
New Insights on Plasmin Long Term Stability and the Mechanism of Its Activity Inhibition Analyzed by Quartz Crystal Microbalance


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Preparation of β-Casein Layers and RQCM Measurements
2.3. The Concept of the Analysis of Protease Inhibition
3. Results and Discussion
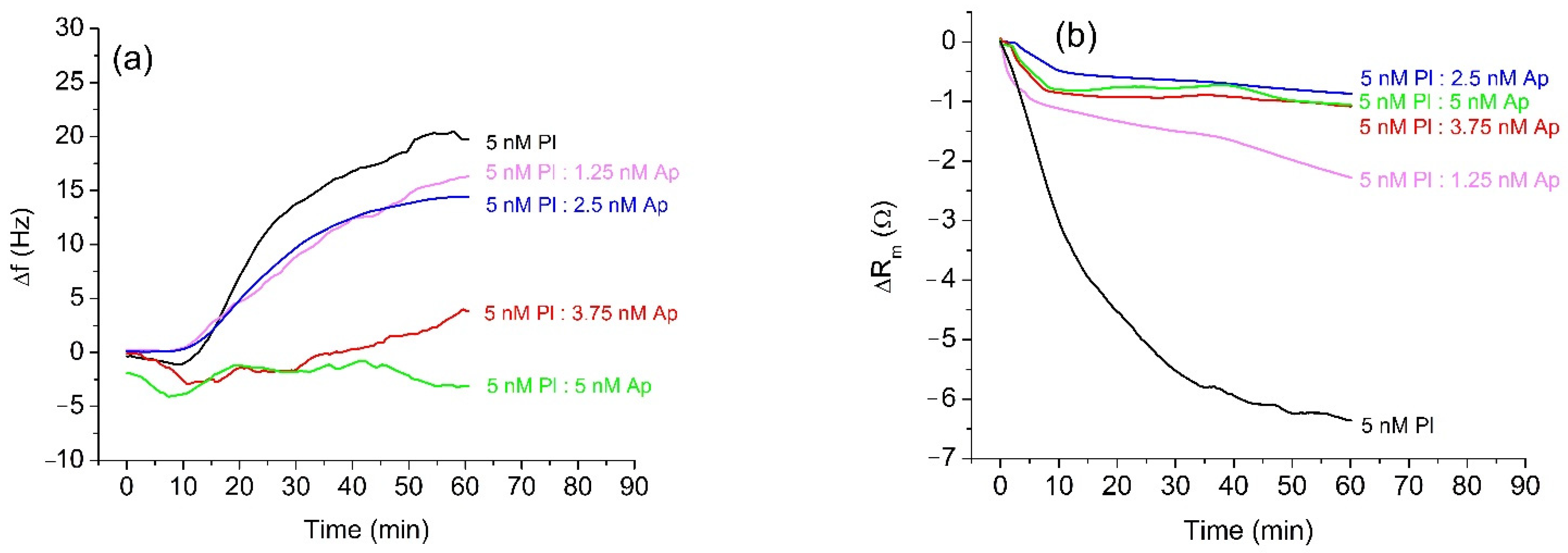
3.1. Inhibition of Trypsin and Plasmin by α2-Antiplasmin
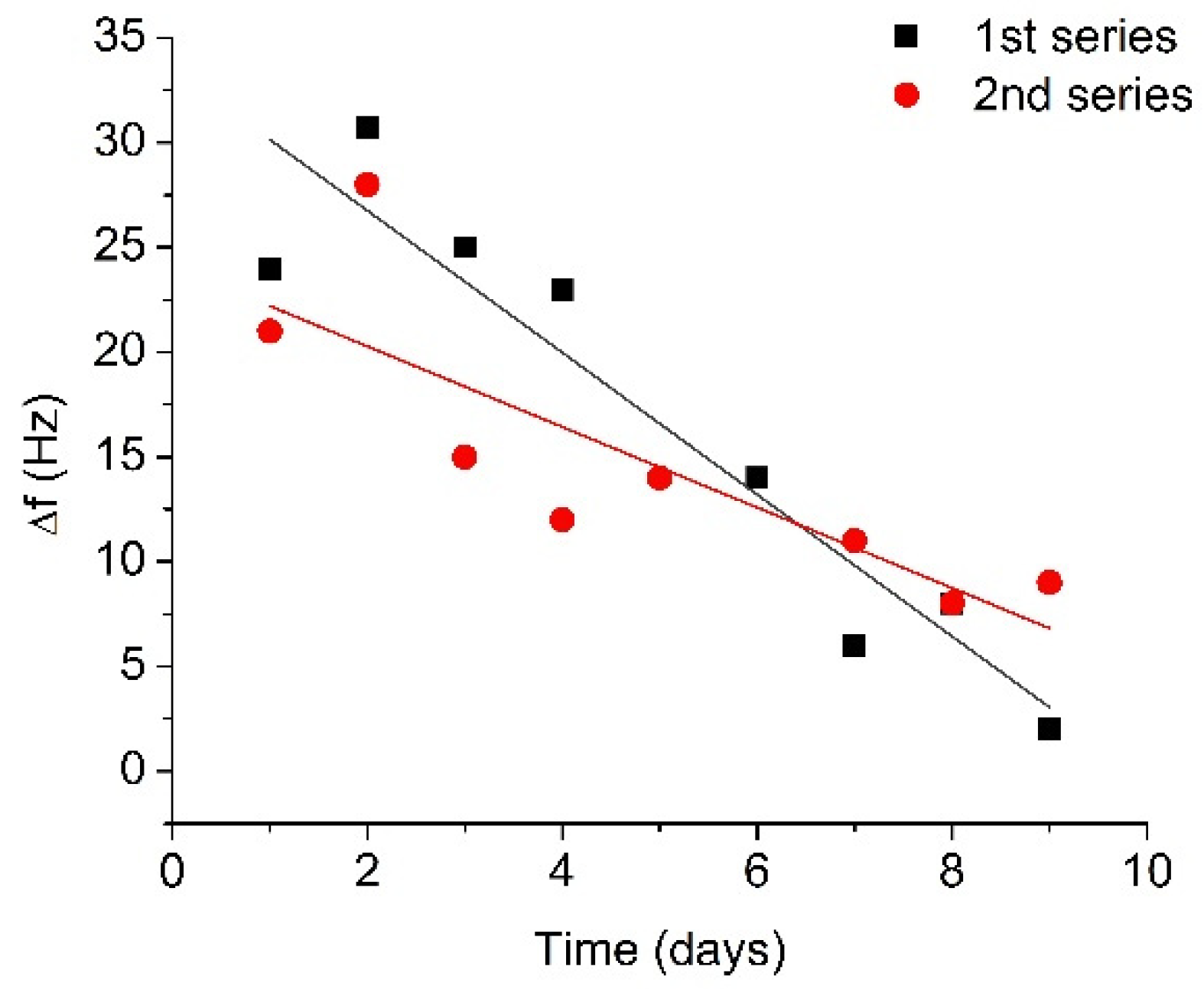
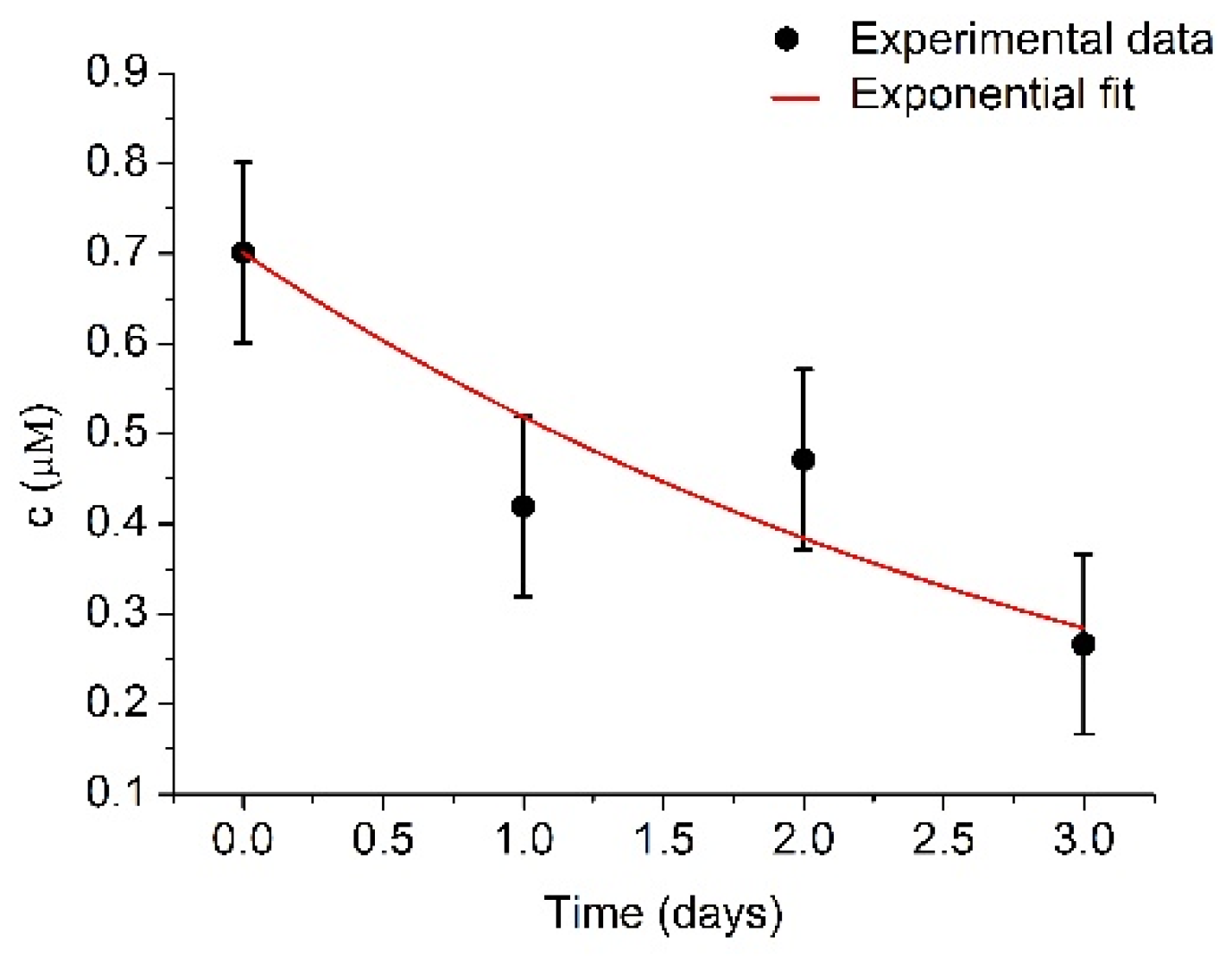
3.2. Monitoring of the Plasmin Autolysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lu, M.; Shiau, Y.; Wong, J.; Lin, R.; Kravis, H.; Blackmon, T.; Pakzad, T.; Jen, T.; Cheng, A.; Chang, J.; et al. Milk spoilage: Methods and practices of detecting milk. Food Nutr. Sci. 2013, 4, 113–123. [Google Scholar] [CrossRef] [Green Version]
- Holt, C.; Carver, J.A.; Ecroyd, H.; Thorn, D.C. Caseins and the casein micelle: Their biological functions, structures, and behavior in foods. J. Dairy Sci. 2013, 96, 6127–6146. [Google Scholar] [CrossRef] [PubMed]
- Creamer, L.K.; Plowman, J.E.; Liddell, M.J.; Smith, M.H.; Hill, J.P. Micelle stability: Kappa-casein structure and function. J. Dairy Sci. 1998, 81, 3004–3012. [Google Scholar] [CrossRef]
- Smith, T.J.; Campbell, R.E.; Jo, Y.; Drake, M.A. Flavor and stability of milk proteins. J. Dairy Sci. 2016, 99, 4325–4346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavan, R.S.; Chavan, S.R.; Khedkar, C.D.; Jana, A.H. UHT milk processing and effect of plasmin activity on shelf life: A review. Compr. Rev. Food Sci. Food Saf. 2011, 10, 251–268. [Google Scholar] [CrossRef]
- Prado, B.M.; Sombers, S.E.; Ismail, B.; Hayes, K.D. Effect of heat treatment on the activity of inhibitors of plasmin and plasminogen activators in milk. Food Sci. Nutr. 2006, 16, 593–599. [Google Scholar] [CrossRef]
- Rauh, V.M.; Johansen, L.B.; Ipsen, R.; Paulsson, M.; Larsem, L.B.; Hammershøj, M. Plasmin activity in UHT milk: Relationship between proteolysis, age, and bitterness. J. Agric. Good Chem. 2014, 62, 6852–6860. [Google Scholar] [CrossRef]
