
Purification and Characterization of a Novel Alginate Lyase from a Marine Streptomyces Species Isolated from Seaweed

 , and
, and
Abstract
:1. Introduction
2. Results and Discussions
2.1. Identification of Marine Actinobacterium
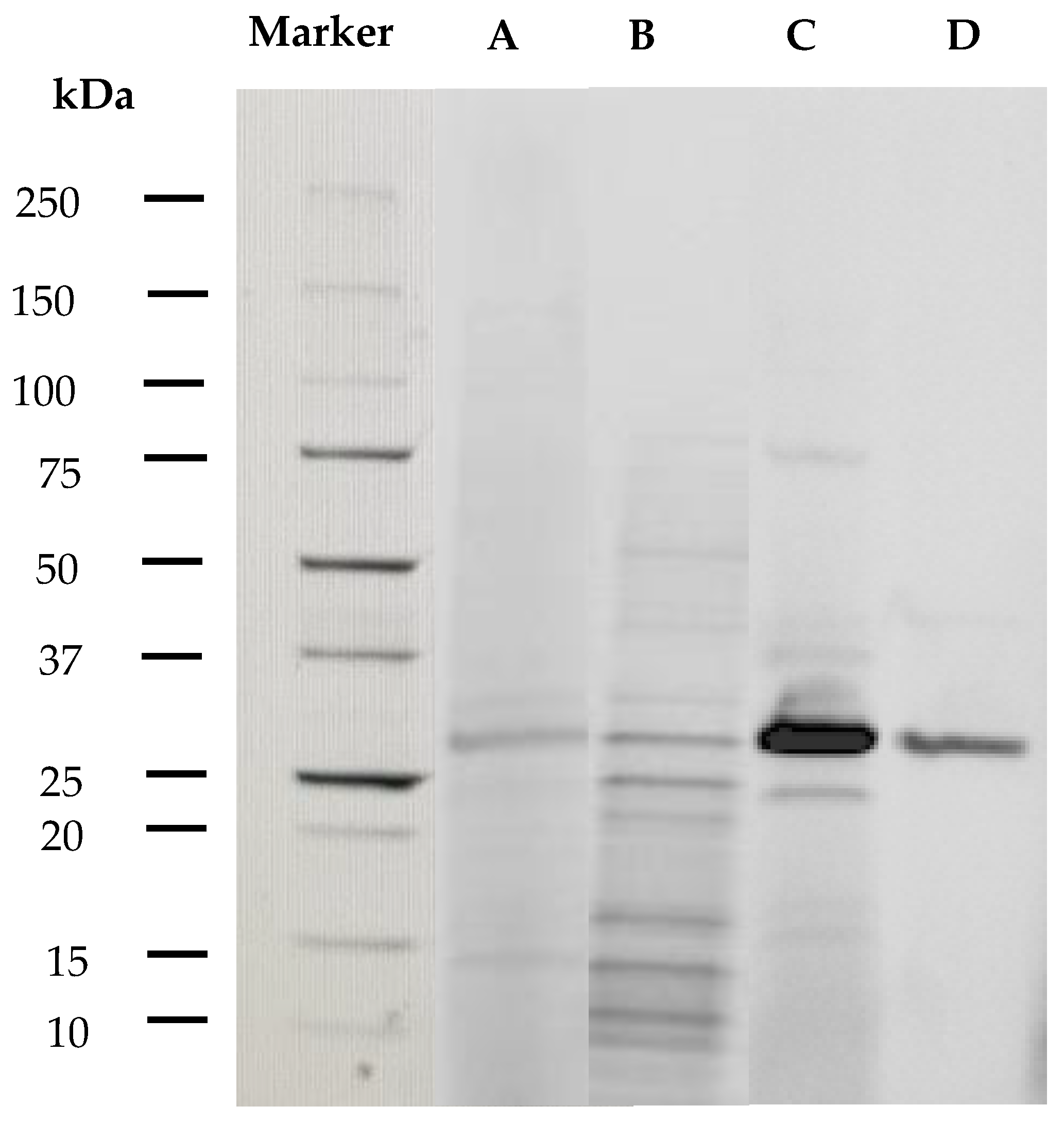
2.2. Purification of AlyDS44
2.3. Characterisation of Enzymes
2.3.1. Molecular Weight of Enzymes
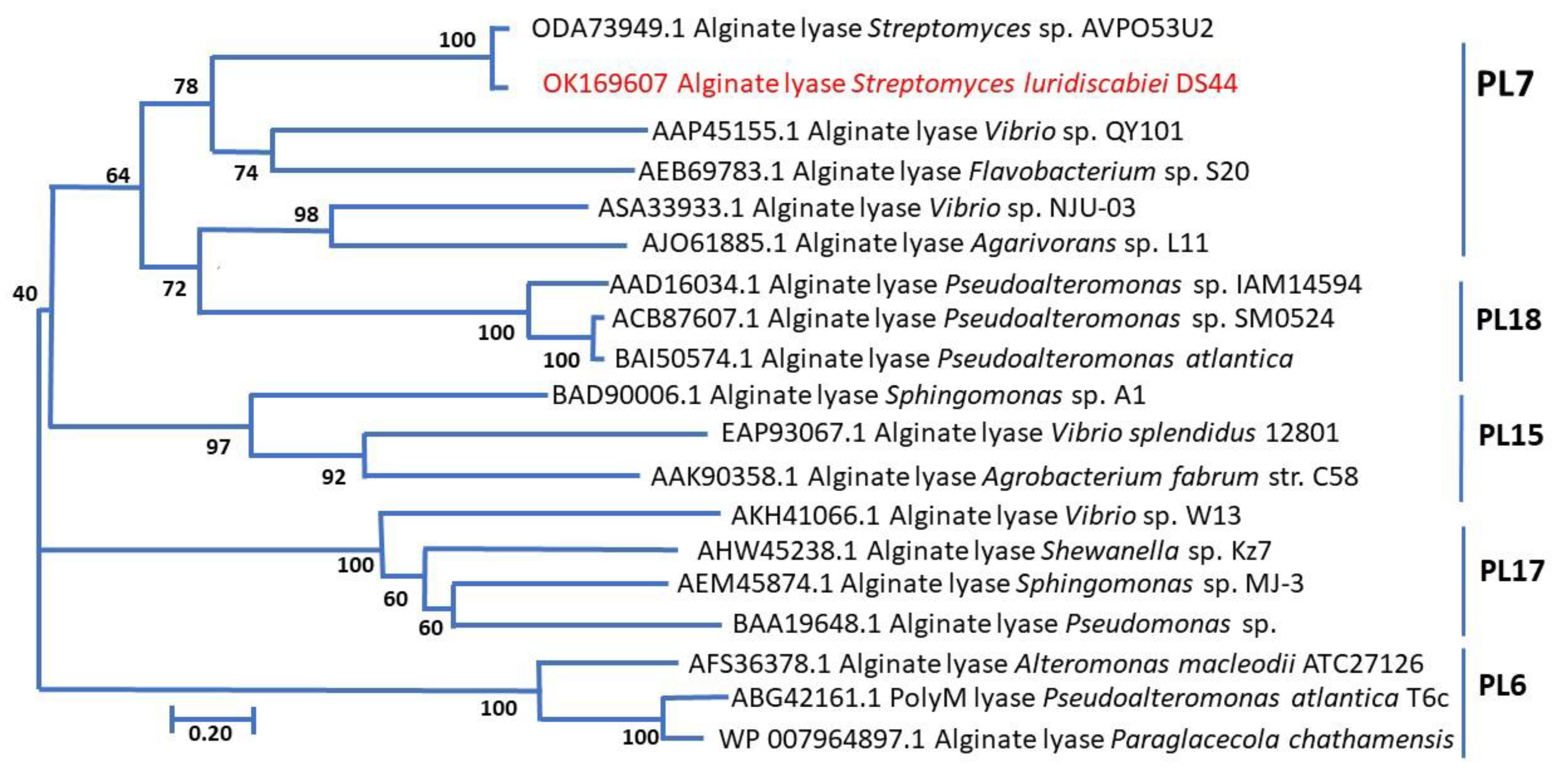
2.3.2. Determination of Amino Acid Sequence of AlyDS44
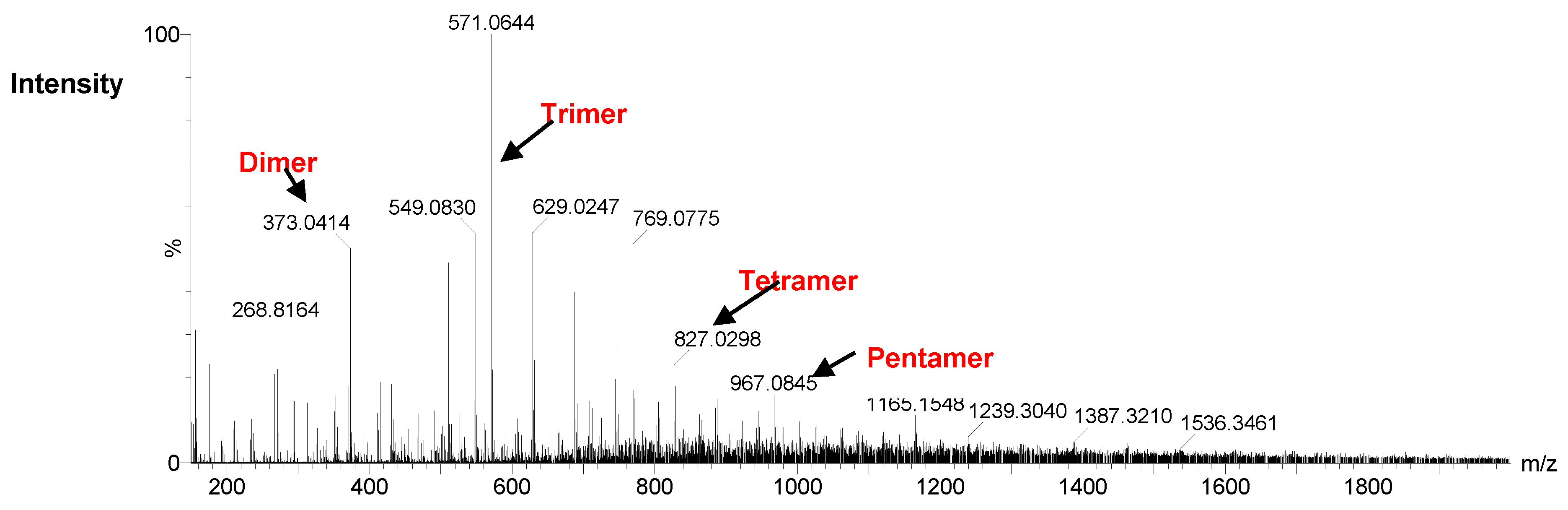
2.4. Analysis of Degradation Products of DS44 Alginate Lyases by ESI-MS
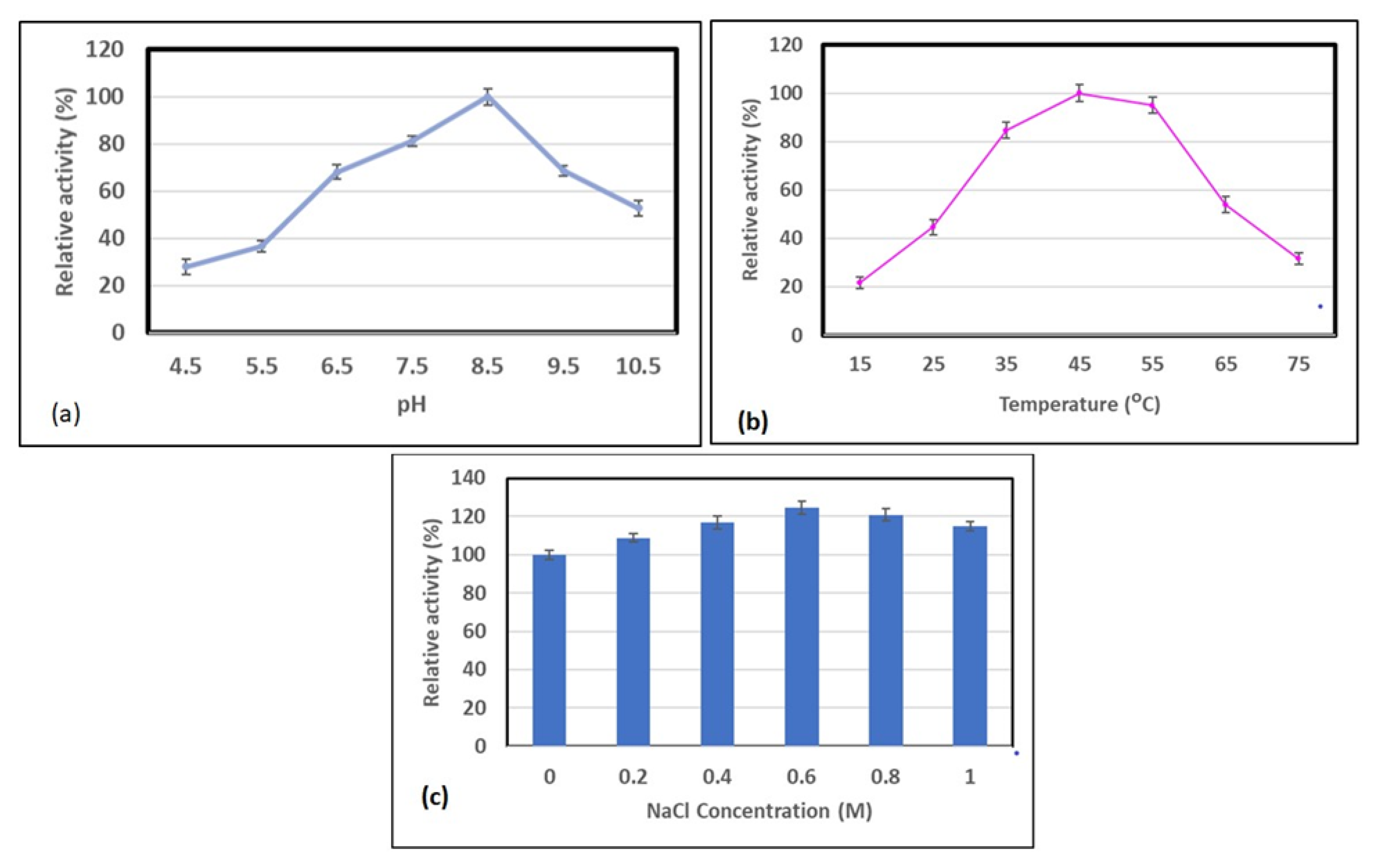
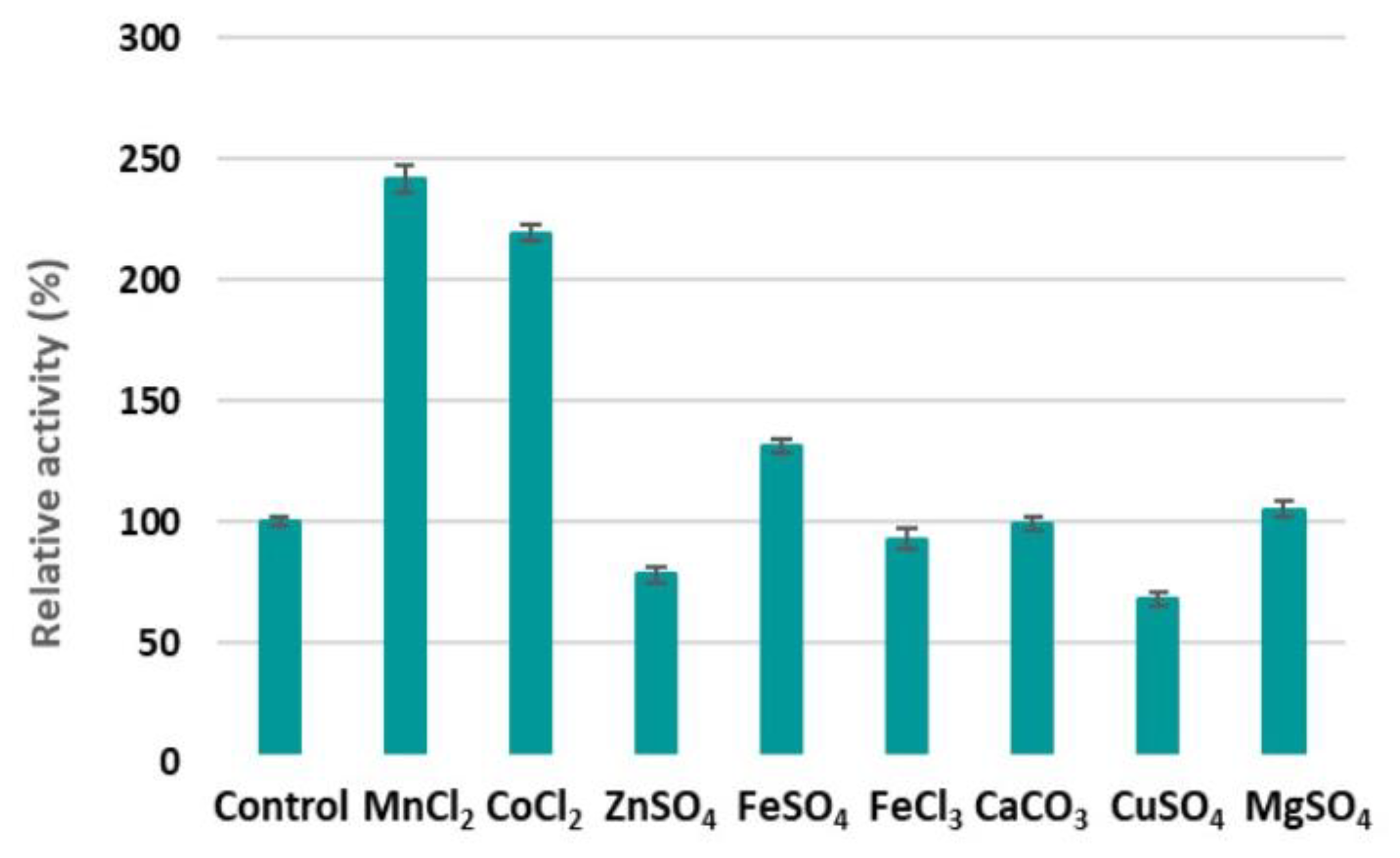
2.5. Optimisation of pH, Temperature, and NaCl, and the Effect of Metal Ions on Alginate Lyase Activity
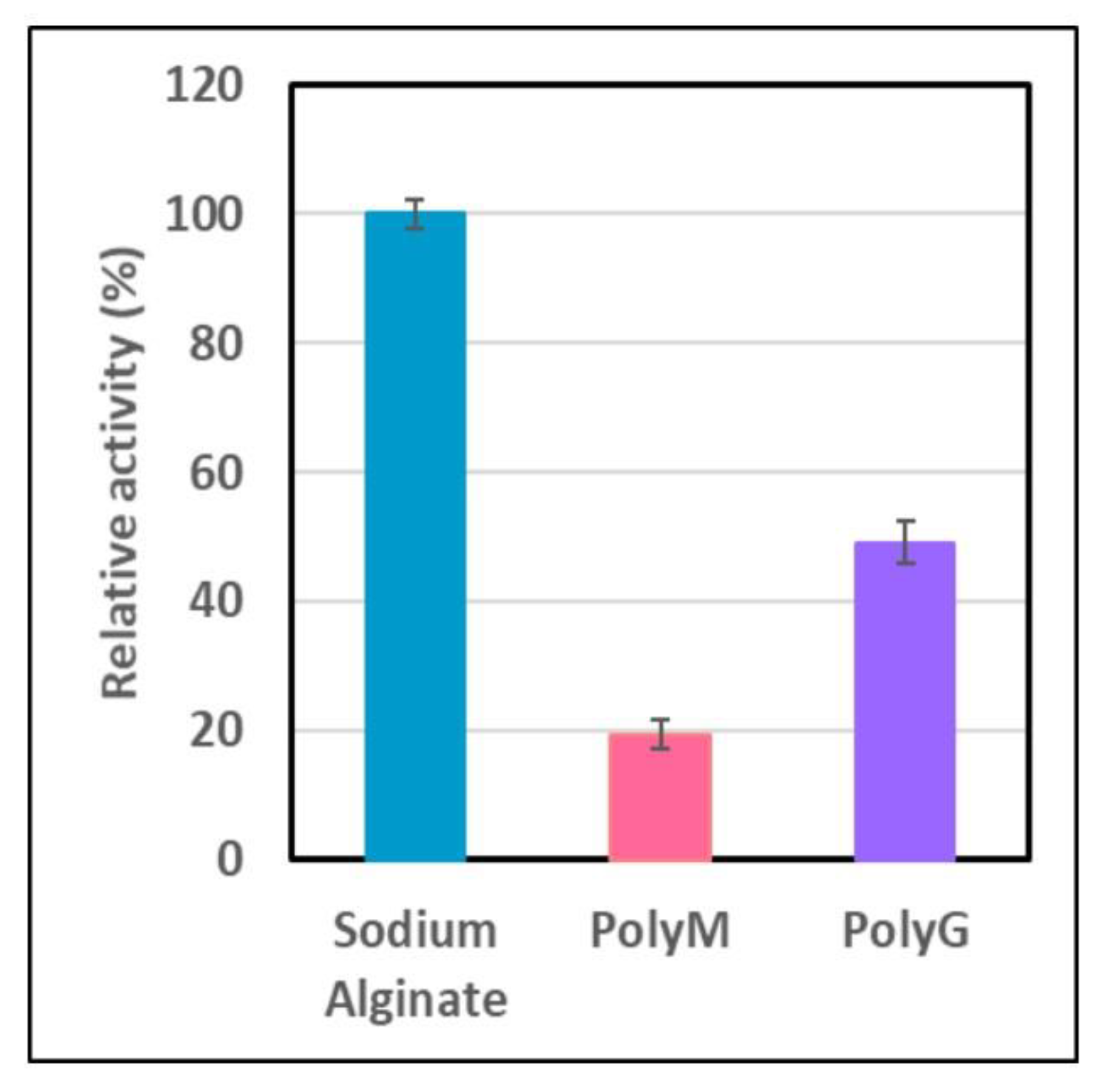
2.6. Hydrolysis of Alginate, and PolyM and PolyG Alginates
3. Materials and Methods
3.1. Materials
3.2. Cultivation and Identification of Actinobacterium DS44
3.3. Purification of Alginate Lyase
3.3.1. Concentration of Crude Enzyme
3.3.2. Ion Exchange Chromatography and Size Exclusion Chromatography
3.4. Alginate Lyase Activity Assay
3.5. Characterisation of Alginate Lyase
3.5.1. SDS-Polyacrylamide Gel Electrophoresis
3.5.2. Amino Acid Sequencing of AlyDS44
3.5.3. Multiple Sequence Alignment
3.6. ESI-MS Analysis of Alginate Lyase Degradation Products
3.7. Effect of Various Factors and Compounds on Alginate Lyase Activity
3.7.1. Determination of the Optimal pH
3.7.2. Determination of the Optimal Temperature
3.7.3. Determination of Salt Tolerance
3.7.4. Influence of Multivalent Metal Ions on Enzyme Activity
3.8. Substrate Specificity
