
Current Advances in the Biodegradation and Bioconversion of Polyethylene Terephthalate

1
Frontier Science Center for Synthetic Biology and Key Laboratory of Systems Bioengineering (Ministry of Education), School of Chemical Engineering and Technology, Tianjin University, Tianjin 300072, China
2
Collaborative Innovation Center of Chemical Science and Engineering (Tianjin), Tianjin University, Tianjin 300072, China
*
Author to whom correspondence should be addressed.
Microorganisms 2022, 10(1), 39; https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10010039
Submission received: 22 November 2021
/
Revised: 15 December 2021
/
Accepted: 20 December 2021
/
Published: 26 December 2021
(This article belongs to the Special Issue Biodegradation and Environmental Microbiomes)
Abstract
:Polyethylene terephthalate (PET) is a widely used plastic that is polymerized by terephthalic acid (TPA) and ethylene glycol (EG). In recent years, PET biodegradation and bioconversion have become important in solving environmental plastic pollution. More and more PET hydrolases have been discovered and modified, which mainly act on and degrade the ester bond of PET. The monomers, TPA and EG, can be further utilized by microorganisms, entering the tricarboxylic acid cycle (TCA cycle) or being converted into high value chemicals, and finally realizing the biodegradation and bioconversion of PET. Based on synthetic biology and metabolic engineering strategies, this review summarizes the current advances in the modified PET hydrolases, engineered microbial chassis in degrading PET, bioconversion pathways of PET monomers, and artificial microbial consortia in PET biodegradation and bioconversion. Artificial microbial consortium provides novel ideas for the biodegradation and bioconversion of PET or other complex polymers. It is helpful to realize the one-step bioconversion of PET into high value chemicals.
1. Introduction
Polyethylene terephthalate (PET) is one of the most widely used synthetic plastics in people’s lives [1]. It is polymerized by terephthalic acid (TPA) and ethylene glycol (EG) through ester bonds [2]. Since PET was first used to produce disposable plastic bottles in the 20th century, it has been welcomed worldwide and has become an indispensable part of people’s lives [3]. As PET is highly resistant to natural degradation, the recycling of PET has been encouraged [4]. At present, the main methods for managing PET waste include landfilling, incineration, as well as physical and chemical recycling [5,6]. These methods usually cause secondary pollution to the environment and consume huge amounts of energy, which is not economical or environmentally friendly. Due to the improper recycling strategies and the strong mechanical properties of plastic products, serious environmental problems, such as soil pollution and the disturbance of marine ecosystems, have occurred [7]. Therefore, PET biodegradation has attracted attention as an environmentally friendly alternative, requiring milder temperatures and lower energy consumption than other recycling methods [8,9]. Additionally, the degradation monomers can easily be recycled, with the hope of converting PET into high value chemicals.
In 1977, several commercial lipases and an esterase were reported to hydrolyze various kinds of polyesters [10]. Since then, many PET hydrolases, such as lipases, cutinases and esterases, have been discovered and characterized by various microorganisms [1,11]. In 2016, Ideonella sakaiensis 201-F6 was isolated from a waste recycling station [12]. It was found to produce PET hydrolase (PETase) and monohydroxyethyl terephthalate (MHET) hydrolase (MHETase), which can degrade PET into intermediate products at 30 °C. Then, the structures of the two enzymes were analyzed and a series of effective enzyme modifications were carried out [13,14,15,16,17], efficiently improving the activity and stability of the two enzymes. The discovery and modification of PETase and MHETase has provided an important basis for the degradation of PET waste under ambient temperatures.
Synthetic biology and metabolic engineering strategies have been applied to the biodegradation and bioconversion of PET waste, especially in the modification of PET hydrolases, optimization of microbial chassis, and reconstruction of degradation pathways. At present, some bacteria, fungi, and marine microalgae have been reported as being good microbial chassis for PET biodegradation. The whole-cell biocatalysts have been able to achieve the initial degradation of PET. The bioconversion pathways of TPA and EG have been identified. Some microorganisms have been engineered to produce high value chemicals from PET monomers, which is an important development direction for PET upcycling. Based on these current advances, developing enhanced microbial chassis and constructing artificial microbial consortia to couple the biodegradation of PET by secreted PET hydrolases with the bioconversion of high value chemicals from monomers is a promising method to realize the circular economy of PET waste.
This review summarizes the current advances in the modified PET hydrolases, engineered microbial chassis in degrading PET, bioconversion pathways of PET monomers, and artificial microbial consortia in PET biodegradation, providing novel ideas for the future degradation of PET, and other types of polymers, by artificial microbial consortia.
2. PET Biodegradation
During PET biodegradation, the microorganisms first adhere onto the surface of PET films and then secrete extracellular PET hydrolases, which bind to the PET films and initiate the biodegradation process [18,19]. PET hydrolases act on the ester bond of PET, hydrolyzing it into TPA and EG and generating incomplete hydrolysis products, such as MHET and Bis-(2-hydroxyethyl) terephthalate (BHET). In I. sakaiensis 201-F6, MHET can be further hydrolyzed into TPA and EG under the action of MHETase [12]. It was reported that MHETase has a hydrolysis activity against the termini-generated PET film, demonstrating the exo-PETase function of the enzyme [20]. PET hydrolases can further hydrolyze BHET to produce MHET, TPA, and EG [12]. The products TPA and EG can be used by different microorganisms and be further metabolized into the tricarboxylic acid cycle (TCA cycle) [21,22,23,24,25,26,27,28,29]. Additionally, these intermediate and final products of PET biodegradation have been identified as competitive inhibitors of PET hydrolases [30,31] (Figure 1).
2.1. Engineered PET Hydrolases
The hydrolases, including lipases [31,32,33,34], cutinases [35,36,37,38,39,40,41,42], esterases [43,44,45,46], PETase [12] and MHETase [12], that can degrade PET have been identified. Among them, lipases have the lowest hydrolysis activity of PET mainly because their catalytic centers are covered by lid structures, which limits the hydrolases’ contact and catalysis with the substrate PET. Cutinases always have a strong PET hydrolysis ability due to their large substrate binding pockets without lid structures, which is conducive to the combination of PET with their active centers. However, cutinases usually degrade PET at high temperatures (50–70 °C), while PETase and MHETase can efficiently and specifically hydrolyze PET at 30 °C [12]. The discovery of PETase and MHETase is helpful in achieving the high efficiency biodegradation of PET at ambient temperatures. At present, the structures of these two enzymes have been studied extensively, and more high activity hydrolases variants have appeared.
The PET hydrolases identified in nature always have poor stability, low activity, and low expression levels, which limit their large-scale industrial application. A series of strategies that could enhance the catalytic activity of PET hydrolases have been proposed [13] (Table 1).
One strategy is to engineer the binding pocket, which can improve the specificity of the PET hydrolases and increase the effective adsorption of enzymes and substrates [15,47,48,49]. Our laboratory previously focused on six key amino acids near the binding of PETase to the substrate and conducted site-directed mutations. The R61A, L88F, and I179F mutants were successfully screened, and the enzyme activity increased 1.4-fold, 2.1-fold, and 2.5-fold, respectively, in comparison to wild-type PETase [50]. Silva et al. [51] modified the cutinase from Thermobifida fusca_0883 by site-directed mutagenesis and constructed a single mutation Ile218Ala and a double mutation Q132A/T101A, which expanded the catalytic space and improved the efficiency of the PET biodegradation. Chen et al. [52] identified the unique amino acids S214 and I218 through the structural analysis of PETase and noted that they are associated with W185 wobbling and β6-β7 loop flexibility. This research is helpful in designing PETase mutants that increase the flexibility of the substrate binding pocket.
Some studies focused on using enzyme engineering strategies to improve the stability of the PET hydrolases to improve PET biodegradation efficiency [53,54]. Methods such as adding Ca2+ or Mg2+ [38,55], introducing a disulfide bond and salt bridge [56,57], and glycosylation have all been proven to improve the stability of PET hydrolases. Researchers added disulfide bonds to improve the thermal stability of leaf-branch compost cutinase (LCC) and performed site-directed mutations on hot amino acids near the substrate binding to obtain the combined mutation F243I/D238C/S283C/Y127G (ICCG) [53]. Finally, 90% of shredded PET plastic bottles were degraded at 72 °C for 10 h, which is by far the most efficient PET hydrolase [53].
Additionally, increasing the substrate accessibility for PET hydrolases by engineering the PET hydrolases has also been widely studied [58,59,60]. It is reported that the fusion expression of Thc_Cut1 from Thermobifida cellulosilytica and hydrophobins (HFB4 and HFB7) from Trichoderma reesei can increase the hydrolysis effect of PET by more than 16 times, while a mixture of the enzyme and the hydrophobins led to only a 4-fold increase at most [61].
The intermediate and final products of PET biodegradation, such as Bis-(2-hydroxyethyl) terephthalate (BHET), monohydroxyethyl terephthalate (MHET), TPA, and EG, are all competitive inhibitors of the PET hydrolases [30]. Therefore, the mixtures of hydrolases that act synergistically or protein engineering strategies that reduce the interaction between the enzymes and products are effective methods for solving the inhibition [62,63,64].
2.2. Engineered PET Biodegradation Chassis
Most of the microorganisms identified that are capable of secreting PET hydrolases are non-model microorganisms and they are difficult to genetically engineer due to their complex genetic background. In addition, the expression level of the PET hydrolases from wild strains is insufficient to satisfy the demand for large-scale degradation. Therefore, it is necessary to develop recombinant expression systems using model microorganisms to express the PET hydrolases efficiently. PET is a high molecular polymer that is polymerized from TPA and EG and cannot enter cells, so in vitro enzymatic degradation of PET has been studied extensively. Owing to the purification and preparation process of PET hydrolases being time-consuming and cost-intensive, the efficient expression PET hydrolases extracellularly for practical applications is necessary [76,77].
At present, some microbial chassis such as bacteria, fungi, and marine microalgae have been applied to the secretion and expression of PET hydrolases, which have been studied and proven to be promising chassis to degrade PET (Table 2). Several whole-cell biocatalysts have been designed to degrade PET, which are able to not only avoid the complicated steps of enzyme purification but also be reused in multi-step reactions, in comparison to the free enzyme-based approach [78,79]. Additionally, the difficulty of the reduced activity of the enzymes, or even enzymes being inactivated, under the influence of environmental factors has been solved. The following is a summary of several microbial chassis that are suitable for PET biodegradation.
2.2.1. Bacteria
Escherichia coli
E. coli is an important model microorganism for the production of recombinant proteins due to its clear genetic background, simple growth conditions, and its advantages in high density cultivation [89]. In recent years, with the continuous discovery of PET hydrolases, more and more enzymes have achieved the heterologous expression in E. coli [12,14,16,50,53,69]. PET hydrolases heterologously expressed in E. coli have been summarized [76] and it is helpful in further analyzing the crystal structures of these enzymes and explore the degradation mechanism for PET.
Recent studies have shown that engineered E. coli can be used as a whole-cell biocatalyst for PET biodegradation. Selecting the optimal signal peptide is a common strategy used to improve the section of heterologous PET hydrolases. A study tested the effects of Sec-dependent and SRP-dependent signal peptides from E. coli in secreting PETase, and successfully produced 6.2 mg/L PETase by fusing SPLamB and PETase [80]. Some other research improved the expression titer and enzymatic activity by modifying the signal peptide. An evolved signal peptide PelB (G58A) obtained by random mutagenesis was successfully used to express heterologous PETase in E. coli and enabled up to 1.7-fold higher PETase secretion [81]. An enhancer of signal peptides B1 (MERACVAV) was studied to mediate the excretion of PETase, and finally, the excretion efficiency of PETase mediated by B1PelB demonstrated a 62-fold increase over that of PelB [82].
Bacillus subtilis
Gram-positive B. subtilis has the advantages of high secretion capacity, fast growth, and the lack of an outer membrane, and it is regarded as an excellent microbial chassis for secreting heterologous proteins compared to E. coli, which usually forms an inclusion body [90,91]. Additionally, B. subtilis has a strong resistance to harsh environments and it has been used to secrete proteins that can degrade many pollutants, which is why it is considered to be a promising microbial chassis for biodegradation [92,93].
In terms of PET biodegradation, B. subtilis has been engineered to secrete PET hydrolases. It is reported that PETase was successfully secreted into the medium by B. subtilis 168 under the direction of its native signal peptide (SPPETase). SPPETase is predicted to be a twin-arginine signal peptide, and the inactivation of twin-arginine translocation (Tat) complexes improved the secretion amount of PETase 3.8-fold [83]. Another two PET hydrolases (BhrPETase and LCC) were also expressed in B. subtilis, and the expression titer of BhrPETase and LCC reached 0.66 g/L and 0.89 g/L in an engineered chaperone-overexpression of B. subtilis, respectively [42]. Additionally, the combinations of signal peptides and promoters were optimized to promote the expression of PETase in B. subtilis WB600, and the combination of the signal peptide SPamy and the weak promoter P43 was proved to be best [84].
Thermophilic Bacteria
Most of the hydrolases capable of degrading PET, including lipases, cutinases and esterases, have higher enzymatic activity at higher temperatures, while the optimal growth temperature of most model microorganisms that can produce heterologous PET hydrolases is usually 30–40 °C. Whole-cell biocatalyst is not compatible with some PET hydrolases that are only functional at high temperatures [94]. Therefore, a thermophilic expression system is necessary to improve the efficiency of PET biodegradation [36,95]. Most thermophilic microorganisms are usually difficult to genetically engineer except for Clostridium thermocellum, which has a mature genetic manipulation platform [94]. C. thermocellum has been engineered for lignocellulose bioconversion [96] and biofuel production [97], which is why it is regarded as a potential microbial chassis for the biodegradation of PET.
LCC has been successfully obtained from an engineered C. thermocellum. This engineered whole-cell biocatalyst realized a high level expression of LCC and more than 60% of a commercial PET film was converted into soluble monomers at 60 °C after 14 days [85]. This thermophilic whole-cell degradation system has the advantage of simultaneous enzyme production and PET degradation compared to only using free enzymes, which is why it is a promising strategy to degrade PET using other high temperature hydrolases [98,99]. In addition to the thermophilic whole-cell degradation system, an alkali-tolerant whole-cell catalytic system has also been reported [100,101,102].
