
Stool Microbiota Diversity Analysis of Blastocystis-Positive and Blastocystis-Negative Individuals

 , , ,
, , ,  , ,
, , {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
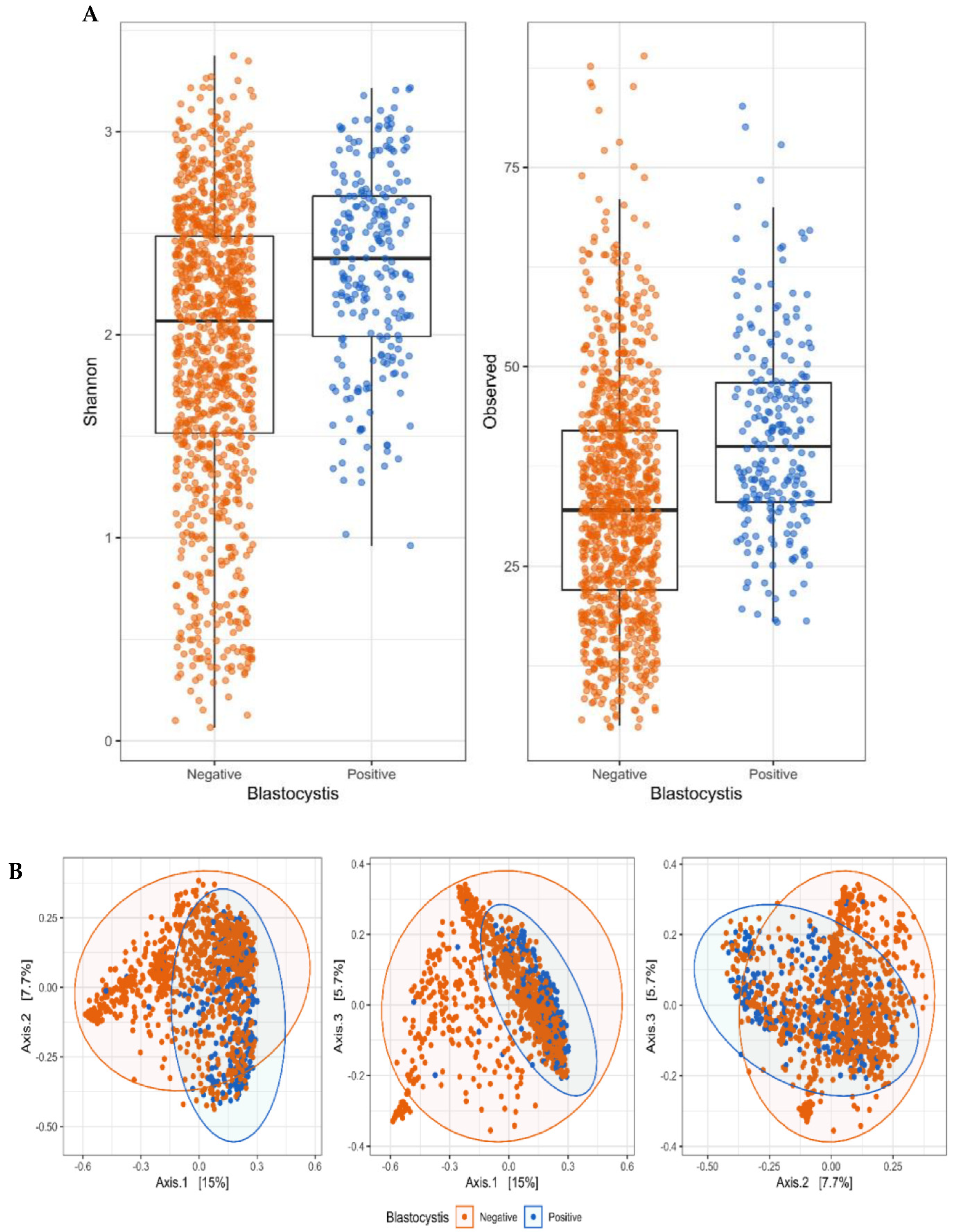
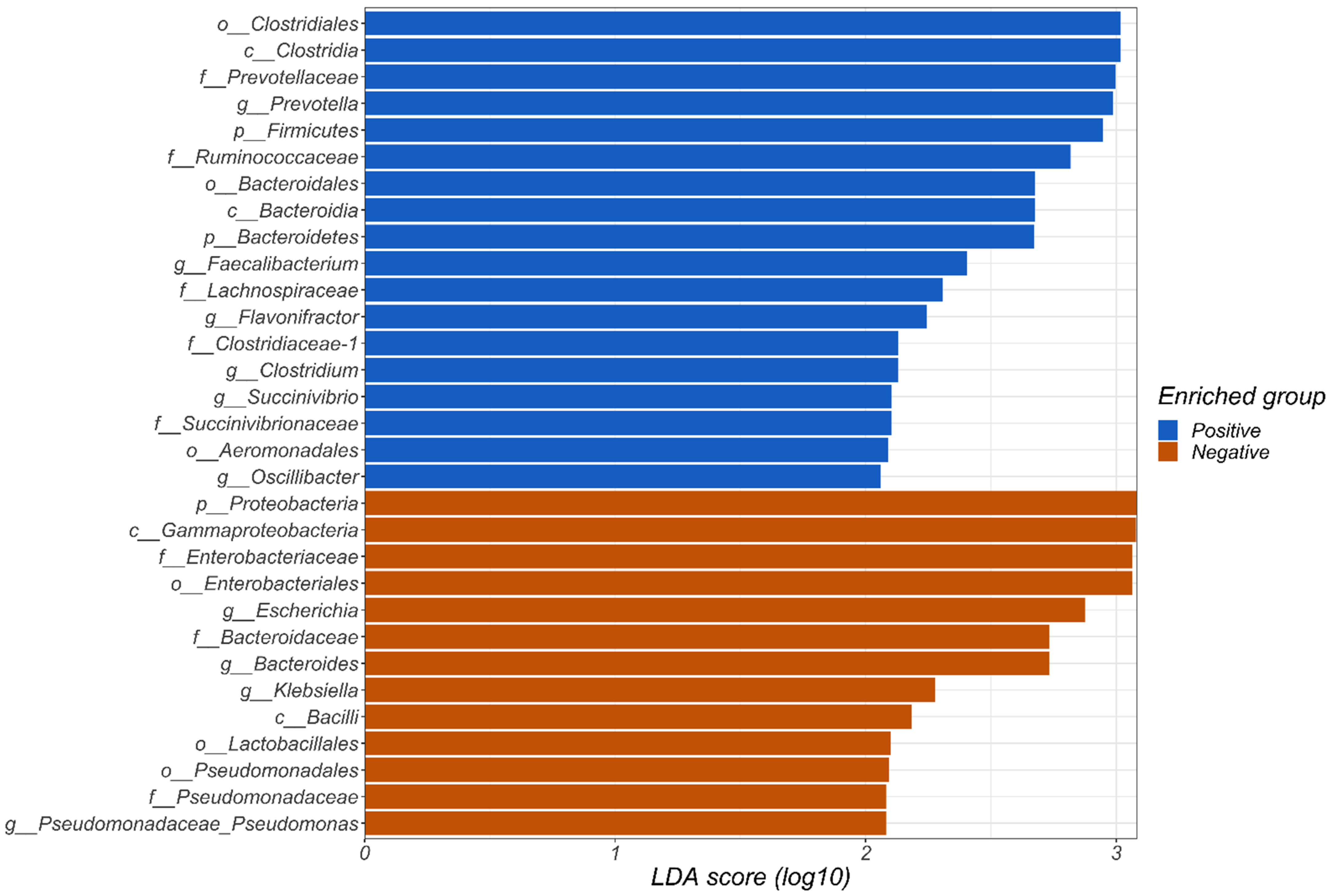
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stensvold, C.R.; Tan, K.S.W.; Clark, C.G. Blastocystis. Trends Parasitol. 2020, 36, 315–316. [Google Scholar] [CrossRef] [PubMed]
- Krogsgaard, L.R.; Engsbro, A.L.; Stensvold, C.R.; Nielsen, H.V.; Bytzer, P. The prevalence of intestinal parasites is not greater among individuals with irritable bowel syndrome: A population-based case-control study. Clin. Gastroenterol. Hepatol. 2015, 13, 507–513.e2. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.O.B.; Karim, A.B.; Roager, H.M.; Vigsnæs, L.K.; Krogfelt, K.A.; Licht, T.R.; Stensvold, C.R. Associations between common intestinal parasites and bacteria in humans as revealed by qPCR. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1427–1431. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.O.B.; Bonde, I.; Nielsen, H.B.; Stensvold, C.R. A retrospective metagenomics approach to studying Blastocystis. FEMS Microbiol. Ecol. 2015, 91, fiv072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Even, G.; Lokmer, A.; Rodrigues, J.; Audebert, C.; Viscogliosi, E.; Ségurel, L.; Chabé, M. Changes in the human gut microbiota associated with colonization by Blastocystis sp. and Entamoeba spp. in non-industrialized populations. Front. Cell. Infect. Microbiol. 2021, 11, 533528. [Google Scholar] [CrossRef] [PubMed]
- Audebert, C.; Even, G.; Cian, A.; Blastocystis Investigation Group; Loywick, A.; Merlin, S.; Viscogliosi, E.; Chabé, M.; El Safadi, D.; Certad, G.; et al. Colonization with the enteric protozoa Blastocystis is associated with increased diversity of human gut bacterial microbiota. Sci. Rep. 2016, 6, 25255. [Google Scholar] [CrossRef] [PubMed]
