
The Role of Chemokines in Cervical Cancers

 and
and
Abstract
:1. Introduction
2. Material and Methods
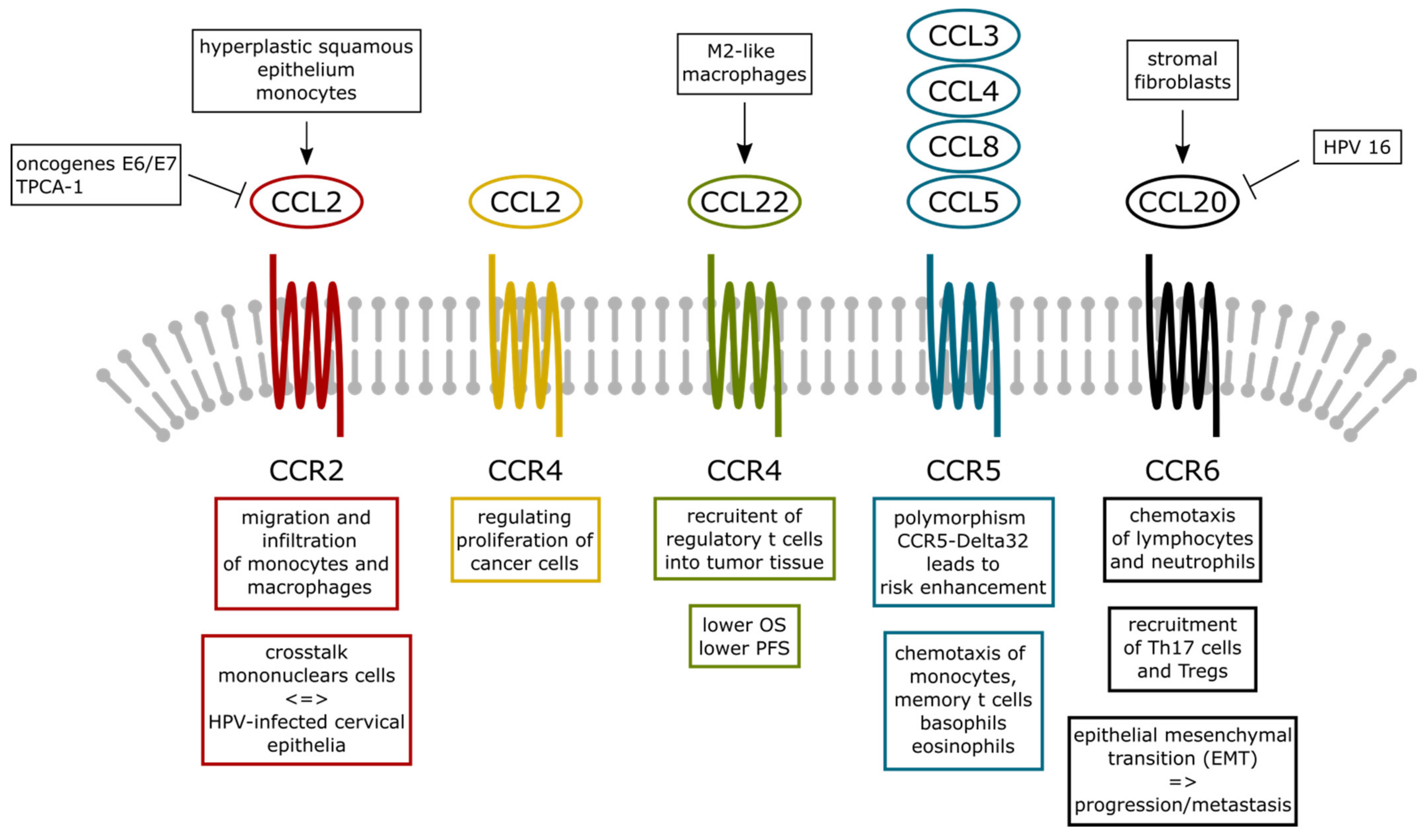
3. CC Chemokine Ligands (CCL) in Cervical Cancer Biology
3.1. CCL2
3.2. CCL3, CCL4, CCL5, and CCL8
3.3. CCL17 and CCL19
3.4. CCL20 and CCL22
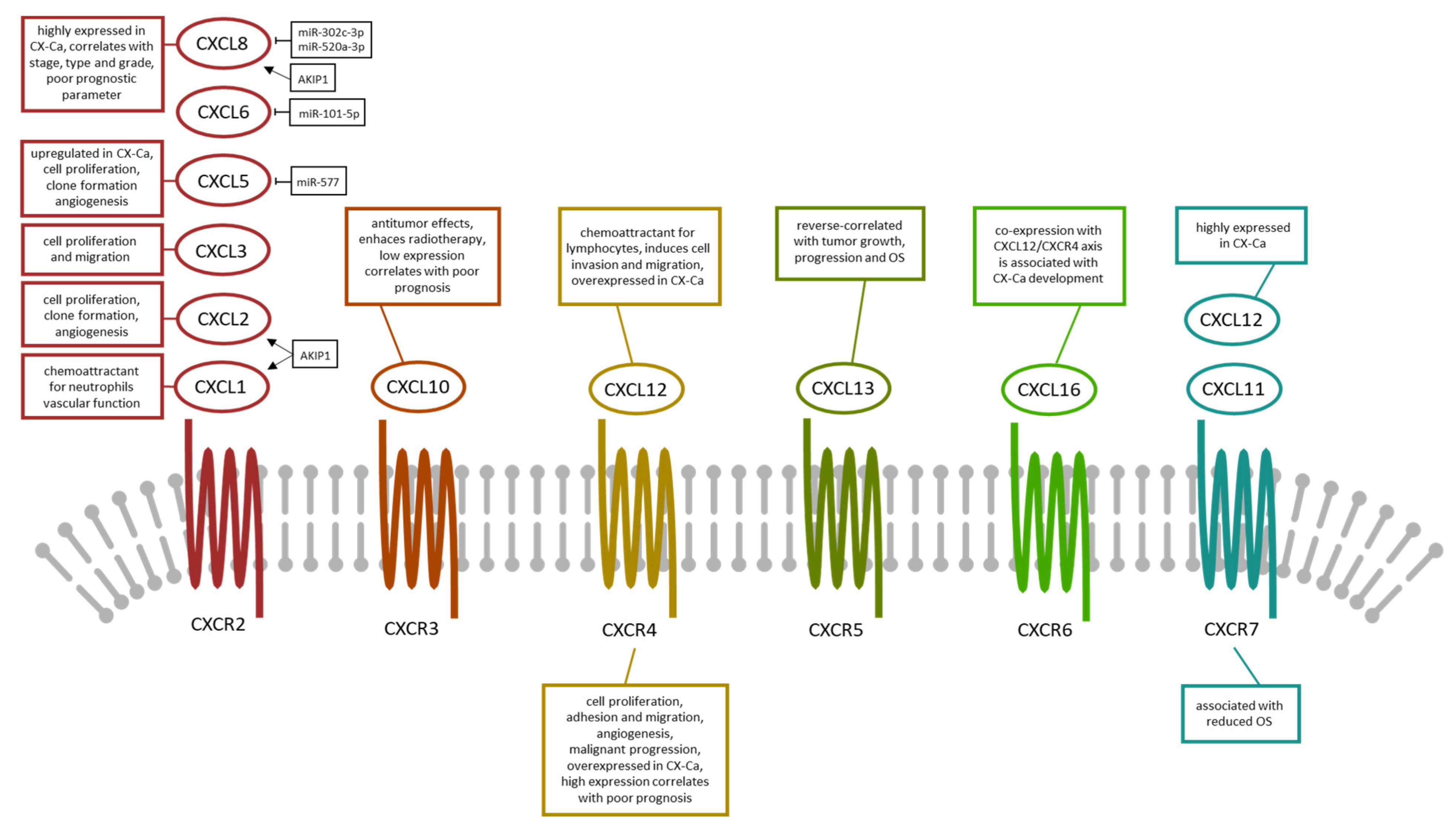
4. CXC Chemokines and Its Receptors in Cervical Cancer
4.1. CXCL1, CXCL2, and CXCL3
4.2. CXCL5, CXCL6, CXCL8, and CXCL10
4.3. CXCL11 and CXCL12
4.4. CXCL13 and CXCL16
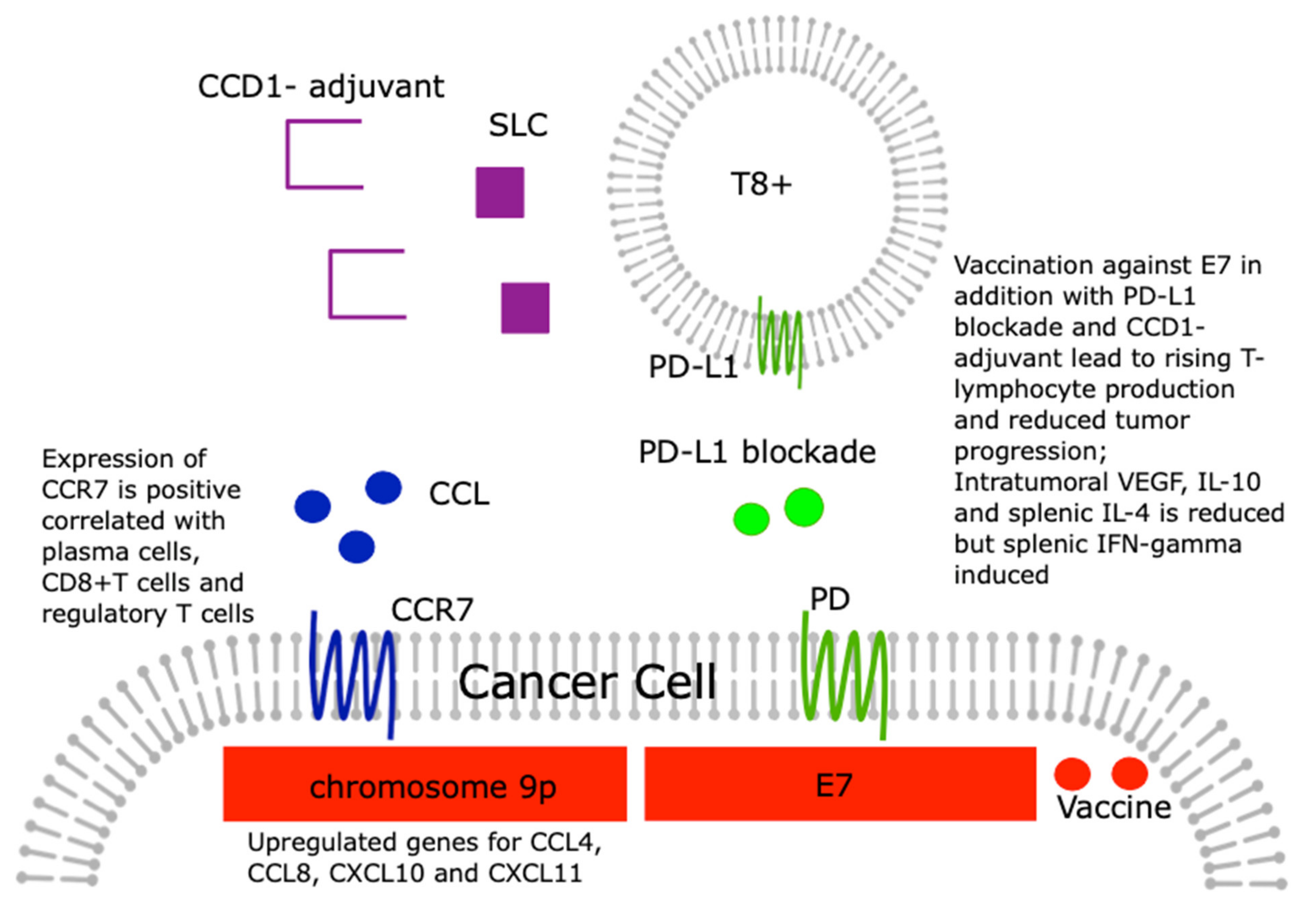
5. The Role of Chemokines in Checkpoint Activation/Inhibition in Cervical Cancer
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Den Boon, J.A.; Pyeon, D.; Wang, S.S.; Horswill, M.; Schiffman, M.; Sherman, M.; Zuna, R.E.; Wang, Z.; Hewitt, S.M.; Pearson, R.; et al. Molecular transitions from papillomavirus infection to cervical precancer and cancer: Role of stromal estrogen receptor signaling. Proc. Natl. Acad. Sci. USA 2015, 112, E3255–E3264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, M.; Li, T.; Niu, M.; Luo, S.; Chu, Q.; Wu, K. Epidemiological trends of women’s cancers from 1990 to 2019 at the global, regional, and national levels: A population-based study. Biomark. Res. 2021, 9, 55. [Google Scholar] [CrossRef] [PubMed]
- Young, R.H.; Clement, P.B. Endocervical adenocarcinoma and its variants: Their morphology and differential diagnosis. Histopathology 2002, 41, 185–207. [Google Scholar] [CrossRef] [PubMed]
- Schiffman, M.; Wentzensen, N.; Wacholder, S.; Kinney, W.; Gage, J.C.; Castle, P.E. Human papillomavirus testing in the prevention of cervical cancer. J. Natl. Cancer Inst. 2011, 103, 368–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Schmoeckel, E.; Kost, B.P.; Kuhn, C.; Vattai, A.; Vilsmaier, T.; Mahner, S.; Mayr, D.; Jeschke, U.; Heidegger, H.H. Higher CCL22+ Cell Infiltration is Associated with Poor Prognosis in Cervical Cancer Patients. Cancers 2019, 11, 2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freier, C.P.; Kuhn, C.; Endres, S.; Mayr, D.; Friese, K.; Jeschke, U.; Anz, D. FOXP3+ Cells Recruited by CCL22 into Breast Cancer Correlates with Less Tumor Nodal Infiltration. Anticancer Res. 2016, 36, 3139–3145. [Google Scholar] [PubMed]
- Freier, C.P.; Kuhn, C.; Rapp, M.; Endres, S.; Mayr, D.; Friese, K.; Anz, D.; Jeschke, U. Expression of CCL22 and Infiltration by Regulatory T Cells are Increased in the Decidua of Human Miscarriage Placentas. Am. J. Reprod. Immunol. 2015, 74, 216–227. [Google Scholar] [CrossRef]
- Liu, L.B.; Xie, F.; Chang, K.K.; Shang, W.Q.; Meng, Y.H.; Yu, J.J.; Li, H.; Sun, Q.; Yuan, M.M.; Jin, L.P.; et al. Chemokine CCL17 induced by hypoxia promotes the proliferation of cervical cancer cell. Am. J. Cancer Res. 2015, 5, 3072–3084. [Google Scholar] [PubMed]
- Ren, H.; Ren, G.; Sun, L.; Fan, X.; Wang, Y.; Li, X. Production and mechanism of CCL5 by macrophages in U14 cervical cancer-bearing mice during infection. Zhonghua Fu Chan Ke Za Zhi 2015, 50, 367–373. [Google Scholar] [PubMed]
- Walch-Ruckheim, B.; Mavrova, R.; Henning, M.; Vicinus, B.; Kim, Y.J.; Bohle, R.M.; Juhasz-Boss, I.; Solomayer, E.F.; Smola, S. Stromal Fibroblasts Induce CCL20 through IL6/C/EBPbeta to Support the Recruitment of Th17 Cells during Cervical Cancer Progression. Cancer Res. 2015, 75, 5248–5259. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Lou, X.M.; He, Y. Preferential recruitment of Th17 cells to cervical cancer via CCR6-CCL20 pathway. PloS ONE 2015, 10, e0120855. [Google Scholar] [CrossRef]
