
Stimulation of Toll-Like Receptor 3 Diminishes Intracellular Growth of Salmonella Typhimurium by Enhancing Autophagy in Murine Macrophages

 , and
, and 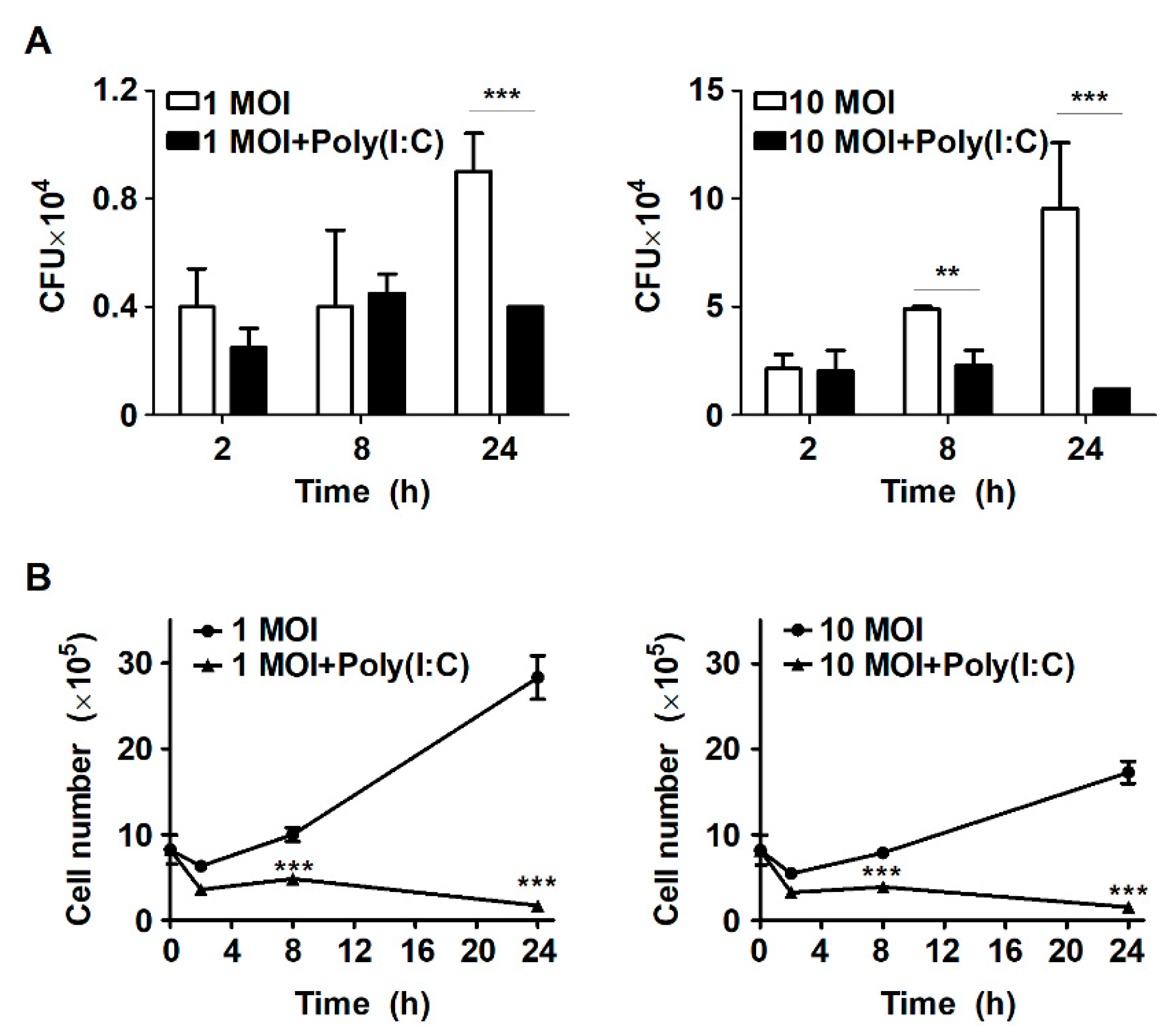{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Treatment with Poly(I:C) Suppresses Intracellular Bacterial Growth in S. Typhimurium-Infected Murine Macrophages
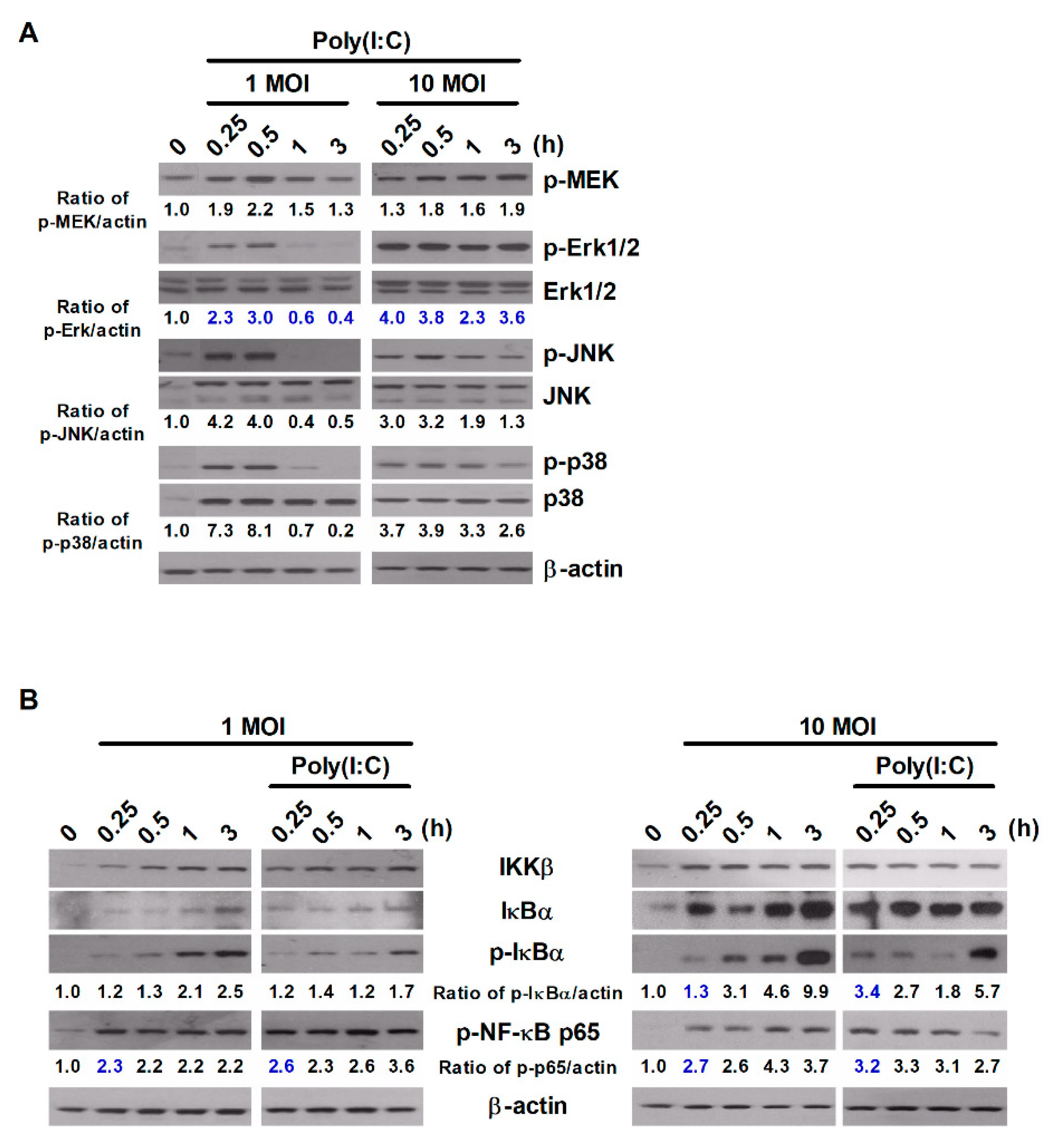
2.2. Poly(I:C) Activates the MAPK and NF-κB Signaling Pathways during S. Typhimurium Infection
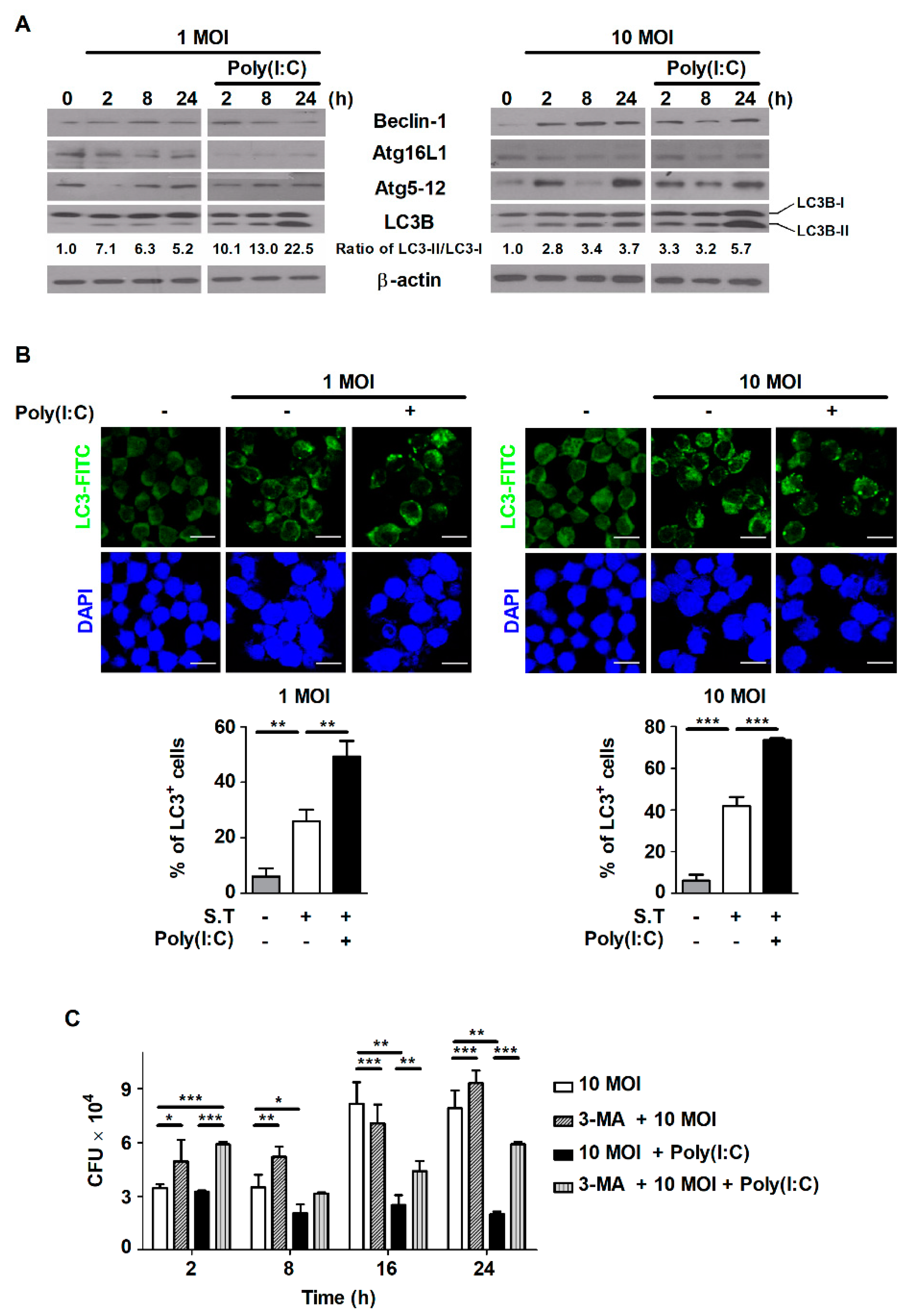
2.3. Poly(I:C) Induces Autophagy in S. Typhimurium-Infected Cells
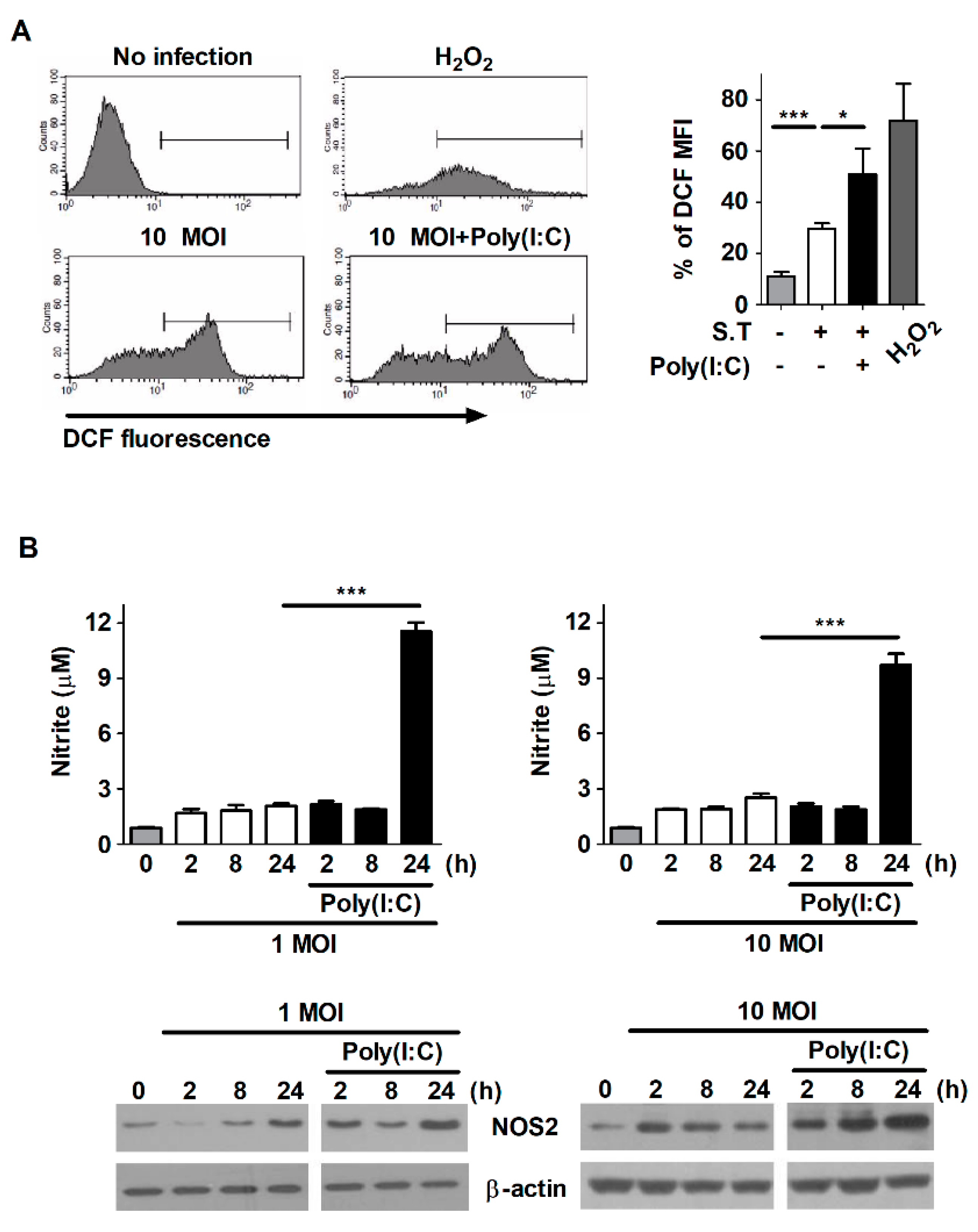
2.4. Poly(I:C) Enhances the Production of ROS and NO in S. Typhimurium-Infected Cells
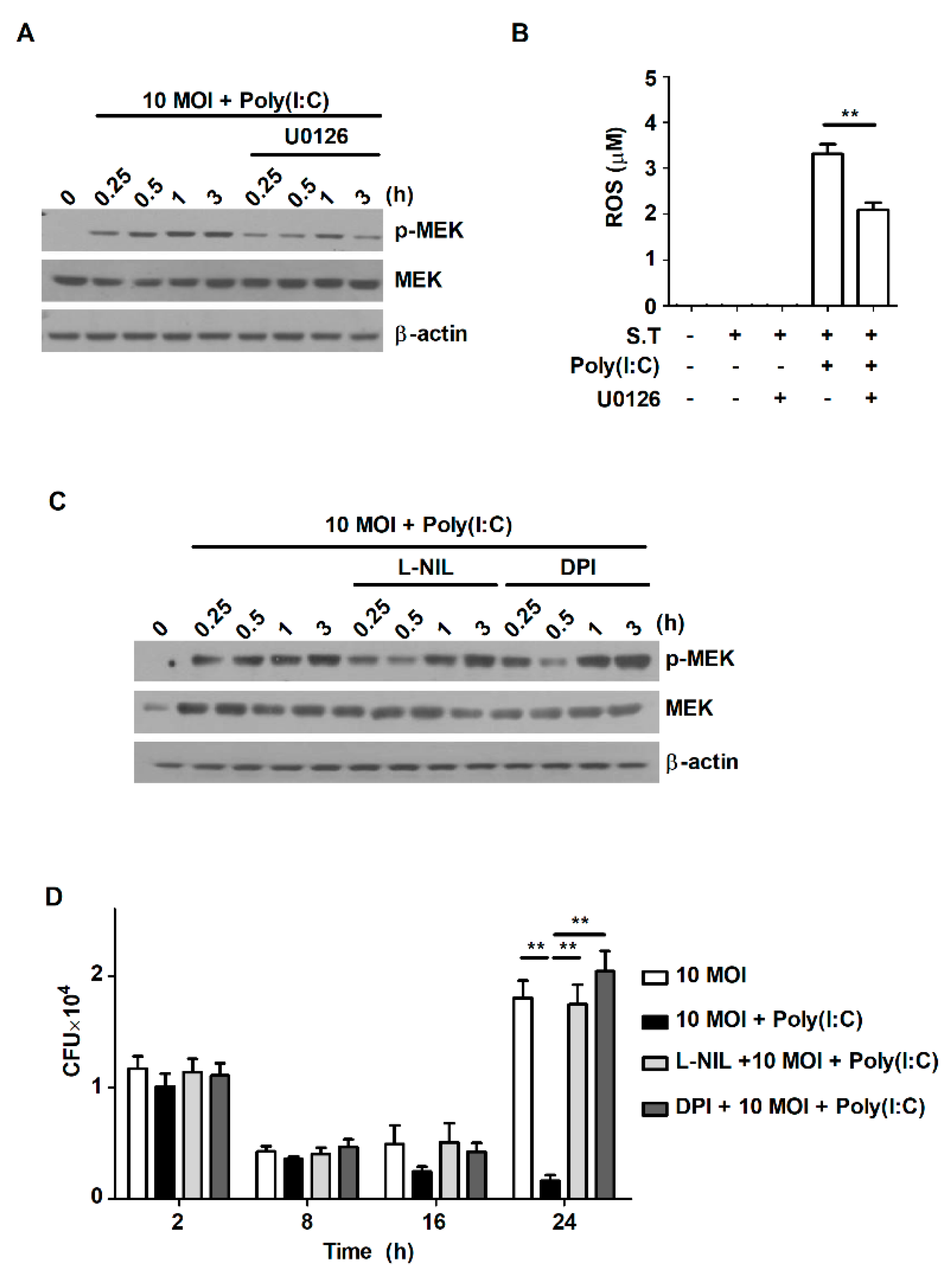
2.5. Poly(I:C) Controls Intracellular Bacterial Growth by Increasing the Production of ROS through the MEK1/2-Mediated Pathway in S. Typhimurium-Infected Macrophages
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. S. Typhimurium Culture Conditions
4.4. S. Typhimurium Infection In Vitro
4.5. Cell Treatments
4.6. Cell Viability Assay
4.7. Colony-Forming Unit (CFU) Assay
4.8. Western Blot Analysis
4.9. Immunofluorescence Analysis
4.10. Flow Cytometry
4.11. ROS Detection Assay
4.12. NO Detection Assay
4.13. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Broz, P.; Ohlson, M.B.; Monack, D.M. Innate immune response to Salmonella typhimurium, a model enteric pathogen. Gut Microbes 2012, 3, 62–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urdaneta, V.; Casadesus, J. Host-pathogen interactions in typhoid fever: The model is the message. Ann. Transl. Med. 2018, 6, S38. [Google Scholar] [CrossRef] [Green Version]