2.2.2. Fungi
In addition to bacteria, the potential of some yeasts, including Pichia pastoris and Yarrowia lipolytica, being used in PET biodegradation has been studied. P. pastoris, with a great secretion expression system and scalable fermentation capability, has become a common strain for protein production in industrial applications. Researchers have expressed BurPL (H344S/F348I) and PETase in P. pastoris and E. coli and noted that both enzymes produced from P. pastoris showed higher activity than that expressed in E. coli because of the protein half-life protection mechanism of P. pastoris [52,103]. A whole-cell biocatalyst was developed by displaying PETase on the surface of P. pastoris and the enzymatic activity of PETase increased 36-fold towards a highly crystalline PET in comparison to that of purified PETase [86]. Additionally, this whole-cell biocatalyst can be reused seven times without obvious activity loss, which is helpful in developing other whole-cell biocatalysts for PET biodegradation [86]. Considering the ability of P. pastoris to perform N-linked glycosylation, some researchers studied the effects of glycosylation on the LCC expressed in P. pastoris and found that the kinetic stability and activity of LCC were both improved [35]. Y. lipolytica is also a great microbial chassis for bioremediation [104]. Researchers isolated Y. lipolytica IMUFRJ 50682 with the ability to convert PET into MHET and verified that the PET monomers may act as inducers in the process of lipase production [105], which showed that Y. lipolytica is a potential microbial chassis for PET biodegradation. Other research expressed PETase in Y. lipolytica Po1f with a signal peptide from lipase and confirmed that the engineered strain could hydrolyze BHET and PET powder into the monomers [106]. Surface display systems and whole-cell biocatalysts provide novel ideas and strategies for achieving the high efficiency expression of PET hydrolases and promoting PET biodegradation [77,107,108]. Yeasts, together with efficient genetic tools, have been used as great microbial chassis for biodegradation and bioconversion [109].
2.2.3. Marine Microalgae
At present, the existing native and engineered microbial chassis that are capable of producing PET hydrolases are usually difficult to adapt to the complexity of the marine environment and produce much PET waste. Recently, some marine microalgae have been used as chassis for PET biodegradation [110]. A photosynthetic microalga Phaeodactylum tricornutum has been reported as being engineered as a chassis capable of secreting a PETase mutant into the culture medium, and the recombinant PETase was able to efficiently degrade different substrates, including PET films, poly (ethylene terephthalateco-1,4-cylclohexylenedimethylene terephthalate) (PETG) film, and shredded PET, at 30 °C or even at mesophilic temperatures (21 °C) [87]. Additionally, Chlamydomonas reinhardtii, the green algae, was also successfully engineered to produce PETase with degrading activity, and the chemical and morphological changes appeared on the PET films after 4 weeks of culture [88]. As environmentally friendly chassis for the biodegradation of PET waste in a saltwater-based environment, marine microalgae have the potential for future biotechnological applications in the degradation of PET polluted seawater [87].
3. Metabolism and Bioconversion of PET Monomers
It is reported that Acetobacterium woodii, Pseudomonas sp., and E. coli have ability to utilize EG. In A. woodii, EG can be utilized by an acetaldehyde/ethanol pathway while it is consumed by a glyoxylic acid pathway in Pseudomonas sp. and E. coli. TPA can be also utilized by some bacteria, such as Rhodococcus sp., etc. Additionally, a series of metabolic engineering strategies have realized the bioconversion of EG and TPA into high value chemicals, such as glycolic acid, PHA, gallic acid, vanillic acid, and β-ketoadipic acid. The metabolism and bioconversion pathways of the PET monomers, TPA and EG, are shown in Figure 1.
3.1. Metabolism of EG
At present, two naturally existing pathways, including the acetaldehyde/ethanol pathway and glyoxylic acid pathway for the utilization of EG by microorganisms, have been reported (Figure 1, green pathway). The use of EG is not commonly reported in metabolic engineering of model microorganisms, except for E. coli.
3.1.1. Acetaldehyde/Ethanol Pathway
The acetogenic bacterium A. woodii can use EG as the sole carbon source for growth, and the EG metabolic pathway has been identified [25]. EG is dehydrated to acetaldehyde, catalyzed by the propane diol dehydratase (PduCDE), then further converted into ethanol and acetyl coenzyme A (acetyl-CoA), catalyzed by CoA-dependent propionaldehyde dehydrogenase (PduP) [111]. PduCDE and PduP are both encoded by the Pdu gene cluster [111]. Acetyl-CoA and a part of the ethanol are converted into acetic acid, and this process is accompanied by the production of adenosine triphosphate (ATP) and nicotinamide adenine dinucleotide (NADH) [111]. The reducing equivalents of the ethanol oxidation are recycled through the reduction of carbon dioxide (CO2) into acetate in the Wood–Ljungdahl pathway [25]. The acetaldehyde/ethanol pathway is commonly found in some Clostridium species and a few other anaerobic organisms because the enzymes that catalyze EG are oxygen sensitive. Additionally, Dragan et al. [25] and Nilanjan et al. [112] proved that the enzymes for EG utilization were encapsulated in bacterial microcompartments.
3.1.2. Glyoxylic Acid Pathway
Glyoxylic Acid Pathway in Pseudomonas sp.
In Pseudomonas aeruginosa and Pseudomonas putida, EG is converted into glyoxylic acid under the action of dehydrogenase and finally enters the TCA cycle through different routes [21,22,23,113]. At present, the metabolic pathway of EG in P. putida KT2440 is the most widely studied. The metabolism pathways in utilizing EG have been well demonstrated in P. putida KT2440, in comparison to other bacteria, and related enzymes have been identified. In P. putida KT2440, two functionally redundant periplasmic quinoproteins, PedE and PedH, catalyze EG into glycolaldehyde [114]. PedE and PedH are both pyrroloquinoline quinone-dependent alcohol dehydrogenases (PQQ-ADHs), and their expression depend on Ca2+ and lanthanide metal ions, respectively [114]. Once glycolaldehyde is produced, the two cytoplasmic aldehyde dehydrogenases, PP_0545 and PedI, catalyze it into glycolic acid, and glyoxylic acid is further generated via the membrane anchored oxidase GlcDEF. The glyoxylic acid is converted into acetyl-CoA and enters the TCA cycle to be catalyzed by a series of enzymes [115]. Additionally, there are another two alternative pathways to convert glyoxylic acid, one of which is catalyzed by isocitrate lyase (AceA) and glyoxylic acid can condense with succinic acid to form isocitrate. The other is catalyzed by malate synthase (GlcB) and glyoxylic acid condenses with acetyl-CoA to form malic acid. However, due to the removal of CO2 and the restriction of the amount of acetyl-CoA, P. putida KT2440 cannot use EG as the sole carbon source for growth [115]. Researchers engineered P. putida KT2440 by overexpressing glycolate oxidase to remove the glycolate metabolic bottleneck and produced an engineered strain that can efficiently metabolize EG [115]. After that, mutants of P. putida KT2440 that utilize EG as the sole carbon source were isolated through adaptive laboratory evolution, and the metabolism and regulation mechanism of EG in P. putida KT2440 was further clarified [116]. P. putida JM37 was reported to be able to use EG as the sole carbon source for growth because there is another pathway to use glyoxylic acid compared to P. putida KT2440. Glyoxylic acid is converted into tartrate semialdehyde under the catalysis of glyoxylate carboxylase (Gcl) and then tartrate semialdehyde is converted into glycerate acid, catalyzed by hydroxypyruvate isomerase (Hyi) and tartrate semialdehyde reductase (GlxR). Glycerate acid can be further converted into 2-phosphoglycerate and enter the TCA cycle [117].
Glyoxylic Acid Pathway in E. coli
Wild-type E. coli cannot use EG as the sole carbon source for growth [118]. In 1983, researchers first reported an E. coli strain capable of using EG as the sole carbon source from the propylene glycol using mutants. They identified the increased activities of propanediol oxidoreductase, glycolaldehyde dehydrogenase, and glycolate oxidase in the mutants [118]. Based on this discovery, researchers began to design and construct engineered E. coli that could use EG to convert PET monomers into high value chemicals.
EG is assimilated and oxidized into glycolaldehyde and, subsequently, into glycolic acid under the catalysis of 1,2-propanediol oxidoreductase mutant (fucO) and glycolaldehyde dehydrogenase (aldA), respectively. Glycolic acid can be metabolized into glyoxylic acid by glycolate dehydrogenase (GlcDEF) [26]. Similar to P. putida, glyoxylic acid is further condensed into acetyl-CoA by the linear glycerate pathway or converted into isocitrate and malate catalyzed by AceA and GlcB, respectively. An engineered E. coli can take EG as the sole carbon source to produce glycolate by expressing fucO mutant (I7L/L8V) and aldA. Experiments identified that oxygen concentration was as an important metabolic valve, and flux to 2-phosphoglycerate was the primary route in the assimilation of EG as a substrate combining modeling [113,119]. Additionally, EG can be efficiently utilized in E. coli by optimizing the gene expression (fucO and aldA) and adding a growth medium with a low concentration of glycerol or a mixture of amino acids [26]. Although E. coli MG1655 contains the endogenous glyoxylic acid metabolism pathway, the EG-utilizing ability of the engineered E. coli still needs to be improved [119]. Introducing a heterologous pathway or unblocking the rate-limiting steps of the EG metabolic pathway in E. coli may further enhance the assimilation of EG. E. coli has a clear genetic background and simple genetic operations compared to other bacteria, so it is easier to engineer it to transform EG into high value chemicals.
3.2. Bioconversion of EG to High Value Chemicals
EG is one of the cheap raw materials for glycolic acid production through incomplete oxidation. Several wild microorganisms, including Pichia naganishii [23], Rhodotorula sp. [23], Burkholderia sp. [120], Gluconobacter oxydans [24], and Hansenula sp. [121], have been reported to produce glycolic acid from EG. Among these microorganisms, G. oxydans has been extensively studied due to its high titer of glycolic acid from EG. It is reported that the overexpression of membrane-bound alcohol dehydrogenase (mADH) in G. oxydans DSM 2003 accelerated cell growth, and 113.8 g/L of glycolic acid was accumulated with a molar yield of 92.9% within 45 h [122]. Two genes encoding recombinant cytosolic oxidoreductases (gox0313 and gox0646) from G. oxydans were heterologously expressed in E. coli and the resulting proteins were purified and characterized [123]. In addition to G. oxydans, engineered E. coli has potential in producing glycolic acid from EG, and 10.4 g/L of glycolic acid was produced from EG after 112 h in a fed-batch bioreactor using a series of oxygen-based strategies [26,118].
EG can also be used to produce polyhydroxyalkanoate (PHA) by P. putida under nitrogen-limiting conditions [124]. An engineered strain P. putida KT2440 realized the conversion of EG into mcl-PHAs [115,124] and some metabolic engineering strategies were developed to enhance medium chain length polyhydroxyalkanoates (mcl-PHAs) production in P. putida [125,126,127]. mcl-PHAs can be upgraded into chemical precursors and fuels via a straightforward catalytic process [128].
3.3. Metabolism of TPA
It was reported that Comamonas sp. [27], Delftia tsuruhatensis [129], Comamonas testosterone [130], and Rhodococcus sp. [28] can use TPA as the sole carbon source for their growth (Figure 1, orange pathway). In these bacteria, TPA enters the cell via the TPA transporters [29]. Generally, TPA can be transformed into 1,6-dihydroxycyclohexa-2,4-diene-dicarboxylate (DCD) under the catalysis of TPA dioxygenase (TphAabc), and DCD is further oxidized by 1,2-dihydroxy-3,5-cyclohexadiene-1,4-dicarboxylate dehydrogenase (TphB) to form protocatechuate (PCA) [130,131,132,133]. The genes responsible for these reactions have been characterized [27,28,129,130]. Comamonas sp. E6 also contains the extra gene TphC, which encodes a permease involved in TPA uptake using the tripartite aromatic acid transporter [29]. There are three main pathways for the metabolism of PCA, the ortho-, meta-, and para-cleavage pathways, which are catalyzed by 3,4-dioxygenase (PCDO), 4,5-dioxygenase, and 2,3-dioxygenase, respectively [134,135,136,137]. At present, the ortho-cleavage pathway is the most extensively studied, and PCA is converted into β-carboxymuconate under the catalysis of protocatechuate 3,4-dioxygenase (PCDO), is finally converted into acetyl-CoA, and enters the TCA cycle [134,138].
3.4. Bioconversion of TPA to High Value Chemicals
It has been demonstrated that the PET monomer TPA is suitable for the biosynthesis of several high value chemicals, such as gallic acid, pyrogallol, catechol, muconic acid, vanillic acid, catechol, adipic acid, PHA, and β-ketoadipic acid [106,139,140,141,142,143]. Since PCA is an important precursor in producing a series of high value aromatic chemicals, the key to the bioconversion of TPA is the acquisition of PCA. By the heterologous expression of TPA, 1,2-dioxygenase (TphAabc), and DCD dehydrogenase (TphB) from Comamonas sp., E. coli was engineered to utilize TPA and produced PCA [133]. Further heterologous expression of different enzymes produced gallic acid, pyrogallol, catechol, muconic acid and vanillic acid from PCA in E. coli [133]. Additionally, a novel pathway for the direct upcycling of TPA into the value-added small molecule vanillin was reported in engineered E. coli and the conversion efficiency reached 79% [143].
PHA can also be produced from TPA. Researchers have isolated P. putida GO16 and P. putida GO19 from a PET bottle processing plant and proved their ability to convert TPA into PHA at a maximal rate of approximately 8.4 mg·L−1·h−1 for 12 h [144]. Recently, researchers engineered Pseudomonas umsongensis GO16 to convert PET into two types of bioplastics, PHA and a novel bio-based poly (amide urethane) (bio-PU), and further achieved the secretion of hydroxyalkanoyloxy alkanoates (HAAs) by introducing the HAA synthesis module into the engineered strain [145]. Poly-(R)-3-hydroxybutyrate (PHB), the first PHA discovered, has also been produced from PET through the heterologous expression of the phbCAB operon from Ralstonia eutropha in Pseudomonas stutzeri [106]. Due to the same synthetic precursors of rhamnolipids and PHA, many microorganisms capable of converting PET into PHA also have the potential to synthesize rhamnolipids [144]. The conversion of PET into biodegradable plastics is a clean and cost-effective way to generate a great market in PET recycling [146].
As for producing β-ketoadipic acid from TPA, four sequential metabolic engineering efforts in P. putida KT2440 were performed to directly convert BHET into β-ketoadipic acid [139]. The engineered P. putida is able to not only degrade BHET into TPA and EG, but also convert TPA into 15.1 g/L of β-ketoadipic acid at 76% molar yield in bioreactors [139]. β-ketoadipic can be further polymerized into a nylon-6,6 analog, or other products [147].
PET waste is depolymerized by microorganisms in nature and converted into CO2 and water, which causes serious resource loss and carbon emissions. Therefore, utilizing PET and its monomers to produce high value chemicals provides a new solution for upgrading and recycling PET and other plastics waste [148].
