Analysis of Bacteriophages with Insulator-Based Dielectrophoresis


 and
and 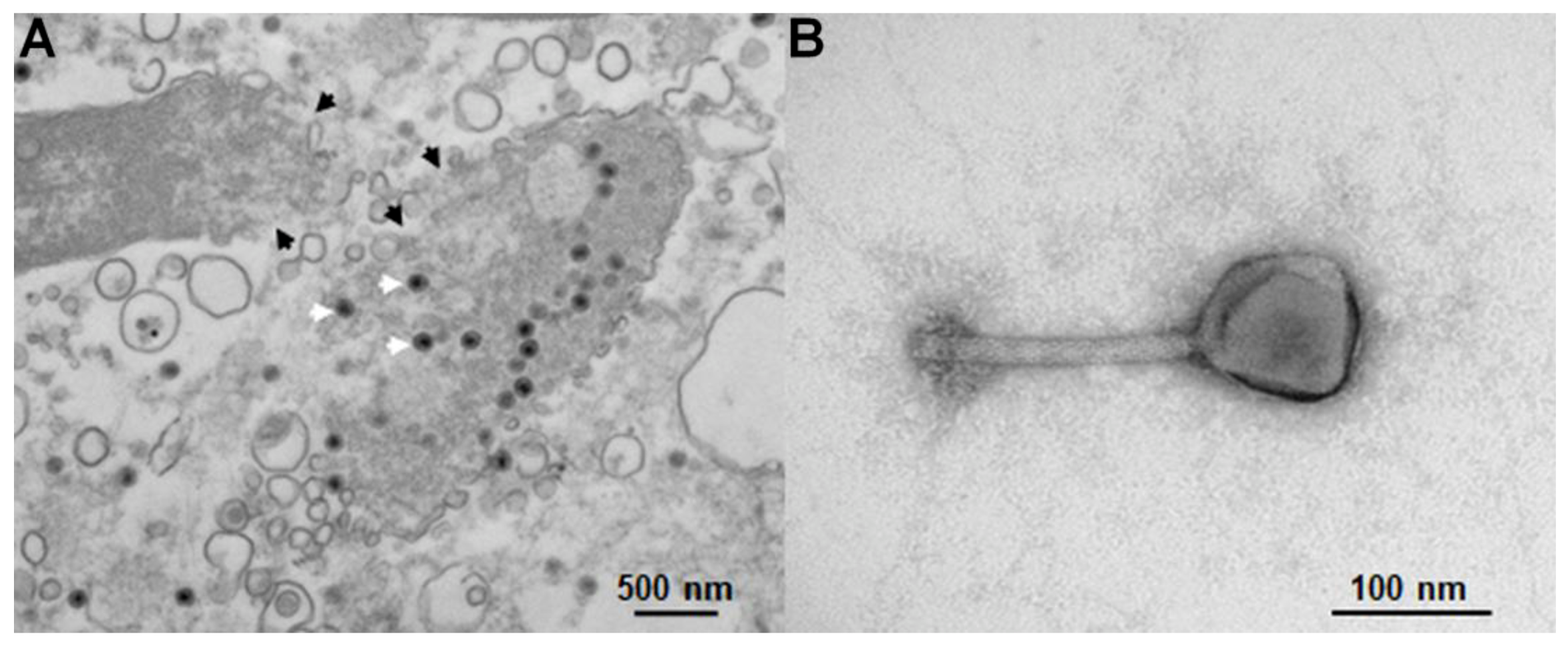{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Theory
3. Materials and Methods
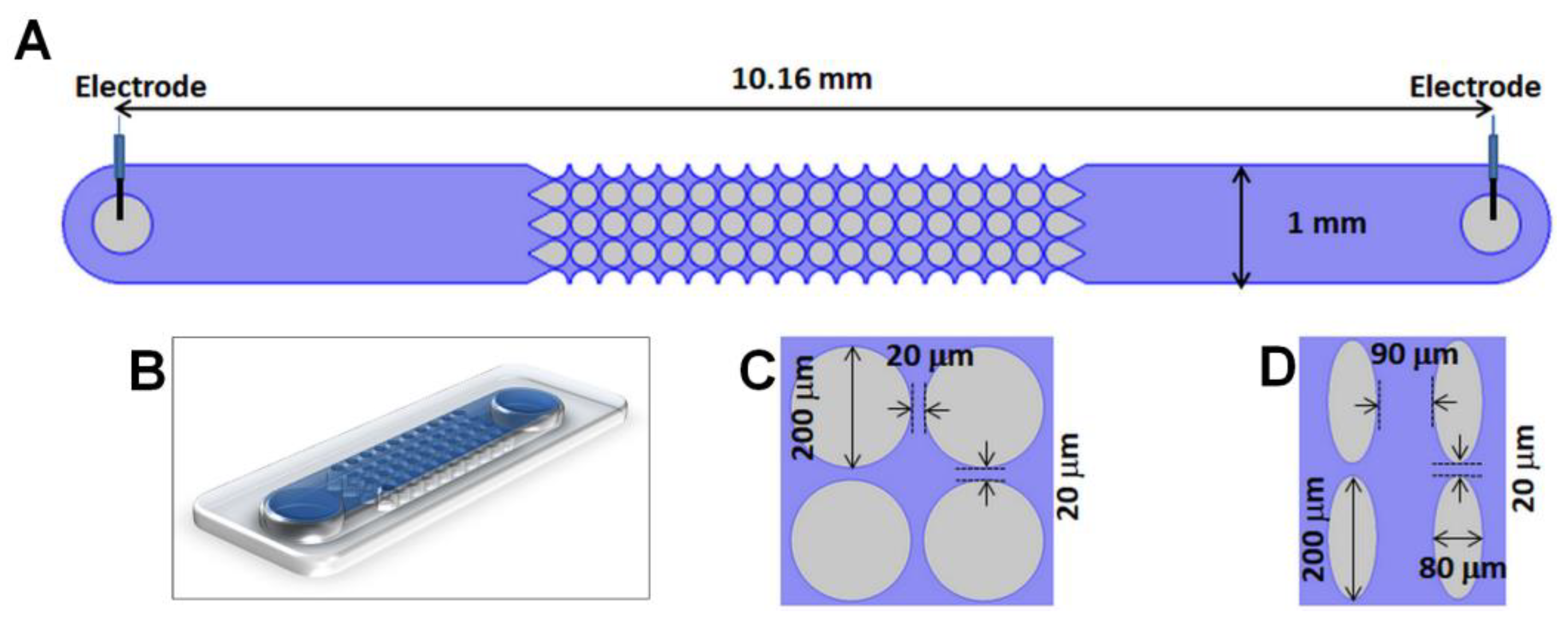
3.1. Microdevices, Viral Samples and Suspending Medium
3.2. Equipment and Experimental Procedure
4. Results and Discussion
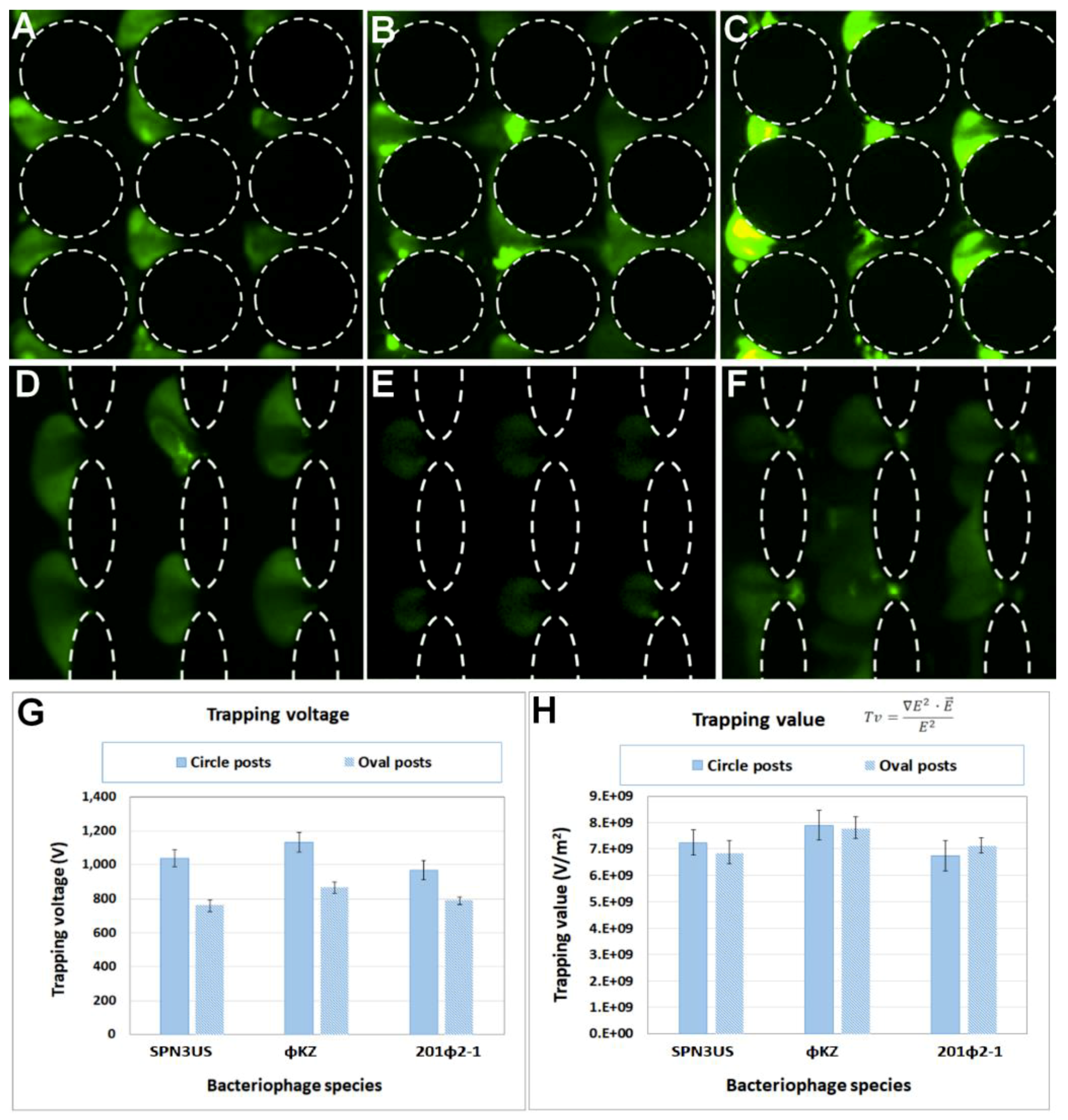
4.1. Experimental Characterization of the Dielectrophoretic Trapping of Phage Virions
4.2. Modeling Predictions for the Trapping of Phage Virus
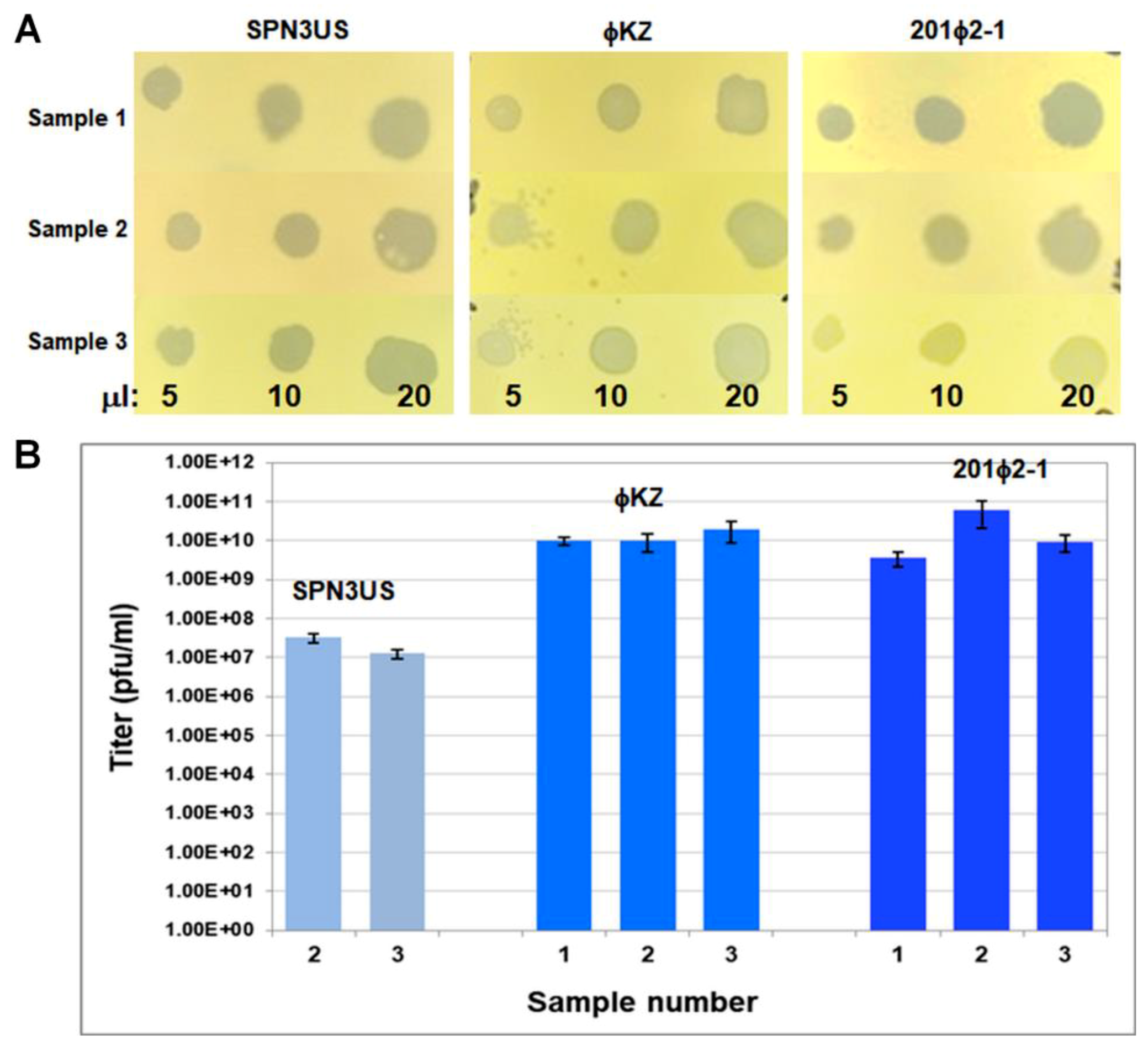
4.3. Viability Assessments after Dielectrophoretic Trapping
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Comeau, A.M.; Hatfull, G.F.; Krisch, H.M.; Lindell, D.; Mann, N.H.; Prangishvili, D. Exploring the prokaryotic virosphere. Res. Microbiol. 2008, 159, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Keen, E.C. A century of phage research: Bacteriophages and the shaping of modern biology. BioEssays 2015, 37, 6–9. [Google Scholar] [CrossRef]
- Nasukawa, T.; Uchiyama, J.; Taharaguchi, S.; Ota, S.; Ujihara, T.; Matsuzaki, S.; Murakami, H.; Mizukami, K.; Sakaguchi, M. Virus purification by CsCl density gradient using general centrifugation. Arch. Virol. 2017, 162, 3523–3528. [Google Scholar] [CrossRef]
- Thomas, J.A.; Hardies, S.C.; Rolando, M.; Hayes, S.J.; Lieman, K.; Carroll, C.A.; Weintraub, S.T.; Serwer, P. Complete genomic sequence and mass spectrometric analysis of highly diverse, atypical Bacillus thuringiensis phage 0305phi8-36. Virology 2007, 368, 405–421. [Google Scholar] [CrossRef] [PubMed]