- Hurley, D.; McCusker, M.P.; Fanning, S.; Martins, M. Salmonella-host interactions-modulation of the host innate immune system. Front. Immunol. 2014, 5, 481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerovic, V.; Bain, C.C.; Mowat, A.M.; Milling, S.W. Intestinal macrophages and dendritic cells: What’s the difference? Trends Immunol. 2014, 35, 270–277. [Google Scholar] [CrossRef]
- Yoon, M.Y.; Lee, K.; Yoon, S.S. Protective role of gut commensal microbes against intestinal infections. J. Microbiol. 2014, 52, 983–989. [Google Scholar] [CrossRef]
- Galan, J.E. Salmonella Typhimurium and inflammation: A pathogen-centric affair. Nat. Rev. Microbiol. 2021. [Google Scholar] [CrossRef]
- de Jong, H.K.; Parry, C.M.; van der Poll, T.; Wiersinga, W.J. Host-pathogen interaction in invasive Salmonellosis. PLoS Pathog. 2012, 8, e1002933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarr, D.; Toth, E.; Gingerich, A.; Rada, B. Antimicrobial actions of dual oxidases and lactoperoxidase. J. Microbiol. 2018, 56, 373–386. [Google Scholar] [CrossRef]
- Lee, H.J.; Woo, Y.; Hahn, T.W.; Jung, Y.M.; Jung, Y.J. Formation and Maturation of the Phagosome: A Key Mechanism in Innate Immunity against Intracellular Bacterial Infection. Microorganisms 2020, 8, 1298. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, N.; Godec, J.; Lau, L.; Sivick, K.E.; McLaughlin, L.M.; Jones, M.B.; Dracheva, T.; Peterson, S.N.; Monack, D.M.; Barton, G.M. TLR Signaling Is Required for Salmonella typhimurium Virulence. Cell 2011, 144, 675–688. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Jia, H.; Jing, Z.; Liu, D. Recognition of pathogen-associated nucleic acids by endosomal nucleic acid-sensing toll-like receptors. Acta Biochim. Biophys. Sin. 2013, 45, 241–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pothlichet, J.; Meunier, I.; Davis, B.K.; Ting, J.P.; Skamene, E.; von Messling, V.; Vidal, S.M. Type I IFN triggers RIG-I/TLR3/NLRP3-dependent inflammasome activation in influenza A virus infected cells. PLoS Pathog. 