3.9. HPLC Analysis of Alginate Hydrolysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Wong, T.; Preston, L.; Schiller, N. Alginate lyase: Review of major sources and enzyme characteristics, structure-function analysis, biological roles, and applications. Ann. Rev. Microbiol. 2000, 54, 289–340. [Google Scholar] [CrossRef] [PubMed]
- Haug, A.; Larsen, B.; Smidsrod, O. Studies on the sequence of uronic acid residues in alginic acid. Acta Chem. Scand. 1967, 21, 691–704. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andriamanantoanina, H.; Rinaudo, M. Characterization of the alginates from five Madagascan brown algae. Carbohydr. Polym. 2010, 82, 555–560. [Google Scholar] [CrossRef]
- Rioux, L.E.; Turgeon, S.L.; Beaulieu, M. Characterization of polysaccharides extracted from brown seaweeds. Carbohydr. Polym. 2007, 69, 530–537. [Google Scholar] [CrossRef]
- Holdt, S.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Puscaselu, R.G.; Lobiuc, A.; Dimian, M.; Covasa, M. Alginate: From food industry to biomedical applications and management of metabolic disorders. Polymers 2020, 12, 2417. [Google Scholar] [CrossRef]
- Sun, J.; Tan, H. Alginate-based biomaterials for regenerative medicine applications. Materials 2013, 6, 1285–1309. [Google Scholar] [CrossRef]
- Takeshita, S.; Oda, T. Usefulness of alginate lyases derived from marine organisms for the preparation of alginate oligomers with various bioactivities. Adv. Food Nutr. Res. 2016, 79, 137–160. [Google Scholar] [CrossRef]
- Zhu, B.; Yin, H. Alginate lyase: Review of major sources and classification, properties, structure-function analysis and applications. Bioengineered 2015, 6, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.J.; Fathoni, A.; Jeong, G.T.; Jeong, H.D.; Nam, T.J.; Kong, I.S.; Kim, J.K. Microbacterium oxydans, a novel alginate- and laminarin-degrading bacterium for the reutilization of brown-seaweed waste. J. Environ. Manag. 2013, 130, 153–159. [Google Scholar] [CrossRef]