- Chang, Y.W.; Chen, M.W.; Chiu, C.F.; Hong, C.C.; Cheng, C.C.; Hsiao, M.; Chen, C.A.; Wei, L.H.; Su, J.L. Arsenic trioxide inhibits CXCR4-mediated metastasis by interfering miR-520h/PP2A/NF-kappaB signaling in cervical cancer. Ann. Surg. Oncol. 2014, 21 (Suppl. 4), S687–S695. [Google Scholar] [CrossRef]
- Kim, B.; Park, B. Baohuoside I suppresses invasion of cervical and breast cancer cells through the downregulation of CXCR4 chemokine receptor expression. Biochemistry 2014, 53, 7562–7569. [Google Scholar] [CrossRef]
- Yadav, S.S.; Prasad, S.B.; Das, M.; Kumari, S.; Pandey, L.K.; Singh, S.; Pradhan, S.; Narayan, G. Epigenetic silencing of CXCR4 promotes loss of cell adhesion in cervical cancer. BioMed Res. Int. 2014, 2014, 581403. [Google Scholar] [CrossRef]
- Yadav, S.S.; Prasad, S.B.; Prasad, C.B.; Pandey, L.K.; Pradhan, S.; Singh, S.; Narayan, G. CXCL12 is a key regulator in tumor microenvironment of cervical cancer: An In Vitro study. Clin. Exp. Metastasis 2016, 33, 431–439. [Google Scholar] [CrossRef]
- Yin, G.; Zhu, T.; Li, J.; Wu, A.; Liang, J.; Zhi, Y. CXCL12 rs266085 and TNF-alpha rs1799724 polymorphisms and susceptibility to cervical cancer in a Chinese population. Int. J. Clin. Exp. Pathol. 2015, 8, 5768–5774. [Google Scholar]
- Zhao, M.; Ma, Q.; Xu, J.; Fu, S.; Chen, L.; Wang, B.; Wu, J.; Yang, L. Combining CXCL10 gene therapy and radiotherapy improved therapeutic efficacy in cervical cancer HeLa cell xenograft tumor models. Oncol. Lett. 2015, 10, 768–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreyer, T.F.; Kuhn, S.; Stange, C.; Heithorst, N.; Schilling, D.; Jelsma, J.; Sievert, W.; Seitz, S.; Stangl, S.; Hapfelmeier, A.; et al. The Chemokine CX3CL1 Improves Trastuzumab Efficacy in HER2 Low-Expressing Cancer In Vitro and In Vivo. Cancer Immunol. Res. 2021, 9, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Gaudin, F.; Nasreddine, S.; Donnadieu, A.C.; Emilie, D.; Combadiere, C.; Prevot, S.; Machelon, V.; Balabanian, K. Identification of the chemokine CX3CL1 as a new regulator of malignant cell proliferation in epithelial ovarian cancer. PloS ONE 2011, 6, e21546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Yi, L.; Liu, P.; Jiang, L.; Wang, H.; Hu, A.; Sun, C.; Dong, J. CX3CL1 involves in breast cancer metastasizing to the spine via the Src/FAK signaling pathway. J. Cancer 2018, 9, 3603–3612. [Google Scholar] [CrossRef]
- Liu, P.; Liang, Y.; Jiang, L.; Wang, H.; Wang, S.; Dong, J. CX3CL1/fractalkine enhances prostate cancer spinal metastasis by activating the Src/FAK pathway. Int. J. Oncol. 2018, 53, 1544–1556. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Liang, Y.; Chan, Q.; Jiang, L.; Dong, J. CX3CL1 promotes lung cancer cell migration and invasion via the Src/focal adhesion kinase signaling pathway. Oncol. Rep. 2019, 41, 1911–1917. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, H.; Dong, T.; Yan, Y.; Sun, L.; Wang, W. Clinical significance of expression level of CX3CL1-CX3CR1 axis in bone metastasis of lung cancer. Clin. Transl. Oncol. Off. Publ. Fed. Span. Oncol. Soc. Natl. Cancer Inst. Mex. 2021, 23, 378–388. [Google Scholar] [CrossRef]
