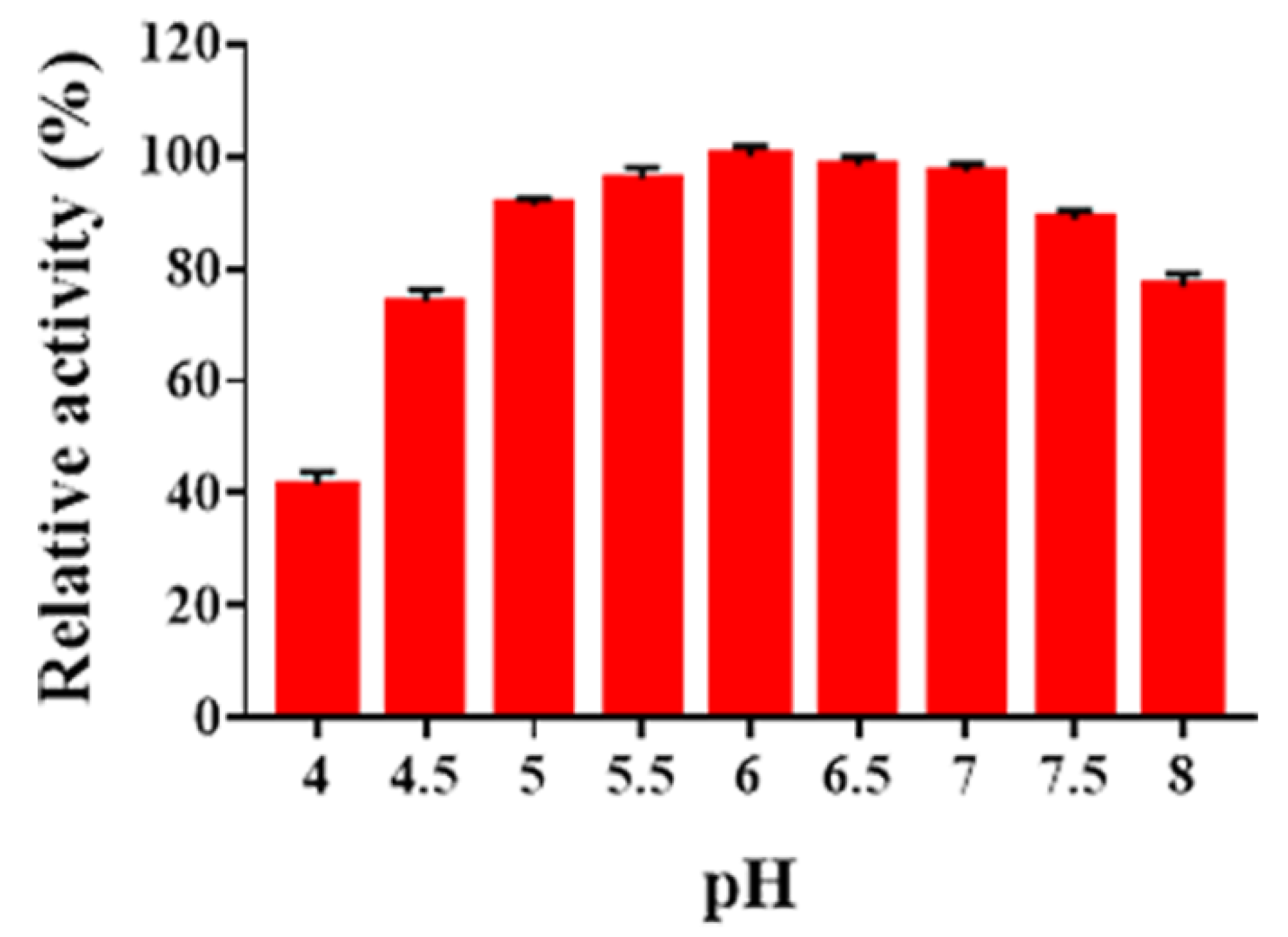
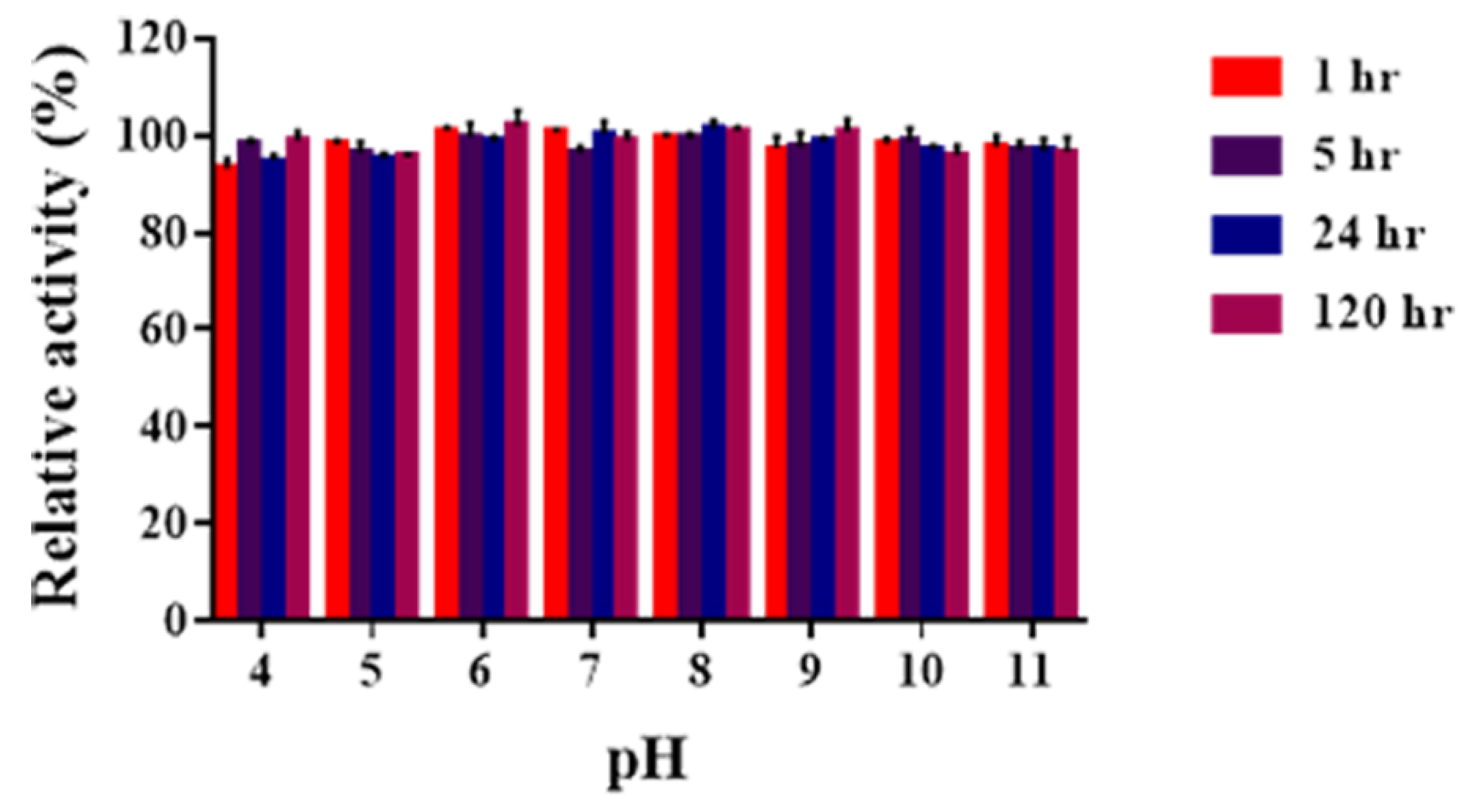
2.3. Determination of pH Optima and pH Stability of Sn1-CSN
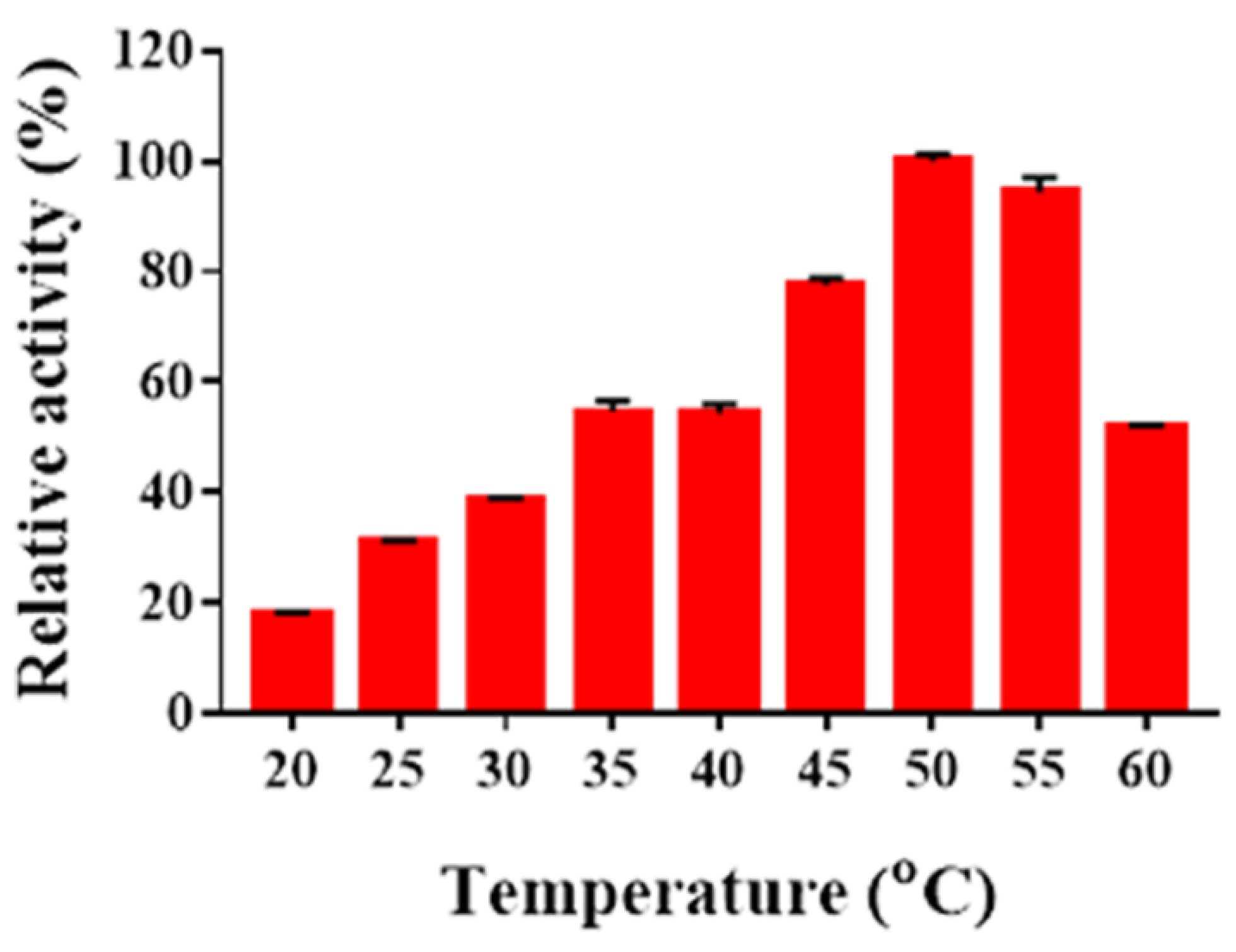
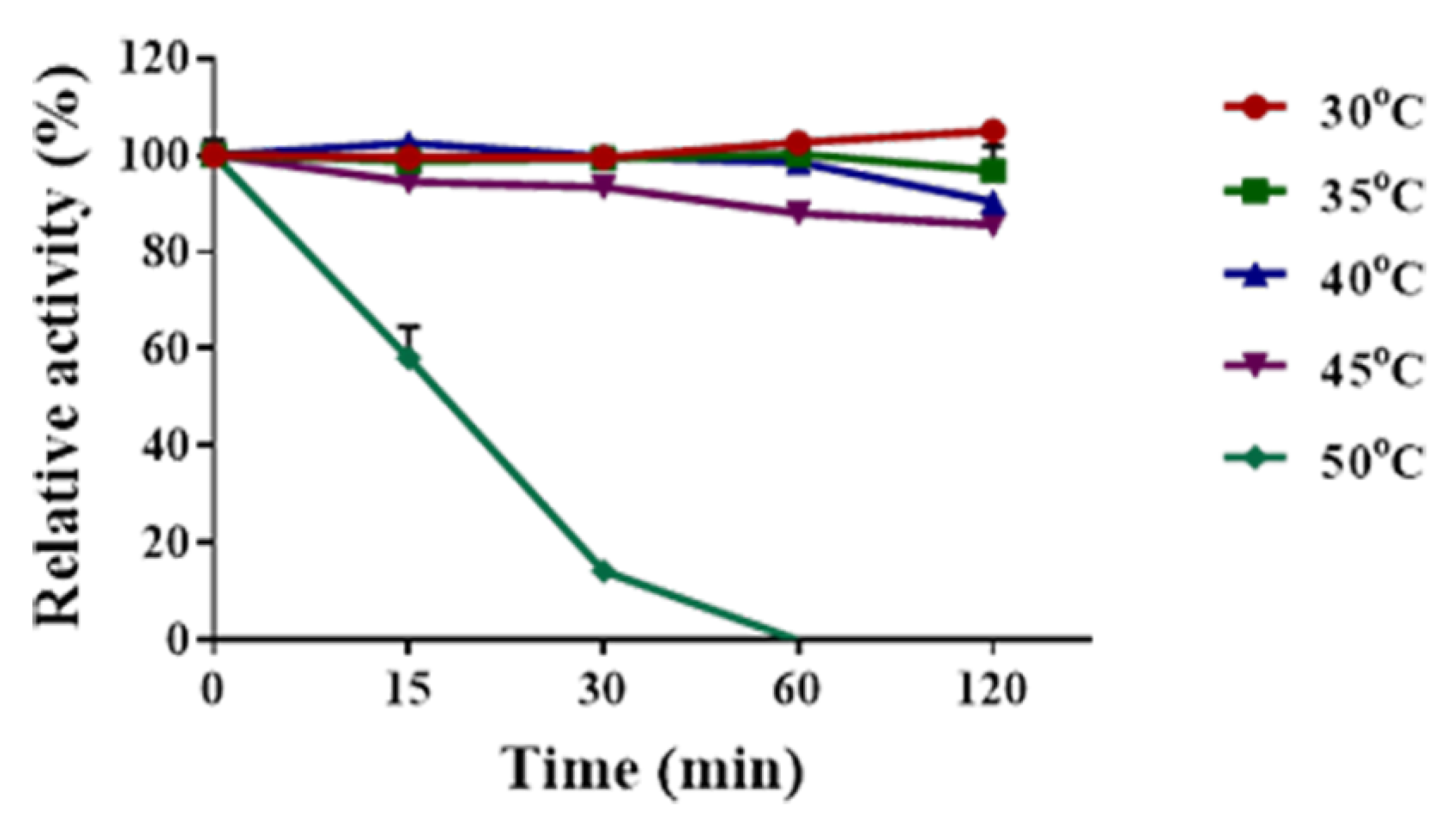
2.4. Determination of Temperature Optima and Thermal Stability of Sn1-CSN
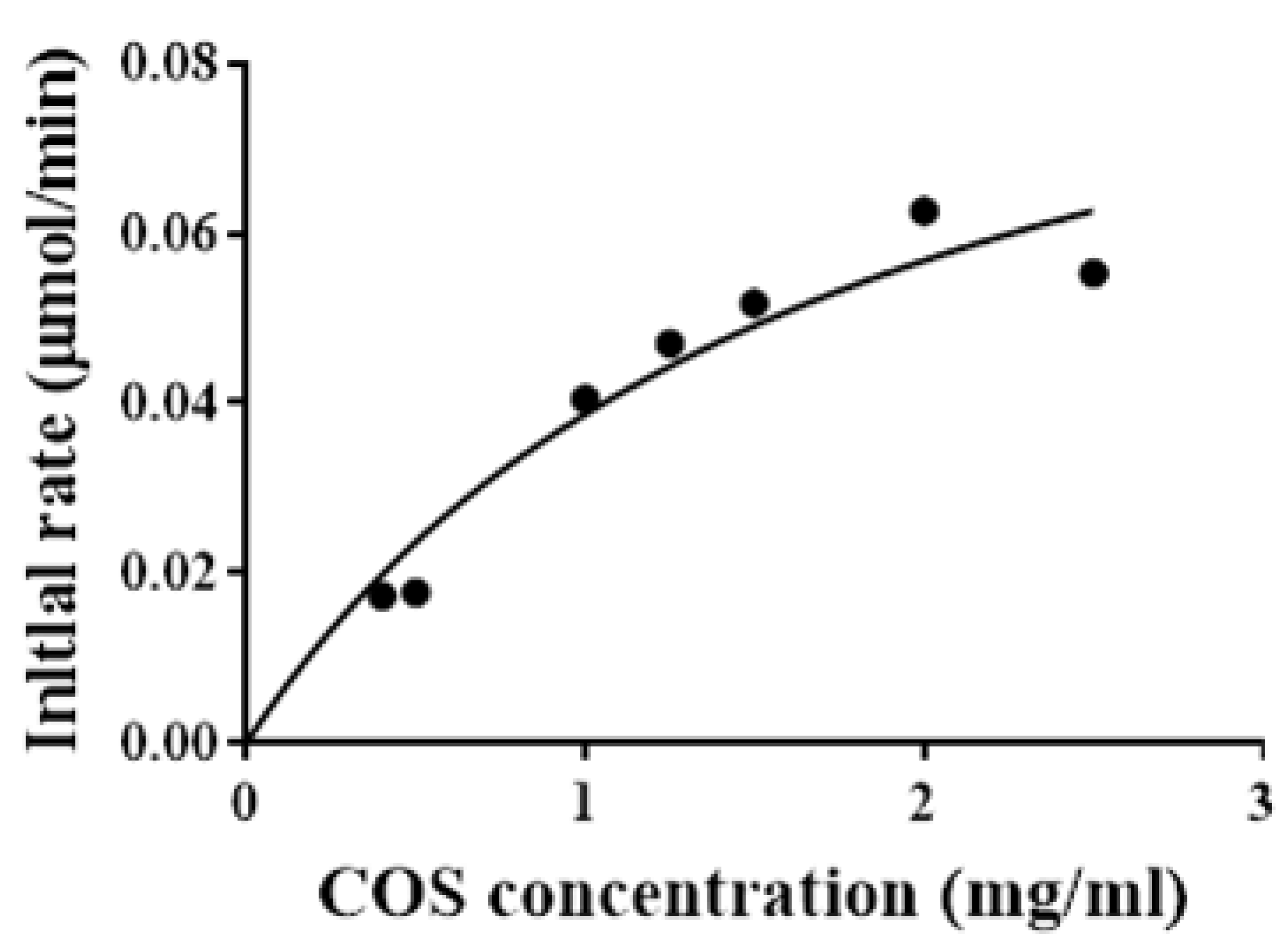
2.5. Determination of Kinetic Parameters of Sn1-CSN
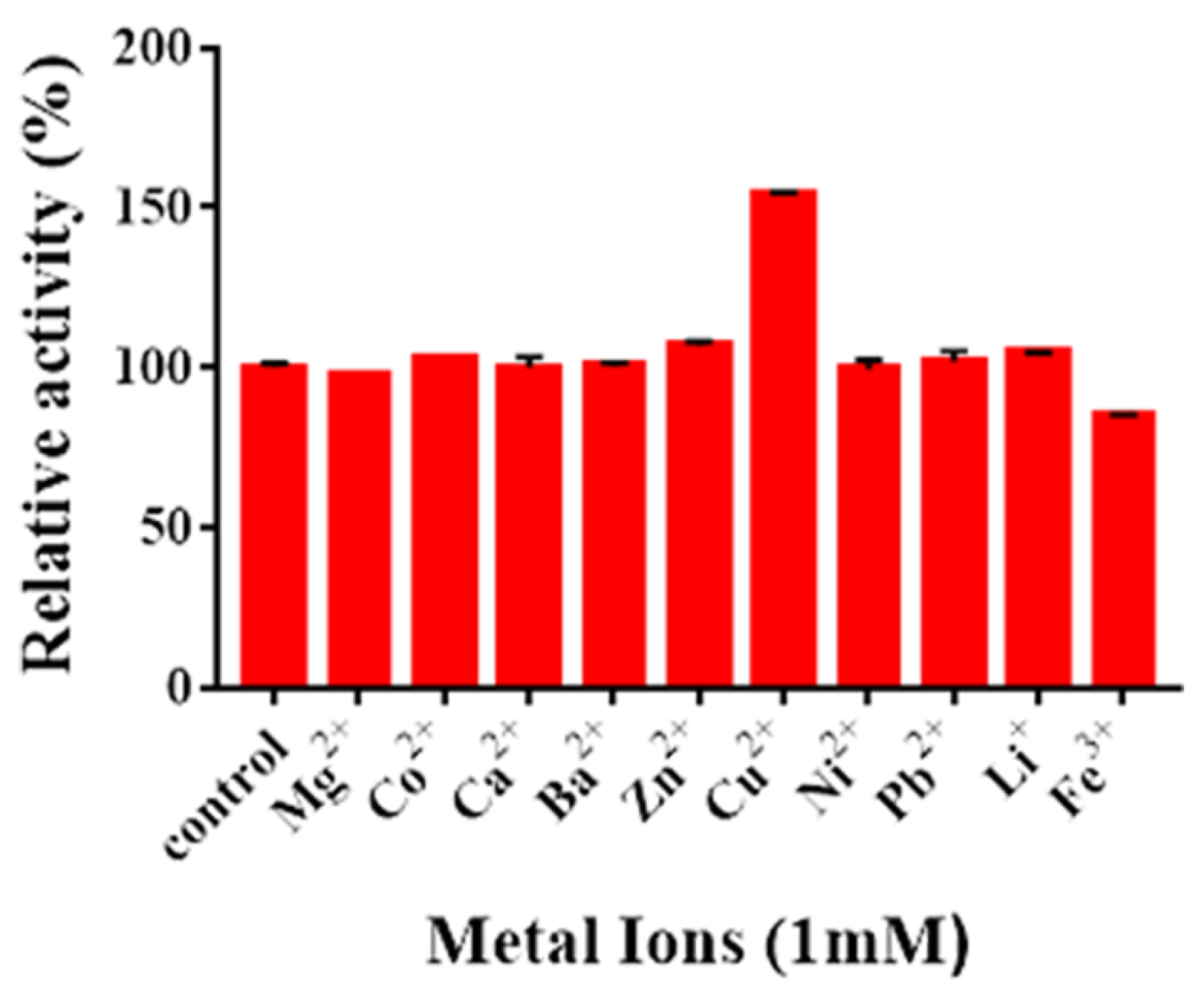
2.6. Effects of Metal Ions on Enzyme Activity
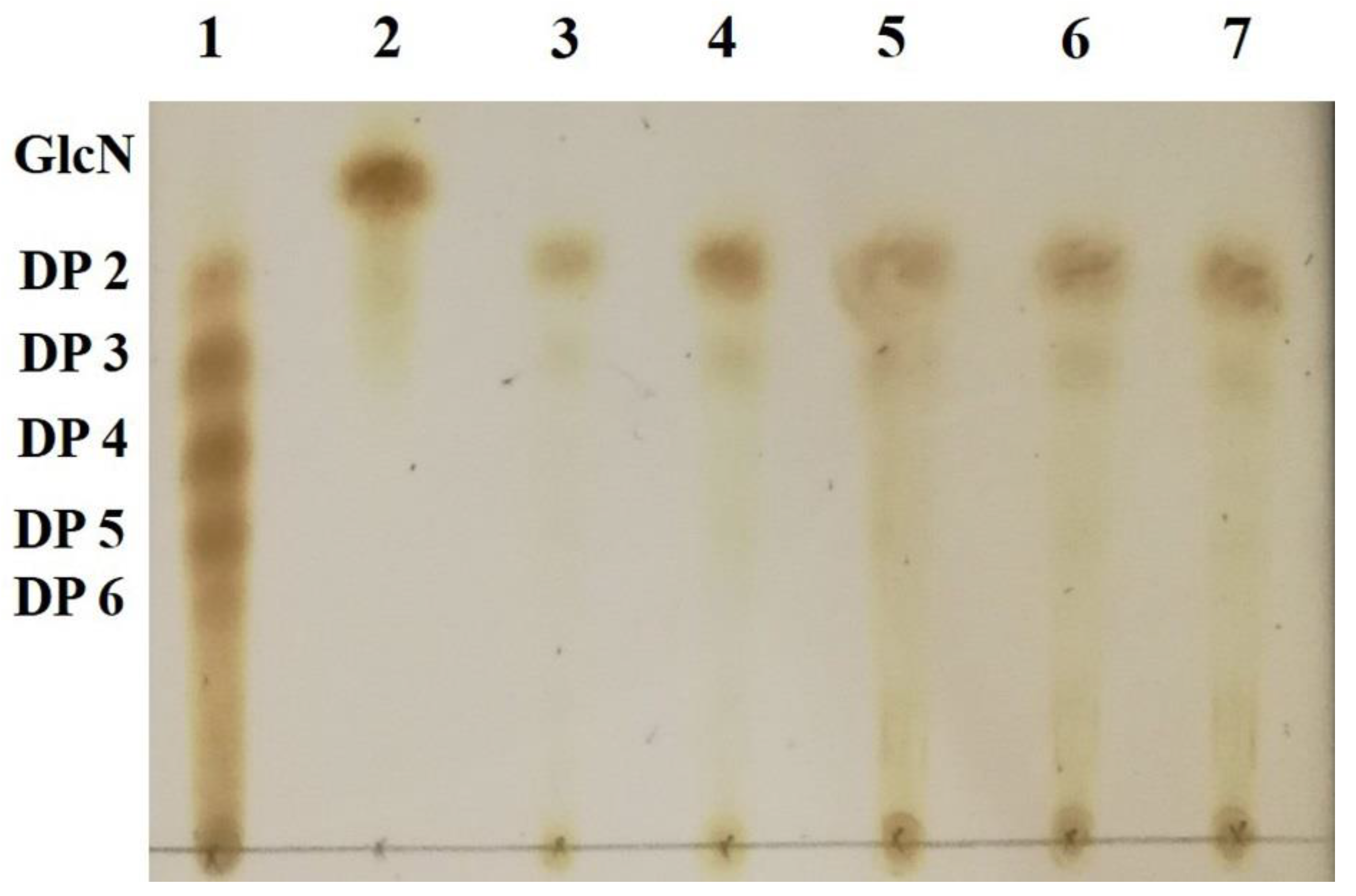
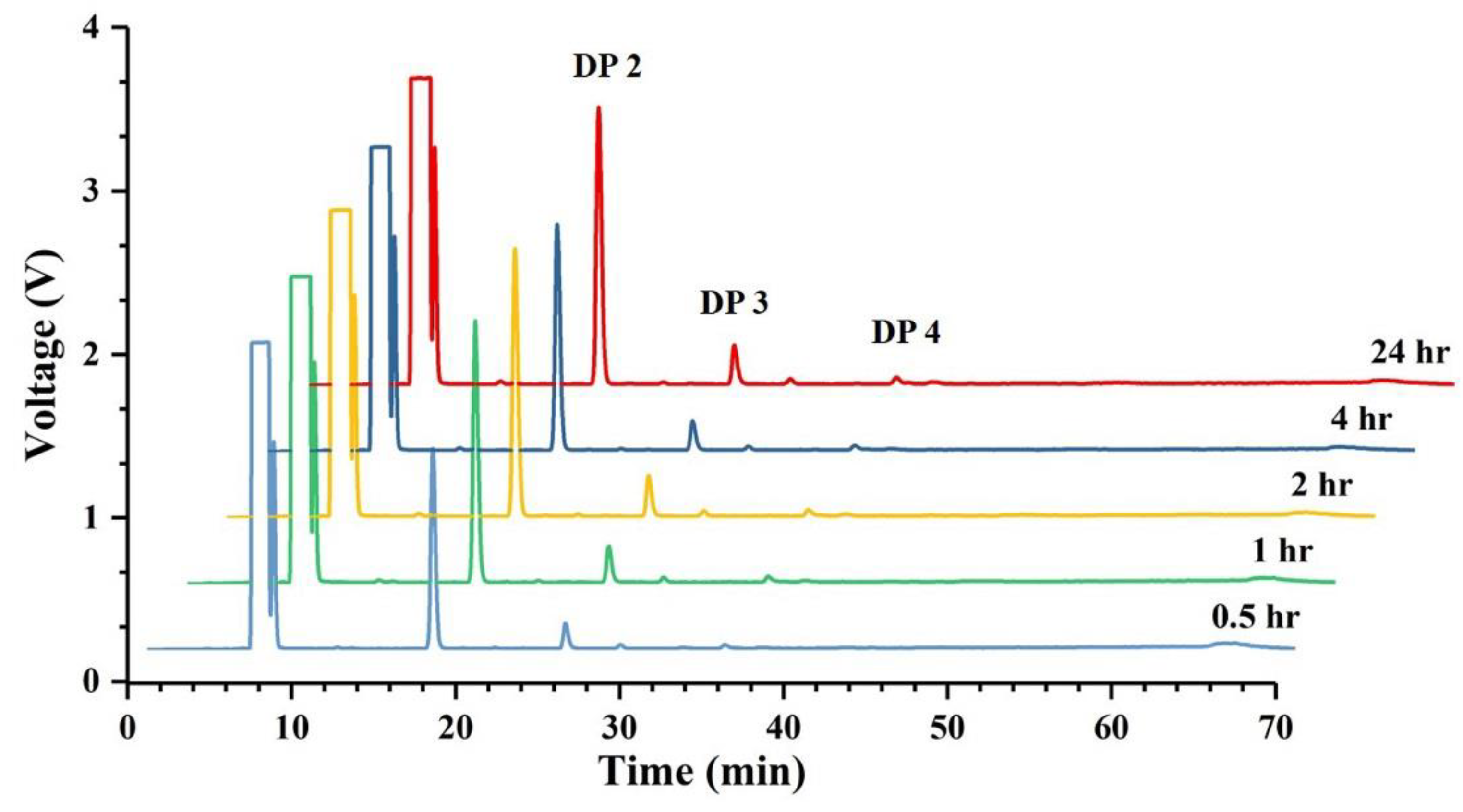
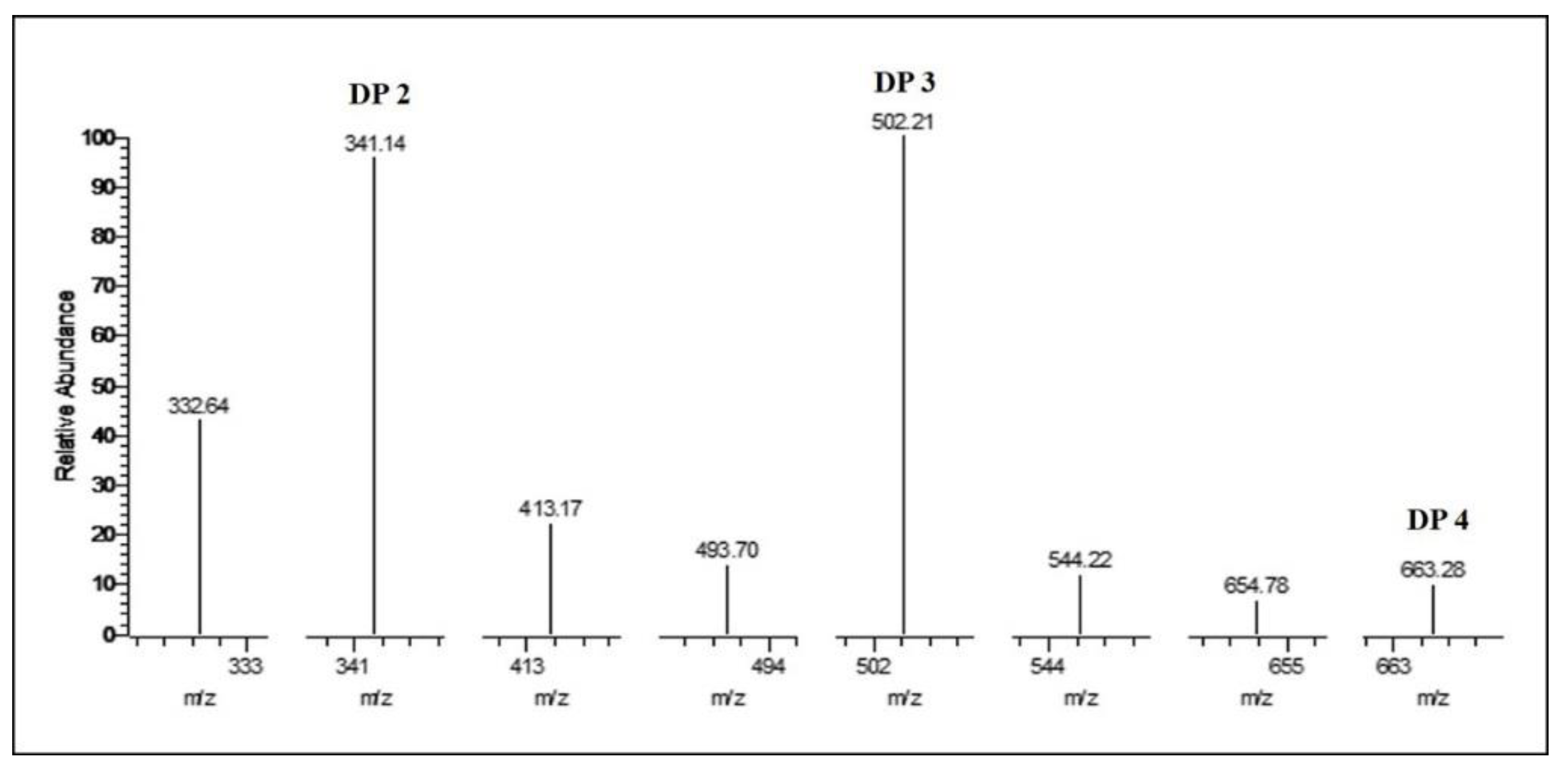
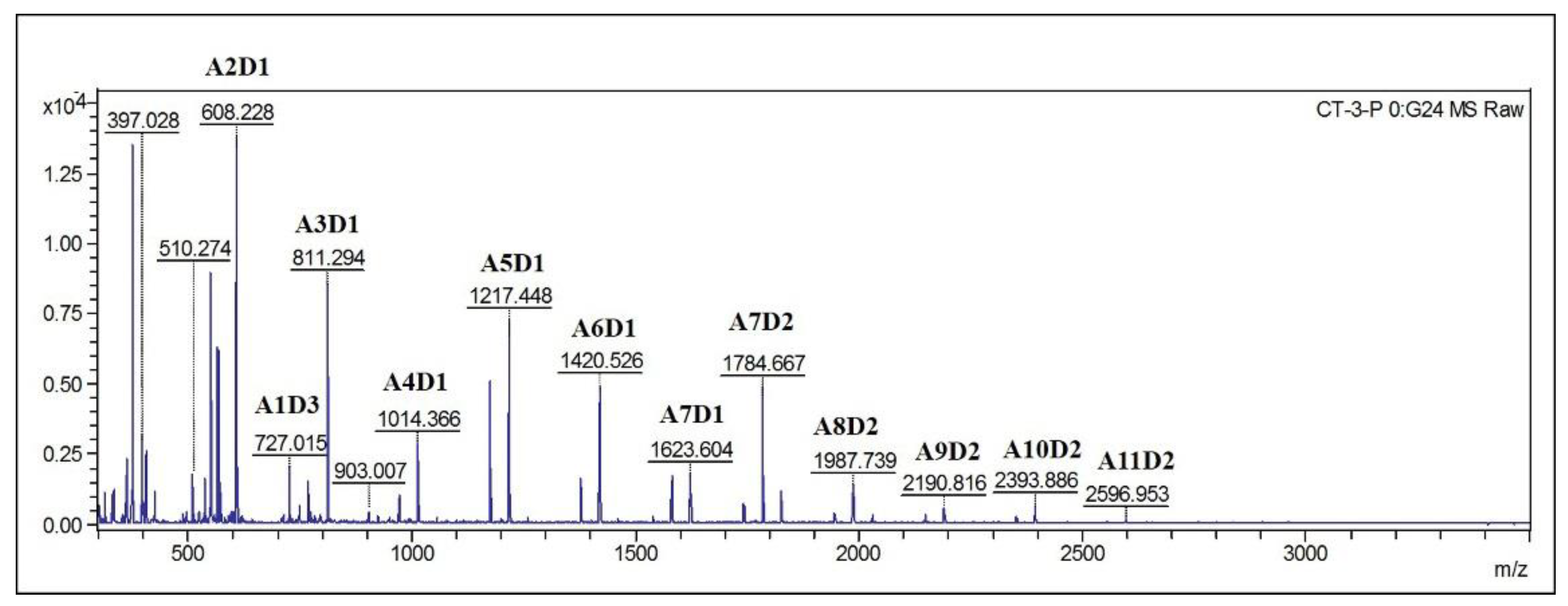
2.7. Identification of Hydrolysis Products of Chitosan Catalyzed by Sn1-CSN
2.8. Analysis of Hydrolysis Products of Partially Acetylated Chitosan Catalyzed by Sn1-CSN
3. Materials and Methods
3.1. Materials
3.2. Bacterial Strains, Plasmids, and Media
3.3. Sequence Analysis and Phylogenetic Tree Analysis
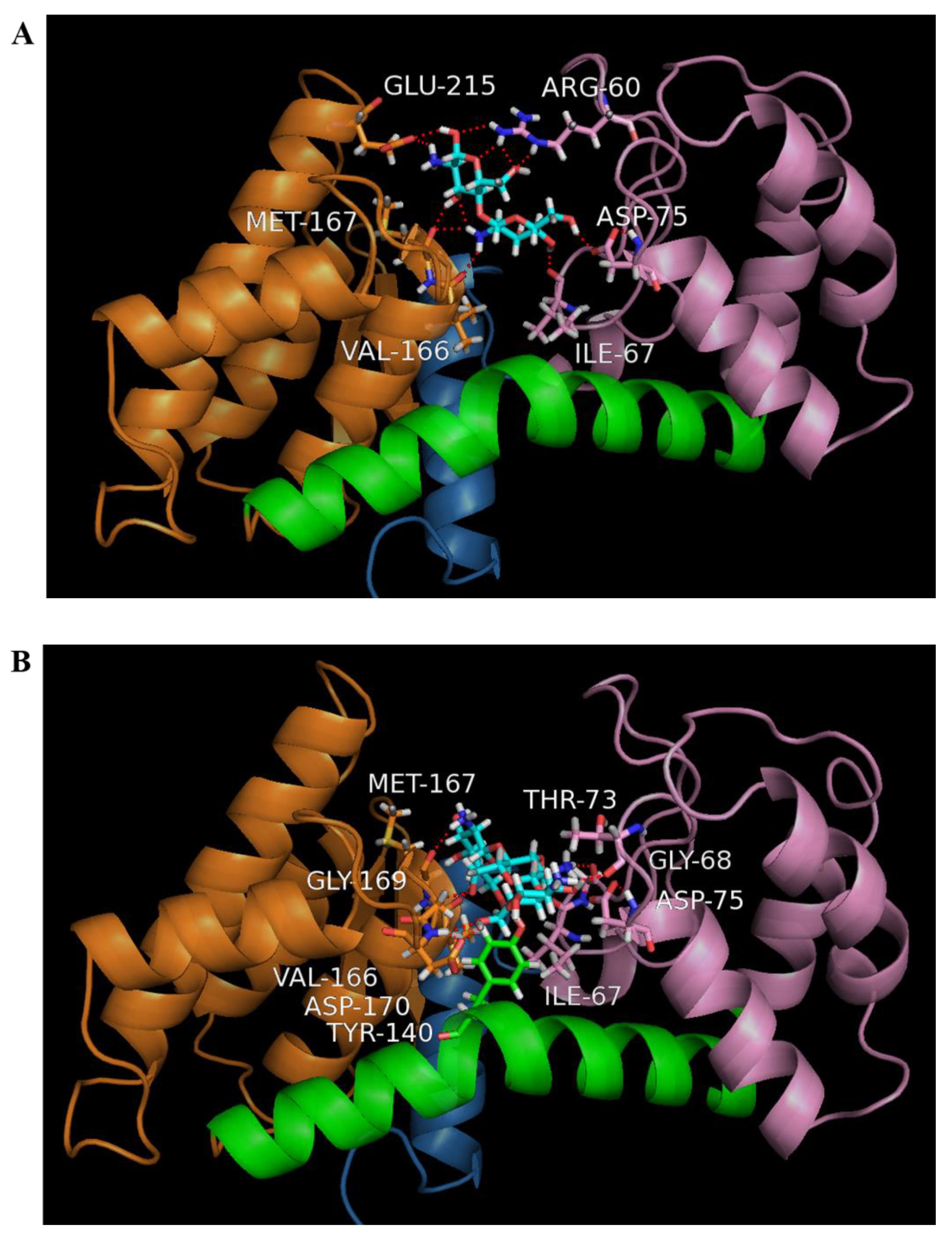
3.4. Homology Modelling of Sn1-CSN and Molecular Docking
3.5. DNA Manipulation
3.6. Protein Over-Expression and Purification
3.7. Enzyme Assay
3.8. Determination of Optimal pH and pH Stability
3.9. Determination of Optimal Temperature and Thermal Stability
3.10. Determination of Kinetic Parameters
3.11. Effects of Metal Ions on Enzyme Activity
3.12. Hydrolysis of Chitosan by Sn1-CSN and Characterization
3.13. Hydrolysis of Partially Acetylated Chitosan by Sn1-CSN and Characterization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Croisier, F.; Jérôme, C. Chitosan-based biomaterials for tissue engineering. Eur. Polym. J. 2013, 49, 780–792. [Google Scholar] [CrossRef] [Green Version]
- Tahtat, D.; Boutrig, H.H.; Khodja, A.N.; Benamer, S.; Hammache, Y.; Mahlous, M. The synergistic effect of gamma irradiation and alkaline soaking at low temperature on the pre-deacetylation of α-chitin: Optimization by design of experiment. Carbohydr. Polym. 2019, 215, 39–46. [Google Scholar] [CrossRef]