- Schaumann, K.; Weide, G. Enzymatic degradation of alginate by marine fungi. In Thirteenth International Seaweed Symposium; Hydrobiologia, Lindstrom, S.C., Gabrielson, P.W., Eds.; Springer: Dordrecht, The Netherlands, 1990; pp. 589–596. [Google Scholar]
- Zhang, C.; Kim, S. Research and application of marine microbial enzymes: Status and prospects. Mar. Drugs 2010, 8, 1920–1934. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Lee, C.; Lee, E. Alginate lyase: Structure, property, and application. Biotechnol. Bioprocess Eng. 2011, 16, 843–851. [Google Scholar] [CrossRef]
- Kim, H.T.; Ko, H.-J.; Kim, N.; Kim, D.; Lee, D.; Choi, I.-G.; Woo, H.C.; Kim, M.D.; Kim, K.H. Characterization of a recombinant endo-type alginate lyase (Alg7D) from Saccharophagus degradans. Biotechnol. Lett. 2012, 34, 1087–1092. [Google Scholar] [CrossRef]
- Park, H.H.; Kam, N.; Lee, E.Y.; Kim, H.S. Cloning and characterization of a novel oligoalginate lyase from a newly isolated bacterium Sphingomonas sp. MJ-3. Mar. Biotechnol. 2012, 14, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, A.; Yamasaki, M.; Mikami, B.; Hashimoto, W.; Murata, K. Crystal structure of exotype alginate lyase Atu3025 from Agrobacterium tumefaciens. J. Biol. Chem. 2010, 285, 24519–24528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Kumar, P.; Das, P.; Solanki, R.; Kapur, M.K. Potential applications of extracellular enzymes from Streptomyces spp. in various industries. Arch. Microbiol. 2020, 202, 1597–1615. [Google Scholar] [CrossRef]
- Gulve, R.; Deshmukh, A. Antimicrobial activity of the marine actinomycetes. Int. Multidiscip. Res. J. 2012, 2, 16–22. [Google Scholar]