- Fantuz, F.; Polidori, P.; Lebboroni, G.; Polidori, F. Characterization of plasmin, plasminogen and plasminogen activator in goat milk. Ital. J. Anim. Sci. 2003, 2, 554–556. [Google Scholar]
- Abdul, S.; Leebeek, F.W.G.; Rijken, D.C.; de Willige, S.U. Natural heterogeneity of α2-antiplasmin: Functional and clinical consequences. Blood 2016, 127, 538–545. [Google Scholar] [CrossRef] [Green Version]
- Rippel, K.M.; Nielsen, S.S.; Hayes, K.D. Effects of native and denatured whey proteins on plasminogen activator activity. J. Dairy Sci. 2004, 87, 2344–2350. [Google Scholar] [CrossRef] [Green Version]
- Monroy, V.; Ruiz, B.H. Participation of the Dengue virus in the fibrinolytic process. Virus Genes 2000, 21, 197–208. [Google Scholar] [CrossRef]
- Dacres, H.; Wang, J.; Anderson, A.; Trowell, S.C. A rapid and sensitive biosensor for measuring plasmin activity in milk. Sens. Act. B. 2019, 301, 127–141. [Google Scholar] [CrossRef]
- Precetti, A.S.; Oria, M.P.; Nielsen, S.S. Presence in bovine milk of two protease inhibitors of the plasmin system. J. Dairy Sci. 1997, 80, 1490–1496. [Google Scholar] [CrossRef]
- Dalhborn, M.; Ahlborg, A.; Sodenborg, B.; Virgin, J.; Wilczek, B. Gamma camera detection of 99mTc-plasmin in the diagnosis of deep-vein thrombosis. Eur. J. Nuclear Med. 1984, 9, 455–501. [Google Scholar] [CrossRef]
- Turner, R.B.; Liu, L.; Szanova, I.Y.; Reed, G.L. Structural elements that govern the substrate specificity of the clot-dissolving enzyme plasmin. J. Biol. Chem. 2002, 277, 33068–33074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, P.H.; Dziobkowski, C.T.; Englert, M.E. A sensitive fluorimetric assay for plasminogen, plasmin and stopkinase. Anal. Biochem. 1974, 61, 200–208. [Google Scholar] [CrossRef]
- Tervo, T.; Honkanen, N.; van Setten, G.B.; Virtanen, T.; Tarkkanen, A.; Harkonen, M. A rapid fluorometric assay for tear fluid plasmin activity. Cornea 1994, 13, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, T.; Konttinen, Y.T.; Honkanen, N.; Harkonen, M.; Tervo, T. Tear fluid plasmin activity of dry eye patients with Sjogren’s syndrome. Acta Ophthalmol. Scand. 1997, 75, 137–141. [Google Scholar] [CrossRef]