2013, 9, e1003256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, C.E.; Sad, S.; Coombes, B.K. Salmonella enterica serovar typhimurium exploits Toll-like receptor signaling during the host-pathogen interaction. Infect. Immun. 2009, 77, 4750–4760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, N. Salmonella Typhimurium infection: Type I Interferons integrate cellular networks to disintegrate macrophages. Cell Stress 2018, 2, 37–39. [Google Scholar] [CrossRef] [Green Version]
- Kocaturk, N.M.; Gozuacik, D. Crosstalk Between Mammalian Autophagy and the Ubiquitin-Proteasome System. Front. Cell Dev. Biol. 2018, 6, 128. [Google Scholar] [CrossRef]
- Escobar-Henriques, M.; Altin, S.; den Brave, F. Interplay between the Ubiquitin Proteasome System and Mitochondria for Protein Homeostasis. Curr. Issues Mol. Biol. 2020, 35, 35–58. [Google Scholar] [CrossRef] [Green Version]
- Chun, Y.; Kim, J. Autophagy: An Essential Degradation Program for Cellular Homeostasis and Life. Cells 2018, 7, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, G.; Shravage, B.V.; Baehrecke, E.H. Regulation and Function of Autophagy during Cell Survival and Cell Death. Cold Spring Harb. Perspect. Biol. 2012, 4, a008813. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.Y.; Lu, J.H. Autophagy and Macrophage Functions: Inflammatory Response and Phagocytosis. Cells 2019, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birmingham, C.L.; Smith, A.C.; Bakowski, M.A.; Yoshimori, T.; Brumell, J.H. Autophagy controls Salmonella infection in response to damage to the Salmonella-containing vacuole. J. Biol. Chem. 2006, 281, 11374–11383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Kang, S.J.; Woo, Y.; Hahn, T.W.; Ko, H.J.; Jung, Y.J. TLR7 Stimulation With Imiquimod Induces Selective Autophagy and Controls Mycobacterium tuberculosis Growth in Mouse Macrophages. Front. Microbiol. 