- Bourdin, G.; Schmitt, B.; Marvin Guy, L.; Germond, J.E.; Zuber, S.; Michot, L.; Reuteler, G.; Brüssow, H. Amplification and purification of T4-like Escherichia coli phages for phage therapy: From laboratory to pilot scale. Appl. Environ. Microbiol. 2014, 80, 1469–1476. [Google Scholar] [CrossRef]
- Castro-Mejía, J.L.; Muhammed, M.K.; Kot, W.; Neve, H.; Franz, C.M.; Hansen, L.H.; Vogensen, F.K.; Nielsen, D.S. Optimizing protocols for extraction of bacteriophages prior to metagenomic analyses of phage communities in the human gut. Microbiome 2015, 3, 64. [Google Scholar] [CrossRef]
- Bonilla, N.; Rojas, M.I.; Cruz, G.N.F.; Hung, S.H.; Rohwer, F.; Barr, J.J. Phage on tap-a quick and efficient protocol for the preparation of bacteriophage laboratory stocks. PeerJ 2016, 4, e2261. [Google Scholar] [CrossRef]
- Masuda, T.; Maruyama, H.; Honda, A.; Arai, F. Virus enrichment for single virus infection by using 3d insulator based dielectrophoresis. PLoS ONE 2014, 9, e94083. [Google Scholar] [CrossRef] [PubMed]
- Whitesides, G.M. The origins and the future of microfluidics. Nature 2006, 442, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Nakano, A.; Camacho-Alanis, F.; Ros, A. Insulator-based dielectrophoresis with [small beta]-galactosidase in nanostructured devices. Analyst 2015, 140, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.V.; Salmon, G.L.; Ros, A. Continuous separation of DNA molecules by size using insulator-based dielectrophoresis. Anal. Chem. 2017, 89, 1531–1539. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Lawrence, R.M.; Jones, P.V.; Hogue, B.G.; Hayes, M.A. Concentration of sindbis virus with optimized gradient insulator-based dielectrophoresis. Analyst 2016, 141, 1997–2008. [Google Scholar] [CrossRef] [PubMed]
