
Downregulation of Squalene Synthase Broadly Impacts Isoprenoid Biosynthesis in Guayule


 ,
,  and
and 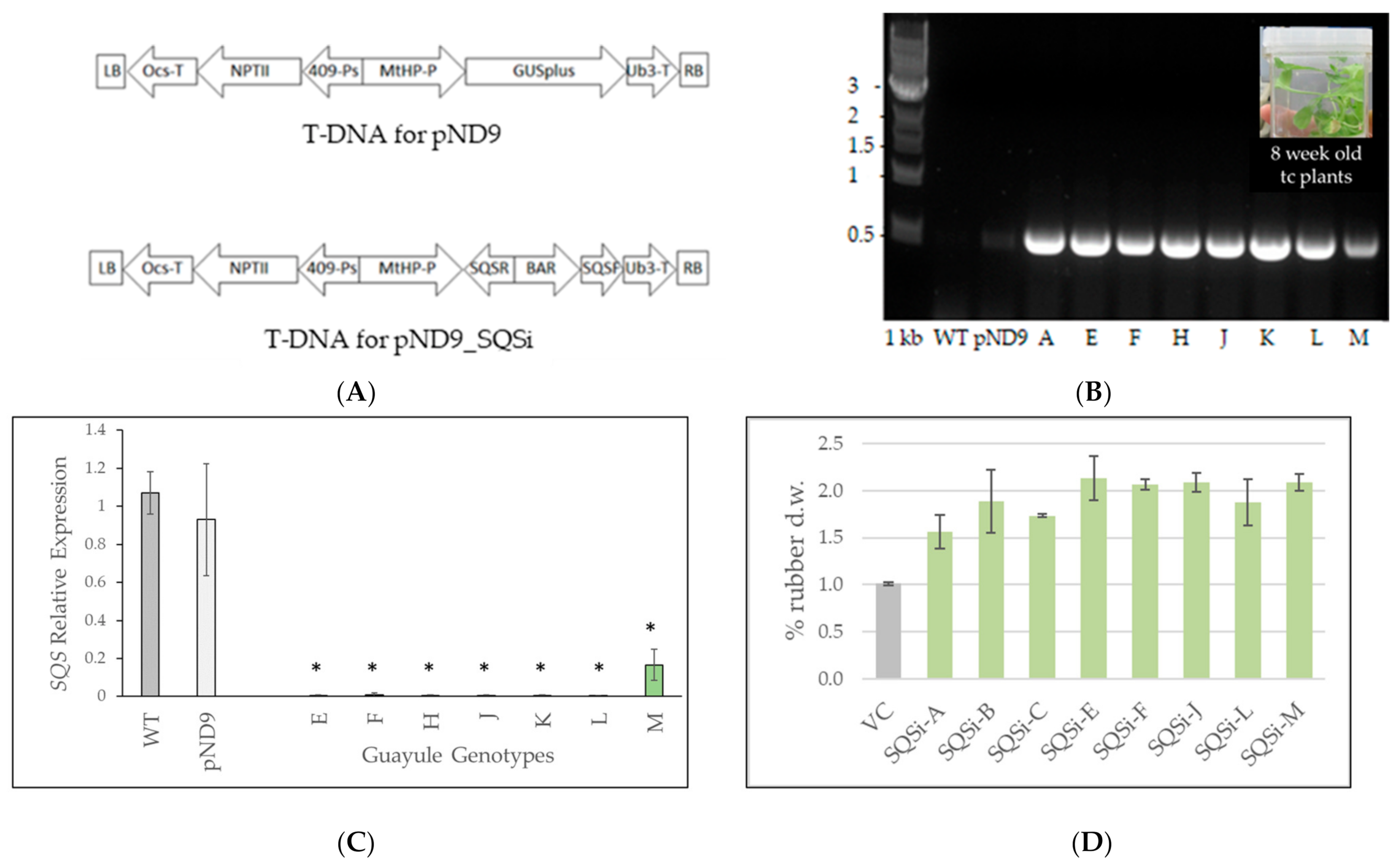{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
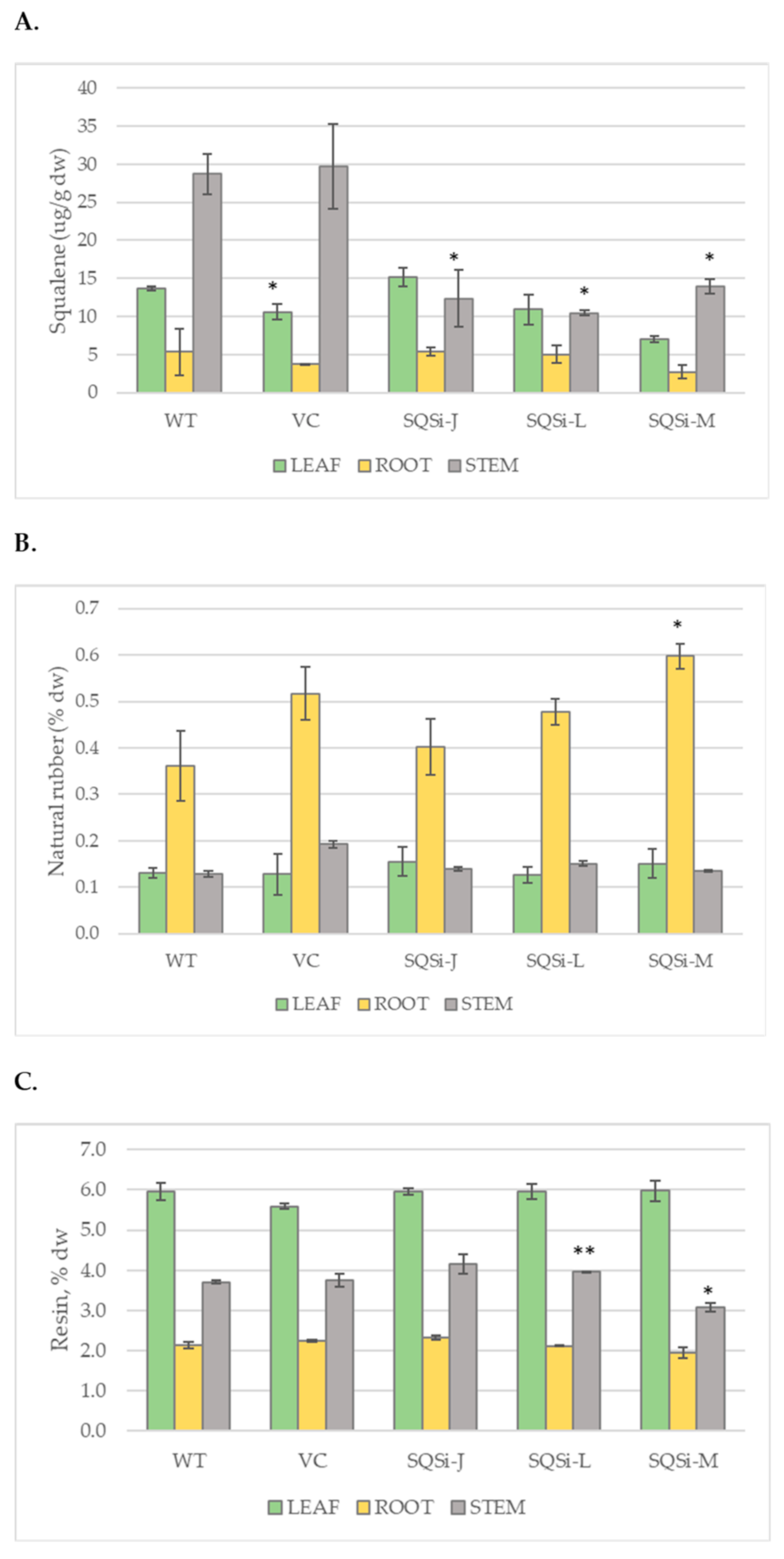
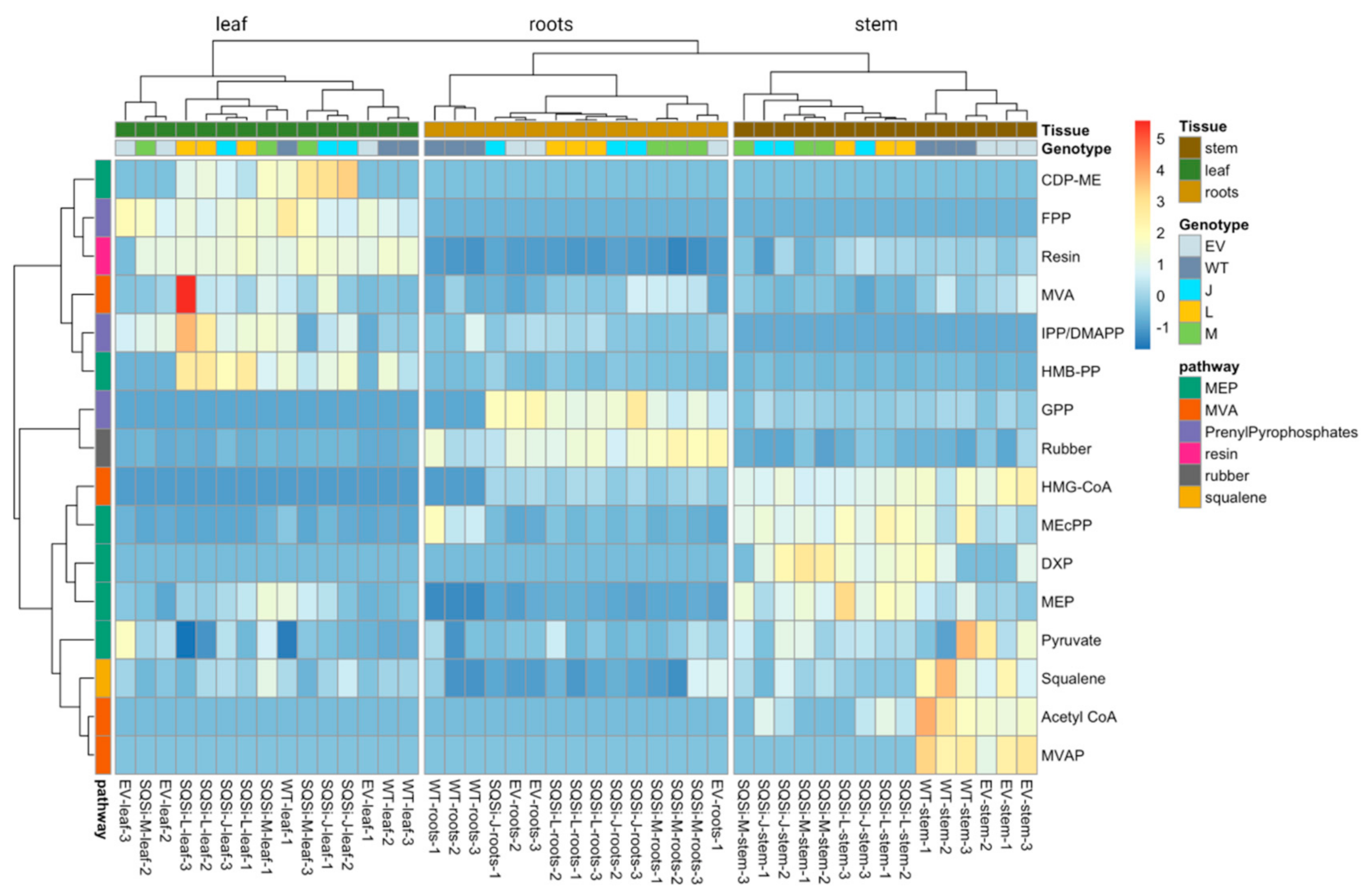
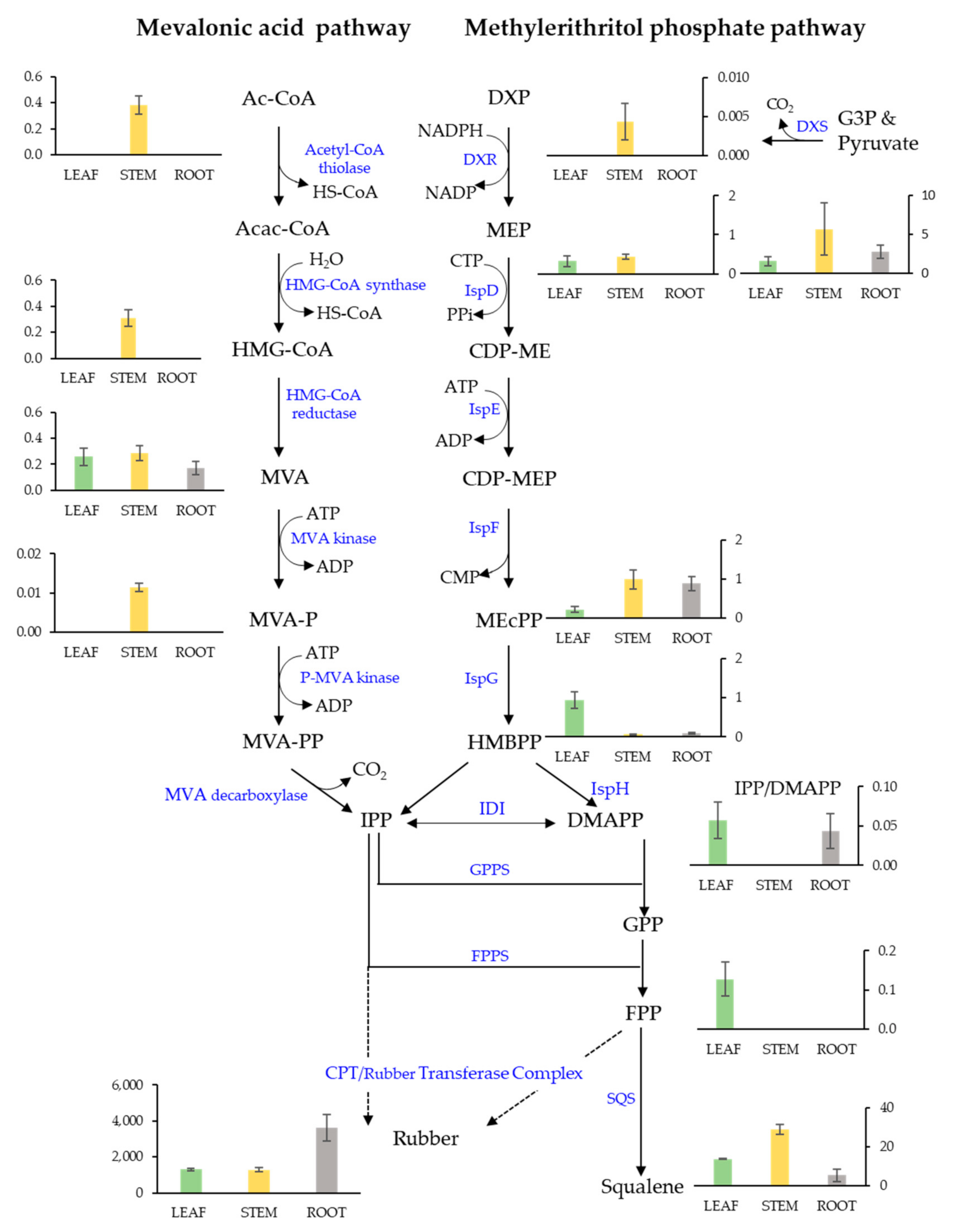
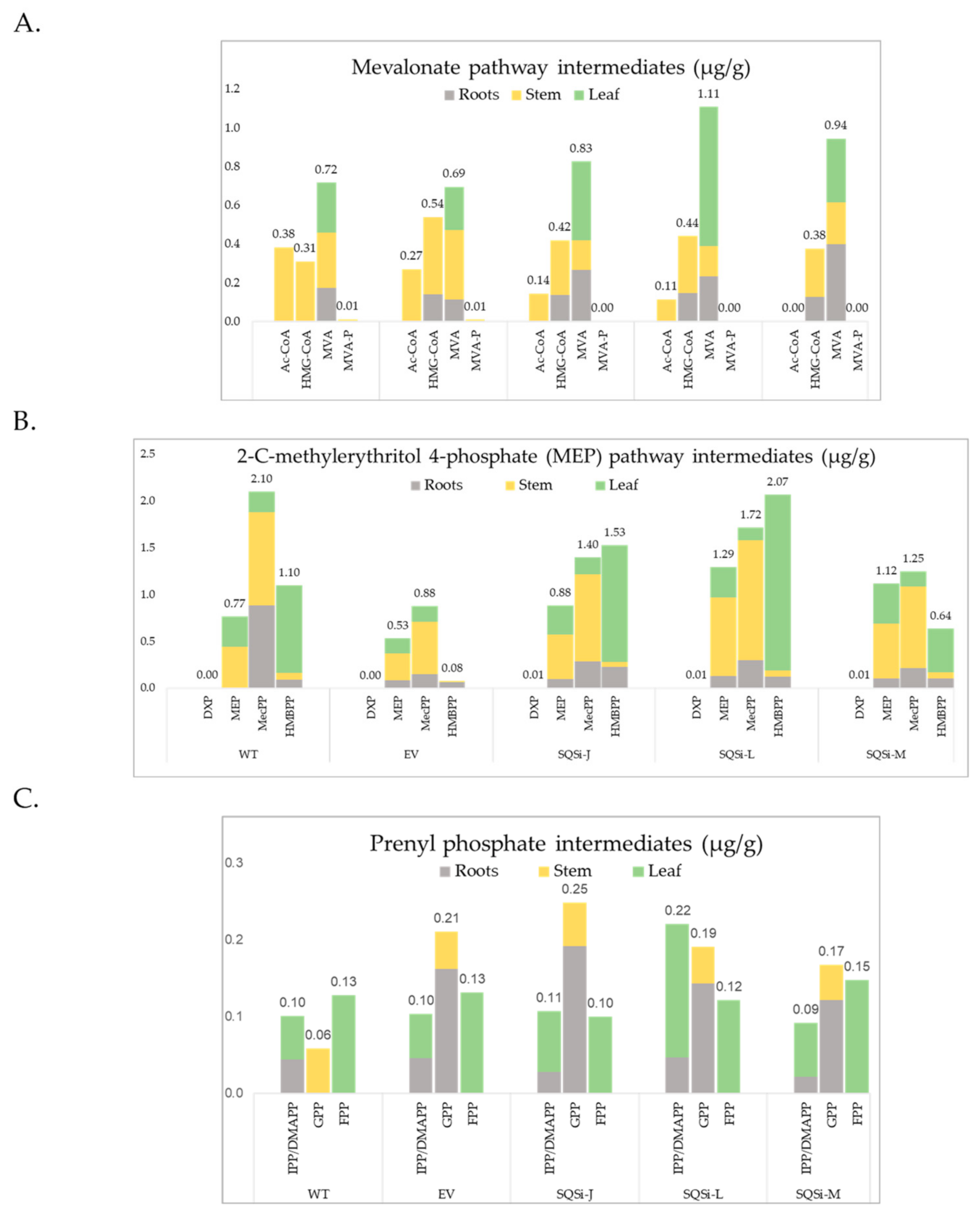
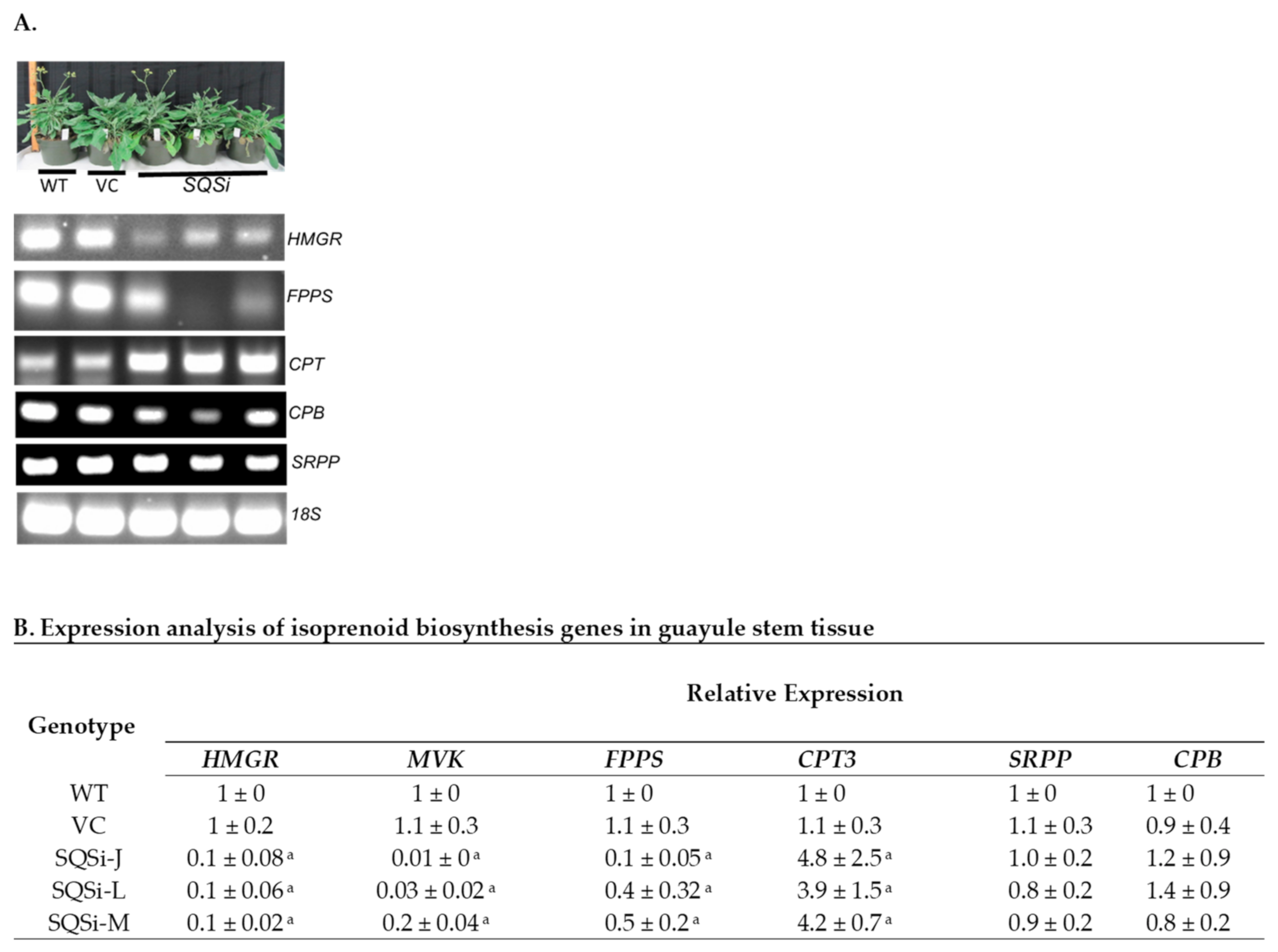
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Transformation Plasmid Construction
4.2. Agrobacterium and Leaf Tissue Transformation
4.3. Genomic DNA and RNA Extractions and PCR and qPCR Analyses
4.4. Rubber and Resin Extractions
4.5. Transplanting and Plant Architecture Measurements
4.6. Squalene Extraction and Purification
4.7. Squalene Quantification by GC-MS
4.8. Quantification of Isoprenoid Intermediates by LC-MS
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonner, J.; Galston, A.W. The physiology and biochemistry of rubber formation in plants. Bot. Rev. 1947, 13, 543–596. [Google Scholar] [CrossRef]
- Bowers, J.E. Natural Rubber-Producing Plants for the United States; USDA, Cooperative State Research Service and National Agricultural Library: Beltsville, MD, USA, 1990; pp. 1–43. [Google Scholar]
- Wood, D.F.; Cornish, K. Microstructure of purified rubber particles. Int. J. Plant Sci. 2000, 161, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Konno, K. Plant latex and other exudates as plant defense systems: Roles of various defense chemicals and proteins contained therein. Phytochemistry 2011, 72, 1510–1530. [Google Scholar] [CrossRef] [PubMed]
- Bonner, J. Effect of temperature on rubber accumulation in guayule plant. Bot. Gaz. 1943, 105, 233–243. [Google Scholar] [CrossRef]
- Benedict, C.R.; Greer, P.J.; Foster, M.A. The physiological and biochemical responses of guayule to the low temperature of the Chihuahuan Desert in the biosynthesis of rubber. Ind. Crop. Prod. 2008, 27, 225–235. [Google Scholar] [CrossRef]
- Ponciano, G.; McMahan, C.M.; Xie, W.; Lazo, G.R.; Coffelt, T.A.; Collins-Silva, J.; Nural-Taban, A.; Gollery, M.; Shintani, D.K.; Whalen, M.C. Transcriptome and gene expression analysis in cold-acclimated guayule (Parthenium argentatum) rubber-producing tissue. Phytochemistry 2012, 79, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Chapelle, J. The genetics and molecular genetics of terpene and sterol origami. Curr. Opin. Plant Biol. 2002, 5, 151–157. [Google Scholar] [CrossRef]