- Yang, G.; Sun, H.; Cao, R.; Liu, Q.; Mao, X. Characterization of a novel glycoside hydrolase family 46 chitosanase, Csn-BAC, from Bacillus sp. MD-5. Int. J. Biol. Macromol. 2020, 146, 518–523. [Google Scholar] [CrossRef]
- Kulikov, S.N.; Chirkov, S.N.; Il’Ina, A.V.; Lopatin, S.A.; Varlamov, V.P. Effect of the molecular weight of chitosan on its antiviral activity in plants. Appl. Biochem. Microbiol. 2006, 42, 200–203. [Google Scholar] [CrossRef]
- Rahman, H.; Hjeljord, L.G.; Aam, B.B.; Sørlie, M.; Tronsmo, A. Antifungal effect of chito-oligosaccharides with different degrees of polymerization. Eur. J. Plant Pathol. 2014, 141, 147–158. [Google Scholar] [CrossRef]
- Cabrera, J.C.; Van Cutsem, P. Preparation of chitooligosaccharides with degree of polymerization higher than 6 by acid or enzymatic degradation of chitosan. Biochem. Eng. J. 2005, 25, 165–172. [Google Scholar] [CrossRef]
- Gohel, V.; Singh, A.; Maisuria, V.; Phadnis, A.; Chhatpar, H.S. Bioprospecting and antifungal potential of chitinolytic microor-ganisms. Afr. J. Biotechnol. 2006, 5, 54–72. [Google Scholar]
- Thadathil, N.; Velappan, S.P. Recent developments in chitosanase research and its biotechnological applications: A review. Food Chem. 2014, 150, 392–399. [Google Scholar] [CrossRef]
- Viens, P.; Lacombe-Harvey, M.-È.; Brzezinski, R. Chitosanases from Family 46 of Glycoside Hydrolases: From Proteins to Phenotypes. Mar. Drugs 2015, 13, 6566–6587. [Google Scholar] [CrossRef]
- Qin, Z.; Chen, Q.; Lin, S.; Luo, S.; Qiu, Y.; Zhao, L. Expression and characterization of a novel cold-adapted chitosanase suitable for chitooligosaccharides controllable preparation. Food Chem. 2018, 253, 139–147. [Google Scholar] [CrossRef]