- Rosl, F.; Lengert, M.; Albrecht, J.; Kleine, K.; Zawatzky, R.; Schraven, B.; zur Hausen, H. Differential regulation of the JE gene encoding the monocyte chemoattractant protein (MCP-1) in cervical carcinoma cells and derived hybrids. J. Virol. 1994, 68, 2142–2150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riethdorf, L.; Riethdorf, S.; Gutzlaff, K.; Prall, F.; Loning, T. Differential expression of the monocyte chemoattractant protein-1 gene in human papillomavirus-16-infected squamous intraepithelial lesions and squamous cell carcinomas of the cervix uteri. Am. J. Pathol. 1996, 149, 1469–1476. [Google Scholar]
- Kleine-Lowinski, K.; Gillitzer, R.; Kuhne-Heid, R.; Rosl, F. Monocyte-chemo-attractant-protein-1 (MCP-1)-gene expression in cervical intra-epithelial neoplasias and cervical carcinomas. Int. J. Cancer 1999, 82, 6–11. [Google Scholar] [CrossRef]
- Stiasny, A.; Kuhn, C.; Mayr, D.; Alexiou, C.; Janko, C.; Wiest, I.; Jeschke, U.; Kost, B. Immunohistochemical Evaluation of E6/E7 HPV Oncoproteins Staining in Cervical Cancer. Anticancer Res. 2016, 36, 3195–3198. [Google Scholar]
- Stiasny, A.; Freier, C.P.; Kuhn, C.; Schulze, S.; Mayr, D.; Alexiou, C.; Janko, C.; Wiest, I.; Dannecker, C.; Jeschke, U.; et al. The involvement of E6, p53, p16, MDM2 and Gal-3 in the clinical outcome of patients with cervical cancer. Oncol. Lett. 2017, 14, 4467–4476. [Google Scholar] [CrossRef] [Green Version]
- Kleine-Lowinski, K.; Rheinwald, J.G.; Fichorova, R.N.; Anderson, D.J.; Basile, J.; Munger, K.; Daly, C.M.; Rosl, F.; Rollins, B.J. Selective suppression of monocyte chemoattractant protein-1 expression by human papillomavirus E6 and E7 oncoproteins in human cervical epithelial and epidermal cells. Int. J. Cancer 2003, 107, 407–415. [Google Scholar] [CrossRef]
- Nakamura, M.; Kyo, S.; Kanaya, T.; Yatabe, N.; Maida, Y.; Tanaka, M.; Ishida, Y.; Fujii, C.; Kondo, T.; Inoue, M.; et al. hTERT-promoter-based tumor-specific expression of MCP-1 effectively sensitizes cervical cancer cells to a low dose of cisplatin. Cancer Gene Ther. 2004, 11, 1–7. [Google Scholar] [CrossRef]
- Zijlmans, H.J.; Fleuren, G.J.; Baelde, H.J.; Eilers, P.H.; Kenter, G.G.; Gorter, A. The absence of CCL2 expression in cervical carcinoma is associated with increased survival and loss of heterozygosity at 17q11.2. J. Pathol. 2006, 208, 507–517. [Google Scholar] [CrossRef]
- Hou, T.; Liang, D.; Xu, L.; Huang, X.; Huang, Y.; Zhang, Y. Atypical chemokine receptors predict lymph node metastasis and prognosis in patients with cervical squamous cell cancer. Gynecol. Oncol. 2013, 130, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Li, S.; Lin, L.; Jiang, Y.; Wan, Y.; Zhou, S.; Cheng, W. Co-expression network analysis identified atypical chemokine receptor 1 (ACKR1) association with lymph node metastasis and prognosis in cervical cancer. Cancer Biomark. Sect. A Dis. Markers 2020, 27, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Pahne-Zeppenfeld, J.; Schroer, N.; Walch-Ruckheim, B.; Oldak, M.; Gorter, A.; Hegde, S.; Smola, S. Cervical cancer cell-derived interleukin-6 impairs CCR7-dependent migration of MMP-9-expressing dendritic cells. Int. J. Cancer 2014, 134, 2061–2073. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Liu, S.; Xiong, J.; Yang, H.; Wu, Y.; Xu, A.; Gong, Y. IL-1beta promotes cervical cancer through activating NF-kappaB/CCL-2. Int. J. Clin. Exp. Pathol. 2021, 14, 426–433. [Google Scholar] [PubMed]