- Bredholt, H.; Fjaervik, E.; Johnsen, G.; Zotchev, S.B. Actinomycetes from sediments in the Trondheim fjord, Norway: Diversity and biological activity. Mar. Drugs 2008, 6, 12–24. [Google Scholar] [CrossRef]
- Ramesh, S.; Mathivanan, N. Screening of marine actinomycetes isolated from the Bay of Bengal, India for antimicrobial activity and industrial enzymes. World J. Microbiol. Biotechnol. 2009, 25, 2103–2111. [Google Scholar] [CrossRef]
- Kumar, V.; Bharti, A.; Negi, Y.K.; Gusain, O.; Pandey, P.; Bisht, G.S. Screening of actinomycetes from earthworm castings for their antimicrobial activity and industrial enzymes. Braz. J. Microbiol. 2012, 43, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Nawani, N.; Aigle, B.; Mandal, A.; Bodas, M.; Ghorbel, S.; Prakash, D. Actinomycetes: Role in biotechnology and medicine. Biomed. Res. Int. 2013, 2013, 687190. [Google Scholar] [CrossRef] [Green Version]
- Holtkamp, A.D.; Kelly, S.; Ulber, R.; Lang, S. Fucoidans and fucoidanases—Focus on techniques for molecular structure elucidation and modification of marine polysaccharides. Appl. Microbiol. Biotechnol. 2008, 82, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zheng, L.; Guo, Z.; Tang, T.; Zhu, B. Alginate degrading enzymes: An updated comprehensive review of the structure, catalytic mechanism, modification method and applications of alginate lyases. Crit. Rev. Biotechnol. 2021, 41, 953–968. [Google Scholar] [CrossRef]
- Cheng, D.; Jiang, C.; Xu, J.; Liu, Z.; Mao, X. Characteristics and applications of alginate lyases: A review. Int. J. Biol. Macromol. 2020, 164, 1304–1320. [Google Scholar] [CrossRef]