4. Microbial Consortia in PET Biodegradation
Artificial microbial consortia that simulate natural microbial consortia to complete complex biological processes have become an important research topic in synthetic biology [86,89,149,150]. It is important to explore the potential and reprogram the functionality of microbial consortium members for specific approaches, especially for the bioconversion of contaminants [151,152]. Previous studies have highlighted the potential for biodegradation and bioconversion using artificial microbial consortia [153,154,155]. Artificial microbial consortia have been used to degrade hydrocarbons [153], organophosphorus pesticides [156], polyaromatic hydrocarbon pollutants [157], and aryl organophosphate flame retardants (aryl-OPFRs) [158] and improve the desulfurization of petroleum sulfides [159]. In addition, some artificial microbial consortia have been constructed in degrading plastic waste, such as polyurethane (PU) [154], polyethylene (PE) [160,161], polypropylene (PP) [162], and polyvinyl chloride (PVC) [163]. These results highlight the potential of artificial microbial consortia in PET biodegradation.
There are specific advantages in PET biodegradation by artificial microbial consortia compared to pure culture: (i) the synergies of different enzymatic systems and combination of the metabolic pathways of various microorganisms can relieve the inhibition of degradation products and improve degradation rate; (ii) PET biodegradation and bioconversion can be realized simultaneously by different microorganisms; and (iii) the construction of artificial microbial consortia is more efficient and time-saving than other metabolic engineering strategies [164]. Therefore, the application of artificial microbial consortia in the biodegradation and bioconversion of PET is regarded as a promising method to realize the circular economy of PET waste.
4.1. Natural Microbial Consortia in PET Biodegradation
At present, most microbial consortia that are capable of degrading PET are natural microbial consortia. Researchers isolated a consortium, including three Pseudomonas sp. And two Bacillus sp., that can reduce the granular PET and they identified that a 100 mg PET granule weighed 3.15 mg less when with the consortium for six weeks, which suggests that the strains can act synergistically to degrade PET [165]. After that, researchers screened for lipase activity associated with PET biodegradation and proved that the secreted enzymes extracted from the consortium could fully convert BHET into TPA and EG [166]. Another study reported a consortium, including Bacillus cereus SEHD031MH and Agromyces mediolanus PNP3, from activated sludge. It noted that the consortium could use PET microplastics (MPs) as the sole carbon source and degrade 17% of PET MPs at 30 °C of the course of 168 days [167]. Additionally, Oberbeckmann et al. [168] analyzed the influence of different seasons, geographic locations, seawater, and substrate material types on the microbial consortia of using single-use PET bottles at multiple stations in the North Sea.
Most PET hydrolases that have been previously reported, such as cutinases, lipases, and esterases, can only accomplish the limited degradation of PET. In 2016, Yoshida et al. [12] successfully isolated a microbial consortium No. 46 that degraded amorphous PET from a waste recycling station completely at ambient temperature. Then, a bacterium capable of degrading and assimilating PET named I. sakaiensis 201-F6 was isolated from No. 46 consortium. It could produce PETase and MHETase to degrade PET, which provided a new direction for the biodegradation of PET under ambient temperatures [4,12].
Marine microbial consortia can colonize PET, form biofilms on its surface, and finally modify its chemical structure [169,170,171,172]. A study demonstrated for the first time that hydrocarbon-degrading marine consortia enriched on tetradecane and diesel have the potential to degrade PET and cause major alterations to the surface structure and hydrophobicity of PET films [173].
4.2. Artificial Microbial Consortia in PET Biodegradation
At present, there are few studies on the construction of artificial microbial consortia to degrade PET. A distinct three-consortium named CAS6 was isolated from an ocean bay and it can make PET films lose sharp morphology, compared to controls after 4 weeks [174]. Then, three bacteria (Exiguobacterium sp., Halomonas sp., and Ochrobactrum sp.) were identified from CAS6 and formed a stable artificial three-microbial consortium in a 1:1:1 ratio to efficiently degrade PET films. PET films were fully degraded into small pieces after 2 weeks of incubation by the three-microbial consortium [174]. Pan et al. [106] designed an engineered Y. lipolytica to secrete PETase in order to degrade PET and an engineered Pseudomonas stutzerithe to convert TPA into PHB. They constructed a microbial consortium with two engineered strains to achieve the conversion of BHET into PHB over the course of 54 h. This was the first attempt at performing the enzymatic hydrolysis of PET and the bioconversion of TPA simultaneously. Although PHB could not be synthesized directly from PET because of the low hydrolyzing efficiency of PETase, it demonstrated the possibility of artificial microbial consortia achieving the simultaneous degradation and upcycling of PET [106]. Our laboratory has constructed a four-species microbial consortium composed of two metabolically engineered B. subtilis, Rhodococcus jostii and P. putida to degrade PET, and the weight loss of PET film reached 23.2% under ambient temperature [149]. The artificial microbial consortium successfully relieved the metabolic inhibition of TPA and EG, and effectively improved the degradation rate [149].
4.3. Prospect of PET Biodegradation by Artificial Microbial Consortia
The artificial microbial consortia are expected to effectively solve the problems in the PET biodegradation process and improve the degradation efficiency. During the biodegradation of PET, the intermediate and final products, such as BHET, MHET, TPA, and EG, have been identified as competitive inhibitors of PET hydrolases [30,31,115]. Constructing artificial microbial consortia can relieve the inhibition of metabolites and promote biodegradation, which has been demonstrated by a two-microbial consortium for corn fiber conversion [175]. By combining the PET-degrading module and the monomer-converting modules to construct an artificial microbial consortium, we speculate that the inhibitory effect of TPA and other substances can be relieved (Figure 2). In addition, only using a single bacterium to degrade PET and convert the degradation products will increase its metabolic burden, while an artificial microbial consortium can solve that problem. A three-microbial consortium, including three engineered E. coli, was constructed to reduce the metabolic burden and synthesize rosmarinic acid [176]. Another three-microbial consortium was constructed to produce short chain fatty acids from lignocellulose and it can reduce the metabolic burden and perform multiple tasks [177].
In view of the current problems of PET biodegradation, the artificial microbial consortium is an important metabolic strategy to solve them. Artificial microbial consortia can be used to perform more complex tasks in a more complex environment [178,179,180,181]. Constructing a PET-degrading module, an EG-converting module, and a TPA-converting module to perform different functions can accelerate the degradation of PET and realize the complete conversion of it [149]. Artificial microbial consortia can couple the depolymerization of PET by secreted enzymes with the biosynthesis of high value chemicals from monomers, which is a promising strategy in realizing the circular economy of PET waste [106].
5. Conclusions
This review summarized the current advances of PET biodegradation and bioconversion from the four aspects of engineered enzymes, chassis, pathways, and consortia, which provide a basis for the construction of artificial microbial consortia to convert PET into high value chemicals. Artificial microbial consortium is a promising strategy in realizing the circular economy of PET waste. On the one hand, the artificial microbial consortia are expected to effectively release the competitive inhibition of monomers in the PET biodegradation and improve the degradation efficiency. On the other hand, the artificial microbial consortia can couple the biodegradation of PET with the bioconversion of high value chemicals from monomers to realize circular economy and sustainability. Owing to the recent advancements in synthetic biology and metabolic engineering, it has now become possible to rationally design and create artificial microbial consortia with a superior metabolic efficiency to degrade PET and convert it into high value chemicals in one step.
Author Contributions
X.Q.: conceptualization, writing—original draft; W.Y.: writing—review and editing; Z.C.: writing—review and editing; M.D.: conceptualization, writing—review and editing, supervision; Y.Y.: supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Key Research and Development Program of China (2019YFA0706900).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mohanan, N.; Montazer, Z.; Sharma, P.K.; Levin, D.B. Microbial and enzymatic degradation of synthetic plastics. Front. Microbiol. 2020, 11, 2837. [Google Scholar] [CrossRef] [PubMed]
- Webb, H.K.; Arnott, J.; Crawford, R.J.; Ivanova, E.P. Plastic degradation and its environmental implications with special reference to Poly(ethylene terephthalate). Polymers 2013, 5, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Koshti, R.; Mehta, L.; Samarth, N. Biological recycling of polyethylene terephthalate: A mini-review. J. Polym. Environ. 2018, 26, 3520–3529. [Google Scholar] [CrossRef]
- Taniguchi, I.; Yoshida, S.; Hiraga, K.; Miyamoto, K.; Kimura, Y.; Oda, K. Biodegradation of PET: Current status and application aspects. ACS Catal. 2019, 9, 4089–4105. [Google Scholar] [CrossRef]
- Peng, R.; Xia, M.; Ru, J.; Huo, Y.; Yang, Y. Microbial degradation of polyurethane plastics. Chin. J. Biotechnol. 2018, 34, 1398–1409. [Google Scholar] [CrossRef]
- Ciobanu, C.S.; Copae, R.; Bulgariu, D.; Bulgariu, L. Comparative study of Pb(II) ions adsorption on pet fibers and flakes: Isotherm, kinetic and mechanism considerations. Desalination Water Treat. 2021, 222, 375–385. [Google Scholar] [CrossRef]
- Yang, H.; Chen, G.; Wang, J. Microplastics in the marine environment: Sources, fates, impacts and microbial degradation. Toxics 2021, 9, 41. [Google Scholar] [CrossRef]
- Wei, R.; Zimmermann, W. Biocatalysis as a green route for recycling the recalcitrant plastic polyethylene terephthalate. Microb. Biotechnol. 2017, 10, 1302–1307. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, W.; Billig, S. Enzymes for the biofunctionalization of Poly(Ethylene Terephthalate). In Biofunctionalization of Polymers and Their Applications; Advances in Biochemical Engineering—Biotechnology; Nyanhongo, G.S., Steiner, W., Gubitz, G.M., Eds.; Springer: Cham, Switzerland, 2011; Volume 125, pp. 97–120. [Google Scholar]
- Tokiwa, Y.; Suzuki, T. Hydrolysis of polyesters by lipases. Nature 1977, 270, 76–78. [Google Scholar] [CrossRef]
- Magalhaes, R.P.; Cunha, J.M.; Sousa, S.F. Perspectives on the role of enzymatic biocatalysis for the degradation of plastic PET. Int. J. Mol. Sci. 2021, 22, 11257. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Hiraga, K.; Takehana, T.; Taniguchi, I.; Yamaji, H.; Maeda, Y.; Toyohara, K.; Miyamoto, K.; Kimura, Y.; Oda, K. A bacterium that degrades and assimilates poly(ethylene terephthalate). Science 2016, 351, 1196–1199. [Google Scholar] [CrossRef] [PubMed]