- Coelho, A.; Matos, A.; Catarino, R.; Pinto, D.; Pereira, D.; Lopes, C.; Medeiros, R. Protective role of the polymorphism CCR2-64I in the progression from squamous intraepithelial lesions to invasive cervical carcinoma. Gynecol. Oncol. 2005, 96, 760–764. [Google Scholar] [CrossRef]
- Coelho, A.; Matos, A.; Catarino, R.; Pinto, D.; Sousa, H.; Pereira, D.; Medeiros, R. The influence of chemokine receptor CCR2 genotypes in the route to cervical carcinogenesis. Gynecol. Obstet. Investig. 2007, 64, 208–212. [Google Scholar] [CrossRef]
- Ivansson, E.L.; Gustavsson, I.M.; Magnusson, J.J.; Steiner, L.L.; Magnusson, P.K.; Erlich, H.A.; Gyllensten, U.B. Variants of chemokine receptor 2 and interleukin 4 receptor, but not interleukin 10 or Fas ligand, increase risk of cervical cancer. Int. J. Cancer 2007, 121, 2451–2457. [Google Scholar] [CrossRef]
- Niwa, Y.; Akamatsu, H.; Niwa, H.; Sumi, H.; Ozaki, Y.; Abe, A. Correlation of tissue and plasma RANTES levels with disease course in patients with breast or cervical cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2001, 7, 285–289. [Google Scholar]
- Singh, H.; Sachan, R.; Jain, M.; Mittal, B. CCR5-Delta32 polymorphism and susceptibility to cervical cancer: Association with early stage of cervical cancer. Oncol. Res. 2008, 17, 87–91. [Google Scholar] [CrossRef]
- Chen, X.J.; Deng, Y.R.; Wang, Z.C.; Wei, W.F.; Zhou, C.F.; Zhang, Y.M.; Yan, R.M.; Liang, L.J.; Zhong, M.; Liang, L.; et al. Hypoxia-induced ZEB1 promotes cervical cancer progression via CCL8-dependent tumour-associated macrophage recruitment. Cell Death Dis. 2019, 10, 508. [Google Scholar] [CrossRef] [Green Version]
- Zehbe, I.; Kaufmann, A.M.; Schmidt, M.; Hohn, H.; Maeurer, M.J. Human papillomavirus 16 E6-specific CD45RA+ CCR7+ high avidity CD8+ T cells fail to control tumor growth despite interferon-gamma production in patients with cervical cancer. J. Immunother. 2007, 30, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Caberg, J.H.; Hubert, P.; Herman, L.; Herfs, M.; Roncarati, P.; Boniver, J.; Delvenne, P. Increased migration of Langerhans cells in response to HPV16 E6 and E7 oncogene silencing: Role of CCL20. Cancer Immunol. Immunother. 2009, 58, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Yao, T.; Lin, Z. MiR-21 is involved in cervical squamous cell tumorigenesis and regulates CCL20. Biochim. Biophys. Acta 2012, 1822, 248–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhu, D.; Lv, Q.; Yi, Y.; Li, F.; Zhang, W. The key role of astrocyte elevated gene-1 in CCR6-induced EMT in cervical cancer. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2015, 36, 9763–9767. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Xue, M. Correlation of E6 and E7 levels in high-risk HPV16 type cervical lesions with CCL20 and Langerhans cells. Genet. Mol. Res. 2015, 14, 10473–10481. [Google Scholar] [CrossRef]
