
Cellular Responses Required for Oxidative Stress Tolerance of the Necrotrophic Fungus Alternaria alternata, Causal Agent of Pear Black Spot

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture Conditions
2.2. Sample Collection of the ROS Metabolism Evaluation
2.3. Assessment of the O2− Generation Rate and H2O2 Content of A. alternata
2.4. ROS Metabolism Key Enzyme Activity and Antioxidant Substances Content of A. alternata Assay
2.5. Real-Time qPCR Analysis
2.6. Statistical Analysis
3. Results
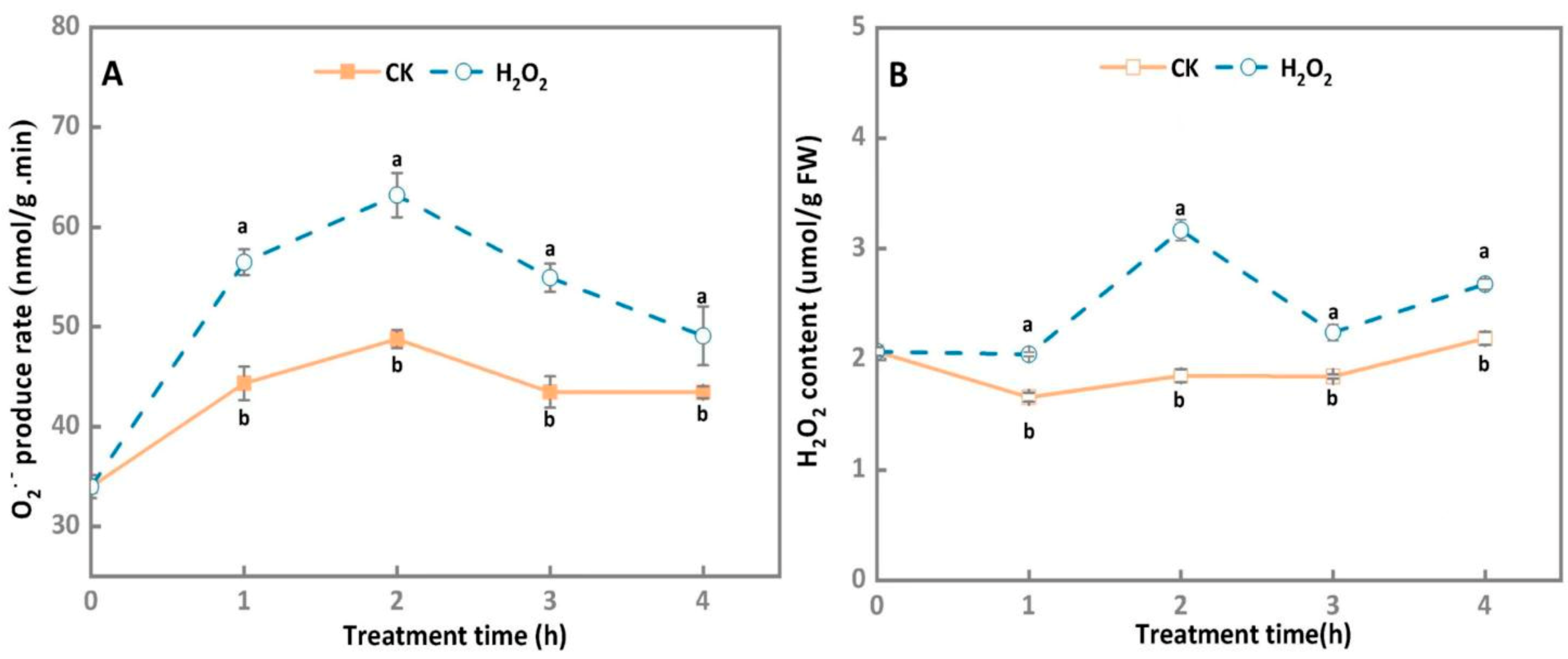
3.1. O2− Generation Rate and H2O2 Content of A. alternata Increased under Exogenous H2O2 Stress
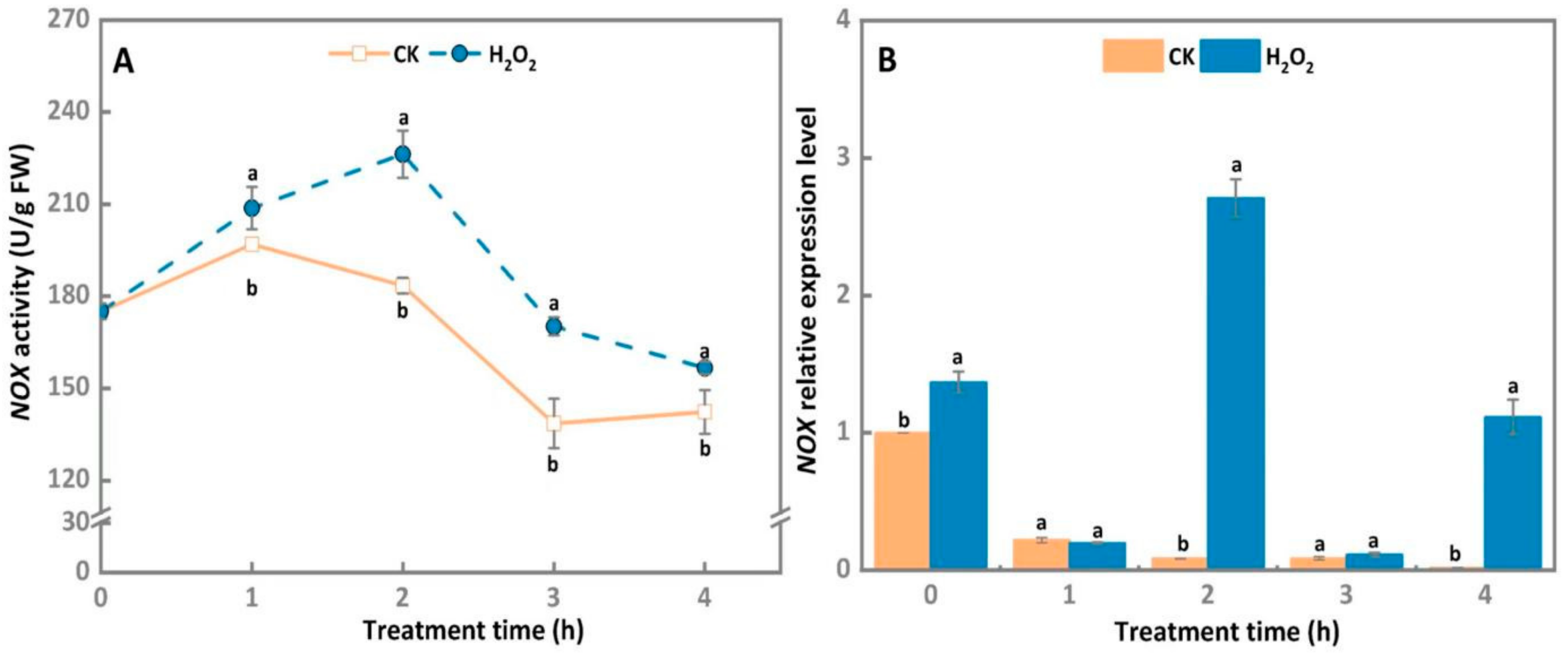
3.2. NOX Activity and Gene Expression Level of A. alternata Increased after Exogenous H2O2 Stress
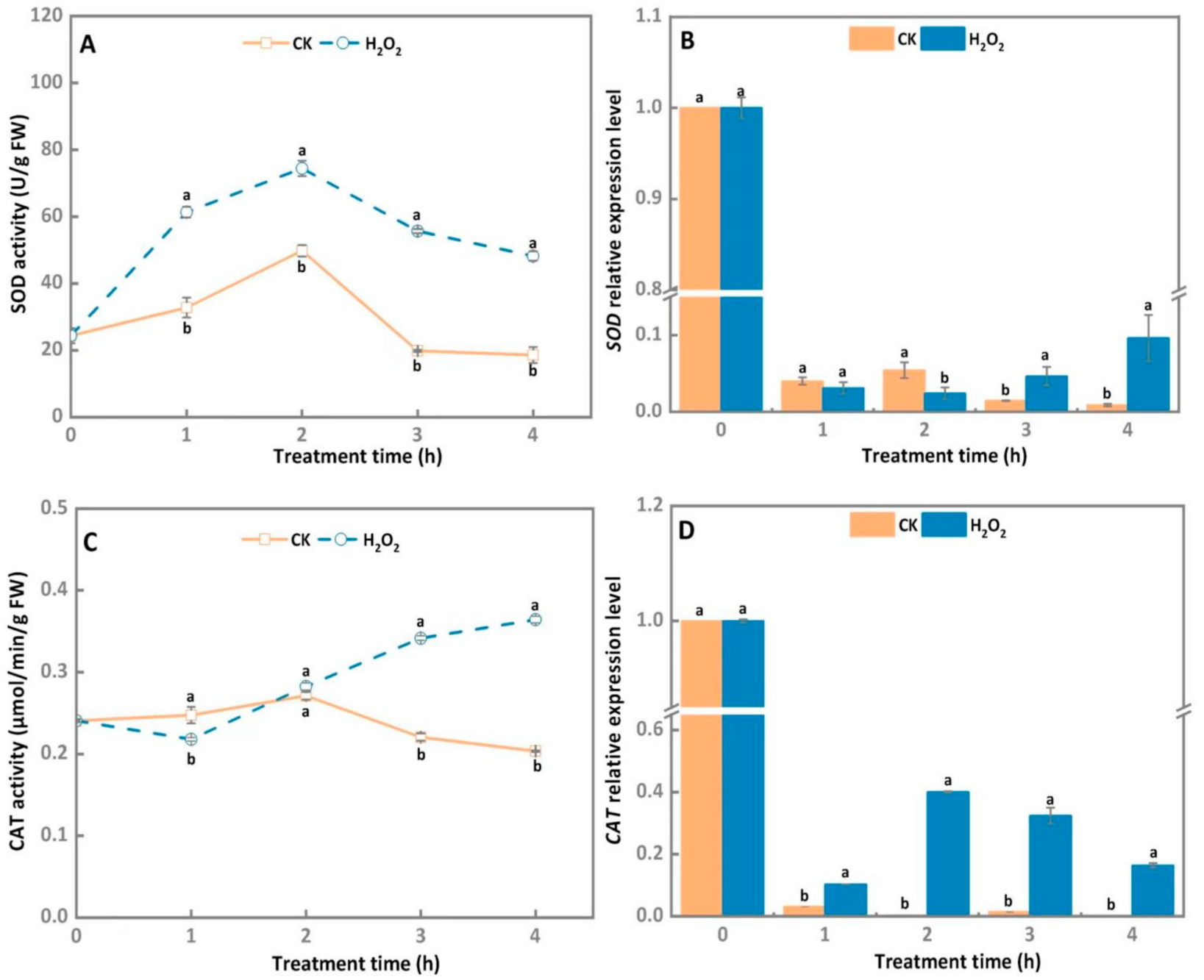
3.3. SOD and CAT Activities, and Transcript Level of A. alternata Increased after Exogenous H2O2 Stress
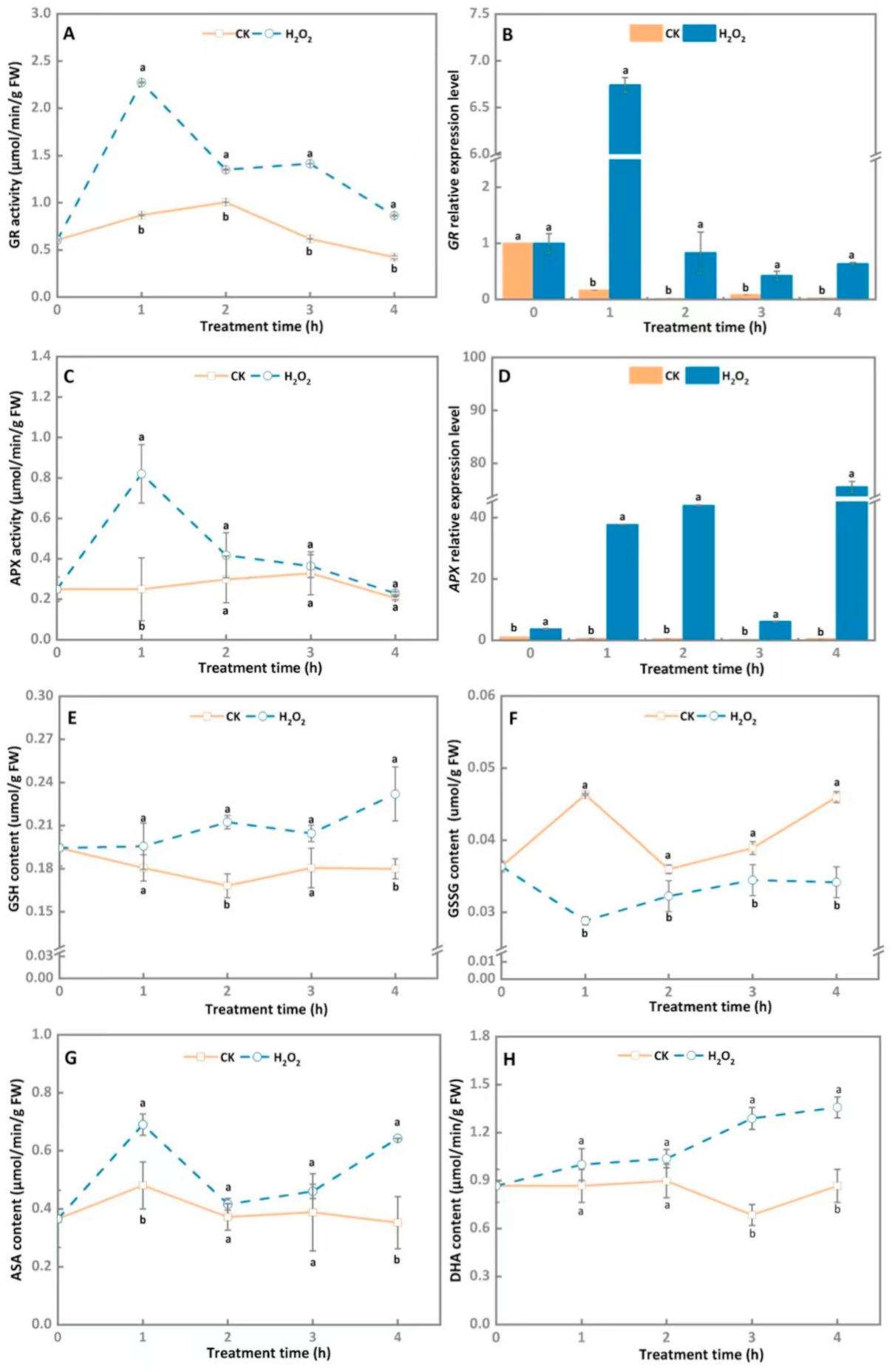
3.4. AsA-GSH Cycle System Involved in Maintaining the Intracellular Redox Balance of A. alternata after Exogenous H2O2 Stress
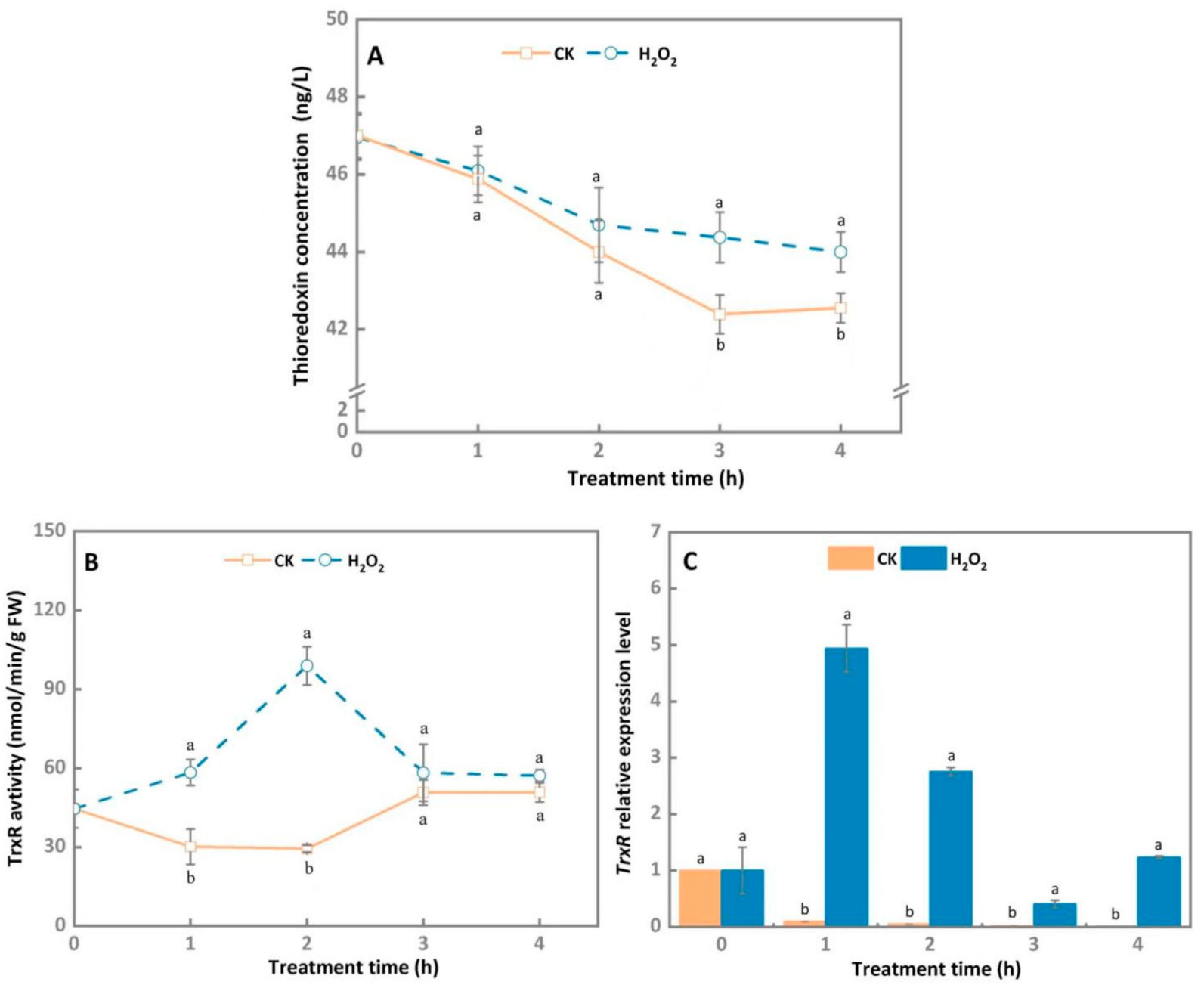
3.5. Thioredoxin System Involved in Maintaining the Intracellular Redox Balance of A. alternata after Exogenous H2O2 Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Y.C.; Yin, Y.; Bi, Y.; Wang, D. Effect of riboflavin on postharvest disease of Asia pear and the possible mechanisms involved. Phytoparasitica 2012, 40, 261–268. [Google Scholar] [CrossRef]