- Yazgan, N.N.; Bulat, T.; Topcu, A.; Dudak, F.C.; Boyaci, I.H.; Tamer, U. Surface-enhanced Raman scattering-based detection of plasmin activity by specific peptide substrate. Food Chem. 2022, 372, 131235. [Google Scholar] [CrossRef]
- Chinnappana, R.; Attas, S.A.; Kamanc, W.E.; Bikker, F.J.; Zourob, M. Development of magnetic nanoparticle based calorimetric assay for the detection of bovine mastitis in cow milk. Anal. Biochem. 2017, 523, 58–64. [Google Scholar] [CrossRef]
- McVey, C.; Logan, N.; Thanh, N.T.K.; Elliott, C.; Cao, C. Unusual switchable peroxidase-mimicking nanozyme for the determination of proteolytic biomarker. Nano Res. 2019, 12, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Chang, B.; Lei, H.; Liu, H.; Liu, C.; Wu, H.; Yeh, T. Antibodies against dengue virus E protein peptide bind to human plasminogen and inhibit plasmin activity. Clin. Exp. Immunol. 1997, 110, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuka, K.; Maekawa, I.; Waki, M.; Takenaka, S. Electrochemical assay of plasmin activity and its kinetic analysis. Anal. Biochem. 2009, 385, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Castillo, G.; Pribransky, K.; Mező, G.; Kocsis, L.; Csámpai, A.; Németh, K.; Keresztes, Z.; Hianik, T. Electrochemical and photometric detection of plasmin by specific peptide substrate. Electroanalysis 2015, 27, 789–798. [Google Scholar] [CrossRef]
- Liu, D.; Xiao, X.; Tang, Z.; Chen, Q.; Li, H.; Wang, X.; Yan, Y. Method for qcm resonator device equivalent circuit parameter extraction and electrode quality assessment. Micromachines 2021, 12, 1086. [Google Scholar] [CrossRef]
- Ghazali, M.; Hayward, G.L. Acoustic determination of performance and equivalence of plasminogen activators. Anal. Bioanal. Chem. 2008, 392, 897–902. [Google Scholar] [CrossRef]
- Poturnayova, A.; Karpisova, I.; Castillo, G.; Mezo, G.; Kocsis, L.; Csápai, A.; Keresztes, Z.; Hianik, T. Detection of plasmin based on specific peptide substrate using acoustic transducer. Sens. Act. B. 2015, 223, 591–598. [Google Scholar] [CrossRef]