2020, 11, 1684. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Zhang, S.P.; Wang, J.F.; Liu, L.; Wang, Y.; Cao, Z.Y.; Hu, Q.K.; Yuan, W.J.; Lin, L. TLR3 contributes to persistent autophagy and heart failure in mice after myocardial infarction. J. Cell Mol. Med. 2018, 22, 395–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Fattah, E.A.; Liu, X.D.; Jagannath, C.; Eissa, N.T. Harnessing of TLR-mediated autophagy to combat mycobacteria in macrophages. Tuberculosis 2013, 93, S33–S37. [Google Scholar] [CrossRef]
- Herb, M.; Schramm, M. Functions of ROS in Macrophages and Antimicrobial Immunity. Antioxidants 2021, 10, 313. [Google Scholar] [CrossRef] [PubMed]
- Jamaati, H.; Mortaz, E.; Pajouhi, Z.; Folkerts, G.; Movassaghi, M.; Moloudizargari, M.; Adcock, I.M.; Garssen, J. Nitric Oxide in the Pathogenesis and Treatment of Tuberculosis. Front. Microbiol. 2017, 8, 2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhen, M. Salmonella and Reactive Oxygen Species: A Love-Hate Relationship. J. Innate Immun. 2019, 11, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, D.L.; Liu, Y.L. Poly (I:C) transfection induces mitochondrial-mediated apoptosis in cervical cancer. Mol. Med. Rep. 2016, 13, 2689–2695. [Google Scholar] [CrossRef]
- Yang, C.S.; Kim, J.J.; Lee, S.J.; Hwang, J.H.; Lee, C.H.; Lee, M.S.; Jo, E.K. TLR3-triggered reactive oxygen species contribute to inflammatory responses by activating signal transducer and activator of transcription-1. J. Immunol. 2013, 190, 6368–6377. [Google Scholar] [CrossRef] [Green Version]