- Biundo, A.; Ribitsch, D.; Guebitz, G.M. Surface engineering of polyester-degrading enzymes to improve efficiency and tune specificity. Appl. Microbiol. Biotechnol. 2018, 102, 3551–3559. [Google Scholar] [CrossRef]
- Palm, G.J.; Reisky, L.; Boettcher, D.; Mueller, H.; Michels, E.A.P.; Walczak, M.C.; Berndt, L.; Weiss, M.S.; Bornscheuer, U.T.; Weber, G. Structure of the plastic-degrading Ideonella sakaiensis MHETase bound to a substrate. Nat. Commun. 2019, 10, 1717. [Google Scholar] [CrossRef]
- Austin, H.P.; Allen, M.D.; Donohoe, B.S.; Rorrer, N.A.; Kearns, F.L.; Silveira, R.L.; Pollard, B.C.; Dominick, G.; Duman, R.; El Omari, K.; et al. Characterization and engineering of a plastic-degrading aromatic polyesterase. Proc. Natl. Acad. Sci. USA 2018, 115, E4350–E4357. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Liu, W.; Huang, J.-W.; Ma, J.; Zheng, Y.; Ko, T.-P.; Xu, L.; Cheng, Y.-S.; Chen, C.-C.; Guo, R.-T. Structural insight into catalytic mechanism of PET hydrolase. Nat. Commun. 2017, 8, 382. [Google Scholar] [CrossRef] [Green Version]
- Joo, S.; Cho, I.J.; Seo, H.; Son, H.F.; Sagong, H.-Y.; Shin, T.J.; Choi, S.Y.; Lee, S.Y.; Kim, K.-J. Structural insight into molecular mechanism of poly (ethylene terephthalate) degradation. Nat. Commun. 2018, 9, 2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, F.; Kawabata, T.; Oda, M. Current knowledge on enzymatic PET degradation and its possible application to waste stream management and other fields. Appl. Microbiol. Biotechnol. 2019, 103, 4253–4268. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, S.; Sharma, B.; Shukla, P. Integrated approaches in microbial degradation of plastics. Environ. Technol. Innov. 2020, 17, 100567. [Google Scholar] [CrossRef]
- Sagong, H.-Y.; Seo, H.; Kim, T.; Son, H.F.; Joo, S.; Lee, S.H.; Kim, S.; Woo, J.-S.; Hwang, S.Y.; Kim, K.-J. Decomposition of the PET Film by MHETase Using Exo-PETase Function. Acs Catal. 2020, 10, 4805–4812. [Google Scholar] [CrossRef]
- Ru, J.; Huo, Y.; Yang, Y. Microbial degradation and valorization of plastic wastes. Front. Microbiol. 2020, 11, 442. [Google Scholar] [CrossRef] [Green Version]
- Child, J.; Willetts, A. Microbial metabolism of aliphatic glycols. Bacterial metabolism of ethylene glycol. Biochim. Biophys. Acta 1978, 538, 316–327. [Google Scholar] [CrossRef]
- Kataoka, M.; Sasaki, M.; Hidalgo, A.; Nakano, M.; Shimizu, S. Glycolic acid production using ethylene glycol-oxidizing microorganisms. Biosci. Biotechnol. Biochem. 2001, 65, 2265–2270. [Google Scholar] [CrossRef] [Green Version]
- Wei, G.; Yang, X.; Zhou, W.; Lin, J.; Wei, D. Adsorptive bioconversion of ethylene glycol to glycolic acid by Gluconobacter oxydans DSM 2003. Biochem. Eng. J. 2009, 47, 127–131. [Google Scholar] [CrossRef]
- Trifunovic, D.; Schuchmann, K.; Mueller, V. Ethylene glycol metabolism in the acetogen Acetobacterium woodii. J. Bacteriol. 2016, 198, 1058–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panda, S.; Fung, V.Y.K.; Zhou, J.F.J.; Liang, H.; Zhou, K. Improving ethylene glycol utilization in Escherichia coli fermentation. Biochem. Eng. J. 2021, 168. [Google Scholar] [CrossRef]
- Sasoh, M.; Masai, E.; Ishibashi, S.; Hara, H.; Kamimura, N.; Miyauchi, K.; Fukuda, M. Characterization of the terephthalate degradation genes of Comamonas sp. strain E6. Appl. Environ. Microbiol. 2006, 72, 1825–1832. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.Y.; Kim, D.; Sul, W.J.; Chae, J.C.; Zylstra, G.J.; Kim, Y.M.; Kim, E. Molecular and biochemical analysis of phthalate and terephthalate degradation by Rhodococcus sp strain DK17. Fems Microbiol. Lett. 2005, 252, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Hosaka, M.; Kamimura, N.; Toribami, S.; Mori, K.; Kasai, D.; Fukuda, M.; Masai, E. Novel Tripartite aromatic acid transporter essential for terephthalate uptake in Comamonas sp. Strain E6. Appl. Environ. Microbiol. 2013, 79, 6148–6155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, M.; Oeser, T.; Wei, R.; Then, J.; Schmidt, J.; Zimmermann, W. Effect of hydrolysis products on the enzymatic degradation of polyethylene terephthalate nanoparticles by a polyester hydrolase from Thermobifida fusca. Biochem. Eng. J. 2015, 93, 222–228. [Google Scholar] [CrossRef]
- Vertommen, M.; Nierstrasz, V.A.; van der Veer, M.; Warmoeskerken, M. Enzymatic surface modification of poly(ethylene terephthalate). J. Biotechnol. 2005, 120, 376–386. [Google Scholar] [CrossRef]
- Muller, R.J.; Schrader, H.; Profe, J.; Dresler, K.; Deckwer, W.D. Enzymatic degradation of poly(ethylene terephthalate): Rapid hydrolyse using a hydrolase from T-fusca. Macromol. Rapid Commun. 2005, 26, 1400–1405. [Google Scholar] [CrossRef]
- Eberl, A.; Heumann, S.; Brueckner, T.; Araujo, R.; Cavaco-Paulo, A.; Kaufmann, F.; Kroutil, W.; Guebitz, G.M. Enzymatic surface hydrolysis of poly(ethylene terephthalate) and bis(benzoyloxyethyl) terephthalate by lipase and cutinase in the presence of surface active molecules. J. Biotechnol. 2009, 143, 207–212. [Google Scholar] [CrossRef]
- Nechwatal, A.; Blokesch, A.; Nicolai, M.; Krieg, M.; Kolbe, A.; Wolf, M.; Gerhardt, M. A contribution to the investigation of enzyme-catalysed hydrolysis of poly(ethylene terephthalate) oligomers. Macromol. Mater. Eng. 2006, 291, 1486–1494. [Google Scholar] [CrossRef]
- Shirke, A.N.; White, C.; Englaender, J.A.; Zwarycz, A.; Butterfoss, G.L.; Linhardt, R.J.; Gross, R.A. Stabilizing leaf and branch compost Cutinase (LCC) with glycosylation: Mechanism and effect on PET hydrolysis. Biochemistry 2018, 57, 1190–1200. [Google Scholar] [CrossRef]
- Acero, E.H.; Ribitsch, D.; Steinkellner, G.; Gruber, K.; Greimel, K.; Eiteljoerg, I.; Trotscha, E.; Wei, R.; Zimmermann, W.; Zinn, M.; et al. Enzymatic surface hydrolysis of PET: Effect of structural diversity on kinetic properties of Cutinases from thermobifida. Macromolecules 2011, 44, 4632–4640. [Google Scholar] [CrossRef] [Green Version]
- Kawai, F.; Oda, M.; Tamashiro, T.; Waku, T.; Tanaka, N.; Yamamoto, M.; Mizushima, H.; Miyakawa, T.; Tanokura, M. A novel Ca2+-activated, thermostabilized polyesterase capable of hydrolyzing polyethylene terephthalate from Saccharomonospora viridis AHK190. Appl. Microbiol. Biotechnol. 2014, 98, 10053–10064. [Google Scholar] [CrossRef] [PubMed]
- Then, J.; Wei, R.; Oeser, T.; Barth, M.; Belisario-Ferrari, M.R.; Schmidt, J.; Zimmermann, W. Ca2+ and Mg2+ binding site engineering increases the degradation of polyethylene terephthalate films by polyester hydrolases from Thermobifida fusca. Biotechnol. J. 2015, 10, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Breite, D.; Song, C.; Graesing, D.; Ploss, T.; Hille, P.; Schwerdtfeger, R.; Matysik, J.; Schulze, A.; Zimmermann, W. Biocatalytic Degradation efficiency of postconsumer polyethylene terephthalate packaging determined by their polymer microstructures. Adv. Sci. 2019, 6, 1900491. [Google Scholar] [CrossRef] [Green Version]
- Ronkvist, A.M.; Xie, W.; Lu, W.; Gross, R.A. Cutinase-catalyzed hydrolysis of Poly(ethylene terephthalate). Macromolecules 2009, 42, 5128–5138. [Google Scholar] [CrossRef]
- Vazquez-Alcantara, L.; Oliart-Ros, R.M.; Garcia-Borquez, A.; Pena-Montes, C. Expression of a Cutinase of Moniliophthora roreri with Polyester and PET-plastic residues degradation activity. Microbiol. Spectr. 2021, 9, e0097621. [Google Scholar] [CrossRef]
- Xi, X.; Ni, K.; Hao, H.; Shang, Y.; Zhao, B.; Qian, Z. Secretory expression in Bacillus subtilis and biochemical characterization of a highly thermostable polyethylene terephthalate hydrolase from bacterium HR29. Enzym. Microb. Technol. 2021, 143, 109715. [Google Scholar] [CrossRef] [PubMed]
- Billig, S.; Oeser, T.; Birkemeyer, C.; Zimmermann, W. Hydrolysis of cyclic poly(ethylene terephthalate) trimers by a carboxylesterase from Thermobifida fusca KW3. Appl. Microbiol. Biotechnol. 2010, 87, 1753–1764. [Google Scholar] [CrossRef] [PubMed]
- Ribitsch, D.; Acero, E.H.; Greimel, K.; Dellacher, A.; Zitzenbacher, S.; Marold, A.; Rodriguez, R.D.; Steinkellner, G.; Gruber, K.; Schwab, H.; et al. A new esterase from Thermobifida halotolerans hydrolyses polyethylene terephthalate (PET) and polylactic acid (PLA). Polymers 2012, 4, 617–629. [Google Scholar] [CrossRef]
- Biundo, A.; Reich, J.; Ribitsch, D.; Guebitz, G.M. Synergistic effect of mutagenesis and truncation to improve a polyesterase from Clostridium botulinum for polyester hydrolysis. Sci. Rep. 2018, 8, 3745. [Google Scholar] [CrossRef] [Green Version]
- Biundo, A.; Braunschmid, V.; Pretzler, M.; Kampatsikas, I.; Darnhofer, B.; Birner-Gruenberger, R.; Rompel, A.; Ribitsch, D.; Guebitz, G.M. Polyphenol oxidases exhibit promiscuous proteolytic activity. Commun. Chem. 2020, 3, 62. [Google Scholar] [CrossRef]
- Wei, R.; Oeser, T.; Barth, M.; Weigl, N.; Luebs, A.; Schulz-Siegmund, M.; Hacker, M.C.; Zimmermann, W. Turbidimetric analysis of the enzymatic hydrolysis of polyethylene terephthalate nanoparticles. J. Mol. Catal. B-Enzym. 2014, 103, 72–78. [Google Scholar] [CrossRef]
- Liu, C.; Shi, C.; Zhu, S.; Wei, R.; Yin, C.-C. Structural and functional characterization of polyethylene terephthalate hydrolase from Ideonella sakaiensis. Biochem. Biophys. Res. Commun. 2019, 508, 289–294. [Google Scholar] [CrossRef]
- Liu, B.; He, L.; Wang, L.; Li, T.; Li, C.; Liu, H.; Luo, Y.; Bao, R. Protein crystallography and site-direct mutagenesis analysis of the Poly(ethylene terephthalate) hydrolase PETase from Ideonella sakaiensis. Chembiochem 2018, 19, 1471–1475. [Google Scholar] [CrossRef]
- Ma, Y.; Yao, M.D.; Li, B.Z.; Ding, M.Z.; He, B.; Chen, S.; Zhou, X.; Yuan, Y.J. Enhanced Poly(ethylene terephthalate) hydrolase activity by protein engineering. Engineering 2018, 4, 888–893. [Google Scholar] [CrossRef]
- Silva, C.; Da, S.; Silva, N.; Matama, T.; Araujo, R.; Martins, M.; Chen, S.; Chen, J.; Wu, J.; Casal, M.; et al. Engineered Thermobifida fusca cutinase with increased activity on polyester substrates. Biotechnol. J. 2011, 6, 1230–1239. [Google Scholar] [CrossRef]
- Chen, C.C.; Han, X.; Li, X.; Jiang, P.C.; Niu, D.; Ma, L.X.; Liu, W.D.; Li, S.Y.; Qu, Y.Y.; Hu, H.B.; et al. General features to enhance enzymatic activity of poly(ethylene terephthalate) hydrolysis. Nat. Catal. 2021, 4, 425–430. [Google Scholar] [CrossRef]
- Tournier, V.; Topham, C.M.; Gilles, A.; David, B.; Folgoas, C.; Moya-Leclair, E.; Kamionka, E.; Desrousseaux, M.L.; Texier, H.; Gavalda, S.; et al. An engineered PET depolymerase to break down and recycle plastic bottles. Nature 2020, 580, 216–219. [Google Scholar] [CrossRef]
- Cui, Y.; Chen, Y.; Liu, X.; Dong, S.; Tian, Y.E.; Qiao, Y.; Mitra, R.; Han, J.; Li, C.; Han, X.; et al. Computational redesign of a PETase for plastic biodegradation under ambient condition by the GRAPE strategy. Acs Catal. 2021, 11, 1340–1350. [Google Scholar] [CrossRef]
- Thumarat, U.; Nakamura, R.; Kawabata, T.; Suzuki, H.; Kawai, F. Biochemical and genetic analysis of a cutinase-type polyesterase from a thermophilic Thermobifida alba AHK119. Appl. Microbiol. Biotechnol. 2012, 95, 419–430. [Google Scholar] [CrossRef]