- Hemmerlin, A.; Harwood, J.L.; Bach, T.J. A raison d’etre for two distinct pathways in the early steps of plant isoprenoid biosynthesis? Prog. Lipid Res. 2012, 51, 95–148. [Google Scholar]
- Tetali, S.D. Terpenes and isoprenoids: A wealth of compounds for global use. Planta 2019, 249, 1–8. [Google Scholar] [CrossRef]
- Veatch, M.E.; Ray, D.T.; Mau, C.J.D.; Cornish, K. Growth, rubber and resin evaluation of two-year-old transgenic guayule. Ind. Crop. Prod. 2005, 22, 65–74. [Google Scholar] [CrossRef]
- Dong, N.; Ponciano, G.; McMahan, C.M.; Coffelt, T.A.; Johnson, L.; Creelman, R.; Whalen, M.C.; Cornish, K. Overexpression of 3-hydroxy-3-methylglutaryl coenzyme A reductase in Parthenium argentatum (guayule). Ind. Crop. Prod. 2013, 46, 15–24. [Google Scholar] [CrossRef]
- Pütter, K.M.; van Deenen, N.; Unland, K.; Prüfer, D.; Schulze Gronover, C. Isoprenoid biosynthesis in dandelion latex is enhanced by the overexpression of three key enzymes involved in the mevalonate pathway. BMC Plant Biol. 2017, 17, 88. [Google Scholar] [CrossRef] [PubMed]
- Ponciano, G.; Chen, G.Q.; McMahan, C. A bicistronic transgene system for genetic modification of Parthenium Argentatum. Plant Biotechnol. Rep. 2018, 12, 149–155. [Google Scholar] [CrossRef]
- Kumar, S.; Hahn, F.M.; Baidoo, E.; Kahlon, T.S.; Wood, D.; McMahan, C.M.; Cornish, K.; Keasling, J.D.; Daniell, H.; Whalen, M. Remodeling the isoprenoid pathway in tobacco by expressing the cytoplasmic mevalonate pathway in chloroplasts. Metab. Eng. 2012, 14, 19–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spanova, M.; Daum, G. Squalene-biochemistry, molecular biology, process biotechnology, and applications. Eur. J. Lipid Sci. Tech. 2011, 113, 1299–1320. [Google Scholar] [CrossRef]
- Lozano-Grande, M.A.; Gorinstein, S.; Espitia-Rangel, E.; Davila-Ortiz, G.; Martinez-Ayala, A.L. Plant sources, extraction methods, and uses of squalene. Int. J. Agron. 2018, 2018, 1829160. [Google Scholar] [CrossRef]
- Clouse, S.D. Arabidopsis mutants reveal multiple roles for sterols in plant development. Plant Cell 2002, 14, 1995–2000. [Google Scholar] [CrossRef] [Green Version]