- Nourrisson, C.; Scanzi, J.; Pereira, B.; NkoudMongo, C.; Wawrzyniak, I.; Cian, A.; Viscogliosi, E.; Livrelli, V.; Delbac, F.; Dapoigny, M.; et al. Blastocystis is associated with decrease of fecal microbiota protective bacteria: Comparative analysis between patients with irritable bowel syndrome and control subjects. PLoS ONE 2014, 9, e111868. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, C.R.; Nielsen, M.; Baraka, V.; Lood, R.; Fuursted, K.; Nielsen, H.V. Entamoeba gingivalis: Epidemiology, genetic diversity and association with oral microbiota signatures in North Eastern Tanzania. J. Oral Microbiol. 2021, 13, 1924598. [Google Scholar] [CrossRef] [PubMed]
- Krogsgaard, L.R.; Andersen, L.O.B.; Johannesen, T.B.; Engsbro, A.L.; Stensvold, C.R.; Nielsen, H.V.; Bytzer, P. Characteristics of the bacterial microbiome in association with common intestinal parasites in irritable bowel syndrome. Clin. Transl. Gastroenterol. 2018, 9, 161. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, C.R.; Lebbad, M.; Hansen, A.; Beser, J.; Belkessa, S.; Andersen, L.O.B.; Clark, C.G. Differentiation of Blastocystis and parasitic archamoebids encountered in untreated wastewater samples by amplicon-based next-generation sequencing. Parasite Epidemiol. Control 2020, 9, e00131. [Google Scholar] [CrossRef] [PubMed]
- Gotfred-Rasmussen, H.; Stensvold, C.R.; Ingham, A.C.; Johannesen, T.B.; Andersen, L.O.B.; Röser, D.; Nielsen, H.V. Impact of metronidazole treatment and Dientamoeba Fragilis colonization on gut microbiota diversity. J. Pediatr. Gastroenterol. Nutr. 2021, 73, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stensvold, C.R.; van der Giezen, M. Associations between gut microbiota and common luminal intestinal parasites. Trends Parasitol. 