- LaLonde, A.; Romero-Creel, M.F.; Lapizco-Encinas, B.H. Assessment of cell viability after manipulation with insulator-based dielectrophoresis. Electrophoresis 2015, 36, 1479–1484. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.H.; Tsegaye, M.; Varhue, W.; Liao, K.T.; Abebe, L.S.; Smith, J.A.; Guerrant, R.L.; Swami, N.S. Quantitative dielectrophoretic tracking for characterization and separation of persistent subpopulations of cryptosporidium parvum. Analyst 2014, 139, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Romero-Creel, M.; Goodrich, E.; Polniak, D.; Lapizco-Encinas, B. Assessment of sub-micron particles by exploiting charge differences with dielectrophoresis. Micromachines 2017, 8, 239. [Google Scholar] [CrossRef] [PubMed]
- Polniak, D.V.; Goodrich, E.; Hill, N.; Lapizco-Encinas, B.H. Separating large microscale particles by exploiting charge differences with dielectrophoresis. J. Chromatogr. A 2018, 1545, 84–92. [Google Scholar] [CrossRef]
- Gallo-Villanueva, R.C.; Perez-Gonzalez, V.H.; Cardenas-Benitez, B.; Jind, B.; Martinez-Chapa, S.O.; Lapizco-Encinas, B.H. Joule heating effects in optimized insulator-based dielectrophoretic devices: An interplay between post geometry and temperature rise. Electrophoresis 2019, 40, 1408–1416. [Google Scholar] [CrossRef]
- Jesús-Pérez, N.M.; Lapizco-Encinas, B.H. Dielectrophoretic monitoring of microorganisms in environmental applications. Electrophoresis 2011, 32, 2331–2357. [Google Scholar] [CrossRef]
- Grom, F.; Kentsch, J.; Müller, T.; Schnelle, T.; Stelzle, M. Accumulation and trapping of hepatitis a virus particles by electrohydrodynamic flow and dielectrophoresis. Electrophoresis 2006, 27, 1386–1393. [Google Scholar] [CrossRef]