- Rogowska, A.; Szakiel, A. The role of sterols in plant response to abiotic stress. Phytochem. Rev. 2020, 19, 1525–1538. [Google Scholar] [CrossRef]
- González-Coloma, A.; López-Balboa, C.; Santana, O.; Reina, M.; Fraga, B.M. Triterpene-based plant defenses. Phytochem. Rev. 2011, 10, 245–260. [Google Scholar] [CrossRef] [Green Version]
- Lindsey, K.; Pullen, M.L.; Topping, J.F. Importance of plant sterols in pattern formation and hormone signaling. Trends Plant Sci. 2003, 8, 521–525. [Google Scholar] [CrossRef]
- Lee, M.-H.; Jeong, J.-H.; Seo, J.-W.; Shin, C.-G.; Kim, Y.-S.; In, J.-G.; Yang, D.-C.; Yi, J.-S.; Choi, Y.-E. Enhanced triterpene and phytosterol biosynthesis in Panax ginseng overexpressing squalene synthase gene. Plant Cell Physiol. 2004, 45, 976–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, J.-W.; Jeong, J.-H.; Shin, C.-G.; Lo, S.-C.; Han, S.-S.; Yu, K.-W.; Harada, E.; Han, J.-Y.; Choi, Y.-E. Overexpression of squalene synthase in Eleutherococcus senticosus increases physterol and triterpene accumulation. Phytochemistry 2005, 66, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Cho, J.H.; Park, S.; Han, J.-Y.; Back, K.; Choi, Y.-E. Gene regulation patterns triterpene biosynthetic pathway driven by overexpression of squalene synthase and methyl jasmonate elicitation in Bupleurum falcatum. Planta 2011, 233, 343–355. [Google Scholar] [CrossRef]
- Mirjalili, M.H.; Moyano, E.; Bonfill, M.; Cusido, R.M.; Palazon, J. Overexpression of the Arabidopsis thaliana squalene synthase gene in Withania coagulans hairy root cultures. Biol. Plant. 2011, 55, 357–360. [Google Scholar] [CrossRef]
- Nguyen, H.T.M.; Neelakadan, A.K.; Quach, T.N.; Valliyodan, B.; Kumar, R.; Zhang, Z.; Nguyen, H.T. Molecular characterization of Glycine max squalene synthase genes in seed phytosterol biosynthesis. Plant Physiol. Biochem. 2013, 73, 23–32. [Google Scholar] [CrossRef]
- Ali, A.; Ahmad, M.M.; Khan, M.A.; Saxena, P.; Khan, S.; Abdin, M.Z. RNAi-mediated modulation of squalene synthase gene expression in Artemisia annua L. and its impact on artemisinin biosynthesis. Rend. Fis. Acc. Lincei 2017, 28, 731–741. [Google Scholar] [CrossRef]