- Then, J.; Wei, R.; Oeser, T.; Gerdts, A.; Schmidt, J.; Barth, M.; Zimmermann, W. A disulfide bridge in the calcium binding site of a polyester hydrolase increases its thermal stability and activity against polyethylene terephthalate. Febs Open Bio 2016, 6, 425–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirke, A.N.; Basore, D.; Holton, S.; Su, A.; Baugh, E.; Butterfoss, G.L.; Makhatadze, G.; Bystroff, C.; Gross, R.A. Influence of surface charge, binding site residues and glycosylation on Thielavia terrestris cutinase biochemical characteristics. Appl. Microbiol. Biotechnol. 2016, 100, 4435–4446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linder, M.B.; Szilvay, G.R.; Nakari-Setala, T.; Penttila, M.E. Hydrophobins: The protein-amphiphiles of filamentous fungi. Fems Microbiol. Rev. 2005, 29, 877–896. [Google Scholar] [CrossRef] [Green Version]
- Puspitasari, N.; Tsai, S.-L.; Lee, C.-K. Fungal Hydrophobin RolA enhanced PETase hydrolysis of polyethylene terephthalate. Appl. Biochem. Biotechnol. 2021, 193, 1284–1295. [Google Scholar] [CrossRef]
- Ribitsch, D.; Yebra, A.O.; Zitzenbacher, S.; Wu, J.; Nowitsch, S.; Steinkellner, G.; Greimel, K.; Doliska, A.; Oberdorfer, G.; Gruber, C.C.; et al. Fusion of binding domains to Thermobifida cellulosilytica Cutinase to tune sorption characteristics and enhancing PET hydrolysis. Biomacromolecules 2013, 14, 1769–1776. [Google Scholar] [CrossRef]
- Ribitsch, D.; Acero, E.H.; Przylucka, A.; Zitzenbacher, S.; Marold, A.; Gamerith, C.; Tscheliessnig, R.; Jungbauer, A.; Rennhofer, H.; Lichtenegger, H.; et al. Enhanced cutinase-catalyzed hydrolysis of polyethylene terephthalate by covalent fusion to hydrophobins. Appl. Environ. Microbiol. 2015, 81, 3586–3592. [Google Scholar] [CrossRef] [Green Version]
- Carniel, A.; Valoni, E.; Nicomedes Junior, J.; Gomes, A.d.C.; de Castro, A.M. Lipase from Candida antarctica (CALB) and cutinase from Humicola insolens act synergistically for PET hydrolysis to terephthalic acid. Process Biochem. 2017, 59, 84–90. [Google Scholar] [CrossRef]
- Wei, R.; Oeser, T.; Schmidt, J.; Meier, R.; Barth, M.; Then, J.; Zimmermann, W. Engineered bacterial polyester hydrolases efficiently degrade polyethylene terephthalate due to relieved product inhibition. Biotechnol. Bioeng. 2016, 113, 1658–1665. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.J.J.; Gao, W.; Rong, Q.X.; Jin, G.J.; Chu, H.Y.; Liu, W.J.; Yang, W.; Zhu, Z.W.; Li, G.H.; Zhu, G.F.; et al. Modular Pathway Engineering of diterpenoid synthases and the mevalonic acid pathway for miltiradiene production. J. Am. Chem. Soc. 2012, 134, 3234–3241. [Google Scholar] [CrossRef]
- Furukawa, M.; Kawakami, N.; Oda, K.; Miyamoto, K. Acceleration of enzymatic degradation of Poly(ethylene terephthalate) by surface coating with anionic surfactants. Chemsuschem 2018, 11, 4018–4025. [Google Scholar] [CrossRef]
- Araujo, R.; Silva, C.; O’Neill, A.; Micaelo, N.; Guebitz, G.; Soares, C.M.; Casal, M.; Cavaco-Paulo, A. Tailoring cutinase activity towards polyethylene terephthalate and polyamide 6,6 fibers. J. Biotechnol. 2007, 128, 849–857. [Google Scholar] [CrossRef] [Green Version]
- Kawabata, T.; Oda, M.; Kawai, F. Mutational analysis of cutinase-like enzyme, Cut190, based on the 3D docking structure with model compounds of polyethylene terephthalate. J. Biosci. Bioeng. 2017, 124, 28–35. [Google Scholar] [CrossRef]
- Joo, S.; Lee, S.; Kim, K. Structural insight into molecular mechanism of poly(ethylene terephthalate) degradation. FEBS J. 2017, 284, 309. [Google Scholar] [CrossRef] [Green Version]
- Bollinger, A.; Thies, S.; Knieps-Gruenhagen, E.; Gertzen, C.; Kobus, S.; Hoeppner, A.; Ferrer, M.; Gohlke, H.; Smits, S.H.J.; Jaeger, K.-E. A novel polyester hydrolase from the marine bacterium Pseudomonas aestusnigri—Structural and functional insights. Front. Microbiol. 2020, 11, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oda, M.; Yamagami, Y.; Inaba, S.; Oida, T.; Yamamoto, M.; Kitajima, S.; Kawai, F. Enzymatic hydrolysis of PET: Functional roles of three Ca2+ ions bound to a cutinase-like enzyme, Cut190*, and its engineering for improved activity. Appl. Microbiol. Biotechnol. 2018, 102, 10067–10077. [Google Scholar] [CrossRef] [PubMed]
- Son, H.F.; Cho, I.J.; Joo, S.; Seo, H.; Sagong, H.-Y.; Choi, S.Y.; Lee, S.Y.; Kim, K.-J. Rational protein engineering of thermo-stable PETase from Ideonella sakaiensis for highly efficient PET degradation. Acs Catal. 2019, 9, 3519–3526. [Google Scholar] [CrossRef]
- Son, H.F.; Joo, S.; Seo, H.; Sagong, H.-Y.; Lee, S.H.; Hong, H.; Kim, K.-J. Structural bioinformatics-based protein engineering of thermo-stable PETase from Ideonella sakaiensis. Enzym. Microb. Technol. 2020, 141, 109656. [Google Scholar] [CrossRef] [PubMed]
- Biundo, A.; Ribitsch, D.; Steinkellner, G.; Gruber, K.; Guebitz, G.M. Polyester hydrolysis is enhanced by a truncated esterase: Less is more. Biotechnol. J. 2017, 12, 109656. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, M.; Kawakami, N.; Tomizawa, A.; Miyamoto, K. Efficient degradation of Poly(ethylene terephthalate) with Thermobifida fusca Cutinase Exhibiting improved catalytic activity generated using mutagenesis and additive-based approaches. Sci. Rep. 2019, 9, 16038. [Google Scholar] [CrossRef] [Green Version]
- Barth, M.; Honak, A.; Oeser, T.; Wei, R.; Belisário-Ferrari, M.R.; Then, J.; Schmidt, J.; Zimmermann, W. A dual enzyme system composed of a polyester hydrolase and a carboxylesterase enhances the biocatalytic degradation of polyethylene terephthalate films. Biotechnol. J. 2016, 11, 1082–1087. [Google Scholar] [CrossRef]
- Samak, N.A.; Jia, Y.; Sharshar, M.M.; Mu, T.; Yang, M.; Peh, S.; Xing, J. Recent advances in biocatalysts engineering for polyethylene terephthalate plastic waste green recycling. Environ. Int. 2020, 145, 106144. [Google Scholar] [CrossRef]
- Gao, R.; Pan, H.; Lian, J. Recent advances in the discovery, characterization, and engineering of poly (ethylene terephthalate) (PET) hydrolases. Enzym. Microb. Technol. 2021, 150, 109868. [Google Scholar] [CrossRef]
- Serra, S.; De Simeis, D. Use of Lactobacillus rhamnosus (ATCC 53103) as whole-cell biocatalyst for the regio- and stereoselective hydration of oleic, linoleic, and linolenic acid. Catalysts 2018, 8, 109. [Google Scholar] [CrossRef] [Green Version]
- Polak, J.; Jarosz-Wilkolazka, A. Whole-cell fungal transformation of precursors into dyes. Microb. Cell Factories 2010, 9, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, H.; Kim, S.; Son, H.F.; Sagong, H.-Y.; Joo, S.; Kim, K.-J. Production of extracellular PETase from Ideonella sakaiensis using sec-dependent signal peptides in E. coli. Biochem. Biophys. Res. Commun. 2019, 508, 250–255. [Google Scholar] [CrossRef]
- Shi, L.; Liu, H.; Gao, S.; Weng, Y.; Zhu, L. Enhanced extracellular production of IsPETase in Escherichia coli via engineering of the pelB signal peptide. J. Agric. Food Chem. 2021, 69, 2245–2252. [Google Scholar] [CrossRef]
- Cui, L.; Qiu, Y.; Liang, Y.; Du, C.; Dong, W.; Cheng, C.; He, B. Excretory expression of IsPETase in E. coli by an enhancer of signal peptides and enhanced PET hydrolysis. Int. J. Biol. Macromol. 2021, 188, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Cao, L.; Qin, Z.; Li, S.; Kong, W.; Liu, Y. Tat-independent secretion of polyethylene terephthalate hydrolase PETase in Bacillus subtilis 168 mediated by its native signal peptide. J. Agric. Food Chem. 2018, 66, 13217–13227. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Guan, F.; Lv, X.; Han, D.; Zhang, Y.; Wu, N.; Xia, X.; Tian, J. Enhancing secretion of polyethylene terephthalate hydrolase PETase in Bacillus subtilis WB600 mediated by the SP(amy)signal peptide. Lett. Appl. Microbiol. 2020, 71, 235–241. [Google Scholar] [CrossRef]
- Yan, F.; Wei, R.; Cui, Q.; Bornscheuer, U.T.; Liu, Y.-J. Thermophilic whole-cell degradation of polyethylene terephthalate using engineered Clostridium thermocellum. Microb. Biotechnol. 2021, 14, 374–385. [Google Scholar] [CrossRef]
- Chen, Z.Z.; Wang, Y.Y.; Cheng, Y.Y.; Wang, X.; Tong, S.W.; Yang, H.T.; Wang, Z.F. Efficient biodegradation of highly crystallized polyethylene terephthalate through cell surface display of bacterial PETase. Sci. Total Environ. 2020, 709, 136138. [Google Scholar] [CrossRef]
- Moog, D.; Schmitt, J.; Senger, J.; Zarzycki, J.; Rexer, K.-H.; Linne, U.; Erb, T.; Maier, U.G. Using a marine microalga as a chassis for polyethylene terephthalate (PET) degradation. Microb. Cell Factories 2019, 18, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.W.; Park, S.-B.; Tran, Q.-G.; Cho, D.-H.; Choi, D.-Y.; Lee, Y.J.; Kim, H.-S. Functional expression of polyethylene terephthalate-degrading enzyme (PETase) in green microalgae. Microb. Cell Factories 2020, 19, 97. [Google Scholar] [CrossRef] [PubMed]
- Touchon, M.; Hoede, C.; Tenaillon, O.; Barbe, V.; Baeriswyl, S.; Bidet, P.; Bingen, E.; Bonacorsi, S.; Bouchier, C.; Bouvet, O.; et al. Organised genome dynamics in the escherichia coli species results in highly diverse adaptive paths. PLoS Genet. 2009, 5, e1000344. [Google Scholar] [CrossRef] [Green Version]
- Van Dijl, J.M.; Hecker, M. Bacillus subtilis: From soil bacterium to super-secreting cell factory. Microb. Cell Factories 2013, 12, 3. [Google Scholar] [CrossRef] [Green Version]
- Westers, L.; Westers, H.; Quax, W.J. Bacillus subtilis as cell factory for pharmaceutical proteins: A biotechnological approach to optimize the host organism. Biochim. Biophys. Acta Mol. Cell Res. 2004, 1694, 299–310. [Google Scholar] [CrossRef]
- Huang, K.; Chen, C.; Shen, Q.; Rosen, B.P.; Zhao, F.-J. Genetically Engineering bacillus subtilis with a heat-resistant arsenite methyltransferase for bioremediation of arsenic-contaminated organic waste. Appl. Environ. Microbiol. 2015, 81, 6718–6724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Song, C.; Freudl, R.; Mulchandani, A.; Qiao, C. Twin-arginine translocation of methyl parathion hydrolase in Bacillus subtilis. Environ. Sci. Technol. 2010, 44, 7607–7612. [Google Scholar] [CrossRef]
- Ng, T.K.; Weimer, T.K.; Zeikus, J.G. Cellulolytic and physiological properties of Clostridium thermocellum. Arch. Microbiol. 1977, 114, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Guyot, S.; Pottier, L.; Hartmann, A.; Ragon, M.; Tiburski, J.H.; Molin, P.; Ferret, E.; Gervais, P. Extremely rapid acclimation of Escherichia coli to high temperature over a few generations of a fed-batch culture during slow warming. Microbiologyopen 2014, 3, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.P.; Mi, L.; Moriok, A.H.; Yoshino, M.M.; Konishi, S.; Xu, S.C.; Papanek, B.A.; Riley, L.A.; Guss, A.M.; Liao, J.C. Consolidated bioprocessing of cellulose to isobutanol using Clostridium thermocellum. Metab. Eng. 2015, 31, 44–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argyros, D.A.; Tripathi, S.A.; Barrett, T.F.; Rogers, S.R.; Feinberg, L.F.; Olson, D.G.; Foden, J.M.; Miller, B.B.; Lynd, L.R.; Hogsett, D.A.; et al. High ethanol titers from cellulose by using metabolically engineered thermophilic, anaerobic microbes. Appl. Environ. Microbiol. 2011, 77, 8288–8294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sooch, B.S.; Kauldhar, B.S.; Puri, M. Isolation and polyphasic characterization of a novel hyper catalase producing thermophilic bacterium for the degradation of hydrogen peroxide. Bioprocess Biosyst. Eng. 2016, 39, 1759–1773. [Google Scholar] [CrossRef]
- Rudolph, B.; Gebendorfer, K.M.; Buchner, J.; Winter, J. Evolution of Escherichia coli for growth at high temperatures. J. Biol. Chem. 2010, 285, 19029–19034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.F.; Wang, X.C.; Gong, J.X.; Gu, Z.Y. A study on the biodegradability of polyethylene terephthalate fiber and diethylene glycol terephthalate. J. Appl. Polym. Sci. 2004, 93, 1089–1096. [Google Scholar] [CrossRef]
- Gong, J.; Duan, N.; Zhao, X. Evolutionary engineering of Phaffia rhodozyma for astaxanthin-overproducing strain. Front. Chem. Sci. Eng. 2012, 6, 174–178. [Google Scholar] [CrossRef]