- West, A.P.; Brodsky, I.E.; Rahner, C.; Woo, D.K.; Erdjument-Bromage, H.; Tempst, P.; Walsh, M.C.; Choi, Y.; Shadel, G.S.; Ghosh, S. TLR signalling augments macrophage bactericidal activity through mitochondrial ROS. Nature 2011, 472, 476–543. [Google Scholar] [CrossRef] [Green Version]
- Rosenberger, C.M.; Finlay, B.B. Macrophages inhibit Salmonella typhimurium replication through MEK/ERK kinase and phagocyte NADPH oxidase activities. J. Biol. Chem. 2002, 277, 18753–18762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabrega, A.; Vila, J. Salmonella enterica serovar Typhimurium skills to succeed in the host: Virulence and regulation. Clin. Microbiol. Rev. 2013, 26, 308–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiner, T.S. How flagellin and toll-like receptor 5 contribute to enteric infection. Infect. Immun. 2007, 75, 545–552. [Google Scholar] [CrossRef] [Green Version]
- Zheng, D.P.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Germic, N.; Frangez, Z.; Yousefi, S.; Simon, H.U. Regulation of the innate immune system by autophagy: Monocytes, macrophages, dendritic cells and antigen presentation. Cell Death Differ. 2019, 26, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Sanjuan, M.A.; Dillon, C.P.; Tait, S.W.; Moshiach, S.; Dorsey, F.; Connell, S.; Komatsu, M.; Tanaka, K.; Cleveland, J.L.; Withoff, S.; et al. Toll-like receptor signalling in macrophages links the autophagy pathway to phagocytosis. Nature 2007, 450, 1253–1257. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Deng, S.; Cao, Y.; Zhang, R.; Wang, Z.; Jiang, X.; Wang, J.; Zhang, X.; Zhang, J.; Liu, G.; et al. Overexpression of Toll-Like Receptor 4 Contributes to Phagocytosis of Salmonella Enterica Serovar Typhimurium via Phosphoinositide 3-Kinase Signaling in Sheep. Cell Physiol. Biochem. 2018, 49, 662–677. [Google Scholar] [CrossRef]