- Singh, A.K.; Dwivedi, V.; Rai, A.; Pal, S.; Reddy, S.G.E.; Rao, D.K.V.; Shasany, A.K.; Nagegowda, D.A. Virus-induced gene silencing of Withania somnifera squalene synthase negatively regulates sterol and defence-related genes resulting in reduced withanolides and biotic stress tolerance. Plant Biotechnol. J. 2015, 13, 1287–1299. [Google Scholar] [CrossRef]
- Navarro Gallon, S.M.; Elejalde-Palmett, C.; Daudu, D.; Liesecke, F.; Jullien, F.; Papon, N.; de Bernonville, T.D.; Courdavault, V.; Lanoue, A.; Oudin, A.; et al. Virus-induced gene silencing of the two squalene synthase isoforms of apple tree (Malus x domestica L.) negatively impacts phytosterol biosynthesis, plastid pigmentatin and leaf growth. Planta 2017, 246, 46–60. [Google Scholar] [CrossRef]
- Manavalan, L.P.; Chen, X.; Clarke, J.; Salmeron, J.; Nguyen, H.T. RNAi-mediated disruption of squalene synthase improves drought tolerance and yield in rice. J. Exp. Bot. 2012, 63, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef]
- Hannon, G. RNA interference. Nature 2002, 418, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.; Dong, C.; Ponciano, G.; Holtman, K.; Placido, D.; Coffelt, T.; Whalen, M.; McMahan, C. Fructan reduction by downregulation of 1-SST in guayule. Ind. Crop. Prod. 2017, 107, 609–617. [Google Scholar] [CrossRef]
- Macrae, S.; Gilliland, M.G.; Van Staden, J. Rubber production in guayule: Determination of rubber producing potential. Plant Physiol. 1986, 81, 1027–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Ponciano, G.; Huo, N.; Gu, Y.; Ilut, D.; McMahan, C. RNASeq analysis of drought-stressed guayule reveals the role of gene transcription for modulating rubber, resin, and carbohydrate synthesis. Sci. Rep. 2021, 11, 21610. [Google Scholar] [CrossRef]
- Unland, K.; Pütter, K.M.; Vorwerk, K.; van Deenen, N.; Twyman, R.M.; Prüfer, D.; Gronover, C.S. Functional characterization of squalene synthase and squalene epoxidase in Taraxacum Koksaghyz. Plant Direct 2018, 2, e00063. [Google Scholar] [CrossRef] [PubMed]
- Wentzinger, L.F.; Bach, T.J.; Hartmann, M.-A. Inhibition of squalene synthase and squalene epoxidase in tobacco cells triggers an up-regulation of 3-hydroxy-3-methylglutaryl coenzyme A reductase. Plant Physiol. 2002, 130, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Espenshade, P.J.; Hughes, A.L. Regulation of Sterol Synthesis in Eukaryotes. Annu. Rev. Genet. 2007, 41, 401–427. [Google Scholar] [CrossRef]