- Heggset, E.B.; Tuveng, T.R.; Hoell, I.A.; Liu, Z.; Eijsink, V.G.H.; Vårum, K.M. Mode of Action of a Family 75 Chitosanase from Streptomyces avermitilis. Biomacromolecules 2012, 13, 1733–1741. [Google Scholar] [CrossRef] [Green Version]
- Yorinaga, Y.; Kumasaka, T.; Yamamoto, M.; Hamada, K.; Kawamukai, M. Crystal structure of a family 80 chitosanase fromMitsuaria chitosanitabida. FEBS Lett. 2017, 591, 540–547. [Google Scholar] [CrossRef] [Green Version]
- Zitouni, M.; Viens, P.; Ghinet, M.G.; Brzezinski, R. Diversity of family GH46 chitosanases in Kitasatospora setae KM-6054. Appl. Microbiol. Biotechnol. 2017, 101, 7877–7888. [Google Scholar] [CrossRef] [Green Version]
- Pechsrichuang, P.; Lorentzen, S.B.; Aam, B.B.; Tuveng, T.R.; Hamre, A.G.; Eijsink, V.G.; Yamabhai, M. Bioconversion of chitosan into chito-oligosaccharides (CHOS) using family 46 chitosanase from Bacillus subtilis (BsCsn46A). Carbohydr. Polym. 2018, 186, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, X.; Li, X.; Han, Y.; Wang, Y.; Yao, R.; Li, S. Purification and Characterization of A New Cold-Adapted and Thermo-Tolerant Chitosanase from Marine Bacterium Pseudoalteromonas sp. SY39. Molecules 2019, 24, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, M.; Zhang, T.; Sun, C.; Zhang, H.; Zhang, Y. A Chitosanase mutant from Streptomyces sp. N174 prefers to produce functional chitopentasaccharide. Int. J. Biol. Macromol. 2020, 151, 1091–1098. [Google Scholar] [CrossRef]
- Hirano, K.; Watanabe, M.; Seki, K.; Ando, A.; Saito, A.; Mitsutomi, M. Classification of Chitosanases by Hydrolytic Specificity toward N1,N4-Diacetylchitohexaose. Biosci. Biotechnol. Biochem. 2012, 76, 1932–1937. [Google Scholar] [CrossRef] [Green Version]