- Doyle, S.E.; O’Connell, R.M.; Miranda, G.A.; Vaidya, S.A.; Chow, E.K.; Liu, P.T.; Suzuki, S.; Suzuki, N.; Modlin, R.L.; Yeh, W.C.; et al. Toll-like receptors induce a phagocytic gene program through p38. J. Exp. Med. 2004, 199, 81–90. [Google Scholar] [CrossRef]
- Zhan, R.; Han, Q.; Zhang, C.; Tian, Z.; Zhang, J. Toll-Like receptor 2 (TLR2) and TLR9 play opposing roles in host innate immunity against Salmonella enterica serovar Typhimurium infection. Infect. Immun. 2015, 83, 1641–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.T.; Coveney, A.P.; Wu, M.; Huang, J.; Blankson, S.; Zhao, H.; O’Leary, D.P.; Bai, Z.J.; Li, Y.P.; Redmond, H.P.; et al. Activation of Both TLR and NOD Signaling Confers Host Innate Immunity-Mediated Protection against Microbial Infection. Front. Immunol. 2019, 9, 3082. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L.A.; Bryant, C.E.; Doyle, S.L. Therapeutic targeting of Toll-like receptors for infectious and inflammatory diseases and cancer. Pharmacol. Rev. 2009, 61, 177–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.H.; Lee, H.J.; Ko, H.J.; Yoon, B.I.; Choe, J.; Kim, K.C.; Hahn, T.W.; Han, J.A.; Choi, S.S.; Jung, Y.M.; et al. The TLR7 agonist imiquimod induces anti-cancer effects via autophagic cell death and enhances anti-tumoral and systemic immunity during radiotherapy for melanoma. Oncotarget 2017, 8, 24932–24948. [Google Scholar] [CrossRef] [Green Version]
- Yi, J.Y.; Jung, Y.J.; Choi, S.S.; Hwang, J.; Chung, E. Autophagy-mediated anti-tumoral activity of imiquimod in Caco-2 cells. Biochem. Biophys. Res. Commun. 2009, 386, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.W.; Liu, K.T.; Chang, C.C.; Chen, Y.J.; Wu, C.Y.; Tsai, J.J.; Lu, W.C.; Wang, Y.T.; Liu, C.M.; Shieh, J.J. Imiquimod simultaneously induces autophagy and apoptosis in human basal cell carcinoma cells. Br. J. Dermatol. 2010, 163, 310–320. [Google Scholar] [CrossRef] [PubMed]