- Mari, M.; Bertolini, P.; Pratella, G.C. Non-conventional methods for the control of post-harvest pear diseases. J. Appl. Microbiol. 2003, 94, 761–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, T.T.; Pu, H.B.; Sun, D.W. Insights into the changes in chemical compositions of the cell wall of pear fruit infected by Alternaria alternata with confocal aman microspectroscopy. Postharvest Biol. Technol. 2017, 132, 119–129. [Google Scholar] [CrossRef]
- Wang, P.H.; Wu, P.C.; Huang, R.; Chung, K.R. The role of a nascent polypeptide associated complex subunit alpha in siderophore biosynthesis, oxidative stress response, and virulence in Alternaria alternata. Mol. Plant Microbe Interact. 2019, 33, 668–679. [Google Scholar] [CrossRef]
- Yang, X.P.; Hu, H.J.; Yu, D.Z.; Sun, Z.H.; He, X.J.; Zhang, J.G.; Chen, Q.L.; Tian, R.; Fan, J.; Liu, J.H. Candidate resistant genes of sand pear (Pyrus pyrifolia nakai) to Alternaria alternata revealed by transcriptome sequencing. PLoS ONE 2015, 10, e0135046. [Google Scholar] [CrossRef]
- Saccon, F.A.M.; Parcey, D.; Paliwal, J.; Sherif, S.S. Assessment of fusarium and deoxynivalenol using optical methods. Food Bioprocess Technol. 2017, 10, 34–50. [Google Scholar] [CrossRef]
- Tanaka, A.; Shiotani, H.; Yamamoto, M.; Tsuge, T. Insertional mutagenesis and cloning of genes required for biosynthesis of the host-specific AK-toxin in the Japanese pear pathotype of Alternaria alternata. Mol. Plant Microbe Interact. 1999, 12, 691–702. [Google Scholar] [CrossRef] [Green Version]
- Kubicek, C.P.; Starr, T.L.; Glass, N.L. Plant cell wall-degrading enzymes and their secretion in plant-pathogenic fungi. Annu. Rev. Phytopathol. 2014, 52, 427–451. [Google Scholar] [CrossRef]
- Takao, K.; Akagi, Y.; Tsuge, T.; Harimoto, Y.; Yamamoto, M.; Kodama, M. The global regulator LaeA controls biosynthesis of host-specific toxins, pathogenicity and development of Alternaria alternata pathotypes. J. Gen. Plant Pathol. 2016, 82, 121–131. [Google Scholar] [CrossRef]