- Weikert, T.; Niehues, A.; Cord-Landwehr, S.; Hellmann, M.J.; Moerschbacher, B.M. Reassessment of chitosanase substrate specificities and classification. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.-S.; Taguchi, T.; Sakai, K.; Matahira, Y.; Wang, M.-W.; Miwa, I. Effect of Chitobiose and Chitotriose on Carbon Tetrachloride-Induced Acute Hepatotoxicity in Rats. Biol. Pharm. Bull. 2005, 28, 1971–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhao, M.; Fan, L.; Cao, X.; Chen, L.; Chen, J.; Lo, Y.M.; Zhao, L. Chitobiose alleviates oleic acid-induced lipid accumulation by decreasing fatty acid uptake and triglyceride synthesis in HepG2 cells. J. Funct. Foods 2018, 46, 202–211. [Google Scholar] [CrossRef]
- Li, K.; Xing, R.; Liu, S.; Qin, Y.; Yu, H.; Li, P. Size and pH effects of chitooligomers on antibacterial activity against Staphylococcus aureus. Int. J. Biol. Macromol. 2014, 64, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Zou, P.; Tian, X.; Dong, B.; Zhang, C. Size effects of chitooligomers with certain degrees of polymerization on the chilling tolerance of wheat seedlings. Carbohydr. Polym. 2017, 160, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Zou, P.; Li, K.; Liu, S.; Xing, R.; Qin, Y.; Yu, H.; Zhou, M.; Li, P. Effect of chitooligosaccharides with different degrees of acetylation on wheat seedlings under salt stress. Carbohydr. Polym. 2015, 126, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Takasuka, T.E.; Bianchetti, C.M.; Tobimatsu, Y.; Bergeman, L.F.; Ralph, J.; Fox, B.G. Structure-guided analysis of catalytic specificity of the abundantly secreted chitosanase SACTE_5457 from Streptomyces sp. SirexAA-E. Proteins Struct. Funct. Bioinform. 2014, 82, 1245–1257. [Google Scholar] [CrossRef] [PubMed]
- Pechsrichuang, P.; Yoohat, K.; Yamabhai, M. Production of recombinant Bacillus subtilis chitosanase, suitable for biosynthesis of chitosan-oligosaccharides. Bioresour. Technol. 2013, 127, 407–414. [Google Scholar] [CrossRef]