- Gong, J.; Kong, T.; Li, Y.; Li, Q.; Li, Z.; Zhang, J. Biodegradation of microplastic derived from Poly(ethylene terephthalate) with bacterial whole-cell biocatalysts. Polymers 2018, 10, 1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Cen, Y.-K.; Zou, S.-P.; Xue, Y.-P.; Zheng, Y.-G. Recent advances in the improvement of enzyme thermostability by structure modification. Crit. Rev. Biotechnol. 2020, 40, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Madzak, C. Yarrowia lipolytica: Recent achievements in heterologous protein expression and pathway engineering. Appl. Microbiol. Biotechnol. 2015, 99, 4559–4577. [Google Scholar] [CrossRef]
- Da Costa, A.M.; de Oliveira Lopes, V.R.; Vidal, L.; Nicaud, J.-M.; de Castro, A.M.; Zarur Coelho, M.A. Poly(ethylene terephthalate) (PET) degradation by Yarrowia lipolytica: Investigations on cell growth, enzyme production and monomers consumption. Process Biochem. 2020, 95, 81–90. [Google Scholar] [CrossRef]
- Liu, P.; Zhang, T.; Zheng, Y.; Li, Q.; Su, T.; Qi, Q. Potential one-step strategy for PET degradation and PHB biosynthesis through co-cultivation of two engineered microorganisms. Eng. Microbiol. 2021, 1, 100003. [Google Scholar] [CrossRef]
- Lebreton, S.; Zurzolo, C.; Paladino, S. Organization of GPI-anchored proteins at the cell surface and its physiopathological relevance. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 403–419. [Google Scholar] [CrossRef]
- Yang, C.-E.; Chu, I.M.; Wei, Y.-H.; Tsai, S.-L. Surface display of synthetic phytochelatins on Saccharomyces cerevisiae for enhanced ethanol production in heavy metal-contaminated substrates. Bioresour. Technol. 2017, 245, 1455–1460. [Google Scholar] [CrossRef]
- Beopoulos, A.; Cescut, J.; Haddouche, R.; Uribelarrea, J.-L.; Molina-Jouve, C.; Nicaud, J.-M. Yarrowia lipolytica as a model for bio-oil production. Progress Lipid Res. 2009, 48, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Barone, G.D.; Ferizovic, D.; Biundo, A.; Lindblad, P. Hints at the applicability of Microalgae and Cyanobacteria for the biodegradation of plastics. Sustainability 2020, 12, 449. [Google Scholar] [CrossRef]
- Bertsch, J.; Siemund, A.L.; Kremp, F.; Mueller, V. A novel route for ethanol oxidation in the acetogenic bacterium Acetobacterium woodii: The acetaldehyde/ethanol dehydrogenase pathway. Environ. Microbiol. 2016, 18, 2913–2922. [Google Scholar] [CrossRef]
- Chowdhury, N.P.; Alberti, L.; Linder, M.; Mueller, V. Exploring bacterial microcompartments in the acetogenic Bacterium Acetobacterium woodii. Front. Microbiol. 2020, 11, 593467. [Google Scholar] [CrossRef] [PubMed]
- Salvador, M.; Abdulmutalib, U.; Gonzalez, J.; Kim, J.; Smith, A.A.; Faulon, J.-L.; Wei, R.; Zimmermann, W.; Jimenez, J.I. Microbial genes for a circular and sustainable Bio-PET economy. Genes 2019, 10, 373. [Google Scholar] [CrossRef] [Green Version]
- Wehrmann, M.; Billard, P.; Martin-Meriadec, A.; Zegeye, A.; Klebensberger, J. Functional role of lanthanides in enzymatic activity and transcriptional regulation of pyrroloquinoline quinone-dependent alcohol dehydrogenases in pseudomonas putida KT2440. Mbio 2017, 8, e00570-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franden, M.A.; Jayakody, L.N.; Li, W.J.; Wagner, N.J.; Cleveland, N.S.; Michener, W.E.; Hauer, B.; Blank, L.M.; Wierckx, N.; Klebensberger, J.; et al. Engineering pseudomonas putida KT2440 for efficient ethylene glycol utilization. Metab. Eng. 2018, 48, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-J.; Jayakody, L.N.; Franden, M.A.; Wehrmann, M.; Daun, T.; Hauer, B.; Blank, L.M.; Beckham, G.T.; Klebensberger, J.; Wierckx, N. Laboratory evolution reveals the metabolic and regulatory basis of ethylene glycol metabolism by Pseudomonas putida KT2440. Environ. Microbiol. 2019, 21, 3669–3682. [Google Scholar] [CrossRef] [PubMed]
- Muckschel, B.; Simon, O.; Klebensberger, J.; Graf, N.; Rosche, B.; Altenbuchner, J.; Pfannstiel, J.; Huber, A.; Hauer, B. Ethylene glycol metabolism by Pseudomonas putida. Appl. Environ. Microbiol. 2012, 78, 8531–8539. [Google Scholar] [CrossRef] [Green Version]
- Boronat, A.; Caballero, E.; Aguilar, J. Experimental evolution of a metabolic pathway for ethylene glycol utilization by Escherichia coli. J. Bacteriol. 1983, 153, 134–139. [Google Scholar] [CrossRef] [Green Version]
- Pandit, A.V.; Harrison, E.; Mahadevan, R. Engineering Escherichia coli for the utilization of ethylene glycol. Microb. Cell Factories 2021, 20, 22. [Google Scholar] [CrossRef]
- Gao, X.; Ma, Z.; Yang, L.; Ma, J. Enhanced Bioconversion of ethylene glycol to glycolic acid by a newly isolated Burkholderia sp EG13. Appl. Biochem. Biotechnol. 2014, 174, 1572–1580. [Google Scholar] [CrossRef]
- Yamada-Onodera, K.; Nakajima, A.; Tani, Y. Purification, characterization, and gene cloning of glycerol dehydrogenase from Hansenula ofunaensis, and its expression for production of optically active diol. J. Biosci. Bioeng. 2006, 102, 545–551. [Google Scholar] [CrossRef]
- Zhang, H.; Shi, L.; Mao, X.; Lin, J.; Wei, D. Enhancement of cell growth and glycolic acid production by overexpression of membrane-bound alcohol dehydrogenase in Gluconobacter oxydans DSM 2003. J. Biotechnol. 2016, 237, 18–24. [Google Scholar] [CrossRef]
- Schweiger, P.; Gross, H.; Zeiser, J.; Deppenmeier, U. Asymmetric reduction of diketones by two Gluconobacter oxydans oxidoreductases. Appl. Microbiol. Biotechnol. 2013, 97, 3475–3484. [Google Scholar] [CrossRef]
- Salvachua, D.; Karp, E.M.; Nimlos, C.T.; Vardon, D.R.; Beckham, G.T. Towards lignin consolidated bioprocessing: Simultaneous lignin depolymerization and product generation by bacteria. Green Chem. 2015, 17, 4951–4967. [Google Scholar] [CrossRef]
- Cai, L.; Yuan, M.Q.; Liu, F.; Jian, J.; Chen, G.Q. Enhanced production of medium-chain-length polyhydroxyalkanoates (PHA) by PHA depolymerase knockout mutant of Pseudomonas putida KT2442. Bioresour. Technol. 2009, 100, 2265–2270. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Luo, G.; Zhou, X.R.; Chen, G.-Q. Biosynthesis of poly(3-hydroxydecanoate) and 3-hydroxydodecanoate dominating polyhydroxyalkanoates by beta-oxidation pathway inhibited Pseudomonas putida. Metab. Eng. 2011, 13, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Poblete-Castro, I.; Binger, D.; Rodrigues, A.; Becker, J.; dos Santos, V.A.P.M.; Wittmann, C. In-silico-driven metabolic engineering of Pseudomonas putida for enhanced production of poly-hydroxyalkanoates. Metab. Eng. 2013, 15, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Linger, J.G.; Vardon, D.R.; Guarnieri, M.T.; Karp, E.M.; Hunsinger, G.B.; Franden, M.A.; Johnson, C.W.; Chupka, G.; Strathmann, T.J.; Pienkos, P.T.; et al. Lignin valorization through integrated biological funneling and chemical catalysis. Proc. Natl. Acad. Sci. USA 2014, 111, 12013–12018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigematsu, T.; Yumihara, K.; Ueda, Y.; Morimura, S.; Kida, K. Purification and gene cloning of the oxygenase component of the terephthalate 1,2-dioxygenase system from Delftia tsuruhatensis strain T7. Fems Microbiol. Lett. 2003, 220, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Z.; Zhou, Y.; Zylstra, G.J. Molecular analysis of isophthalate and terephthalate degradation by Comamonas testosteroni YZW-D. Environ. Health Perspect. 1995, 103 (Suppl. S5), 9–12. [Google Scholar] [CrossRef] [Green Version]
- Ward, P.G.; de Roo, G.; O’Connor, K.E. Accumulation of polyhydroxyalkanoate from styrene and phenylacetic acid by Pseudomonas putida CA-3. Appl. Environ. Microbiol. 2005, 71, 2046–2052. [Google Scholar] [CrossRef] [Green Version]
- Ward, P.G.; Goff, M.; Donner, M.; Kaminsky, W.; O’Connor, K.E. A two step chemo-biotechnological conversion of polystyrene to a biodegradable thermoplastic. Environ. Sci. Technol. 2006, 40, 2433–2437. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.T.; Kim, J.K.; Cha, H.G.; Kang, M.J.; Lee, H.S.; Khang, T.U.; Yun, E.J.; Lee, D.-H.; Song, B.K.; Park, S.J.; et al. Biological valorization of Poly(ethylene terephthalate) monomers for upcycling waste PET. Acs Sustain. Chem. Eng. 2019, 7, 19396–19406. [Google Scholar] [CrossRef]
- Harwood, C.S.; Parales, R.E. The beta-ketoadipate pathway and the biology of self-identity. Annu. Rev. Microbiol. 1996, 50, 553–590. [Google Scholar] [CrossRef]
- Kasai, D.; Fujinami, T.; Abe, T.; Mase, K.; Katayama, Y.; Fukuda, M.; Masai, E. Uncovering the Protocatechuate 2,3-Cleavage Pathway Genes. J. Bacteriol. 2009, 191, 6758–6768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frazee, R.W.; Livingston, D.M.; LaPorte, D.C.; Lipscomb, J.D. Cloning, sequencing, and expression of the Pseudomonas putida protocatechuate 3,4-dioxygenase genes. J. Bacteriol. 1993, 175, 6194–6202. [Google Scholar] [CrossRef] [Green Version]
- Noda, Y.; Nishikawa, S.; Shiozuka, K.; Kadokura, H.; Nakajima, H.; Yoda, K.; Katayama, Y.; Morohoshi, N.; Haraguchi, T.; Yamasaki, M. Molecular cloning of the protocatechuate 4,5-dioxygenase genes of Pseudomonas paucimobilis. J. Bacteriol. 1990, 172, 2704–2709. [Google Scholar] [CrossRef] [Green Version]
- Wells, T.; Ragauskas, A.J. Biotechnological opportunities with the beta-ketoadipate pathway. Trends Biotechnol. 2012, 30, 627–637. [Google Scholar] [CrossRef]
- Werner, A.Z.; Clare, R.; Mand, T.D.; Pardo, I.; Ramirez, K.J.; Haugen, S.J.; Bratti, F.; Dexter, G.N.; Elmore, J.R.; Huenemann, J.D.; et al. Tandem chemical deconstruction and biological upcycling of poly(ethylene terephthalate) to beta-ketoadipic acid by Pseudomonas putida KT2440. Metab. Eng. 2021, 67, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Kunjapur, A.M.; Hyun, J.C.; Prather, K.L.J. Deregulation of S-adenosylmethionine biosynthesis and regeneration improves methylation in the E-coli de novo vanillin biosynthesis pathway. Microb. Cell Factories 2016, 15, 61. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Shen, X.; Wang, J.; Wang, J.; Yuan, Q.; Yan, Y. Rational engineering of p-Hydroxybenzoate hydroxylase to enable efficient gallic acid synthesis via a novel artificial biosynthetic pathway. Biotechnol. Bioeng. 2017, 114, 2571–2580. [Google Scholar] [CrossRef]
- Jimenez, N.; Antonio Curiel, J.; Reveron, I.; de las Rivas, B.; Munoz, R. Uncovering the Lactobacillus plantarum WCFS1 gallate decarboxylase involved in tannin degradation. Appl. Environ. Microbiol. 2013, 79, 4253–4263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckham, G.T.; Johnson, C.W.; Karp, E.M.; Salvachua, D.; Vardon, D.R. Opportunities and challenges in biological lignin valorization. Curr. Opin. Biotechnol. 2016, 42, 40–53. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Mawgoud, A.M.; Lepine, F.; Deziel, E. A Stereospecific pathway diverts beta-oxidation intermediates to the biosynthesis of rhamnolipid biosurfactants. Chem. Biol. 2014, 21, 156–164. [Google Scholar] [CrossRef]
- Tiso, T.; Narancic, T.; Wei, R.; Pollet, E.; Beagan, N.; Schroeder, K.; Honak, A.; Jiang, M.; Kenny, S.T.; Wierckx, N.; et al. Towards bio-upcycling of polyethylene terephthalate. Metab. Eng. 2021, 66, 167–178. [Google Scholar] [CrossRef]
- Hwang, K.R.; Jeon, W.; Lee, S.Y.; Kim, M.S.; Park, Y.K. Sustainable bioplastics: Recent progress in the production of bio-building blocks for the bio-based next-generation polymer PEF. Chem. Eng. J. 2020, 390, 124636. [Google Scholar] [CrossRef]
- Cywar, R.M.; Rorrer, N.A.; Hoyt, C.B.; Beckham, G.T.; Chen, E.Y.X. Bio-based polymers with performance-advantaged properties. Nat. Rev. Mater. 2021. [Google Scholar] [CrossRef]