- De Waele, J.; Verhezen, T.; van der Heijden, S.; Berneman, Z.N.; Peeters, M.; Lardon, F.; Wouters, A.; Smits, E. A systematic review on poly(I:C) and poly-ICLC in glioblastoma: Adjuvants coordinating the unlocking of immunotherapy. J. Exp. Clin. Cancer Res. 2021, 40, 213. [Google Scholar] [CrossRef]
- Lantier, L.; Drouet, F.; Guesdon, W.; Mancassola, R.; Metton, C.; Lo-Man, R.; Werts, C.; Laurent, F.; Lacroix-Lamande, S. Poly(I:C)-induced protection of neonatal mice against intestinal Cryptosporidium parvum infection requires an additional TLR5 signal provided by the gut flora. J. Infect. Dis. 2014, 209, 457–467. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Shen, Y.; Zhang, S.; Xiao, Y.; Shi, S. Salmonella Interacts With Autophagy to Offense or Defense. Front. Microbiol. 2020, 11, 721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, J.E.; Lee, H.K. Pattern recognition receptors and autophagy. Front. Immunol 2014, 5, 300. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Jagannath, C.; Liu, X.D.; Sharafkhaneh, A.; Kolodziejska, K.E.; Eissa, N.T. Toll-like receptor 4 is a sensor for autophagy associated with innate immunity. Immunity 2007, 27, 135–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Zhuang, J.; Jiang, Y.Y.; Sun, J.; Prinz, R.A.; Sun, J.; Jiao, X.N.; Xu, X.L. Toll-like receptor signalling cross-activates the autophagic pathway to restrict Salmonella Typhimurium growth in macrophages. Cell Microbiol. 2019, 21, e13095. [Google Scholar] [CrossRef]