- Marcotte, E.M.; Monzingo, A.F.; Ernst, S.R.; Brzezinski, R.; Robertas, J.D. X-ray structure of an anti-fungal chitosanase from streptomyces N174. Nat. Genet. 1996, 3, 155–162. [Google Scholar] [CrossRef]
- Guo, N.; Sun, J.; Wang, W.; Gao, L.; Liu, J.; Liu, Z.; Xue, C.; Mao, X. Cloning, expression and characterization of a novel chitosanase from Streptomyces albolongus ATCC 27414. Food Chem. 2019, 286, 696–702. [Google Scholar] [CrossRef]
- Luo, S.; Qin, Z.; Chen, Q.; Fan, L.; Jiang, L.; Zhao, L. High level production of a Bacillus amlyoliquefaciens chitosanase in Pichia pastoris suitable for chitooligosaccharides preparation. Int. J. Biol. Macromol. 2020, 149, 1034–1041. [Google Scholar] [CrossRef]
- Sun, H.; Cao, R.; Li, L.; Zhao, L.; Liu, Q. Cloning, purification and characterization of a novel GH46 family chitosanase, Csn-CAP, from Staphylococcus capitis. Process. Biochem. 2018, 75, 146–151. [Google Scholar] [CrossRef]
- Yoon, H.-G.; Lee, K.-H.; Kim, H.-Y.; Kim, H.-K.; Shin, D.-H.; Hong, B.-S.; Cho, H.-Y. Gene Cloning and Biochemical Analysis of Thermostable Chitosanase (TCH-2) fromBacillus coagulansCK108. Biosci. Biotechnol. Biochem. 2002, 66, 986–995. [Google Scholar] [CrossRef]
- Lee, Y.S.; Yoo, J.S.; Chung, S.Y.; Cho, Y.S.; Choi, Y.L. Cloning, purification, and characterization of chitosanase from Bacillus sp. DAU101. Appl. Microbiol. Biotechnol. 2006, 73, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.-G.; Kim, H.-Y.; Lim, Y.-H.; Kim, H.-K.; Shin, D.-H.; Hong, B.-S.; Cho, H.-Y. Thermostable Chitosanase from Bacillus sp. Strain CK4: Cloning and Expression of the Gene and Characterization of the Enzyme. Appl. Environ. Microbiol. 2000, 66, 3727–3734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimosaka, M.; Fukumori, Y.; Zhang, X.-Y.; He, N.-J.; Kodaira, R.; Okazaki, M. Molecular cloning and characterization of a chitosanase from the chitosanolytic bacterium Burkholderia gladioli strain CHB101. Appl. Microbiol. Biotechnol. 2000, 54, 354–360. [Google Scholar] [CrossRef]