- Yoshida, R.; Imai, T.; Hieshima, K.; Kusuda, J.; Baba, M.; Kitaura, M.; Nishimura, M.; Kakizaki, M.; Nomiyama, H.; Yoshie, O. Molecular cloning of a novel human CC chemokine EBI1-ligand chemokine that is a specific functional ligand for EBI1, CCR7. J. Biol. Chem. 1997, 272, 13803–13809. [Google Scholar] [CrossRef] [Green Version]
- Yung, M.M.; Tang, H.W.; Cai, P.C.; Leung, T.H.; Ngu, S.F.; Chan, K.K.; Xu, D.; Yang, H.; Ngan, H.Y.; Chan, D.W. GRO-alpha and IL-8 enhance ovarian cancer metastatic potential via the CXCR2-mediated TAK1/NFkappaB signaling cascade. Theranostics 2018, 8, 1270–1285. [Google Scholar] [CrossRef] [PubMed]
- Sales, K.J.; Sutherland, J.R.; Jabbour, H.N.; Katz, A.A. Seminal plasma induces angiogenic chemokine expression in cervical cancer cells and regulates vascular function. Biochim. Biophys. Acta 2012, 1823, 1789–1795. [Google Scholar] [CrossRef] [Green Version]
- Gavamukulya, Y.; Maina, E.N.; El-Shemy, H.A.; Meroka, A.M.; Kangogo, G.K.; Magoma, G.; Wamunyokoli, F. Annona muricata silver nanoparticles exhibit strong anticancer activities against cervical and prostate adenocarcinomas through regulation of CASP9 and the CXCL1/CXCR2 genes axis. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2021, 43, 37–55. [Google Scholar] [CrossRef]
- Zhang, W.; Wu, Q.; Wang, C.; Yang, L.; Liu, P.; Ma, C. AKIP1 promotes angiogenesis and tumor growth by upregulating CXC-chemokines in cervical cancer cells. Mol. Cell. Biochem. 2018, 448, 311–320. [Google Scholar] [CrossRef]
- Qi, Y.L.; Li, Y.; Man, X.X.; Sui, H.Y.; Zhao, X.L.; Zhang, P.X.; Qu, X.S.; Zhang, H.; Wang, B.X.; Li, J.; et al. CXCL3 overexpression promotes the tumorigenic potential of uterine cervical cancer cells via the MAPK/ERK pathway. J. Cell Physiol. 2020, 235, 4756–4765. [Google Scholar] [CrossRef]
- Feng, X.; Zhang, D.; Li, X.; Ma, S.; Zhang, C.; Wang, J.; Li, Y.; Liang, L.; Zhang, P.; Qu, Y.; et al. CXCL5, the upregulated chemokine in patients with uterine cervix cancer, in vivo and in vitro contributes to oncogenic potential of Hela uterine cervix cancer cells. Biomed. Pharmacother. 2018, 107, 1496–1504. [Google Scholar] [CrossRef]
- Bai, L.; Yao, N.; Qiao, G.; Wu, L.; Ma, X. CXCL5 contributes to the tumorigenicity of cervical cancer and is post-transcriptionally regulated by miR-577. Int. J. Clin. Exp. Pathol. 2020, 13, 2984–2993. [Google Scholar]
- Shen, W.; Xie, X.Y.; Liu, M.R.; Wang, L.L. MicroRNA-101-5p inhibits the growth and metastasis of cervical cancer cell by inhibiting CXCL6. Eur. Rev. Med Pharmacol. Sci. 2019, 23, 1957–1968. [Google Scholar] [CrossRef]
- Yan, R.; Shuai, H.; Luo, X.; Wang, X.; Guan, B. The clinical and prognostic value of CXCL8 in cervical carcinoma patients: Immunohistochemical analysis. Biosci. Rep. 2017, 37, BSR20171021. [Google Scholar] [CrossRef] [Green Version]
- Ding, H.M.; Zhang, H.; Wang, J.; Zhou, J.H.; Shen, F.R.; Ji, R.N.; Shi, J.Y.; Chen, Y.G. miR302c3p and miR520a3p suppress the proliferation of cervical carcinoma cells by targeting CXCL8. Mol. Med. Rep. 2021, 23, 322. [Google Scholar] [CrossRef]