- Zhu, Y.; Wu, L.; Chen, Y.; Ni, H.; Xiao, A.; Cai, H. Characterization of an extracellular biofunctional alginate lyase from marine Microbulbifer sp. ALW1 and antioxidant activity of enzymatic hydrolysates. Microbiol. Res. 2016, 182, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Dou, W.; Wei, D.; Li, H.; Li, H.; Rahman, M.M.; Shi, J.; Xu, Z.; Ma, Y. Purification and characterisation of a bifunctional alginate lyase from novel Isoptericola halotolerans CGMCC 5336. Carbohydr. Polym. 2013, 98, 1476–1482. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.E.; Lee, E.Y.; Kim, H.S. Cloning and characterization of alginate lyase from a marine bacterium Streptomyces sp. ALG-5. Mar. Biotechnol. 2009, 11, 10. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Yu, Y.; Yang, S.; Shi, X.; Mou, H.; Li, L. Expression and characterization of a new polyg-specific alginate lyase from marine bacterium Microbulbifer sp. Q7. Front. Microbiol. 2018, 9, 2894. [Google Scholar] [CrossRef]
- Thomas, F.; Lundqvist, L.C.; Jam, M.; Jeudy, A.; Barbeyron, T.; Sandstrom, C.; Michel, G.; Czjzek, M. Comparative characterization of two marine alginate lyases from Zobellia galactanivorans reveals distinct modes of action and exquisite adaptation to their natural substrate. J. Biol. Chem. 2013, 288, 23021–23037. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Chen, P.; Zeng, Y.; Men, Y.; Mu, S.; Zhu, Y.; Chen, Y.; Sun, Y. The characterization and modification of a novel bifunctional and robust alginate lyase derived from Marinimicrobium sp. H1. Mar. Drugs 2019, 17, 545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, F.; Bibb, M.J. Codon usage in the G+C-rich Streptomyces genome. Gene 1992, 113, 55–65. [Google Scholar] [CrossRef]
- Inoue, A. Characterization of PL-7 Family alginate lyases from marine organisms and their applications. Methods Enzymol. 2018, 605, 499–524. [Google Scholar] [CrossRef]