- Masson, J.-Y.; Boucher, I.; Neugebauer, W.A.; Ramotar, D.; Brzezinski, R. A new chitosanase gene from a Nocardioides sp. is a third member of glycosyl hydrolase family 46. Microbiology 1995, 141, 2629–2635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ando, A.; Saito, A.; Arai, S.; Usuda, S.; Furuno, M.; Kaneko, N.; Shida, O.; Nagata, Y. Molecular Characterization of a Novel Family-46 Chitosanase fromPseudomonassp. A-01. Biosci. Biotechnol. Biochem. 2008, 72, 2074–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghinet, M.G.; Roy, S.; Poulin-Laprade, D.; Lacombe-Harvey, M.-È.; Morosoli, R.; Brzezinski, R. Chitosanase fromStreptomyces coelicolorA3(2): Biochemical properties and role in protection against antibacterial effect of chitosan. Biochem. Cell Biol. 2010, 88, 907–916. [Google Scholar] [CrossRef]
- Li, H.; Fei, Z.; Gong, J.; Yang, T.; Xu, Z.; Shi, J. Screening and characterization of a highly active chitosanase based on metagenomic technology. J. Mol. Catal. B Enzym. 2015, 111, 29–35. [Google Scholar] [CrossRef]
- Sun, H.; Mao, X.; Guo, N.; Zhao, L.; Cao, R.; Liu, Q. Discovery and Characterization of a Novel Chitosanase fromPaenibacillus dendritiformisby Phylogeny-Based Enzymatic Product Specificity Prediction. J. Agric. Food Chem. 2018, 66, 4645–4651. [Google Scholar] [CrossRef]
- Yang, Y.; Zheng, Z.; Xiao, Y.; Zhang, J.; Zhou, Y.; Li, X.; Li, S.; Yu, H. Cloning and Characterization of a Cold-adapted Chitosanase from Marine Bacterium Bacillus sp. BY01. Molecules 2019, 24, 3915. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Li, X.; Yang, K.; Li, S. Characterization of a New Chitosanase from a Marine Bacillus sp. and the Anti-Oxidant Activity of Its Hydrolysate. Mar. Drugs 2020, 18, 126. [Google Scholar] [CrossRef] [Green Version]