2018, 34, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Beghini, F.; Pasolli, E.; Truong, T.D.; Putignani, L.; Cacciò, S.M.; Segata, N. Large-scale comparative metagenomics of Blastocystis, a common member of the human gut microbiome. ISME J. 2017, 11, 2848–2863. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Asnicar, F.; Berry, S.E.; Valdes, A.M.; Nguyen, L.H.; Piccinno, G.; Drew, D.A.; Leeming, E.; Gibson, R.; Le Roy, C.; Khatib, H.A.; et al. Microbiome connections with host metabolism and habitual diet from 1,098 deeply phenotyped individuals. Nat. Med. 2021, 27, 321–332. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stensvold, C.R.; Sørland, B.A.; Berg, R.P.K.D.; Andersen, L.O.; van der Giezen, M.; Bowtell, J.L.; El-Badry, A.A.; Belkessa, S.; Kurt, Ö.; Nielsen, H.V. Stool Microbiota Diversity Analysis of Blastocystis-Positive and Blastocystis-Negative Individuals. Microorganisms 2022, 10, 326. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020326
Stensvold CR, Sørland BA, Berg RPKD, Andersen LO, van der Giezen M, Bowtell JL, El-Badry AA, Belkessa S, Kurt Ö, Nielsen HV. Stool Microbiota Diversity Analysis of Blastocystis-Positive and Blastocystis-Negative Individuals. Microorganisms. 2022; 10(2):326. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020326
Chicago/Turabian StyleStensvold, Christen Rune, Brede Aksdal Sørland, Rebecca P. K. D. Berg, Lee O’Brien Andersen, Mark van der Giezen, Joanna L. Bowtell, Ayman A. El-Badry, Salem Belkessa, Özgür Kurt, and Henrik Vedel Nielsen. 2022. "Stool Microbiota Diversity Analysis of Blastocystis-Positive and Blastocystis-Negative Individuals" Microorganisms 10, no. 2: 326. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020326