- Camejo, D.; Guzman-Cedeno, A.; Moreno, A. Reactive oxygen species, essential molecules, during plant-pathogen interactions. Plant Physiol. Biochem. 2016, 103, 10–23. [Google Scholar] [CrossRef]
- Bhattacharjee, S. An inductive pulse of hydrogen peroxide pretreatment restores redox- homeostasis and oxidative membrane damage under extremes of temperature in two rice cultivars. Plant Growth Regul. 2012, 68, 395–410. [Google Scholar] [CrossRef]
- Tsukagoshi, H.; Busch, W.; Benfey, P.N. Transcriptional regulation of ROS controls transition from proliferation to differentiation in the root. Cell 2010, 143, 606–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaragoza, O.; Chrisman, C.J.; Castelli, M.V.; Frases, S.; Casadevall, A. Capsule enlargement in Cryptococcus neoformans confers resistance to oxidative stress suggesting a mechanism for intracellular survival. Cell Microbiol. 2008, 10, 2043–2057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganetta, E.; Walker, G.M.; Adya, A.K. Nanoscopic morphological changes in Yyt cell surfaces caused by oative stress: An atomic force microscopic study. Microbiol. Biotechnol. 2009, 19, 547–555. [Google Scholar] [CrossRef]
- Buron-Moles, G.; Torres, R.; Teixidó, N.; Usall, J.; Vilanova, L.; Vinas, I. Characterization of H2O2 production to study compatible and non-host pathogen interactions in orange and apple fruit at different maturity stages. Postharvest Biol. Technol. 2015, 99, 27–36. [Google Scholar] [CrossRef]