- Yamasaki, M.; Ogura, K.; Hashimoto, W.; Mikami, B.; Murata, K. A structural basis for depolymerization of alginate by polysaccharide lyase family-7. J. Mol. Biol. 2005, 352, 11–21. [Google Scholar] [CrossRef]
- Kawamoto, H.; Horibe, A.; Miki, Y.; Kimura, T.; Tanaka, K.; Nakagawa, T.; Kawamukai, M.; Matsuda, H. Cloning and sequencing analysis of alginate lyase genes from the marine bacterium Vibrio sp. O2. Mar. Biotechnol. 2006, 8, 481–490. [Google Scholar] [CrossRef]
- Uchimura, K.; Miyazaki, M.; Nogi, Y.; Kobayashi, T.; Horikoshi, K. Cloning and sequencing of alginate lyase genes from deep-sea strains of Vibrio and Agarivorans and characterization of a new Vibrio enzyme. Mar. Biotechnol. 2010, 12, 526–533. [Google Scholar] [CrossRef]
- Zhu, B.; Sun, Y.; Ni, F.; Ning, L.; Yao, Z. Characterization of a new endo-type alginate lyase from Vibrio sp. NJU-03. Int. J. Biol. Macromol. 2018, 108, 1140–1147. [Google Scholar] [CrossRef]
- Han, F.; Gong, Q.H.; Song, K.; Li, J.B.; Yu, W.G. Cloning, sequence analysis and expression of gene alyVI encoding alginate lyase from marine bacterium Vibrio sp. QY101. DNA Seq. J DNA Seq. Mapp. 2004, 15, 344–350. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Bi, X.; Ren, Y.; Han, Q.; Zhou, Y.; Han, Y.; Yao, R.; Li, S. Characterization of an alkaline alginate lyase with pH-stable and thermo-tolerance property. Mar. Drugs 2019, 17, 308. [Google Scholar] [CrossRef] [Green Version]
- Park, D.; Jagtap, S.; Nair, S.K. Structure of a PL17 family alginate lyase demonstrates functional similarities among exotype depolymerases. J. Biol. Chem. 2014, 289, 8645–8655. [Google Scholar] [CrossRef] [Green Version]