- Park, J.; Zielinski, M.; Magder, A.; Tsantrizos, Y.S.; Berghuis, A.M. Human farnesyl pyrophosphate synthase is allosterically inhibited by its own product. Nat. Commun. 2017, 8, 14132. [Google Scholar] [CrossRef] [Green Version]
- Nyati, P.; Rivera-Perez, C.; Noriega, F.G. Negative Feedbacks by Isoprenoids on a Mevalonate Kinase Expressed in the Corpora Allata of Mosquitoes. PLoS ONE 2015, 10, e0143107. [Google Scholar] [CrossRef] [Green Version]
- Dorsey, J.K.; Porter, J.W. The inhibition of melavonic kinases by geranyl and farnesyl pyrophosphates. J. Biol. Chem. 1968, 243, 4667–4670. [Google Scholar] [CrossRef]
- Flint, A.P.F. The activity and kinetic properties of mevalonate kinase in superovulated rat ovary. Biochem. J. 1970, 120, 145–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, V.; Pitera, D.; Withers, S.; Newman, J.D.; Keasling, J.D. Engineering a mevalonate pathway in Escherichia coli for production of terpenoids. Nat. Biotechnol. 2003, 21, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Quan, S.; Xiao, H. Towards efficient terpenoid biosynthesis: Manipulating IPP and DMAPP supply. Bioresour. Bioprocess. 2019, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Hepper, C.M.; Audley, B.G. The biosynthesis of rubber from β-hydroxy-β-methylglutaryl-coenzyme A in Hevea brasiliensis latex. Biochem. J. 1969, 114, 379–386. [Google Scholar] [CrossRef]
- Gray, J.C.; Kekwick, R.G.O. The inhibition of plant mevalonate kinase preparations by prenyl phosphates. Biochim. Biophys. Acta 1972, 279, 290–296. [Google Scholar] [CrossRef]
- Xie, W.; McMahan, C.M.; DeGraw, A.J.; Distefano, M.D.; Cornish, K.; Whalen, M.C.; Shintani, D.K. Initiation of rubber biosynthesis: In vitro comparisons of benzophenone-modified diphosphate analogues in three rubber-producing species. Phytochemistry 2008, 69, 2539–2545. [Google Scholar] [CrossRef] [PubMed]
- Lakusta, A.M.; Kwon, M.; Kwon, E.-J.G.; Stonebloom, S.; Scheller, H.; Ro, D.-K. Molecular studies of the protein complexes involving cis-prenyltransferase in Guayule (Parthenium argentatum), an alternative rubber-producing plant. Front. Plant Sci. 2019, 10, 165. [Google Scholar] [CrossRef]