- Dissanayake, L.; Jayakody, L.N. Engineering microbes to bio-upcycle Polyethylene Terephthalate. Front. Bioeng. Biotechnol. 2021, 9, 656465. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Ma, Y.; Chang, H.; Li, B.; Ding, M.; Yuan, Y. Evaluation of PET Degradation Using Artificial Microbial Consortia. Front. Microbiol. 2021, 12, 778828. [Google Scholar] [CrossRef]
- Ding, M.-Z.; Song, H.; Wang, E.-X.; Liu, Y.; Yuan, Y.-J. Design and construction of synthetic microbial consortia in China. Synth. Syst. Biotechnol. 2016, 1, 230–235. [Google Scholar] [CrossRef] [Green Version]
- Dangi, A.K.; Sharma, B.; Hill, R.T.; Shukla, P. Bioremediation through microbes: Systems biology and metabolic engineering approach. Crit. Rev. Biotechnol. 2019, 39, 79–98. [Google Scholar] [CrossRef] [PubMed]
- Carpio, I.E.M.; Franco, D.C.; Zanoli Sato, M.I.; Sakata, S.; Pellizari, V.H.; Ferreira Filho, S.S.; Rodrigues, D.F. Biostimulation of metal-resistant microbial consortium to remove zinc from contaminated environments. Sci. Total Environ. 2016, 550, 670–675. [Google Scholar] [CrossRef]
- Ibrar, M.; Zhang, H. Construction of a hydrocarbon-degrading consortium and characterization of two new lipopeptides biosurfactants. Sci. Total Environ. 2020, 714, 136400. [Google Scholar] [CrossRef] [PubMed]
- Utomo, R.N.C.; Li, W.-J.; Tiso, T.; Eberlein, C.; Doeker, M.; Heipieper, H.J.; Jupke, A.; Wierckx, N.; Blank, L.M. Defined microbial mixed culture for utilization of polyurethane monomers. Acs Sustain. Chem. Eng. 2020, 8, 17466–17474. [Google Scholar] [CrossRef]
- Skariyachan, S.; Taskeen, N.; Kishore, A.P.; Krishna, B.V.; Naidu, G. Novel consortia of enterobacter and pseudomonas formulated from cow dung exhibited enhanced biodegradation of polyethylene and polypropylene. J. Environ. Manag. 2021, 284, 112030. [Google Scholar] [CrossRef]
- Sun, X.; Chen, L.; Liu, C.; Xu, Y.; Ma, W.; Ni, H. Biodegradation of CP/TCP by a constructed microbial consortium after comparative bacterial community analysis of long-term CP domesticated activated sludge. J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes 2020, 55, 898–908. [Google Scholar] [CrossRef]
- Tang, X.; He, L.Y.; Tao, X.Q.; Dang, Z.; Guo, C.L.; Lu, G.N.; Yi, X.Y. Construction of an artificial microalgal-bacterial consortium that efficiently degrades crude oil. J. Hazard. Mater. 2010, 181, 1158–1162. [Google Scholar] [CrossRef]
- Wang, J.H.; Hlaing, T.S.; Nwe, M.T.; Aung, M.M.; Ren, C.; Wu, W.; Yan, Y.C. Primary biodegradation and mineralization of aryl organophosphate flame retardants by Rhodococcus-Sphingopyxis consortium. J. Hazard. Mater. 2021, 412, 125238. [Google Scholar] [CrossRef]
- Martinez, I.; Mohamed, M.E.S.; Rozas, D.; Garcia, J.L.; Diaz, E. Engineering synthetic bacterial consortia for enhanced desulfurization and revalorization of oil sulfur compounds. Metab. Eng. 2016, 35, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Skariyachan, S.; Manjunatha, V.; Sultana, S.; Jois, C.; Bai, V.; Vasist, K.S. Novel bacterial consortia isolated from plastic garbage processing areas demonstrated enhanced degradation for low density polyethylene. Environ. Sci. Pollut. Res. 2016, 23, 18307–18319. [Google Scholar] [CrossRef]
- Syranidou, E.; Karkanorachaki, K.; Amorotti, F.; Repouskou, E.; Kroll, K.; Kolvenbach, B.; Corvini, P.F.X.; Fava, F.; Kalogerakis, N. Development of tailored indigenous marine consortia for the degradation of naturally weathered polyethylene films. PLoS ONE 2017, 12, e0183984. [Google Scholar] [CrossRef] [Green Version]
- Aravinthan, A.; Arkatkar, A.; Juwarkar, A.A.; Doble, M. Synergistic growth of Bacillus and Pseudomonas and its degradation potential on pretreated polypropylene. Prep. Biochem. Biotechnol. 2016, 46, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Giacomucci, L.; Raddadi, N.; Soccio, M.; Lotti, N.; Fava, F. Biodegradation of polyvinyl chloride plastic films by enriched anaerobic marine consortia. Mar. Environ. Res. 2020, 158, 104949. [Google Scholar] [CrossRef] [PubMed]
- Ballerstedt, H.; Tiso, T.; Wierckx, N.; Wei, R.; Averous, L.; Bornscheuer, U.; O’Connor, K.; Floehr, T.; Jupke, A.; Klankermayer, J.; et al. MIXed plastics biodegradation and UPcycling using microbial communities: EU Horizon 2020 project MIX-UP started January 2020. Environ. Sci. Eur. 2021, 33, 99. [Google Scholar] [CrossRef]
- Leon-Zayas, R.; Roberts, C.; Vague, M.; Mellies, J.L. Draft genome sequences of five environmental bacterial isolates that degrade polyethylene terephthalate plastic. Microbiol. Resour. Announc. 2019, 8, e00237-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, C.; Edwards, S.; Vague, M.; Leon-Zayas, R.; Scheffer, H.; Chan, G.; Swartz, N.A.; Mellies, J.L. Environmental consortium containing pseudomonas and bacillus species synergistically degrades polyethylene terephthalate plastic. Msphere 2020, 5, e01151-20. [Google Scholar] [CrossRef]
- Torena, P.; Alvarez-Cuenca, M.; Reza, M. Biodegradation of polyethylene terephthalate microplastics by bacterial communities from activated sludge. Can. J. Chem. Eng. 2021, 99, S69–S82. [Google Scholar] [CrossRef]
- Oberbeckmann, S.; Osborn, A.M.; Duhaime, M.B. Microbes on a bottle: Substrate, season and geography influence community composition of microbes colonizing marine plastic debris. PLoS ONE 2016, 11, e0159289. [Google Scholar] [CrossRef]
- Webb, H.K.; Crawford, R.J.; Sawabe, T.; Ivanova, E.P. Poly(ethylene terephthalate) Polymer Surfaces as a substrate for bacterial attachment and biofilm formation. Microbes Environ. 2009, 24, 39–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.K.; Pal, S.; Ray, S. Study of microbes having potentiality for biodegradation of plastics. Environ. Sci. Pollut. Res. 2013, 20, 4339–4355. [Google Scholar] [CrossRef] [PubMed]
- Oberbeckmann, S.; Kreikemeyer, B.; Labrenz, M. Environmental factors support the formation of specific bacterial assemblages on microplastics. Front. Microbiol. 2018, 8, 2709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, M.; Langer, T.M.; Hueffer, T.; Hofmann, T.; Herndl, G.J. The composition of bacterial communities associated with plastic biofilms differs between different polymers and stages of biofilm succession. PLoS ONE 2019, 14, e0217165. [Google Scholar] [CrossRef] [Green Version]
- Denaro, R.; Aulenta, F.; Crisafi, F.; Di Pippo, F.; Viggi, C.C.; Matturro, B.; Tomei, P.; Smedile, F.; Martinelli, A.; Di Lisio, V.; et al. Marine hydrocarbon-degrading bacteria breakdown poly(ethylene terephthalate) (PET). Sci. Total Environ. 2020, 749, 141608. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Sun, C. A marine bacterial community capable of degrading poly(ethylene terephthalate) and polyethylene. J. Hazard. Mater. 2021, 416, 125928. [Google Scholar] [CrossRef]
- Beri, D.; York, W.S.; Lynd, L.R.; Pena, M.J.; Herring, C.D. Development of a thermophilic coculture for corn fiber conversion to ethanol. Nat. Commun. 2020, 11, 1937. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Wang, X.; Zhang, H. Balancing the non-linear rosmarinic acid biosynthetic pathway by modular co-culture engineering. Metab. Eng. 2019, 54, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Shahab, R.L.; Brethauer, S.; Davey, M.P.; Smith, A.G.; Vignolini, S.; Luterbacher, J.S.; Studer, M.H. A heterogeneous microbial consortium producing short-chain fatty acids from lignocellulose. Science 2020, 369, eabb1214. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.A.; Wang, X. Use of bacterial co-cultures for the efficient production of chemicals. Curr. Opin. Biotechnol. 2018, 53, 33–38. [Google Scholar] [CrossRef]
- Krause, S.M.B.; Johnson, T.; Karunaratne, Y.S.; Fu, Y.; Beck, D.A.C.; Chistoserdova, L.; Lidstrom, M.E. Lanthanide-dependent cross-feeding of methane-derived carbon is linked by microbial community interactions. Proc. Natl. Acad. Sci. USA 2017, 114, 358–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, X.; Chen, L.; Sui, Y.; Chen, C.; Zhang, W.; Zhou, J.; Dong, W.; Jiang, M.; Xin, F.; Ochsenreither, K. Biotechnological potential and applications of microbial consortia. Biotechnol. Adv. 2020, 40, 107500. [Google Scholar] [CrossRef]
- Jones, J.A.; Vernacchio, V.R.; Collins, S.M.; Shirke, A.N.; Xiu, Y.; Englaender, J.A.; Cress, B.F.; McCutcheon, C.C.; Linhardt, R.J.; Gross, R.A.; et al. Complete biosynthesis of anthocyanins using E. coli Polycultures. Mbio 2017, 8, e00621-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
PET metabolic pathways and conversion of high value chemicals. Enzymes implicated in the pathways, PedH: quinoprotein alcohol dehydrogenase; PedE: quinoprotein alcohol dehydrogenase; PedI: aldehyde dehydrogenase family protein; PP_0545: aldehyde dehydrogenase family protein; GlcDEF: glycolate oxidase; Gcl: glyoxylate carboligase; GlxR: tartronate semialdehyde reductase; PP_3178: glycerate kinase; Eno: Enolase; PykA: Pyruvate kinase; GlcB: malate synthase; AceA: isocitrate lyase; Hyi: Hydroxypyruvate isomerase; PduCDE: propane diol dehydratase; PduP: CoA-dependent propionaldehyde dehydrogenase; TphAabc: TPA 1,2-dioxygenase; TphB: 1,2-dihydroxy-3,5-cyclohexadiene-1,4-dicarboxylate dehydrogenase; PCDO: protocatechuate 3,4-dioxygenase; CMLE: β-carboxy-cis, cis-muconate lactonizing enzyme; CMD: β-carboxymuconolactone decarboxylase; ELH: enollactone hydrolase; TR: β-ketoadipate:succinyl-CoA transferase; TH: β-ketoadipyl-CoA thiolase; PhaA: acetyl-CoA acetyltransferase; PhaB: acetoacetyl-CoA reductase; PhaC: poly(3-hydroxyalkanoate) polymerase; PobA: p-hydroxybenzoate hydroxylase; AroY: PCA decarboxylase; OMT: catechol O-methyltransferase; PhKLMNOPQ: phenol hydroxylase; CatA: a catechol 1,2-dioxygenase originating; CatBC: Muconate cycloisomerase 1/Muconolactone Delta-isomerase; ER: Enoate reductase. Steps in PET degradation; EG metabolism, TPA metabolism, TCA cycle, and high value chemical synthesis are indicated by dark blue, green, orange, light blue, and red, respectively.
Figure 1.
PET metabolic pathways and conversion of high value chemicals. Enzymes implicated in the pathways, PedH: quinoprotein alcohol dehydrogenase; PedE: quinoprotein alcohol dehydrogenase; PedI: aldehyde dehydrogenase family protein; PP_0545: aldehyde dehydrogenase family protein; GlcDEF: glycolate oxidase; Gcl: glyoxylate carboligase; GlxR: tartronate semialdehyde reductase; PP_3178: glycerate kinase; Eno: Enolase; PykA: Pyruvate kinase; GlcB: malate synthase; AceA: isocitrate lyase; Hyi: Hydroxypyruvate isomerase; PduCDE: propane diol dehydratase; PduP: CoA-dependent propionaldehyde dehydrogenase; TphAabc: TPA 1,2-dioxygenase; TphB: 1,2-dihydroxy-3,5-cyclohexadiene-1,4-dicarboxylate dehydrogenase; PCDO: protocatechuate 3,4-dioxygenase; CMLE: β-carboxy-cis, cis-muconate lactonizing enzyme; CMD: β-carboxymuconolactone decarboxylase; ELH: enollactone hydrolase; TR: β-ketoadipate:succinyl-CoA transferase; TH: β-ketoadipyl-CoA thiolase; PhaA: acetyl-CoA acetyltransferase; PhaB: acetoacetyl-CoA reductase; PhaC: poly(3-hydroxyalkanoate) polymerase; PobA: p-hydroxybenzoate hydroxylase; AroY: PCA decarboxylase; OMT: catechol O-methyltransferase; PhKLMNOPQ: phenol hydroxylase; CatA: a catechol 1,2-dioxygenase originating; CatBC: Muconate cycloisomerase 1/Muconolactone Delta-isomerase; ER: Enoate reductase. Steps in PET degradation; EG metabolism, TPA metabolism, TCA cycle, and high value chemical synthesis are indicated by dark blue, green, orange, light blue, and red, respectively.

Figure 2.
Artificial microbial consortia for PET biodegradation and bioconversion.


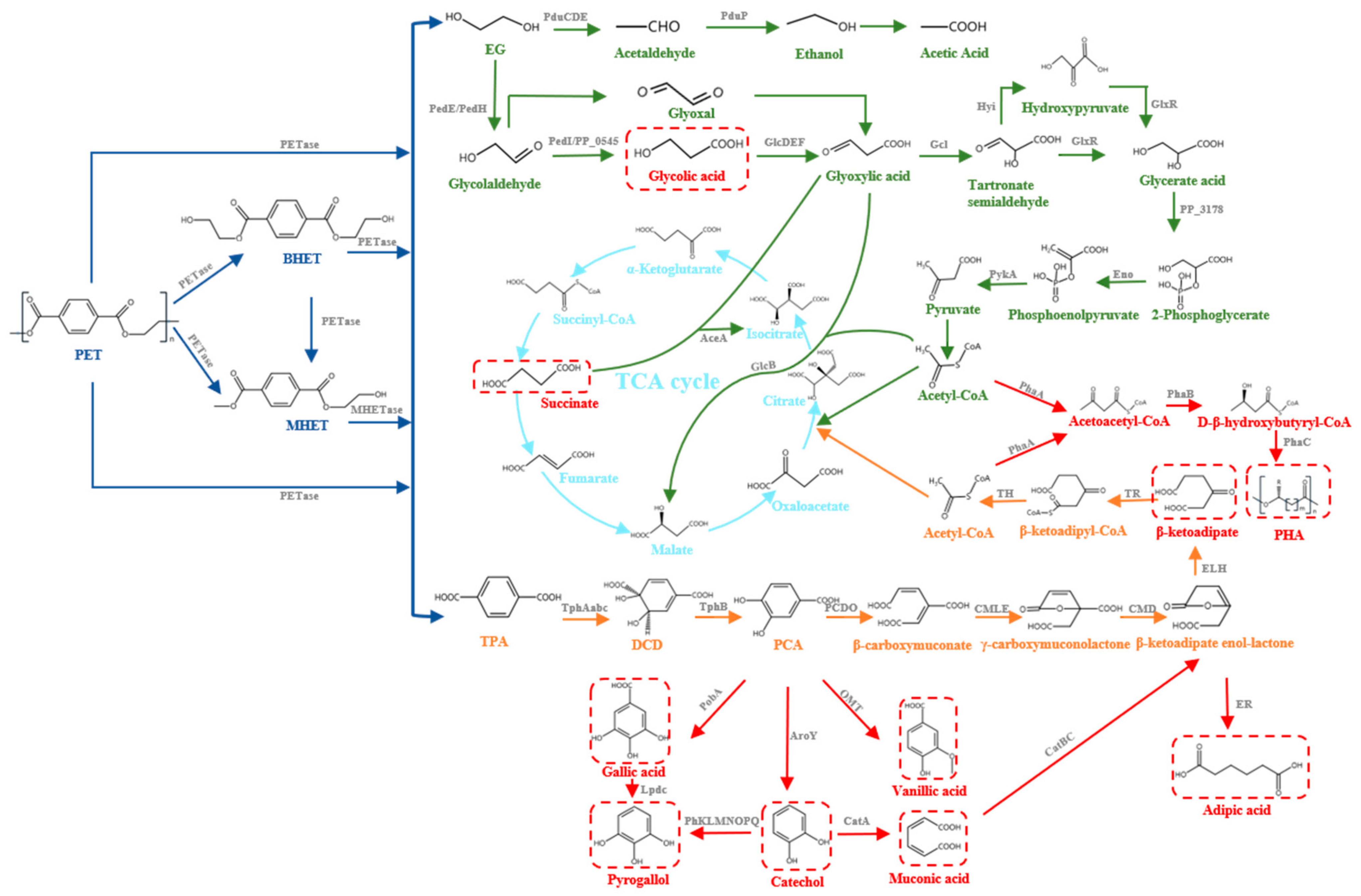{kind=link}
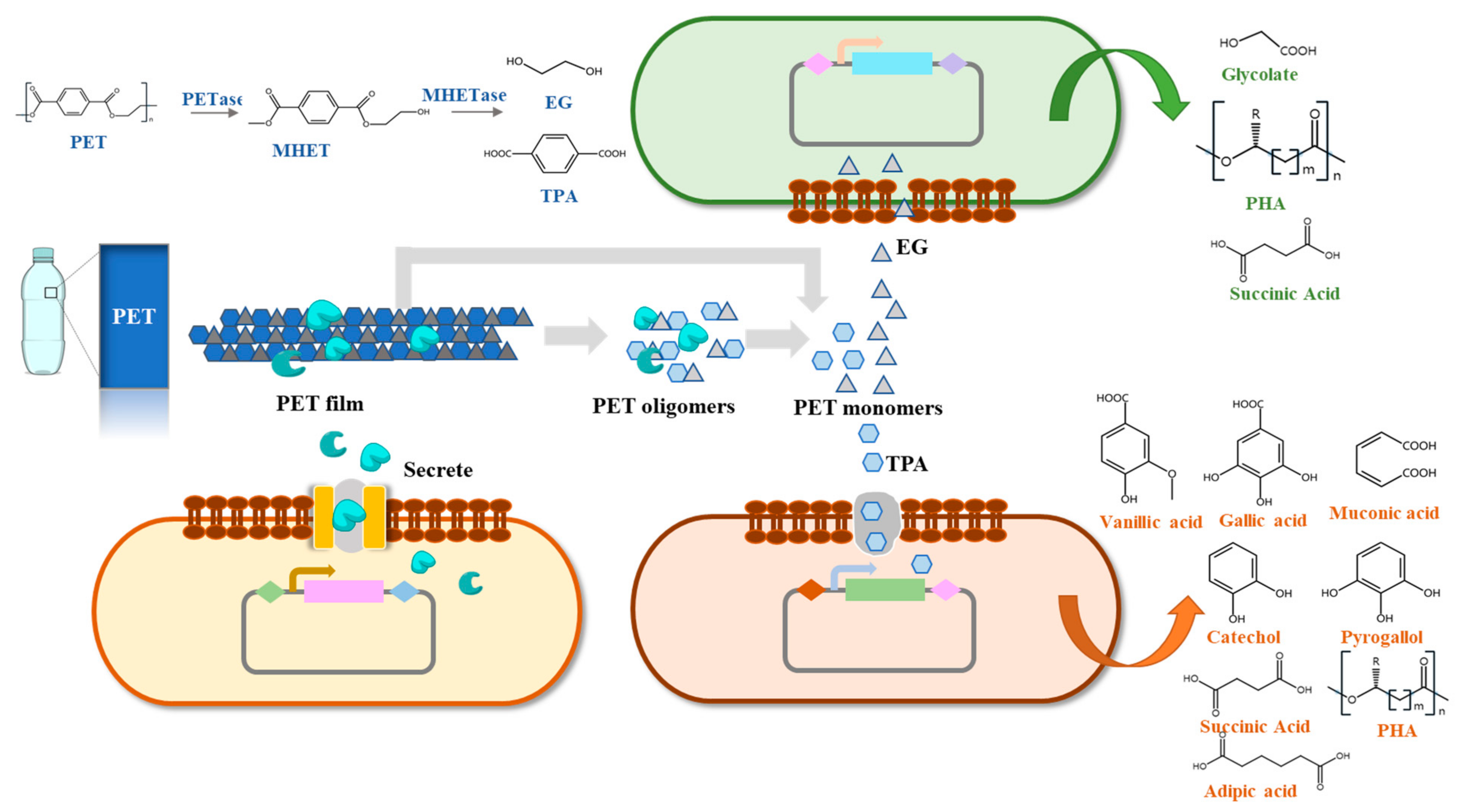{kind=link}
Table 1.