- Tatarko, M.; Muckley, E.; Šubjaková, V.; Goswami, M.; Sumpter, B.; Hianik, T.; Ivanov, I.N. Machine learning enabled acoustic detection of subnanomolar concentration of trypsin and plasmin in solution. Sens. Actuat. B. 2018, 272, 282–288. [Google Scholar] [CrossRef]
- Románszki, L.; Tatarko, M.; Jiao, M.; Keresztes, Z.; Hianik, T.; Thompson, M. Casein probe-based fast plasmin determination in the picomolar range by an ultra-high frequency acoustic wave biosensor. Sens. Act. B. 2018, 275, 206–214. [Google Scholar] [CrossRef]
- Poturnayova, A.; Szabo, K.; Tatarko, M.; Hucker, A.; Kocsis, R.; Hianik, T. Determination of plasmin in milk using QCM and ELISA methods. Food Control 2021, 123, 107774. [Google Scholar] [CrossRef]
- Melikishvili, S.; Dizon, M.; Hianik, T. Application of high-resolution ultrasonic spectroscopy for real-time monitoring of trypsin activity in β-casein solution. Food Chem. 2021, 337, 127759. [Google Scholar] [CrossRef] [PubMed]
- Dizon, M.; Tatarko, M.; Szabo, K.; Hianik, T. Application of high-resolution ultrasonic spectroscopy for detection of the plasmin activity toward β-casein. Food Chem. 2021, 353, 129373. [Google Scholar] [CrossRef]
- Jatwani, M.; Swain, S. Is small scale dairy farming dying out? An in-depth study. Indian, J. Community Med. 2020, 45, 47–51. [Google Scholar] [CrossRef]
- Melikishvili, S.; Piovarci, I.; Hianik, T. Advances in colorimetric assay based on AuNPs modified by proteins and nucleic acid aptamers. Chemosensors 2021, 9, 281. [Google Scholar] [CrossRef]
- Piovarci, I.; Melikishvili, S.; Tatarko, M.; Hianik, T.; Thompson, M. Detection of sub-nanomolar concentration of trypsin by thickness-shear mode acoustic biosensor and spectrophotometry. Biosensors 2021, 11, 117. [Google Scholar] [CrossRef] [PubMed]
- Parsajoo, C.; Kaufmann, J.M.; Elkaoutit, M. 9—Biosensors for drug testing and discovery. In Biosensors for Medical Applications, 1st ed.; Higson, S., Ed.; Woodhead Publishing: Sawston, UK, 2012; pp. 233–262. [Google Scholar] [CrossRef]