- Epping, J.; van Deenan, N.; Niephaus, E.; Stolze, A.; Fricke, J.; Huber, C.; Eisenreich, W.; Twyman, R.M.; Prufer, D.; Prüfer, D.; et al. A rubber transferase activator is necessary for natural rubber biosynthesis in dandelion. Nat. Plants 2015, 1, 1–9. [Google Scholar] [CrossRef]
- Collins-Silva, J.; Nural-Taban, A.; Skaggs, A.; Scott, D.; Hathwaik, U.; Woolsey, R.; Schegg, K.; McMahan, C.; Whalen, M.; Cornish, K.; et al. Altered levels of the Taraxacum kok-saghyz (Russian dandelion) small rubber particle protein, TkSRPP3, result in qualitative and quantitative changes in rubber metabolism. Phytochemistry 2012, 79, 46–56. [Google Scholar] [CrossRef]
- Hillebrand, A.; Post, J.J.; Wurbs, D.; Wahler, D.; Lenders, M.; Krzyzanek, V.; Prüfer, D.; Gronover, C.S. Down-regulation of small rubber particle protein expression affects integrity of rubber particles and rubber content in Taraxacum brevicorniculatum. PLoS ONE 2012, 7, e41874. [Google Scholar] [CrossRef] [Green Version]
- Scora, R.W.; Kumamoto, J. Essential leaf oils of Parthenium argentatum A. Gray. J. Agric. Food Chem. 1979, 27, 642–643. [Google Scholar] [CrossRef]
- Kumamoto, J.; Scora, R.W.; Clerx, W.A. Composition of leaf oils in the genus Parthenium, L. compositae. J. Agric. Food Chem. 1985, 33, 650–652. [Google Scholar] [CrossRef]
- Spano, N.; Meloni, P.; Idda, I.; Mariani, A.; Pilo, I.M.; Nurchi, M.V.; Lachowicz, I.J.; Rivera, E.; Orona-Espino, A.; Sanna, G. Assessment, validation and application to real samples of an RP-HPLC method for the determination of guayulins a, B, C and d in Guayule Shrub. Separations 2018, 5, 23. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Concepción, M.; Boronat, A. Breaking new ground in the regulation of the early steps of plant isoprenoid biosynthesis. Curr. Opin. Plant Biol. 2006, 25, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Bick, J.A.; Lange, B.M. Metabolic cross talk between cytosolic and plastidial pathways of isoprenoid biosynthesis: Unidirectional transport of intermediates across the chloroplast envelope membrane. Arch. Biochem. Biophys. 2003, 415, 146–154. [Google Scholar] [CrossRef]
- Flügge, U.-I.; Gao, W. Transport of isoprenoid intermediates across chloroplast envelope membranes. Plant Biol. 2005, 7, 91–97. [Google Scholar] [CrossRef]