Engineered hydrolases for PET biodegradation.
| Strategy | Hydrolase | Source | Variant | Substract | pH | Temperature | Time | Effect 1 | References |
|---|---|---|---|---|---|---|---|---|---|
| Engineering the key binding sites | MHETase | Ideonella sakaiensis 201-F6 | F424Q/F424N | BHET | 7.5 | 30 °C | 19.25 h | Turnover rate: >0.12 s−1 | [14] |
| MHETase | Ideonella sakaiensis 201-F6 | W397A, F415H, H488A | MHET | 7.5 | 30 °C | 19.25 h | Turnover rates: 2.2-, 1.6-, and 1.15-fold, respectively | [14] | |
| MHETase | Ideonella sakaiensis 201-F6 | S416A/F424N, R411A/S419G/F424N | BHET | 7.5 | 30 °C | 19.25 h | Turnover rate: >0.12 s−1 (120-fold) | [14] | |
| PETase | Ideonella sakaiensis 201-F6 | S238F/W159H | PET of ~15% crystallinity | 7.2 | 30 °C | 96 h | Crystallinity loss: 4.13%; TPA release rate: 1.2-fold | [15] | |
| PETase | Ideonella sakaiensis 201-F6 | S238F/W159H | Semi-aromatic polyester PEF | 7.2 | 30 °C | 96 h | Crystallinity loss: 1.3%; FDCA release rate: 1.2-fold | [15] | |
| MHETase | Ideonella sakaiensis 201-F6 | F424N, F424V, F424I, R411K, R411K/F424N, R411K/F424V, R411K/F424I, R411K/S416A/F424I | BHET | 8.0 | 30 °C | 4 h | Activity: 3.9-, 3.0-, 3.4-, 1.7-, 8.7-, 10.5-, 11.1-, and 15.3-fold, respectively | [20] | |
| MHETase | Ideonella sakaiensis 201-F6 | R411K/S416A/F424I | PETaseS121A/D186H/R280A-treated PET films | 8.0 | 30 °C | 72 h | Activity: 2-fold | [20] | |
| PETase | Ideonella sakaiensis 201-F6 | Y58A, W130A, W130H, A180I, S185H | Drinking bottle | 9.0 | 30 °C | 48 h | Activity: 3.5-, 1.9-, 3.3-, 2.0-, and 2.3-fold, respectively | [49] | |
| PETase | Ideonella sakaiensis 201-F6 | R61A, L88F, I179F, Y58A | Amorphous PET films | 8.5 | 30 °C | 48 h | Activity: 1.4-, 2.1-, and 2.5-fold, respectively | [50] | |
| Tfu_0883 | Thermobifida fusca | Q132A/T101A, I218A | PET fabric (100% polyester) | 7.5 | 60 °C | 48 h | Productivity of TPA: 19.3 ± 0.1 mM (1.6-fold increase) and 15.4 ± 0.1 mM (1.3-fold increase), respectively | [51] | |
| BurPL | Burkholderiales bacterium RIFCSPLOWO2_02_FULL_57_36) | H344S/F348I | PET powder/Goodfellow PET | 9.0 | 35 °C | 18 h | Productivity of the MHET and TPA: ~3-fold increase | [52] | |
| PbPL | Polyangium brachysporum | H216S/F220I | PET powder/Goodfellow PET | 9.0 | 40 °C | 18 h | Productivity of the MHET and TPA: ~10-fold increase | [52] | |
| CtPL | Caldimonas taiwanensis | H210S/F214I | PET powder | 9.0 | 60 °C | 18 h | Productivity of the MHET: 23.12 ± 2.14 μM; Productivity of the TPA: 6.74 ± 1.21 μM | [52] | |
| PET2 | Metagenomics analysis | H229S/F233I | PET powder | 9.0 | 50 °C | 18 h | Productivity of the TPA: 30.31 ± 0.13 μM | [52] | |
| LCC | Leaf-branch compost | H218S/F222I | PET powder | 9.0 | 60 °C | 18 h | Productivity of the MHET: ~900 μM; Productivity of the TPA: ~450 μM | [52] | |
| TfCut2 | Thermobifida fusca22 | H185S/F189I | PET powder | 9.0 | 60 °C | 18 h | Productivity of the MHET: ~120 μM; Productivity of the TPA: ~80 μM | [52] | |
| FsC | Fusarium solani pisi | L81A | PET fibers | 7.5 | 37 °C | 24 h | Activity: 5-fold increase | [66] | |
| Cut190 | Saccharomonospora viridis AHK190 | I224A/Q138A | BHET | 8.2 | 37 °C | Catalytic activity: 150 ± 0.2 s−1 | [67] | ||
| PETase | Ideonella sakaiensis 201-F6 | R280A | Commercial PET films | 9.0 | 30 °C | 18 h/36 h | Activity: increased by 22.4% in 18 h and 32.4% in 36 h | [68] | |
| PE-H | Pseudomonas aestusnigri | Y250S | Amorphous PET films | 7.4 | 30 °C | 48 h | Productivity of MHET: >5 mg/L | [69] | |
| PE-H | Pseudomonas aestusnigri | Y250S | Commercial single use bottle | 7.4 | 30 °C | 48 h | Productivity of MHET: >0.12 mg/L | [69] | |
| Improving the stability | LCC | Leaf-branch compost | LCC-G | PET films of 7% crystallinity | 8.0 | 70 °C | 48 h | Weight loss: ~95% | [35] |
| Cut190 | Saccharomonospora viridis AHK190 | S226P/R228S | Amorphous PET films | 8.2 | 63 °C | 3 d | Weight loss: 14% for PET-GF and 27% for PET-S | [37] | |
| TfCut2 | Thermobifida fusca | D174R, D204R, E253R | Low crystallinity PET films | 8.5 | 65 °C | 48 h | Weight loss: 6.9 ± 0.0%, 11.3 ± 0.3%, and 10.1 ± 0.3%, respectively | [38] | |
| LCC | Leaf-branch compost | F243I/D238C/S283C/Y127G | Post-consumer colored-flake PET waste | 8.0 | 72 °C | 9.3 h | Degradation rate: 90%; Productivity of TPA: 16.7 g·L−1·h−1 | [53] | |
| LCC | Leaf-branch compost | F243W/D238C/S283C/Y127G | Post-consumer colored-flake PET waste | 8.0 | 72 °C | 10.5 h | Degradation rate: 90%; Productivity of TPA: 16.7 g·L−1·h−1 | [53] | |
| PETase | Ideonella sakaiensis 201-F6 | S214H/I168R/W159H/S188Q/R280A/A180/G165A/Q119Y/L117/T140D | PET films of 49.2% crystallinity | 9.0 | 40 °C | 10 d | Activity: 400-fold | [54] | |
| Est119 | Thermobifida alba AHK119 | A68V/S219P | p-nitrophenyl butyrate | 7.0 | 37 °C | 16 h | Activity: 50-fold increase | [55] | |
| TfCut2 | Thermobifida fusca | D204C/E253C/D174R | Low crystallinity PET films | 8.0 | 70 °C | 48 h | Weight loss: 25.0 ± 0.8% (WT: 0.3 ± 0.1%) | [56] | |
| TfCut2 | Thermobifida fusca KW3 | G62A/I213S | Amorphous PET films | 8.0 | 65 °C | 50 h | Weight loss: 42% (2.7-fold) | [63] | |
| Cut190 | Saccharomonospora viridis AHK190 | Q138A/D250C-E296C/Q123H/N202H | Microfiber amorphous PET | 8.5 | 70 °C | 3 d | Degradation rate: > 30% | [70] | |
| PETase | Ideonella sakaiensis 201-F6 | S121E/D186H/R280A, S121D/D186H, S121E/D186H | Commercial PET films | 9.0 | 40 °C | 24 h | Activity: 9.1-, 3.4-, and 4.5-fold, respectively | [71] | |
| PETase | Ideonella sakaiensis 201-F6 | S121E/D186H/R280A, S121D/D186H, S121E/D186H | Commercial PET films | 9.0 | 40 °C | 72 h | Activity: 13.9-, 4.4-, and 6.0-fold, respectively | [71] | |
| PETase | Ideonella sakaiensis 201-F6 | S242T, N246D, S121E/D186H/S242T/N246D | Bottle-grade PET films | 9.0 | 37 °C | 24 h | Activity: 2.5-, 2.6-, and 58-fold, respectively | [72] | |
| Cbotu_EstA | Clostridium botulinum | del71Cbotu_EstA | Amorphous PET films | 7.0 | 50 °C | 120 h | Activity: > 8-fold | [73] | |
| Increasing the substrate accessibility | PETase | Ideonella sakaiensis 201-F6 | S93M, W159F, N241F | L-naphthyl butyrate | 8.0 | 30 °C | Activity: 2.5-, 4.3-, and 3.3-fold, respectively | [48] | |
| Thc_Cut1 | Thermobifida cellulosylitica | Fusion to CBM | Amorphous PET films | 7.0 | 50 °C | 72 h | Productivity of TPA and MHET: 1.7 mol/mol (WT: 1.2 mol/mol) | [60] | |
| Thc_Cut1 | Thermobifida cellulosylitica | Fusion to CBM | Amorphous PET films | 7.0 | 50 °C | 72 h | Productivity of TPA and MHET: 4.5 mol/mol (WT: 1.2 mol/mol) | [60] | |
| Thc_Cut1 | Thermobifida cellulosylitica | Fusion to HFB4 | Amorphous PET films | 7.0 | 50 °C | 24 h | Degradation rate: >16-fold | [61] | |
| Thc_Cut1 | Thermobifida cellulosylitica | Fusion to HFB7 | Amorphous PET films | 7.0 | 50 °C | 24 h | Degradation rate: >16-fold | [61] | |
| PETase | Ideonella sakaiensis 201-F6 | R53E | Low crystallinity PET films | 8.0, 9.0 | 30 °C | Degradation rate: >0.2 nmol/min−1cm−2 | [65] | ||
| TfCut2 | Thermobifida fusca KW3 | G62A/F209A | Low crystallinity PET films | 9.0 | 65 °C | 30 h | Degradation rate: 97 ± 1.8% | [74] | |
| Reducing the interaction between enzymes and products | HiC | Humicola insolens | Non-carbonated mineral water bottles | 7.0 | 50 °C | 14 d | Degradation rate: 7.7-fold increase | [62] | |
| TfCut2 | Thermobifida fusca KW3 | G62A/I213S | Amorphous PET films | 8.0 | 65 °C | 50 h | Weight loss: 42% (2.7-fold) | [63] | |
| TfCut2 | Thermobifida fusca KW3 | TfCa-TfCut2 | Amorphous PET films | 8.0 | 60 °C | 24 h | Productivity: increased by 91% | [75] | |
| LCC | Leaf-branch compost | TfCa-LCC | Amorphous PET films | 8.0 | 60 °C | 24 h | Total products: increased by 104% | [75] |
1 All multiples without explanation are compared to wild-type (WT).
Table 2.
Engineered microbial chassis for PET biodegradation.
| Organism | Strain | Hydrolase | Strategy | Signal Peptide | Reaction Temperature | Substrate | Effects | Ref. |
|---|---|---|---|---|---|---|---|---|
| Bacteria | B. subtilis CBS2 | BhrPETase | Overexpression of molecular chaperones (△ hrcA) | AprE | 70 °C | Amorphous PET films | Expression titer: 0.66 g/L; 6.3 mM products (0.17 mM BHET, 3.66 mM MHET, 2.47 mM TPA) for 20 h | [42] |
| LCC | Expression titer: 0.89 g/L | |||||||
| E. coli BL21(DE3)-T1R | PETase | Selection of signal peptides | SPLamB | 30 °C | Commercial PET films | Expression titer: 6.2 mg/L; MHET/TPA: 2.3 mg/L for 24 h, 3.7 mg/L for 72 h | [80] | |
| E. coli BL21-Gold (DE3) | PETase | Selection and random mutagenesis of signal peptides | PM3 (Evolved PelB, G58A) | 30 °C | BHET | Enzymatic activity: 1.7-fold more than PelB | [81] | |
| PET powder/ | Enzymatic activity: produced 1117 μM MHET and TPA (2.1-fold more than PelB) for 18 h | |||||||
| Amorphous PET films | Enzymatic activity: obvious morphological changes and pores appeared for 168 h | |||||||
| E. coli BL21 (DE3) | PETase | Signal peptide modification | PelB modified by enhancer B1 (MERACVAV) | 30 °C | Amorphous PET films | 62-fold more excretion than PelB | [82] | |
| Fusion hydrophobin (HFBII) | MHET/TPA: 2.7-fold increase for 42 h | |||||||
| B. subtilis 168 | PETase | Selection of signal peptides; Delete Tat translocases | SPPETase | 30 °C | Amorphous PET films | 3.8-fold more excretion than not deleted Tat translocases; Expression titer: 15 mg/L for 20 h; | [83] | |
| B. subtilis WB600 | PETase | Selection of signal peptides; Promoter optimization (P43 promoter) | SPamy | 28 °C | PET films | Pores and serious corrosion appeared for 36 h | [84] | |
| C. thermocellum | LCC | Thermophilic whole-cell degradation system | Signal peptide from exoglucanase Cel48S | 60 °C | Commercial PET films | Soluble monomer feedstocks: >30 mg (60%) for 14 d | [85] | |
| Fungi | Y. lipolytica Po1f | PETase | Selection of signal peptides | SPLIP2 | 30 °C | BHET | 3.68 mM BHET was degraded for 1 h | [76] |
| P. pastoris | PETase | Surface display; Whole-cell biocatalyst | ND | 30 °C | Commercial PET bottles | Enzymatic activity: 36-fold increase | [86] | |
| Marine microalgae | P. tricornutum | PETaseR280A | Fusion proteins and localization | Alkaline phosphatase (AP) | 21 °C | Commercial PET beverage bottle | Almost all MHET was converted into TPA for 10 d | [87] |
| C. reinhardtii CC-124 | PETaseR280A | Cell lysis catalysis | ND | 30 °C | Commercial PET beverage bottle | Many holes and dents appeared and TPA was detected for 4 weeks | [88] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Qi, X.; Yan, W.; Cao, Z.; Ding, M.; Yuan, Y. Current Advances in the Biodegradation and Bioconversion of Polyethylene Terephthalate. Microorganisms 2022, 10, 39. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10010039
AMA Style
Qi X, Yan W, Cao Z, Ding M, Yuan Y. Current Advances in the Biodegradation and Bioconversion of Polyethylene Terephthalate. Microorganisms. 2022; 10(1):39. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10010039
Chicago/Turabian StyleQi, Xinhua, Wenlong Yan, Zhibei Cao, Mingzhu Ding, and Yingjin Yuan. 2022. "Current Advances in the Biodegradation and Bioconversion of Polyethylene Terephthalate" Microorganisms 10, no. 1: 39. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10010039
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.