- Schnelle, T.; Muller, T.; Fiedler, S.; Shirley, S.G.; Ludwig, K.; Herrmann, A.; Fuhr, G.; Wagner, B.; Zimmermann, U. Trapping of viruses in high-frequency electric field cages. Naturwissenschaften 1996, 83, 172–176. [Google Scholar] [CrossRef]
- Müller, T.; Fiedler, S.; Schnelle, T.; Ludwig, K.; Jung, H.; Fuhr, G. High frequency electric fields for trapping of viruses. Biotechnol. Tech. 1996, 10, 221–226. [Google Scholar] [CrossRef]
- Hughes, M.P.; Morgan, H.; Rixon, F.J. Characterization of herpes simplex virus particles by dielectrophoretic crossover methods. In Proceedings of the 20th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Hong Kong, China, 1 November 1998; pp. 2816–2819. [Google Scholar]
- Hughes, M.P.; Morgan, H.; Rixon, F.J.; Burt, J.P.H.; Pethig, R. Manipulation of herpes simplex virus type 1 by dielectrophoresis. Biochim. Biophys. Acta (BBA) Gen. Subj. 1998, 1425, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Hughes, M.P.; Morgan, H.; Rixon, F.J. Measuring the dielectric properties of herpes simplex virus type 1 virions with dielectrophoresis. Biochim. Biophys. Acta (BBA) Gen. Subj. 2002, 1571, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Akin, D.; Li, H.; Bashir, R. Real-time virus trapping and fluorescent imaging in micro-fluidic devices. Nano Letters. 2004, 4, 257–259. [Google Scholar] [CrossRef]
- Prakash, R.; Pabbaraju, K.; Wong, S.; Wong, A.; Tellier, R.; Kaler, K. Multiplex, quantitative, reverse transcription pcr detection of influenza viruses using droplet microfluidic technology. Micromachines 2015, 6, 63. [Google Scholar] [CrossRef]
- Singh, R.; Sharma, A.; Hong, S.; Jang, J. Electrical immunosensor based on dielectrophoretically-deposited carbon nanotubes for detection of influenza virus h1n1. Analyst 2014, 139, 5415–5421. [Google Scholar] [CrossRef] [PubMed]