- Newton, M.I.; Atherton, S.; Morris, R.H.; Stanley, S.M.; Evans, C.R.; Hughes, D.C.; McHale, G. Low-cost QCM sensor system for screening semen samples. J. Sensors 2010, 2010, 326365. [Google Scholar] [CrossRef]
- Tatarko, M.; Spagnolo, S.; Oravczová, V.; Süle, J.; Hun, M.; Hucker, A.; Hianik, T. Changes of viscoelastic properties of aptamer-based sensing layers following interaction with List. Innocua. Sens. 2021, 21, 5585. [Google Scholar] [CrossRef]
- Kolev, K.; Léránt, I.; Tenekejiev, K.; Machovich, R. Regulation of fibrinolytic activity of neutrophil leukocyte elastase, plasmin, and miniplasmin by plasma protease inhibitors. J. Biol. Chem. 1994, 269, 17030–17034. [Google Scholar] [CrossRef]
- Cavic, B.A.; Thompson, M. Interfacial nucleic acid chemistry studied by acoustic shear wave propagation. Anal. Chim. Acta 2002, 469, 101–113. [Google Scholar] [CrossRef]
- Kull, T.; Nylander, T.; Tiberg, F.; Wahlgren, N.M. Effect of surface properties and added electrolyte on the layer structure of β-casein layers adsorbed at the solid/aqueous interface. Langmuir 1997, 13, 5141–5147. [Google Scholar] [CrossRef]
- Sauerbrey, G. Verwendung von schwingquarzen zur wägung dünner schichten und zur mikrowägung. Zeitschrift für Physik 1959, 155, 206–222. [Google Scholar] [CrossRef]
- Ellis, J.S.; Thompson, M. Viscoelastic modeling with interfacial slip of a protein monolayer electrode-adsorbed on an acoustic wave biosensor. Langmuir 2010, 26, 11558–11567. [Google Scholar] [CrossRef] [PubMed]
- Maron, B.A.; Loscalzo, J. The role of platelets in fibrinolysis. In Platelets, 2nd ed.; Michelson, A.D., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2007; pp. 415–430. [Google Scholar] [CrossRef]
- Takada, Y. Potential role of Kringle-integrin interaction in plasmin and uPA actions (A Hypothesis). J. Biomed. Biotech. 2012, 2012, 136302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edy, J.; Collen, D. The interaction in human plasma of antiplasmin, the fast-reacting plasmin inhibitor, with plasmin, trypsin and chymotrypsin. Biochim. Biophys. Acta 1977, 484, 423–432. [Google Scholar] [CrossRef]