- Kitamikado, M.; Tseng, C.H.; Yamaguchi, K.; Nakamura, T. Two types of bacterial alginate lyases. Appl. Environ. Microbiol. 1992, 58, 2474–2478. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Ni, F.; Ning, L.; Sun, Y.; Yao, Z. Cloning and characterization of a new pH-stable alginate lyase with high salt tolerance from marine Vibrio sp. NJ-04. Int. J. Biol. Macromol. 2018, 115, 1063–1070. [Google Scholar] [CrossRef]
- Shin, J.W.; Choi, S.H.; Kim, D.E.; Kim, H.S.; Lee, J.H.; Lee, I.S.; Lee, E.Y. Heterologous expression of an alginate lyase from Streptomyces sp. ALG-5 in Escherichia coli and its use for preparation of the magnetic nanoparticle-immobilized enzymes. Bioprocess. Biosyst. Eng. 2011, 34, 113–119. [Google Scholar] [CrossRef]
- Kim, H.S. Characterization of recombinant polyg-specific lyase from a marine bacterium, Streptomyces sp. M3. J. Life Sci. 2010, 20, 1582–1588. [Google Scholar] [CrossRef] [Green Version]
- Hu, F.; Li, Q.; Zhu, B.; Ni, F.; Sun, Y.; Yao, Z. Effects of module truncation on biochemical characteristics and products distribution of a new alginate lyase with two catalytic modules. Glycobiology 2019, 29, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Kao, S.S.; Bassiouni, A.; Ramezanpour, M.; Finnie, J.; Chegeni, N.; Colella, A.D.; Chataway, T.K.; Wormald, P.J.; Vreugde, S.; Psaltis, A.J. Proteomic analysis of nasal mucus samples of healthy patients and patients with chronic rhinosinusitis. J. Allergy Clin. Immunol. 2021, 147, 168–178. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Steps | Volume (mL) | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) | Yield (%) | Purification (Fold) |
|---|---|---|---|---|---|---|
| Culture broth | 300 | 604.5 | 695.9 | 1.15 | 100 | 1 |