- Liu, X.; Williams, C.E.; Nemacheck, J.A.; Wang, H.Y. Reactive oxygen species are involved in plant defense against a gall midge. Plant Physiol. 2010, 152, 985–999. [Google Scholar] [CrossRef] [Green Version]
- Heller, J.; Tudzynski, P. Reactive oxygen species in phytopathogenic fungi: Signaling, development and disease. Annu. Rev. Phytopathol. 2011, 49, 369–390. [Google Scholar] [CrossRef]
- Kou, Y.; Qiu, J.; Tao, Z. Every coin has two sides: Reactive oxygen species during rice Magnaporthe oryzae interaction. Int. J. Mol. Sci. 2019, 20, 1191. [Google Scholar] [CrossRef] [Green Version]
- Eloy, Y.R.; Vasconcelos, I.M.; Barreto, A.L.; Freire-filho, F.R.; Oliveira, J.T.A. H2O2 plays an important role in the lifestyle of Colletotrichum gloeosporioides during interaction with cowpea Vigna unguiculata (L.) walp. Fungal Biol. 2015, 119, 747–757. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.Z.; Guo, L.J.; Yang, L.Y.; Huang, J. Foatf1, a ZIP transcription factor of Fusarium oxysporum f. sp. cubense, is involved in pathogenesis by regulating the oxidative stress responses of Cavendish banana (Musa spp.). Physiol. Mol. Plant Path. 2013, 84, 76–85. [Google Scholar] [CrossRef]
- Li, G.F.; Peng, G.X.; Keyhani, N.O.; Xin, J.; Cao, Y.; Xia, Y.X. A bifunctional catalase-peroxidase, MakatG1, contributes to virulence of Metarhizium acridum by overcoming oxidative stress on the host insect cuticle. Environ. Microbiol. 2017, 19, 4365–4378. [Google Scholar] [CrossRef] [PubMed]
- Russell, E.G.; Cotter, T.G. New insight into the role of reactive oxygen species (ROS) in cellular signal-transduction processes. Int. Rev. Cell Mol. Biol. 2015, 319, 221. [Google Scholar] [CrossRef] [PubMed]