- Sato, E.; Fujimoto, J.; Toyoki, H.; Sakaguchi, H.; Alam, S.M.; Jahan, I.; Tamaya, T. Expression of IP-10 related to angiogenesis in uterine cervical cancers. Br. J. Cancer 2007, 96, 1735–1739. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.L.; Chen, P.; Luo, S.; Li, J.; Liu, K.; Hu, H.Z.; Wei, Y.Q. CXC-chemokine-ligand-10 gene therapy efficiently inhibits the growth of cervical carcinoma on the basis of its anti-angiogenic and antiviral activity. Biotechnol. Appl. Biochem. 2009, 53, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.L.; Wang, B.Q.; Chen, L.L.; Luo, H.Q.; Wu, J.B. CXCL10 enhances radiotherapy effects in HeLa cells through cell cycle redistribution. Oncol. Lett. 2012, 3, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, L.; Teng, X.; Liu, Z.; Liu, C.; Zhang, L.; Liu, Z. The chemokine receptor CXCR7 is a critical regulator for the tumorigenesis and development of papillary thyroid carcinoma by inducing angiogenesis In Vitro and In Vivo. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 2415–2423. [Google Scholar] [CrossRef]
- Schrevel, M.; Karim, R.; ter Haar, N.T.; van der Burg, S.H.; Trimbos, J.B.; Fleuren, G.J.; Gorter, A.; Jordanova, E.S. CXCR7 expression is associated with disease-free and disease-specific survival in cervical cancer patients. Br. J. Cancer 2012, 106, 1520–1525. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Qin, W.; Zhao, C.; Long, J.; Li, M. CXCR7, a Prognostic Biomarker in Cervical Squamous Cell Carcinoma, May Be a Screening Index for Treatment Options at Stages IB1 and IIA1. Cancer Manag. Res. 2019, 11, 10287–10296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Li, C.; Hua, F.; Liu, X. The CXCL12/CXCR7 signalling axis promotes proliferation and metastasis in cervical cancer. Med Oncol. 2021, 38, 58. [Google Scholar] [CrossRef]
- Kodama, J.; Hasengaowa; Kusumoto, T.; Seki, N.; Matsuo, T.; Ojima, Y.; Nakamura, K.; Hongo, A.; Hiramatsu, Y. Association of CXCR4 and CCR7 chemokine receptor expression and lymph node metastasis in human cervical cancer. Ann. Oncol. Off. J. Eur. Soc. Med Oncol. 2007, 18, 70–76. [Google Scholar] [CrossRef]
- Zhang, J.P.; Lu, W.G.; Ye, F.; Chen, H.Z.; Zhou, C.Y.; Xie, X. Study on CXCR4/SDF-1alpha axis in lymph node metastasis of cervical squamous cell carcinoma. Int. J. Gynecol. Cancer Off. J. Int. Gynecol. Cancer Soc. 2007, 17, 478–483. [Google Scholar] [CrossRef]
- Yang, Y.C.; Lee, Z.Y.; Wu, C.C.; Chen, T.C.; Chang, C.L.; Chen, C.P. CXCR4 expression is associated with pelvic lymph node metastasis in cervical adenocarcinoma. Int. J. Gynecol. Cancer Off. J. Int. Gynecol. Cancer Soc. 2007, 17, 676–686. [Google Scholar] [CrossRef] [PubMed]
- Walenkamp, A.M.; Boer, I.G.; Bestebroer, J.; Rozeveld, D.; Timmer-Bosscha, H.; Hemrika, W.; van Strijp, J.A.; de Haas, C.J. Staphylococcal superantigen-like 10 inhibits CXCL12-induced human tumor cell migration. Neoplasia 2009, 11, 333–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amine, A.; Rivera, S.; Opolon, P.; Dekkal, M.; Biard, D.S.; Bouamar, H.; Louache, F.; McKay, M.J.; Bourhis, J.; Deutsch, E.; et al. Novel anti-metastatic action of cidofovir mediated by inhibition of E6/E7, CXCR4 and Rho/ROCK signaling in HPV tumor cells. PloS ONE 2009, 4, e5018. [Google Scholar] [CrossRef] [Green Version]
- Chaudary, N.; Mujcic, H.; Wouters, B.G.; Hill, R.P. Hypoxia and metastasis in an orthotopic cervix cancer xenograft model. Radiother. Oncol. J. Eur. Soc. Ther. Radiol. Oncol. 2013, 108, 506–510. [Google Scholar] [CrossRef]