- Lee, H.J.; Hong, W.G.; Woo, Y.; Ahn, J.H.; Ko, H.J.; Kim, H.; Moon, S.; Hahn, T.W.; Jung, Y.M.; Song, D.K.; et al. Lysophosphatidylcholine Enhances Bactericidal Activity by Promoting Phagosome Maturation via the Activation of the NF-kappaB Pathway during Salmonella Infection in Mouse Macrophages. Mol. Cells 2020, 43, 989–1001. [Google Scholar] [CrossRef]
- Woo, Y.; Lee, H.J.; Kim, J.; Kang, S.G.; Moon, S.; Han, J.A.; Jung, Y.M.; Jung, Y.J. Rapamycin Promotes ROS-Mediated Cell Death via Functional Inhibition of xCT Expression in Melanoma Under gamma-Irradiation. Front. Oncol. 2021, 11, 665420. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Ko, H.J.; Kim, S.H.; Jung, Y.J. Pasakbumin A controls the growth of Mycobacterium tuberculosis by enhancing the autophagy and production of antibacterial mediators in mouse macrophages. PLoS ONE 2019, 14, e0199799. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-J.; Lee, S.-H.; Jeon, J.-H.; Kim, H.-J.; Jeong, E.-K.; Kim, M.-J.; Jung, Y.M.; Jung, Y.-J. Stimulation of Toll-Like Receptor 3 Diminishes Intracellular Growth of Salmonella Typhimurium by Enhancing Autophagy in Murine Macrophages. Metabolites 2021, 11, 602. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11090602
Lee H-J, Lee S-H, Jeon J-H, Kim H-J, Jeong E-K, Kim M-J, Jung YM, Jung Y-J. Stimulation of Toll-Like Receptor 3 Diminishes Intracellular Growth of Salmonella Typhimurium by Enhancing Autophagy in Murine Macrophages. Metabolites. 2021; 11(9):602. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11090602
Chicago/Turabian StyleLee, Hyo-Ji, Sun-Hye Lee, Ji-Hui Jeon, Hyo-Jung Kim, Eui-Kwon Jeong, Min-Jeong Kim, Young Mee Jung, and Yu-Jin Jung. 2021. "Stimulation of Toll-Like Receptor 3 Diminishes Intracellular Growth of Salmonella Typhimurium by Enhancing Autophagy in Murine Macrophages" Metabolites 11, no. 9: 602. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11090602