- Madiyar, F.R.; Haller, S.L.; Farooq, O.; Rothenburg, S.; Culbertson, C.; Li, J. Ac dielectrophoretic manipulation and electroporation of vaccinia virus using carbon nanoelectrode arrays. Electrophoresis 2017, 38, 1515–1525. [Google Scholar] [CrossRef]
- Morgan, H.; Green, N. Dielectrophoretic manipulation of rod-shaped viral particles. J. Electrost. 1997, 42, 279–293. [Google Scholar] [CrossRef] [Green Version]
- Ermolina, I.; Milner, J.; Morgan, H. Dielectrophoretic investigation of plant virus particles: Cow pea mosaic virus and tobacco mosaic virus. Electrophoresis 2006, 27, 3939–3948. [Google Scholar] [CrossRef]
- Ermolina, I.; Morgan, H.; Green, N.G.; Milner, J.J.; Feldman, Y. Dielectric spectroscopy of tobacco mosaic virus. Biochim. Biophys. Acta (BBA) Gen. Subj. 2003, 1622, 57–63. [Google Scholar] [CrossRef]
- Lapizco-Encinas, B.H.; Davalos, R.; Simmons, B.A.; Cummings, E.B.; Fintschenko, Y. An insulator-based (electrodeless) dielectrophoretic concentrator for microbes in water. J. Microbiol. Methods 2005, 62, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Sonnenberg, A.; Marciniak, J.Y.; McCanna, J.; Krishnan, R.; Rassenti, L.; Kipps, T.J.; Heller, M.J. Dielectrophoretic isolation and detection of cfc-DNA nanoparticulate biomarkers and virus from blood. Electrophoresis 2013, 34, 1076–1084. [Google Scholar] [CrossRef] [PubMed]
- Madiyar, F.R.; Syed, L.U.; Culbertson, C.T.; Li, J. Manipulation of bacteriophages with dielectrophoresis on carbon nanofiber nanoelectrode arrays. Electrophoresis 2013, 34, 1123–1130. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Zare, M.J.; Madadi, H.; Sellarès, J.; Casals-Terré, J. A new approach to design an efficient micropost array for enhanced direct-current insulator-based dielectrophoretic trapping. Anal. Bioanal. Chem. 2016, 408, 5285–5294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowther, C.V.; Hayes, M.A. Refinement of insulator-based dielectrophoresis. Analyst 2017, 142, 1608–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, T.B. Electromechanics of Particles; Cambridge University Press: New York, NY, USA, 1995; p. 265. [Google Scholar]
- Gencoglu, A.; Olney, D.; LaLonde, A.; Koppula, K.S.; Lapizco-Encinas, B.H. Dynamic microparticle manipulation with an electroosmotic flow gradient with low frequency alternating current dielectrophoresis. Electrophoresis 2014, 35, 363–373. [Google Scholar] [CrossRef]
- Saucedo-Espinosa, M.A.; Lapizco-Encinas, B.H. Experimental and theoretical study of dielectrophoretic particle trapping in arrays of insulating structures: Effect of particle size and shape. Electrophoresis 2015, 36, 1086–1097. [Google Scholar] [CrossRef] [PubMed]
- Saucedo-Espinosa, M.A.; Lapizco-Encinas, B.H. Design of insulator-based dielectrophoretic devices: Effect of insulator posts characteristics. J. Chromatogr. A 2015, 1422, 325–333. [Google Scholar] [CrossRef]
- Lee, J.H.; Shin, H.; Kim, H.; Ryu, S. Complete genome sequence of salmonella bacteriophage spn3us. J. Virol. 2011, 85, 13470–13471. [Google Scholar] [CrossRef]
- Mesyanzhinov, V.V.; Robben, J.; Grymonprez, B.; Kostyuchenko, V.A.; Bourkaltseva, M.V.; Sykilinda, N.N.; Krylov, V.N.; Volckaert, G. The genome of bacteriophage φKZ of Pseudomonas aeruginosa. J. Mol. Biol. 2002, 317, 1–19. [Google Scholar] [CrossRef]