- Mir, A.A. Systematic characterization of the peroxidase gene family provides new insights into fungal pathogenicity in Magnaporthe oryzae. Sci. Rep. 2015, 5, 11831. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.B.; Tang, L.; Ying, S.H.; Feng, M.G. Regulative roles of glutathione reductase and four glutaredoxins in glutathione redox, antioxidant activity, and iron homeostasis of Beauveria bassiana. Appl. Microbiol. Biotechnol. 2016, 100, 5907–5917. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Li, Y.C.; Bi, Y.; Wang, Y. Role of pear fruit cuticular wax and surface hydrophobicity in regulating the prepenetration phase of Alternaria alternata infection. J. Phytopathol. 2017, 165, 313–322. [Google Scholar] [CrossRef]
- Zhang, X.M.; Zong, Y.Y.; Gong, D.; Yu, L.R.; Sionov, E.; Bi, Y.; Prusky, D. NADPH oxidase regulates the growth and pathogenicity of Penicillium expansum. Front. Plant Sci. 2021, 12, 696210. [Google Scholar] [CrossRef]
- Ge, Y.H.; Deng, H.W.; Bi, Y.; Li, Y.C.; Liu, Y.Y. The role of reactive oxygen species in ASM-induced disease resistance in apple fruit. In Postharvest Pathology; Prusky, D., Gullino, M.L., Eds.; Springer International Publishing: Cham, Switzerland, 2014; pp. 39–52. [Google Scholar] [CrossRef]
- Ren, Y.L.; Wang, Y.F.; Bi, Y.; Deng, H. Postharvest BTH treatment induced disease resistance and enhanced reactive oxygen species metabolism in muskmelon (Cucumis melo L.) fruit. Eur. Food Res. Technol. 2012, 234, 963–971. [Google Scholar] [CrossRef]
- Sun, J.Z.; Lin, H.T.; Zhang, S.; Lin, Y.F.; Wang, H.; Lin, M.S.; Chen, Y.H. The roles of ROS production-scavenging system in Lasiodiplodia theobromae (Pat.) Griff. & Maubl. -induced pericarp browning and disease development of harvested longan fruit. Food Chem. 2018, 247, 16–22. [Google Scholar] [CrossRef]