- Brule, S.; Friand, V.; Sutton, A.; Baleux, F.; Gattegno, L.; Charnaux, N. Glycosaminoglycans and syndecan-4 are involved in SDF-1/CXCL12-mediated invasion of human epitheloid carcinoma HeLa cells. Biochim. Biophys. Acta 2009, 1790, 1643–1650. [Google Scholar] [CrossRef]
- Jaafar, F.; Righi, E.; Lindstrom, V.; Linton, C.; Nohadani, M.; Van Noorden, S.; Lloyd, T.; Poznansky, J.; Stamp, G.; Dina, R.; et al. Correlation of CXCL12 expression and FoxP3+ cell infiltration with human papillomavirus infection and clinicopathological progression of cervical cancer. Am. J. Pathol. 2009, 175, 1525–1535. [Google Scholar] [CrossRef] [Green Version]
- Cai, C.; Rodepeter, F.R.; Rossmann, A.; Teymoortash, A.; Lee, J.S.; Quint, K.; Di Fazio, P.; Ocker, M.; Werner, J.A.; Mandic, R. SIVmac(2)(3)(9)-Nef down-regulates cell surface expression of CXCR4 in tumor cells and inhibits proliferation, migration and angiogenesis. Anticancer Res. 2012, 32, 2759–2768. [Google Scholar] [PubMed]
- Wen, W.W.; Xie, S.; Xin, X.L.; Geng, M.Y.; Ding, J.; Chen, Y. Oligomannurarate sulfate inhibits CXCL12/SDF-1-mediated proliferation and invasion of human tumor cells in vitro. Acta Pharmacol. Sin. 2013, 34, 1554–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dillenburg-Pilla, P.; Patel, V.; Mikelis, C.M.; Zarate-Blades, C.R.; Doci, C.L.; Amornphimoltham, P.; Wang, Z.; Martin, D.; Leelahavanichkul, K.; Dorsam, R.T.; et al. SDF-1/CXCL12 induces directional cell migration and spontaneous metastasis via a CXCR4/Galphai/mTORC1 axis. FASEB J. 2015, 29, 1056–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecavalier-Barsoum, M.; Chaudary, N.; Han, K.; Koritzinsky, M.; Hill, R.; Milosevic, M. Targeting the CXCL12/CXCR4 pathway and myeloid cells to improve radiation treatment of locally advanced cervical cancer. Int. J. Cancer 2018, 143, 1017–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Z.; Zhang, P.; Luo, H.; Huang, H.; Wang, F. CXCL12 modulates the radiosensitivity of cervical cancer by regulating CD44. Mol. Med. Rep. 2018, 18, 5101–5108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecavalier-Barsoum, M.; Chaudary, N.; Han, K.; Pintilie, M.; Hill, R.P.; Milosevic, M. Correction: Targeting CXCL12/CXCR4 and myeloid cells to improve the therapeutic ratio in patient-derived cervical cancer models treated with radio-chemotherapy. Br. J. Cancer 2019, 121, 626. [Google Scholar] [CrossRef] [PubMed]
- Hartimath, S.V.; Draghiciu, O.; Daemen, T.; Nijman, H.W.; van Waarde, A.; Dierckx, R.; de Vries, E.F.J. Therapy-Induced Changes in CXCR4 Expression in Tumor Xenografts Can Be Monitored Noninvasively with N-[(11)C]Methyl-AMD3465 PET. Mol. Imaging Biol. 2020, 22, 883–890. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Fan, S.B.; Hua, N.; Li, G.H.; Chang, Q.; Liu, X. Hypermethylation of Single CpG Dinucleotides at the Promoter of CXCL13 Gene Promoting Cell Migration in Cervical Cancer. Curr. Cancer Drug Targets 2020, 20, 355–363. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, J.; Cui, Z.M.; Zhao, J.; Zheng, Y. Expression of the CXCL12/CXCR4 and CXCL16/CXCR6 axes in cervical intraepithelial neoplasia and cervical cancer. Chin. J. Cancer 2013, 32, 289–296. [Google Scholar] [CrossRef] [Green Version]
- CXCL6 C-X-C Motif Chemokine Ligand 6. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/gene/6372 (accessed on 26 July 2021).