| 60% Ammonium sulfate precipitate (after dialysis) | 12 | 13.52 | 370.8 | 27.54 | 53.3 | 23.9 |
| Mono-Q Active fractions | 1 | 0.327 | 28.88 | 93.78 | 4.15 | 81.5 |
| Superdex 75 Purified product | 1 | 0.115 | 12.49 | 108.6 | 1.79 | 94.4 |
| Peak 1 | Peak 2 | Peak 3 | ||||
|---|---|---|---|---|---|---|
| % | MW (kDa) | % | MW (kDa) | % | MW (kDa) | |
| Sodium alginate | ||||||
| Control | 44.3 | 577.8 | 43.5 | 305.0 | 0.2 | 2.1 |
| 1 h | 10.2 | 350.6 | 60.7 | 64.9 | 21.8 | 15.5 |
| 24 h | 15.2 | 18.7 | 15.3 | 6.6 | 35.0 | 3.4 |
| Poly M | ||||||
| Control | 79.9 | 17.7 | 2.9 | 2.2 | 1.7 | 1.0 |
| 1 h | 65.5 | 15.8 | 3.7 | 2.2 | 1.6 | 1.0 |
| 24 h | 54.7 | 9.8 | 8.3 | 4.1 | 5.8 | 2.2 |
| Poly G | ||||||
| Control | 80.9 | 14.4 | 0.4 | 2.1 | 16.7 | 0.6 |
| 1 h | 65.8 | 10.9 | 2.5 | 2.1 | 31.9 | 0.5 |
| 24 h | 51.0 | 3.1 | 14.3 | 2.1 | 34.5 | 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, T.N.T.; Chataway, T.; Araujo, R.; Puri, M.; Franco, C.M.M. Purification and Characterization of a Novel Alginate Lyase from a Marine Streptomyces Species Isolated from Seaweed. Mar. Drugs 2021, 19, 590. https://0-doi-org.brum.beds.ac.uk/10.3390/md19110590
Nguyen TNT, Chataway T, Araujo R, Puri M, Franco CMM. Purification and Characterization of a Novel Alginate Lyase from a Marine Streptomyces Species Isolated from Seaweed. Marine Drugs. 2021; 19(11):590. https://0-doi-org.brum.beds.ac.uk/10.3390/md19110590
Chicago/Turabian StyleNguyen, Thi Nhu Thuong, Timothy Chataway, Ricardo Araujo, Munish Puri, and Christopher Milton Mathew Franco. 2021. "Purification and Characterization of a Novel Alginate Lyase from a Marine Streptomyces Species Isolated from Seaweed" Marine Drugs 19, no. 11: 590. https://0-doi-org.brum.beds.ac.uk/10.3390/md19110590