- Laule, O.; Fürholz, A.; Chang, H.S.; Zhu, T.; Wang, X.; Heifetz, P.B.; Gruissem, W.; Lange, M. Crosstalk between cytosolic and plastidial pathways of isoprenoid biosynthesis in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2003, 100, 6866–6871. [Google Scholar] [CrossRef] [Green Version]
- Hajdukiewicz, P.; Svab, Z.; Maliga, P. The small, versatile pPZP family of Agrobacterium binary vectors for plant transformation. Plant Mol. Biol. 1994, 25, 989–994. [Google Scholar] [CrossRef]
- Rockhold, D.R.; Chang, S.; Taylor, N.; Allen, P.V.; McCue, K.F.; Belknap, W.R. Structure of Two Solanum bulbocastanum Polyubiquitin Genes and Expression of Their Promoters in Transgenic Potatoes. Am. J. Potato Res. 2008, 85, 219–226. [Google Scholar] [CrossRef]
- Herrera-Estrella, L.; De Block, M.; Messens, E.; Hernalsteens, J.-P.; Van Montagu, M.; Schell, J. Chimeric genes as dominant selectable markers in plant cells. Embo J. 1983, 2, 987–995. [Google Scholar] [CrossRef]
- De Greve, H.; Dhaese, P.; Seurinck, J. Nucleotide sequence and transcript map of the Agrobacterium tumefaciens Ti plasmid-encoded octopine synthase gene. J. Mol. Appl. Genet. 1982, 1, 499–511. [Google Scholar]
- Xiao, K.; Zhang, C.; Harrison, M.; Wang, Z.-Y. Isolation and characterization of a novel plant promoter that directs strong constitutive expression of transgenes in plants. Mol. Breed. 2005, 15, 221–231. [Google Scholar] [CrossRef]
- Jefferson, A.R. Assaying the chimeric genes in plants: The GUS gene fusion system. Plant Mol. Biol. Rep. 1987, 5, 387–405. [Google Scholar] [CrossRef]
- Garbarino, J.E.; Belknap, W.R. Isolation of a ubiquitin-ribosomal protein gene (ubi3) from potato and expression of its promoter in transgenic plants. Plant Mol. Biol. 1994, 24, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Christensen, A.H.; Quail, P.H. Ubiquitin promoter-based vectors for high-level expression of selectable and/or screenable marker genes in monocotyledonous plants. Transgenic Res. 1996, 5, 213–218. [Google Scholar] [CrossRef]
- Hood, E.E.; Helmer, G.L.; Fraley, R.T.; Chilton, M.D. The hypervirulence of Agrobacterium tumefaciens A281 is encoded in a region of pTiBo542 outside of T=DNA. J. Bacteriol. 1986, 168, 1291–1301. [Google Scholar] [CrossRef] [Green Version]