- Moeini, S.; Saeidi, M.; Fotouhi, F.; Mondanizadeh, M.; Shirian, S.; Mohebi, A.; Gorji, A.; Ghaemi, A. Synergistic effect of programmed cell death protein 1 blockade and secondary lymphoid tissue chemokine in the induction of anti-tumor immunity by a therapeutic cancer vaccine. Arch. Virol. 2017, 162, 333–346. [Google Scholar] [CrossRef]
- Budczies, J.; Denkert, C.; Gyorffy, B.; Schirmacher, P.; Stenzinger, A. Chromosome 9p copy number gains involving PD-L1 are associated with a specific proliferation and immune-modulating gene expression program active across major cancer types. BMC Med. Genom. 2017, 10, 74. [Google Scholar] [CrossRef] [Green Version]
- Tian, W.J.; Feng, P.H.; Wang, J.; Yan, T.; Qin, Q.F.; Li, D.L.; Liang, W.T. CCR7 Has Potential to Be a Prognosis Marker for Cervical Squamous Cell Carcinoma and an Index for Tumor Microenvironment Change. Front. Mol. Biosci. 2021, 8, 583028. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tiruthani, K.; Li, S.; Hu, M.; Zhong, G.; Tang, Y.; Roy, S.; Zhang, L.; Tan, J.; Liao, C.; et al. mRNA Delivery of a Bispecific Single-Domain Antibody to Polarize Tumor-Associated Macrophages and Synergize Immunotherapy against Liver Malignancies. Adv. Mater. 2021, 33, e2007603. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Yu, W.; Chang, C.; Miyamoto, H.; Liu, X.; Jiang, K.; Yeh, S. Estrogen receptor alpha promotes lung cancer cell invasion via increase of and cross-talk with infiltrated macrophages through the CCL2/CCR2/MMP9 and CXCL12/CXCR4 signaling pathways. Mol. Oncol. 2020, 14, 1779–1799. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Chemokine. | Effects of the Chemokine | Effects in Cervical Cancer | Receptor | References |
|---|---|---|---|---|
| CCL2 (MCP-1) | Migration, infiltration of monocytes and macrophages | Crosstalk between mononuclear cells and HPV-infected epithelia | CCR-2 | [24,26] |
| CCL5 (RANTES) | Chemoattractant for monocytes, memory T-cells, basophils, eosinophils | Elevated in primary tumor and metastatic lesions | CCR-5 | [39] |
| CCL8 (MCP-2) | Recruitment of macrophages via CCR-2- NFκB-pathway | Independent prognosticator for cervical cancer progression | CCR-2 | [41] |
| CCL17 (TARC) | Dose-dependent cell proliferation | High level regulates proliferation of cervical cancer cells via JNK/STAT5 pathways | CCR-4 | [8] |
| CCL19 (MIP-3ß), CCL21 | Proinflammatory | Dysfunctional T-cells in HPV16 positive cervical cancer cells | CCR-7 | [42,47] |
| CCL20 (MIP-3A) | Chemoattractant for lymphocytes and neutrophils, recruitment of Th17 and Treg cells | Down-regulation by HPV16 E6/E7 | CCR-6 | [11] |
| CCL22 | Recruitment of Treg cells | Independent predictor for shorter OS | CCR-4 | [5] |
| Chemokine (Alternate Names) | Effects of the Chemokine | Effects in Cervical Cancer | Receptor | References |
|---|---|---|---|---|
| CXCL1 (GRO1 oncogene, GROα, NAP-3, MGSA-α) | Chemoattractant for immune cells, endothelial tube formation | Increased cervical cancer angiogenesis (AKIP1-dependent) | CXCR2 | [49] |
| CXCL2 (GROβ) | Endothelial tube formation | Increased cervical cancer angiogenesis (AKIP1-dependent) - > increased Cervical cancer cell proliferation | CXCR2 | [51] |
| CXCL3 (GROγ) | Enhanced proliferation and migration activities | Potential tumor marker and interference target | CXCR2 | [52] |
| CXCL5 (ENA78) | Cell proliferation and migration, regulation of expression of tumor-related genes | Contributes to the tumorigenicity of cervical cancer | CXCR2 | [53,54] |
| CXCL6 (GCP2) | Chemotactic for neutrophil granulocytes | Inhibition via Micro-RNA-101-5p leads to Inhibition of tumor growth and metastasis | CXCR1, CXCR2 | [55,82] |
| CXCL8 (IL-8) | Proinflammatory, endothelial tube formation | Increased cervical cancer angiogenesis (AKIP1-dependent), Expression is correlated with clinical stage, distant metastasis, histological type and grade | CXCR1, CXCR2 | [56,57] |
| CXCL10 (IP-10, small-inducible cytokine B10) | Suppression of angiogenesis, modulating formation of micro vessel and expression of E6 and E7 | Prognostic indicator for cervical cancer, May be used as gene therapy in combination with radiotherapy | CXCR3 | [58,59] |
| CXCL11 (I-TAC) | Chemotactic for interleukin-activated T-cells | Higher expression of CXCR7 is associated with shorter DFS and OS | CXCR3, CXCR7 | [63,82] |
| CXCL12 (SDF1) | Induction of directed cell migration | Chemotaxis and rescue from apoptosis, linked to tumor dissemination and poor prognosis | CXCR4, CXCR7 | [65,66,74] |
| CXCL13 (BCA1) | Inhibits cell migration | Low expression is associated with risk of death | CXCR3, CXCR5 | [80] |
| CXCL16 | Co-expression with CXCL12/CXCR4 - > durative process in cervical cancer development | Biomarker, prognostic factor | CXCR6 | [81] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garrido, F.; Wild, C.M.; Mittelberger, J.; Dobler, F.; Schneider, M.; Ansorge, N.; Köpke, M.; Strieder, A.; Ditsch, N.; Jeschke, U.; et al. The Role of Chemokines in Cervical Cancers. Medicina 2021, 57, 1141. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57111141
Garrido F, Wild CM, Mittelberger J, Dobler F, Schneider M, Ansorge N, Köpke M, Strieder A, Ditsch N, Jeschke U, et al. The Role of Chemokines in Cervical Cancers. Medicina. 2021; 57(11):1141. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57111141
Chicago/Turabian StyleGarrido, Fabian, Carl Mathis Wild, Johanna Mittelberger, Franziska Dobler, Mariella Schneider, Nadine Ansorge, Melitta Köpke, Annamarie Strieder, Nina Ditsch, Udo Jeschke, and et al. 2021. "The Role of Chemokines in Cervical Cancers" Medicina 57, no. 11: 1141. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57111141