- Turcsanyi, E.; Lyons, T.; Plöchl, M.; Barnes, J. Does ascorbate in the mesophyll cell walls form the first line of defence against ozone? Testing the concept using broad bean (Vicia faba L.). J. Exp. Bot. 2000, 51, 901–910. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Brown, D.I.; Griendling, K.V.K. Nox proteins in signal transduction. Free Radic. Biol. Med. 2009, 47, 1239–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, G.; Shulaev, V.; Mittler, R. Reactive oxygen signaling and abiotic stress. Physiol. Plant. 2008, 133, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Segal, L.M.; Wilson, R.A. Reactive oxygen species metabolism and plant-fungal interactions. Fungal Genet. Biol. 2018, 110, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.L.; Chung, K.R. The NADPH oxidase-mediated production of hydrogen peroxide (H2O2) and resistance to oxidative stress in the necrotrophic pathogen Alternaria alternata of citrus. Mol. Plant Microbe Interact. 2012, 13, 900–914. [Google Scholar] [CrossRef]
- Angelova, M.B.; Pashova, S.B.; Spasova, B.K.; Vassilev, S.V.; Slokoska, L.S. Oxidative stress response of filamentous fungi induced by hydrogen peroxide and paraquat. Mycol. Res. 2005, 109, 150–158. [Google Scholar] [CrossRef]
- Wei, M.L.; Ge, Y.H.; Li, C.Y.; Han, X.; Qin, S.C.; Chen, Y.R.; Tang, Q.; Li, J.R. G6PDH regulated NADPH production and reactive oxygen species metabolism to enhance disease resistance against blue mold in apple fruit by acibenzolar-S-methyl. Postharvest Biol. Technol. 2019, 148, 228–235. [Google Scholar] [CrossRef]