- Johnsen, M.G.; Hansen, O.C.; Stougaard, P. Isolation, characterization and heterologous expression of a novel chitosanase from Janthinobacterium sp. strain 4239. Microb. Cell Factories 2010, 9, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wattjes, J.; Niehues, A.; Cord-Landwehr, S.; Hoßbach, J.; David, L.; Delair, T.; Moerschbacher, B.M. Enzymatic Production and Enzymatic-Mass Spectrometric Fingerprinting Analysis of Chitosan Polymers with Different Nonrandom Patterns of Acetylation. J. Am. Chem. Soc. 2019, 141, 3137–3145. [Google Scholar] [CrossRef]
- Schmitz, C.; Auza, L.G.; Koberidze, D.; Rasche, S.; Fischer, R.; Bortesi, L. Conversion of Chitin to Defined Chitosan Oligomers: Current Status and Future Prospects. Mar. Drugs 2019, 17, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, B.; Cheng, G.; Li, J.; Wang, Z.A.; Du, Y. Inhibition of Liver Tumor Cell Metastasis by Partially Acetylated Chitosan Oligosaccharide on A Tumor-Vessel Microsystem. Mar. Drugs 2019, 17, 415. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinform. 2008, 9, 40–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kidibule, P.E.; Santos-Moriano, P.; Jiménez-Ortega, E.; Ramírez-Escudero, M.; Limón, M.C.; Remacha, M.; Plou, F.J.; Sanz-Aparicio, J.; Fernández-Lobato, M. Use of chitin and chitosan to produce new chitooligosaccharides by chitinase Chit42: Enzymatic activity and structural basis of protein specificity. Microb. Cell Factories 2018, 17, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, G.; Ahmad, E.; Khan, M.V.; Ashraf, M.T.; Bhat, R.; Khan, R.H. Impact of structural stability of cold adapted Candida antarctica lipase B (CaLB): In relation to pH, chemical and thermal denaturation. RSC Adv. 2015, 5, 20115–20131. [Google Scholar] [CrossRef]