- Thomas, J.A.; Rolando, M.R.; Carroll, C.A.; Shen, P.S.; Belnap, D.M.; Weintraub, S.T.; Serwer, P.; Hardies, S.C. Characterization of Pseudomonas chlororaphis myovirus 201phi2-1 via genomic sequencing, mass spectrometry, and electron microscopy. Virology 2008, 376, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.P.; Morgan, H.; Flynn, M.F. The dielectrophoretic behavior of submicron latex spheres: Influence of surface conductance. J. Colloid Interface Sci. 1999, 220, 454–457. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.P.; Morgan, H.; Rixon, F.J. Dielectrophoretic manipulation and characterization of herpes simplex virus-1 capsids. Eur. Biophys. J. 2001, 30, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.A.; Benítez Quintana, A.D.; Bosch, M.A.; Coll De Peña, A.; Aguilera, E.; Coulibaly, A.; Wu, W.; Osier, M.V.; Hudson, A.O.; Weintraub, S.T.; et al. Identification of essential genes in the Salmonella phage SPN3US reveals novel insights into giant phage head structure and assembly. J. Virol. 2016, 90, 10284–10298. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coll De Peña, A.; Mohd Redzuan, N.H.; Abajorga, M.K.; Hill, N.; Thomas, J.A.; Lapizco-Encinas, B.H. Analysis of Bacteriophages with Insulator-Based Dielectrophoresis. Micromachines 2019, 10, 450. https://0-doi-org.brum.beds.ac.uk/10.3390/mi10070450
Coll De Peña A, Mohd Redzuan NH, Abajorga MK, Hill N, Thomas JA, Lapizco-Encinas BH. Analysis of Bacteriophages with Insulator-Based Dielectrophoresis. Micromachines. 2019; 10(7):450. https://0-doi-org.brum.beds.ac.uk/10.3390/mi10070450
Chicago/Turabian StyleColl De Peña, Adriana, Nurul Humaira Mohd Redzuan, Milky K. Abajorga, Nicole Hill, Julie A. Thomas, and Blanca H. Lapizco-Encinas. 2019. "Analysis of Bacteriophages with Insulator-Based Dielectrophoresis" Micromachines 10, no. 7: 450. https://0-doi-org.brum.beds.ac.uk/10.3390/mi10070450