- Li, Q.; Mcneil, B.; Harvey, L.M. Adaptive response to oxidative stress in the filamentous fungus Aspergillus niger B1-D. Free Radic. Biol. Med. 2008, 44, 394–402. [Google Scholar] [CrossRef] [Green Version]
- Chumyam, A.; Shank, L.; Faiyue, B.; Uthaibutra, J.; Saengnil, K. Effects of chlorine dioxide fumigation on redox balancing potential of antioxidative ascorbate-glutathione cycle in ‘Daw’ longan fruit during storage. Sci. Hortic. 2017, 222, 76–83. [Google Scholar] [CrossRef]
- Emri, T.; Pocsi, I.; Szentirmai, A. Glutathione metabolism and protection against oxidative stress caused by peroxides in Penicillium chrysogenum. Free Radic. Biol. Med. 1997, 23, 809–814. [Google Scholar] [CrossRef]
- Jiang, H.; Wang, Y.; Li, C.J.; Wang, B.; Ma, L.; Ren, Y.Y.; Bi, Y.; Li, Y.C.; Xue, H.L.; Prusky, D. The effect of benzo-(1,2,3)-thiadiazole-7-carbothioic acid S-methyl ester (BTH) treatment on regulation of reactive oxygen species metabolism involved in wound healing of potato tubers during postharvest. Food Chem. 2020, 309, 125608. [Google Scholar] [CrossRef]
- Ying, W.H. NAD+/NADH and NADP+/NADPH in cellular functions and cell death: Regulation and biological consequences. Antioxid. Redox Signal. 2008, 10, 179–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dantas, A.D.S.; Patterson, M.J.; Smith, D.A.; Maccallum, D.M.; Quinn, J. Thioredoxin regulates multiple hydrogen peroxide-induced signaling pathways in Candida albicans. Cell Mol. Biol. 2010, 30, 4550–4563. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Wang, H.; Xiong, X.; Chen, S.; Zhang, D. Mitochondria thioredoxin"s backup role in oxidative stress resistance in Trichoderma reesei. Microbiol. Res. 2015, 171, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Viefhues, A.; Heller, J.; Temme, N.; Tudzynski, P. Redox systems in Botrytis cinerea: Impact on development and virulence. Mol. Plant Mocrobe Interact. 