- Rabbani, G.; Ahmad, E.; Zaidi, N.; Fatima, S.; Khan, R.H. pH-Induced Molten Globule State of Rhizopus niveus Lipase is More Resistant Against Thermal and Chemical Denaturation Than Its Native State. Cell Biophys. 2012, 62, 487–499. [Google Scholar] [CrossRef]
- Keisuke, K.T.S.; Iwakura, Y. Studies on Chitin, 4: Evidence for Formation of Block and Random Copolymers of N-Acetyl-D-glucosamine and D-Giucosamine by Hetero- and Homogeneous Hydrolyses. Makromol. Chem. 1977, 178, 3197–3202. [Google Scholar]
- Popa-Nita, S.; Lucas, J.-M.; Ladavière, C.; David, L.; Domard, A. Mechanisms Involved During the Ultrasonically Induced Depolymerization of Chitosan: Characterization and Control. Biomacromolecules 2009, 10, 1203–1211. [Google Scholar] [CrossRef] [PubMed]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, T.; Cheng, G.; Jiao, S.; Ren, L.; Zhao, C.; Wei, J.; Han, J.; Pei, M.; Du, Y.; Li, J.-J. Expression and Biochemical Characterization of a Novel Marine Chitosanase from Streptomyces niveus Suitable for Preparation of Chitobiose. Mar. Drugs 2021, 19, 300. https://0-doi-org.brum.beds.ac.uk/10.3390/md19060300
Chen T, Cheng G, Jiao S, Ren L, Zhao C, Wei J, Han J, Pei M, Du Y, Li J-J. Expression and Biochemical Characterization of a Novel Marine Chitosanase from Streptomyces niveus Suitable for Preparation of Chitobiose. Marine Drugs. 2021; 19(6):300. https://0-doi-org.brum.beds.ac.uk/10.3390/md19060300
Chicago/Turabian StyleChen, Tong, Gong Cheng, Siming Jiao, Lishi Ren, Chuanfang Zhao, Jinhua Wei, Juntian Han, Meishan Pei, Yuguang Du, and Jian-Jun Li. 2021. "Expression and Biochemical Characterization of a Novel Marine Chitosanase from Streptomyces niveus Suitable for Preparation of Chitobiose" Marine Drugs 19, no. 6: 300. https://0-doi-org.brum.beds.ac.uk/10.3390/md19060300