- Ray, D.T.; Dierig, D.A.; Thompson, A.E.; Coffelt, T.A. Registration of six guayule germplasms with high yielding ability. Crop Sci. 1999, 39, 300–301. [Google Scholar] [CrossRef]
- Dong, N.; Montanez, B.; Creelman, R.A.; Cornish, K. Low light and low ammonium are key factors for guayule leaf tissue shoot organogenesis and transformation. Plant Cell Rep. 2006, 25, 26–34. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plantarum. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Placido, D.; Dong, N.; Dong, C.; Cruz, V.M.V.; Dierig, D.A.; Cahoon, R.E.; Kang, B.; Huynh, T.; Whalen, M.; Ponciano, G.; et al. Downregulation of a CYP74 rubber particle protein increases natural rubber production in Parthenium argentatum. Front. Plant Sci. 2019, 760, 1–18. [Google Scholar] [CrossRef]
- Baidoo, E.E.K.; Wang, G.; Joshua, C.; Teixeira Benites, V.; Keasling, J.D. Liquid Chromatography and Mass Spectrometry Analysis of Isoprenoid Intermediates in Escherichia coli. In Microbial Metabolomics: Methods and Protocols. Methods in Molecular Biology; Baidoo, E.E.K., Ed.; Humana Press Inc.: New York, NY, USA, 2019; pp. 209–224. [Google Scholar] [CrossRef]
- Kim, J.; Baidoo, E.E.K.; Amer, B.; Mukhopadhyay, A.; Adams, P.D.; Simmons, B.A.; Lee, T.S. Engineering Saccharomyces cerevisiae for isoprenol production. Metab. Eng. 2021, 64, 154–166. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Placido, D.; Dong, N.; Amer, B.; Dong, C.; Ponciano, G.; Kahlon, T.; Whalen, M.; Baidoo, E.E.K.; McMahan, C. Downregulation of Squalene Synthase Broadly Impacts Isoprenoid Biosynthesis in Guayule. Metabolites 2022, 12, 303. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo12040303
Placido D, Dong N, Amer B, Dong C, Ponciano G, Kahlon T, Whalen M, Baidoo EEK, McMahan C. Downregulation of Squalene Synthase Broadly Impacts Isoprenoid Biosynthesis in Guayule. Metabolites. 2022; 12(4):303. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo12040303
Chicago/Turabian StylePlacido, Dante, Niu Dong, Bashar Amer, Chen Dong, Grisel Ponciano, Talwinder Kahlon, Maureen Whalen, Edward E. K. Baidoo, and Colleen McMahan. 2022. "Downregulation of Squalene Synthase Broadly Impacts Isoprenoid Biosynthesis in Guayule" Metabolites 12, no. 4: 303. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo12040303