2014, 27, 858–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence of Primer (5′ to 3′) | TM |
|---|---|---|
| NOX | F: AACGAAGTCGCAGTTCTTATTGR: GGCGGAGGTGGTAGATAGATT | 60 |
| SOD | F: AACAACTTCAGCGAGCAAATCR: TTGATGGCAGCAGATAGCG | 60 |
| CAT | F: CCACGGCACCTTTGTTTCTR: ATCTCGCACTGTGTCAGCACT | 60 |
| TrxR | F: GCGGTATCGTCAGGCTATCAR: CTATCCCTATGCTGTCATCTTGC | 60 |
| GR | F: GCCAAACACGGTGCAAAAGTR: TTTGAAGGTCTCGGCAATCG | 60 |
| APX | F: AATGCTGGTCTCAAGGCTGCR: GACCCTGCATCTCCTGGATG | 60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Zhang, Y.; Li, Y.; Bi, Y.; Mao, R.; Yang, Y.; Jiang, Q.; Prusky, D. Cellular Responses Required for Oxidative Stress Tolerance of the Necrotrophic Fungus Alternaria alternata, Causal Agent of Pear Black Spot. Microorganisms 2022, 10, 621. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10030621
Zhang M, Zhang Y, Li Y, Bi Y, Mao R, Yang Y, Jiang Q, Prusky D. Cellular Responses Required for Oxidative Stress Tolerance of the Necrotrophic Fungus Alternaria alternata, Causal Agent of Pear Black Spot. Microorganisms. 2022; 10(3):621. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10030621
Chicago/Turabian StyleZhang, Miao, Yandong Zhang, Yongcai Li, Yang Bi, Renyan Mao, Yangyang Yang, Qianqian Jiang, and Dov Prusky. 2022. "Cellular Responses Required for Oxidative Stress Tolerance of the Necrotrophic Fungus Alternaria alternata, Causal Agent of Pear Black Spot" Microorganisms 10, no. 3: 621. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10030621