- Bumberger, R.; Belitz, H.D. Bitter taste of enzymic hydrolysates of casein. Z. Lebensm. Unters. Forsch. 1993, 197, 14–19. [Google Scholar] [CrossRef]
- Tandon, O.P.; Tripathi, Y. Best & Taylor’s Physiological Basis of Medical Practice, 13th ed.; Kluwer India Pvt. Ltd.: Gurugram, India, 2018. [Google Scholar]
- Thomas, A.; Kohler, M.; Walpurgis, K.; Schänzer, W.; Thevis, M. Proteolysis and autolysis of proteases and the detection of degradation products in doping control. Drug Test. Anal. 2009, 1, 81–86. [Google Scholar] [CrossRef]
- Jiang, L.; Yuan, C.; Huang, M. A general strategy to inhibit serine protease by targeting its autolysis loop. FASEB J. 2021, 35, e21259. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| α2-Antiplasmin (nM) | Trypsin | Plasmin | ||||
|---|---|---|---|---|---|---|
| Δf (Hz) | Δm/A (μg/cm2) | εi (%) | Δf (Hz) | Δm/A (μg/cm2) | εi (%) | |
| 0 | 28.97 ± 0.31 | −0.513 ± 0.005 | 0 | 19.8 ± 1.01 | −0.350 ± 0.018 | 0 |
| 1.25 | 23.94 ± 0.36 | −0.424 ± 0.006 | 15.29 | 16.26 ± 1.37 | −0.288 ± 0.024 | 28.17 |
| 2.5 | 15.46 ± 0.8 | −0.274 ± 0.014 | 49.48 | 14.4 ± 0.13 | −0.255 ± 0.02 | 49.73 |
| 3.75 | 12.59 ± 0.47 | −0.223 ± 0.008 | 80.06 | 3.9 ± 0.65 | −0.069 ± 0.011 | 75.84 |
| 5.0 | −3.98 ± 0.6 | 0.07 ± 0.01 | 100 | −3.11 ± 0.81 | 0.055 ± 0.014 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tatarko, M.; Ivanov, I.N.; Hianik, T. New Insights on Plasmin Long Term Stability and the Mechanism of Its Activity Inhibition Analyzed by Quartz Crystal Microbalance. Micromachines 2022, 13, 55. https://0-doi-org.brum.beds.ac.uk/10.3390/mi13010055
Tatarko M, Ivanov IN, Hianik T. New Insights on Plasmin Long Term Stability and the Mechanism of Its Activity Inhibition Analyzed by Quartz Crystal Microbalance. Micromachines. 2022; 13(1):55. https://0-doi-org.brum.beds.ac.uk/10.3390/mi13010055
Chicago/Turabian StyleTatarko, Marek, Ilia N. Ivanov, and Tibor Hianik. 2022. "New Insights on Plasmin Long Term Stability and the Mechanism of Its Activity Inhibition Analyzed by Quartz Crystal Microbalance" Micromachines 13, no. 1: 55. https://0-doi-org.brum.beds.ac.uk/10.3390/mi13010055